
Abstract
For desert species, human development may buffer against resource scarcity by providing reliable resources in an otherwise stark environment. We used high-throughput tracking technology to explore the movement patterns of a social desert passerine bird (the Arabian babbler—Argya squamiceps, Leiothrichidae) in a mosaic of human-modified and semi-natural habitats. From late summer to early winter, as natural food and water resources increase, we tracked 21 individuals representing 10 groups. Toward winter, groups spent less time within villages, had smaller home ranges, and were less territorial. In general, birds showed a preference for spending time in the vegetated semi-natural habitat. We further found that even a small section of uncultivated agricultural land (~2 km stretch of mostly bare dirt) can act as a movement barrier for babblers. Altogether, we highlight how the complex interaction between resource availability and anthropogenic changes to habitats, can shape animals’ responses to our changing planet.
Similar content being viewed by others

Introduction
Natural selection is driven by the ever-changing environment and can lead to species’ evolution. However, the intense and rapid environmental changes that characterise the Anthropocene are negatively affecting these natural processes1,2. These changes include habitat loss and modification, the spread of exotic species, overexploitation by humans, and climate change3,4,5,6,7. Consequently, for species to survive they require diverse adaptation mechanisms8. One such adaptation is behavioural flexibility, a mechanism that allows animals to quickly respond to a changing environment (e.g. by changing their movement patterns).
Human modifications to the environment impact organisms across all biomes, due to vast changes to resource abundance and composition, predator abundance or composition, and the creation of novel barriers to movement9,10,11,12. One of the most striking aspects of these modifications to arid desert biomes is the extreme differences between human-altered habitats and natural habitats13. In addition, deserts are facing a more pronounced decline in habitat and biodiversity compared to other biomes14,15. However, despite the fact that terrestrial lands consist of ~40% drylands (e.g. deserts, temperate grasslands, savanna woodlands16), they have received a disproportionate lack of scientific attention17. Desert species may possess greater behavioural flexibility that allows them to adjust quickly to any changes to their environment as a response to the temporally and spatially unpredictable resource availability that characterises most deserts18,19.
Here, we explored the movement patterns of a social desert passerine bird—the Arabian babbler (Argya squamiceps, Leiothrichidae, henceforth ‘babbler’) in the Negev desert, Israel. Specifically, we quantified how group movements changed over time in a mosaic of natural and human-modified habitats. Babblers are territorial group-living birds that are resident in the Negev desert. The tight-knit social structure of babblers makes them a model system to understand fundamental questions in evolutionary biology and behavioural ecology, as group living has been theorised to evolve as an adaptation to coping with harsh environments20,21. In more natural habitat, such as that of a long-term study site in the Arava valley, Israel, babblers are generally limited to the higher resource green patches generally found within the wadis (i.e. dry riverbeds) that interweave the desert landscape22,23. In these wadis, babblers are highly territorial and only occasionally cross territory boundaries24. Nevertheless, despite 50 years of intensive study of various aspects of babblers in the wild [e.g. refs. 24,25,26,27,28.], much of their behaviour remains unknown, especially in modified habitats.
High-throughput tracking systems (specifically, the ATLAS; Advanced Tracking and Localisation of Animals in real-life Systems29) have already revealed key behaviours for several animal species30,31,32,33. However, such high-throughput tracking in desert ecosystems, as well as on social animals, remains unexplored. This approach can be particularly pertinent to understanding whether animals exploit the additional and consistent food and water resources present in human-modified habitats34,35,36. To understand how resource availability around human settlements may be affecting movement patterns and resource use in a territorial social bird, we used the ATLAS to explore babbler group behaviour in a complex mosaic of human-modified and semi-natural habitats. Resources in the human villages and orchards interspersing the babblers’ natural habitat remain available during the hottest and driest periods of the year37,38. These resources can thus potentially provide babblers with a buffering effect against the most extreme weather. We analysed high-resolution bird movements with respect to daily climate, seasonality, and the degree of human alterations of the habitats. We further explored the degree of home-range overlap across the different babbler groups. During summer, when resources are scarcer in the semi-natural desert habitat, we expected groups to spend more time and be more territorial within villages.
Results
In total, we collected 727,605 localisations from 21 tagged birds, for a total of 1542 tag days. This gave us a per-tag average of 59.8 days of data and 31,985 localisations. Once data were cleaned so that each group was represented by a single tag per day, we had an average of 560.4 localisations per day per group (Table S1).
Overall home-range patterns
Daily average distance travelled by groups was 18.77 km, while average home-ranges were 0.31 km2. Distance travelled (km) increased as the date progressed (estimate = 3.56e-2 ± 1.98e-2, t = 1.799, p = 0.072), and decreased with smaller relative group size (estimate = −1.93e-1 ± 1.02e-1, t = −1.897, p = 0.058), although neither relationship was significant. Daily home-ranges (km2) were significantly smaller as the date progressed (estimate = −6.57e-2 ± 1.83e-2, t = −3.589, p < 0.001; Fig. 1) but were not related to relative group size (estimate = 1.21e-1 ± 1.12e-1, t = 1.087, p = 0.277).

Model fits are LMER with linear regression and shaded 95% confidence intervals with individual groups indicated by colour (see legend) and light grey shading and overall average of all groups indicated by overlayed solid black line and dark grey shading. The x-axis has been modified to show the actual date as opposed to scaled date values.
Habitat use
Proportional habitat use and total habitat use (km2) were correlated for all habitat types, although less strongly for plateau (plateau: r = 0.555, village: r = 0.817, orchard: r = 0.781). As the date progressed, groups increased their habitat use of plateau (estimate = 6.66e-4 ± 1.48e-4, t = 4.493, p < 0.001) and decreased their use of village (estimate = −9.26e-4 ± 1.33e-4, t = −6.956, p < 0.001), but did not significantly change their use of orchard (estimate = 7.52e-5 ± 1.38e-4, t = 0.546, p = 0.585; Fig. 2). There were no differences in NDVI among the three time periods for the overall region covered by the ATLAS system (sum of squares = 7.27e-5, F = 8.70e-3, p = 0.991). There were significant differences in NDVI among habitats (Sum of Squares = 2.50e-2, F = 525.81, p < 0.001); orchard habitat had the highest NDVI compared to both other habitats, and village had higher NDVI than the plateau (orchard-plateau: mean difference = 1.28e-1 ± 2.68e-1, p < 0.001; orchard-village: mean difference = 7.99e-2 ± 1.72e-1, p < 0.001; village-plateau: mean difference = 4.79e-2 ± 1.11e-2, p < 0.001).

Model fit is a LMER with linear regression and shaded 95% confidence intervals with habitat indicated by colour (see legend). The x-axis has been modified to show the actual date as opposed to scaled date values.
Within home-ranges, there were differences in NDVI during the three time periods (sum of squares = 6.76e-1, F = 127.49, p < 0.001), with NDVI from within the babbler home-range’s higher during December compared to both August and November (Dec–Aug: estimate = 9.12e-2, SE = 6.92e-3, t = −13.182, p < 0.001; Aug–Nov: estimate = 4.00e-3, SE = 6.86e-3, t = 0.584, p = 0.829; Dec–Nov: estimate = 9.52e-2, SE = 6.86e-3, t = 13.870, p < 0.001; Fig. 3A).

A Boxplots showing overall average NDVI from within babbler home-ranges for each time period in our study (significance indicated by asterisk). B Boxplots showing the difference between actual NDVI values of area used by babbler groups (i.e. from within home-ranges) as compared to the overall NDVI (set to ‘0’ and indicated by dotted line), by habitat type for each time period in our study.
The NDVI of the village habitat from within babbler home-ranges compared to the overall village NDVI was not significantly different in August, was lower in November, and was higher in December (August: mean NDVIdiff = -1.40e-2, SD = 5.45e-2, p = 0.145; November: NDVIdiff = −2.30e-2, SD = 3.39e-2, p = 0.001; December: NDVIdiff = 7.60e-2, SD = 3.39e-2, p < 0.001). The NDVI of orchard habitat from within babbler home-ranges was lower than expected in August and November, with no difference in December (August: NDVIdiff = -8.43e-2, SD = 4.78e-2, p < 0.01; November: NDVIdiff = −1.01e-1, SD = 3.80e-2, p < 0.001; December: NDVIdiff = 9.67e-3, SD = 5.01e-2, p = 0.254). However, the NDVI of the plateau habitat from within babbler home-ranges was always higher than the overall plateau NDVI (August: NDVIdiff = 5.18e-2, SD = 7.83e-2, p < 0.001; November: NDVIdiff = 4.15e-2, SD = 5.22e-2, p < 0.001; December: NDVIdiff = 1.26e-1, SD = 4.16e-2, p < 0.001; Fig. 3B).
Group overlap
We found a significant change in the extent of home-range overlap (km2) over the study period (estimate = 8.72e-4 ± 4.30e-4, t = 2.033, p = 0.0426) when we analysed home-range overlap across all pairs of groups. We further examined changes in overlap separately for the two groups around the Midrasha and the eight groups around the Kibbutz (based on Fig. S1). We found no change in the amount of home-range overlap (km2) for the two groups around the Midrasha (estimate = −1.44e-3 ± 2.42e-4, t = −0.595, p = 0.554) but a significant increase in the amount of daily home-range overlap for the groups around the Kibbutz as the season changed (estimate = 9.27e-4 ± 4.61e-4, t = 2.010, p = 0.045; Fig. 4A). The groups around the Kibbutz also increased the number of daily home-range overlaps that occurred with other groups as the season changed from summer to winter (estimate = 3.44e-3 ± 8/35e-4, t = 4.116, p = 4.44e-5; Fig. 4B, C, D).

A The total amount of home-range overlap (km2) per day. B The total number of home-range overlaps each group had per day. C Graphical depiction of the number of overlaps between babbler groups during the first week of the study (Aug 18–24, 2022). D Graphical depiction of the number of overlaps between babbler groups during the last week of the study (Dec 25–31, 2022). Data shown are only for the 8 groups of Arabian babblers around Kibbutz Sde Boker in the Negev desert. *Kibbutz7 had not yet separated as its own group for the first week of our study. The X-axis (A, B) has been modified to show the actual date as opposed to scaled date values.
Discussion
As predicted, we found evidence that reliable resources in human villages were especially important during the summer when natural resources in the desert habitat were scarce. Despite no seasonal change in vegetation across the highlands (based on NDVI), as the season changed from summer to winter, groups spent more time outside of villages, had smaller home-ranges, and were less territorial (i.e. had greater overall area and number of home-range overlaps). We also show how human-induced disruptions to the landscape present regions of attraction (e.g. villages) and repulsion (e.g. the ‘no babbler’s land’ mentioned below). These areas potentially create a mosaic that is similar to the wadis (areas of attraction) and surrounding barren desert (areas of repulsion) in the babblers’ more natural habitat in the Arava.
Accurate data for the total distance an individual bird travels in a day is key to provide quantifiable links between behaviour and energetics39. High-throughput localisation data at high spatial resolution will necessarily lead to more accurate speed and distance calculations40. Many past studies have produced calculations for bird flight speeds during or outside of migration41,42, as well as greatest distance travelled by birds during migration42,43,44 or outside migration45,46. Nevertheless, there are currently very little data on the daily distance travelled outside of migration and breeding for passerine birds. The only somewhat comparable data are from Kirtland’s Warblers (Setophaga kirtlandii) where birds travelled up to 77 km between detections47. However, some of these data included measurements from >24 h periods, and the precision was at the scale of kilometres as opposed to our finer scale of 10–20 m47. Our use of such data enabled us to provide an improved idea of the total daily distance travelled by our tracked babblers, which in turn allowed us to better understand daily babbler group behaviour.
On average, babblers travelled 18 km a day, with a large amount of individual variation (min = 1.79 km, max = 97.32 km). Nevertheless, despite the distance travelled each day showing no significant change with the season, we did see a trend in winter toward less daily distance travelled alongside the significantly smaller home-ranges. This suggests a few possibilities: 1—in summer, resources are spread over larger distances, and birds are required to move further distances and cover more territory to fulfill their daily energy requirements, 2—birds are more protective of resources and recently fledged young in summer and their movement reflects safeguarding (or expanding) the edges of their main territory as a buffer against other groups, or 3—groups need more resources due to the presence of young birds (3–6 months old) during the summer period. The uncultivated agricultural fields (fallow fields consisting of mostly bare dirt) between Kibbutz Sde Boker and Midreshet Ben-Gurion seemed to present a ‘no babbler’s land’, where not one localisation was recorded over the 135-day period (see Fig. S4). Further, no babbler groups crossed this area over the following 6 months of monitoring (unpublished data), and so this ~2 km stretch of uncultivated agricultural land seemed to act as a barrier to group interactions, dispersal, and habitat use. While there is a large amount of literature on cleared land in forest ecosystems causing movement barriers48,49,50, cleared agricultural land in deserts may also act as barriers due to vegetative homogeneity and lack of high vegetation and shade51. Arabian babblers have fairly limited flight capabilities and use an elevated group ‘sentinel’ to act as a lookout for potential predators, and so are reliant on at least some vegetation and local flora25. Our discovery of babbler’s avoidance of such an open landscape reinforces the idea that Arabian babblers need a continuous expanse of at least a modicum of verdant land, such as well-spaced trees (≤500 m apart) and low bushes found within the naturally occurring wadis25. Thus, agricultural fields that remove natural vegetation, that could act as perches for sentinels, could hinder the dispersal and mobility of babblers between neighbouring regions.
The movement of babbler groups from villages to the plateau as the season changed from summer to winter was somewhat expected, as we had assumed babblers would make use of village resources only when the resources on the plateau were very limited. While we found no evidence for an overall ‘greening’ of the highlands based on NDVI, we did find evidence that in all habitats, babblers were using areas of higher NDVI in December compared to August. Specifically, groups were spending time in areas of the plateau habitat with the highest NDVI, and, thus, presumably, the highest levels of food availability. Studies on the seasonal behaviour of native arthropods in the desert have shown beetle activity increases within daylight hours during the winter when temperatures are cooler52,53. Thus, in winter, babbler food would become more abundant during their hours of foraging activity in the surrounding desert. These factors suggest that in winter, birds would have to depend less on resources from human development (i.e. villages and orchards) as there may be an increase in key resources found in the babbler’s more natural habitat.
As babblers are mainly insectivorous, NDVI may not accurately reflect their specific food availability, as it is solely a measure of vegetation. However, the fact that in December, babblers began to spend more time in areas of the plateau with higher-than-average NDVI, indicates there was some aspect of the plateau habitat that increased in importance or attractiveness as the seasons changed. We suggest a few possibilities for the change in importance, being 1—the plateau reached a threshold of productivity in areas with higher NDVI (e.g. a change in insect activity such as mentioned above), 2—as breeding success tends to be higher for nests outside of villages54 birds may prefer favourable nest-sites options in the semi-natural plateau habitat as the breeding season approaches, 3—the ease and availability of open water in villages may have attracted them during the hotter and drier summer, when groups had fledglings or juveniles and was no longer as essential in the cooler winter. While data for this study were predominantly collected remotely, this species has been studied in the broader region (~50 km east in the Arava valley) for over 50 years24. However, we expect food type and availability from the Arava to reflect more natural food type and availability and suggest that future research into actual food type availability in each of our modified habitats could provide further insights into real applications of babblers’ habitat choice.
We found no evidence for larger group size resulting in larger home-ranges or increased use of high resource areas (i.e. villages). Group living is thought to be an adaptation to living in a harsh environment20,21. For babblers the benefits from larger group size may not only be due to resource/area defence, but instead, larger groups may lead to predation dilution (e.g. ref. 55) or occur solely during the breeding season (e.g. increased provisioning;56, and predator defence57). For this species, group size has been shown to have equivocal effects on various breeding dynamics (e.g. provisioning rate24,27,54,58,59,60), however, the effects of group size on predation have yet to be studied. We had expected to see either smaller groups having smaller home-ranges, or perhaps larger groups having more resource-abundant areas (i.e. greater use of villages), but neither was found in this study.
At its most basic level, territoriality involves protecting resources that maximise survival and reproduction61. During winter, groups spent less time concentrated in villages, so we expected less overlap among groups as they moved to the more expansive plateau habitat surrounding the villages. Instead, we found both the amount of overlap among groups and the number of group overlaps increased in the winter. This supports the idea that groups were more territorial in summer when key resources (e.g. open water) became scarce. Similar behaviour has been recorded in the ecologically similar Southern pied babbler (Turdoides bicolour) in the Kalahari, where groups became more territorial when resources were low or unreliable62.
The decrease in territoriality that we found (i.e. increase in a number of group overlaps) may have been a by-product of groups extending the edges of their main territories and exploring into neighbouring areas to scout out the resource situation while that group was occupied elsewhere—such as investigating potential nest sites for the upcoming breeding season. It may also be that as juveniles become fully bonded with their natal group, there is less risk of them being lured away, and so there is less reason to avoid overlapping home-ranges. Territoriality comes with a number of potential costs that involve energy expenditure on behalf of the group or individual: patrolling effort, advertising ownership, physical defence, increased risk of predation, and time away from other activities63. As such, the decrease in territoriality in winter could also allow birds to optimise their body condition as they prepare for the breeding season64. While our study occurred outside the breeding season, future work could examine group dynamics during the breeding season, where we would expect very high levels of territoriality shown by little to no overlap in home-ranges.
Anthropogenic modifications to habitats are increasing in scale and intensity, especially in desert environments13. Here we identified two main aspects of this anthropogenic change that may affect desert specialists such as babblers, namely, agricultural expansion and urbanisation.
Agricultural fields without at least a modicum of connectivity (e.g. vegetative margins, trees as perches), can create barriers to animal movements50,65,66,67. Here, we found that unvegetated fields may create ‘behavioural barriers’45 to babbler movement as they may disrupt their regular behavioural patterns (e.g. being unable to have a sentinel on lookout from a perch). A minor intervention, such as encouraging farmers to leave small patches of native vegetation (e.g. Vachellia tortilis, Retema raetam) at their field margins, may enable babbler movement between regions while using this vegetation. Such interventions would benefit overall gene flow and help avoid potential population bottlenecks that could be caused due to fragmented habitat.
The implications of small-scale urbanisation are more difficult to interpret in our system. The Sde Tsin highlands have a greater amount of modified habitats and support a greater abundance of babblers than the more natural Arava region. In the Arava, babbler group density ranged from 0.31–0.73 groups/km2 and 1.25–5.62 individuals/km2 from 1986–201268. Our study has less precise data on group-size. Nevertheless, we found that babbler group density was 0.81 groups/km2 and 7.40 individuals/km2. While this may seem beneficial for babblers, increased density is likely to have unexpected consequences, especially if it comes due to artificially provided resources. Resource availability can alter inter- and intra-group dynamics, in some cases causing increased aggression69,70,71,72, which we may expect to see in babblers26.
Using our unique combination of technology and study site, we were able to uncover some of the more complex group behavioural dynamics and highlight some key elements that link group behaviour to seasonality and habitat use. We were able to tease apart some of the complexities inherent in social behaviour, showing not only how groups make use of different habitats seasonally but also how resource availability alters inter-group interactions such as territoriality. We found no direct evidence that the overall level of vegetation increased on the semi-natural plateau habitat in winter, nevertheless, we did find groups moved onto the plateau in winter. Hence, village resources may be exploited by babblers predominantly during the harsher summer months. This exploitation is accompanied by increased territoriality, with groups less territorial in winter as they are able to find resources in the more expansive plateau habitat.
It is also noteworthy that the rising accessibility of high-throughput data collection techniques such as the ATLAS system was necessary to uncover both the small-scale daily movements and large-scale dynamics among babbler groups. Traditional methods are likely unable to uncover instances where groups made use of the same place but at different times of the day, as it is impractical for researchers to follow groups all day, every day. Moreover, traditional techniques would be unable to uncover the fact that even a small section of agricultural land may act as a movement barrier. As human modifications of the environment increase, new technologies could be instrumental to better understand how animals cope with a modified planet, and how best they can be protected.
Methods
Study site and species
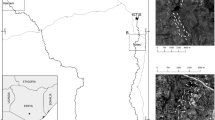
This study took place from 18 Aug–31 Dec 2022 on the Sde Tzin highlands (~12 km2) around Sde Boker, Israel (30°52'25“N 34°47'35“E, 486 m.a.s.l.). The Sde Tzin highlands encompass three villages of ~500–2000 residents: Midreshet Ben-Gurion, Kibbutz Sde Boker, and Merhav Am, along with infrastructure (water treatment plants, solar panels), agricultural fields, orchards, and semi-natural desert habitat (‘plateau’; Fig. 5A). This region was covered by an ATLAS system consisting of seven receiver stations placed around the Sde Tzin highlands in locations that maximise the ability to triangulate and record localisations of tags (Fig. 5). The ATLAS system provides localisation data with similar accuracy and precision to traditional GPS (within ~5 m of traditional GPS), at a faster sampling rate (~1–8 s), while being comparatively inexpensive, and with the ability to use very small tags (weighing as little as ~1 g73,74). The reverse-GPS technology of the ATLAS relies on the installation of a system of receivers usually limited to a regional scale (~100 km wide) that trilaterate (use distances to determine the coordinates of a point of interest) and record localisations of transmitting tags affixed to the study animals74.

A Map of the region covered by the ATLAS network showing simplified habitat type designations and locations of the ATLAS receiving stations. B An adult male babbler wearing an ATLAS tag attached via leg-loop harness. C Schematic showing placement of the ATLAS tag and leg-loop harness.
During this study, there were seven ATLAS receiver stations placed on the plateau surrounding Kibbutz Sde Boker (Fig. 5). Tags used in this study consisted of a CR1620 battery connected to a Vildehaye tag75 V 2.8, which was set to emit a signal on an 8-s interval. The tags and batteries were coated with epoxy resin for protection. Tags were activated in the field using a gas-powered soldering iron used to complete the battery circuit. ATLAS technology is already yielding novel insights into how animals interact with their environments through quantitative tracking resulting in copious amounts of big-data30,31,32,33. The technology allows for remote tracking of multiple animals, with tags small enough and light enough to place on ~20 g organisms74.
Based on our observations of babbler groups as part of a previous study in the same area54, we decided to focus on the 10 groups of babblers whose territories occurred solely within the region covered by the ATLAS system. We monitored babblers outside their main breeding season (Feb–June; see Supplementary Materials Fig. S1 for a visual representation of the territories). While an additional three groups made occasional use of the ATLAS region, they were not localised often enough for us to reliably include them in our analysis. Initial analysis of all localisations for each group indicated a large geographic divide occurring between the two southern groups centred around Midreshet Ben-Gurion and the eight groups around Kibbutz Sde Boker, and so our group designations were labelled as Midrasha1–2, and Kibbutz1–8 (Fig. S1; see Supplementary Materials Table S1 for group designations, and information on tag deployment, localisations, and days of data).
We divided the Sde Tzin highlands region into seven simplified habitat types based on high-resolution 2021 satellite imagery (govmap.gov.il). These habitats include: 1—plateau, semi-natural desert habitat with low human presence and no agriculture or irrigation, that included degraded land and more natural habitat adjacent to a nearby National Park; 2—agriculture, areas with seasonal cultivation grown by members of Kibbutz Sde Boker (e.g. cabbage, lettuce, etc.); 3—orchard, predominantly olive groves with one small argan grove; 4—village, the three regions with permanent human habitation including Midreshet Ben-Gurion, Kibbutz Sde Boker, and Merhav Am; 5—solar, fenced regions of solar panels; 6—poultry, fenced regions of poultry production; and 7—water treatment, fenced region with three large open water treatment pools (Fig. 5A). During the dates of this study, the agricultural fields were not cultivated (personal observations).
Capturing and tagging
Babblers were caught using 40 × 40 cm flap traps baited with giant mealworms (Tenebrio molitor). In total, we captured and tagged 21 babblers. Fourteen of these birds (three females, three males, eight juveniles) had been banded previously. The other seven were caught as unbanded birds (two females, three males, two juveniles) so these were first banded with a standardised aluminium ring (see Supplementary Materials Table S2 for demographic information on captured birds). We secured the ATLAS tags to each bird via leg-loop harness (tag and harness average mass = 2.7 g; average 3.6% of individual body mass; Fig. 5B, C). We made multiple trapping attempts to try and maintain at least one active tag in each of the 10 groups over the 134-day period—from Aug 18 to Dec 31, 2022. In seven groups, we collected continuous data throughout the 134-day period. Nevertheless, due to trapping difficulties, two groups had a single period with no active tag (Midrasha2 from Nov 15 to Dec 31 and Kibbutz2 from Nov 25 to Dec 9), and we began monitoring one additional group (Kibbutz7) on Nov 12 after it split from another group (Kibbutz4).
Bird harnesses were made using 0.8 mm silicone thread, thin wire shaped into a figure-eight loop, and a flexible jewellery fastener (see Supplementary Materials Fig. S2A). The harness was prepared by securing the middle of a ~ 40 mm piece of silicone thread and the middle of the wire to the tag. Each side of the remaining pieces of silicone thread was cut to be equal in length to ensure an even placement on the bird. The end silicone threads were then threaded through the wire figure-eight loop and gathered in the jewellery fastener, which was pinched to be tight enough the threads did not easily pull out but remained loose enough to be drawn firmly to the bird (Fig. S2A). To attach the harness to each bird, the tag was placed in the middle of the babbler’s back, and each leg-loop was drawn over its respective leg, using a 0.5 mm knitting needle to move the loop into the thigh if necessary. The threads were then pulled tight enough to remain secure but loose enough not to press into the skin, and the jewellery fastener was pinched tight enough the threads could no longer move (Fig. S2B). We then placed a small piece of cardboard under the tag, flattened the wire figure-eight loop and the exposed wires, and covered the entire tag and harness setup in epoxy (Fig. S2B). The entire process after capture took 5–10 min.
All birds were captured and handled with ethics approval by the Israeli National Parks Authority (permit number: 2022/43033). Ringing of birds was done with authorisation from the Israel Bird Ringing Centre (https://www.birds.org.il/en/birding-center/The-Israeli-Bird-Ringing-Center).
Climate data
Daily climate data for Aug 18–Dec 31, 2022, were downloaded from the Israeli Meteorological Service (ims.gov.il/en/data_gov) for the Sde Boker weather station. Maximum daily temperatures (Tmax, °C) were highly correlated with the day of the study (r = −0.941), where days became progressively cooler as the season changed from summer to winter. Consequently, we did not include Tmax as a separate predictor variable in our analysis. The first rainfall event occurred during late October (Oct 20 to 21, 2022; 7.4 mm), which is typical for the region76,77.
We downloaded mean NDVI (normalised difference vegetation index) data for the region from the Copernicus Sentinal2 satellite using Google Earth Engine (https://earthengine.google.com). Data were obtained for three time periods: A—August 18–24, 2022 (‘August’), B—October 27–November 3, 2022 (i.e. a week after the first rainfall; ‘November’), and C—December 15–31, 2022 (‘December’; see Fig. S3 for an overview of NDVI in each period). Data were downloaded at a 3 m2 resolution and filtered for cloudless data. We then extracted mean NDVI values for all three periods for each habitat type from within the home-ranges of each group of babblers for the days of each time period (see below) using ArcGIS Pro78. We further calculated the difference between the NDVI of each habitat type from within each group’s home-range (defined below) and the overall NDVI of that habitat type (NDVIdiff; see Table S3 for sample data).
Data analysis
All the following analyses were performed in the R Statistical Environment v 4.3.179 using RStudio 2023.06.280.
The ATLAS provides raw data that allows for a simple filter-smoothing process73. As we were interested in diurnal area use, we filtered out nocturnal localisations for each day based on sunrise and sunset data for Sde Boker. Following ref. 81 we then performed an initial exploratory analysis of data tracks, and minimised location error by filtering based on two location attributes added with toolsForAtlas: (1) we filtered the overall location quality for each localisation (‘traceNorm’; calculated from the variance in Easting and Northing) by creating histograms and removing the outliers (based on the ‘tail’), and (2) we filtered location error estimates (‘stdVarY’; variance-covariance matrix of each position) using a standard deviation of 20. We further filtered unrealistic movement by assuming a maximum speed of 5 m/s based on the behavioural traits of babblers (e.g. mainly hopping or walking on the ground) and observations from the nearby long-term study on babblers. We are further convinced 5 m/s is a realistic maximum speed as a study on Eurasian blackcap (Sylvia atricapilla) found tagged birds had a maximum speed of 3 m/s during non-migratory periods82.
We examined overall movement during the study period. We calculated a daily total distance travelled (km) per group as a sum of the distance between consecutive GPS points. For days when a group had more than one bird with an active tag, the more recently tagged bird was chosen to represent the group on that day. We calculated a 100% Local Convex Hull (LoCoH; km2, hereafter referred to as ‘home-range’) per day per group, to determine overall movement patterns throughout the study period. We chose LoCoHs, to create home-ranges, as this method takes time into account when creating density kernels and can more accurately represent habitat use in a heterogeneous landscape83,84,85. Home-ranges were calculated only for days with at least 100 localisations, specifying k = 12 (number of nearest neighbours to include) and setting duplicates to ‘random’ (where duplicated points are moved slightly by setting the duplicate value ‘d’, and randomly adding or subtracting d/5 degrees to their latitude/longitude).
We first investigated how season and group size predicted daily distance travelled and home-range. To address identifiability issues in our models resulting from large eigenvalues we first rescaled the variables ‘day of the study’ and ‘relative group size’ to have a standardised mean of 0 and standard deviation of 1. We then fitted a Generalised Linear Mixed-Effects Model (GLMER, with a gamma distribution, and a log-link function) separately to distance travelled (km) and home-range (km2) as predicted by the day of the study and the relative group size, with group ID as a random effect.
To see whether groups changed their use of habitat types throughout the study period, we calculated the total and proportional use of each habitat type (village, orchard, plateau) within each home-range per group per day. We focused solely on the three main habitats in the region as personal observations showed other habitats were either not used (e.g. agriculture creating the ‘no babbler’s land’ mentioned below in the discussion) or were not well-covered by the ATLAS (e.g. solar farms caused interference but also were seldom used by babblers; pers. obs.). We then fitted linear mixed-effects models (LMERs) separately to the total habitat use and proportional habitat use as predicted by the day of the study, with group ID as a random effect and a Gaussian distribution.
To identify if group seasonal movements were following vegetative productivity, we overlayed each group’s daily home-ranges for each time period (August, November, December) over mean NDVI data from each specific time period. We then fitted an LMER to NDVI as predicted by the time period, with the group as a random effect and a Gaussian distribution.
To see if babbler groups were potentially using areas of each habitat with either higher or lower NDVI than the overall NDVI of the habitat type, we ran one-sample t-tests to test if NDVIdiff was statistically different from ‘0’—with ‘0’ being set to the mean NDVI of each habitat type. We did this for each habitat type separately by time period.
To explore whether groups seasonally changed their home-range overlap with other groups throughout the study period, we calculated the overlap in home-ranges (km2) across all groups. We further calculated the number of overlaps each group had with any other group for each day. We fitted linear models (LMs) separately to the amount of overlap and number of group overlaps per day as predicted by the day of the study.
We used Wald tests (GLMER) Satterthwaite’s method t-tests (LMER), and standard t-tests (LM, one-sample t-test) to determine significance based on p < 0.05.
We used the following R packages for pre-processing and cleaning of individual tag data: amt86, dplyr87, suncalc88, lubridate89, and toolsForAtlas90; spatial data analysis and visualisations: amt, dplyr, toolsForAtlas, sp91, rgeos92, adehabitatHR93, rgdal94, geosphere95, raster96, and leaflet97; and mixed-effects model analyses and visualisations: amt, rgdal, sp, car98, lme499, lmerTest100, and ggplot2101.
Data availability
The datasets supporting this article have been uploaded as part of the supplementary materials.
Code availability
The dataset and R code supporting this article have been uploaded as part of the Supplementary Materials.
Change history
18 August 2025
A Correction to this paper has been published: https://doi.org/10.1038/s44185-025-00106-5
References
Sih, A., Ferrari, M. C. & Harris, D. J. Evolution and behavioural responses to human-induced rapid environmental change. Evol. Appl. 4, 367–387 (2011).
Otto, S. P. Adaptation, speciation and extinction in the Anthropocene. Proc. R. Soc. B Biol. Sci. 285, 20182047 (2018).
Bellard, C., Bertelsmeier, C., Leadley, P., Thuiller, W. & Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 15, 365–377 (2012).
IPCC. Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation (Cambridge University Press, Cambridge, UK and New York, USA, 2012).
Lockwood J. L., Hoopes M. F. & Marchetti M. P. Invasion Ecology (John Wiley & Sons, West Sussex, UK, 2013).
Salo, P., Korpimäki, E., Banks, P. B., Nordström, M. & Dickman, C. R. Alien predators are more dangerous than native predators to prey populations. Proc. R. Soc. B Biol. Sci. 274, 1237–1243 (2007).
Murali, G., Iwamura, T., Meiri, S. & Roll, U. Future temperature extremes threaten land vertebrates. Nature 615, 461–467 (2023).
Palumbi, S. R. Humans as the world’s greatest evolutionary force. Science 293, 1786–1790 (2001).
Berger-Tal, O. & Saltz, D. Invisible barriers: anthropogenic impacts on inter- and intra-specific interactions as drivers of landscape-independent fragmentation. Philos. Trans. R. Soc. B Biol. Sci. 374, 20180049 (2019).
Geffroy, B. et al. Evolutionary dynamics in the Anthropocene: Life history and intensity of human contact shape antipredator responses. PLoS Biol. 18, e3000818 (2020).
Leblond, M., Dussault, C. & Ouellet, J. -P. Avoidance of roads by large herbivores and its relation to disturbance intensity. J. Zool. 289, 32–40 (2013).
Murray, M. et al. Greater consumption of protein-poor anthropogenic food by urban relative to rural coyotes increases diet breadth and potential for human–wildlife conflict. Ecography 38, 1235–1242 (2015).
Sullivan B. K., Haverbeke D. R. V. & Chambers C. in Design with the Desert (CRC Press, Boca Raton, USA, 2013).
Reynolds, J. F. et al. Global Desertification: building a science for dryland development. Science 316, 847–851 (2007).
Lewin, A., Murali, G., Rachmilevitch, S. & Roll, U. Global evaluation of current and future threats to drylands and their vertebrate biodiversity. Nat. Ecol. Evol 1–11 https://doi.org/10.1038/s41559-024-02450-4 (2024).
Prăvălie, R. Drylands extent and environmental issues. A global approach. Earth-Sci. Rev. 161, 259–278 (2016).
Davison, C. W., Rahbek, C. & Morueta-Holme, N. Land-use change and biodiversity: Challenges for assembling evidence on the greatest threat to nature. Glob. Change Biol. 27, 5414–5429 (2021).
Crawford, C. S. & Gosz, J. R. Desert ecosystems: their resources in space and time. Environ. Conserv. 9, 181–195 (1982).
Nandintsetseg, D., Kaczensky, P., Ganbaatar, O., Leimgruber, P. & Mueller, T. Spatiotemporal habitat dynamics of ungulates in unpredictable environments: The khulan (Equus hemionus) in the Mongolian Gobi desert as a case study. Biol. Conserv. 204, 313–321 (2016).
Zahavi, A. & Zahavi, A. The Handicap Principle: A Missing Piece of Darwin’s Puzzle. (Oxford University Press, Cambridge, UK, 1999).
Keynan, O., Ridley, A. R. & Lotem, A. Social foraging strategies and acquisition of novel foraging skills in cooperatively breeding Arabian babblers. Behav. Ecol. 26, 207–214 (2015).
Anava, A., Kam, M., Shkolnik, A. & Degen, A. A. Effect of group size on field metabolic rate of Arabian babblers provisioning nestlings. Condor 103, 376–380 (2001).
Naguib, M. et al. Cooperatively breeding Arabian babblers call differently when mobbing in different predator-induced situations. Behav. Ecol. 10, 636–640 (1999).
Zahavi, A. in Cooperative Breeding in Birds: Long Term Studies of Ecology and Behaviour (eds P. B. Stacey & W. D. Koenig) 103–130 (Cambridge University Press, Cambridge, UK, 1990).
Wright, J., Berg, E., De Kort, S. R., Khazin, V. & Maklakov, A. A. Cooperative sentinel behaviour in the Arabian babbler. Anim. Behav. 62, 973–979 (2001).
Ostreiher, R. in The Arava—Landscape, Nature. and People in the Arave Valley (ed. Y. Giladi) 262–273 (Arava Publishing, Ein Yahav, Israel (in Hebrew), 2013).
Carlisle, T. R. & Zahavi, A. Helping at the nest, allofeeding and social status in immature arabian babblers. Behav. Ecol. Sociobiol. 18, 339–351 (1986).
Zahavi, A. Communal nesting by the Arabian Babbler: a case of individual selection. Ibis 116, 84–87 (1974).
Toledo, S. et al. Cognitive map–based navigation in wild bats revealed by a new high-throughput tracking system. Science 369, 188–193 (2020).
Corl, A. et al. Movement ecology and sex are linked to barn owl microbial community composition. Mol. Ecol. 29, 1358–1371 (2020).
Lourie, E., Schiffner, I., Toledo, S. & Nathan R. 2021 Memory and conformity, but not competition, explain spatial partitioning between two neighboring fruit bat colonies. Front. Ecol. Evol. 9, 732514 (2021).
Vilk, O. et al. Ergodicity breaking in area-restricted search of avian predators. Phys. Rev. X 12, 031005 (2022).
Luisa Vissat, L., Cain, S., Toledo, S., Spiegel, O. & Getz, W. M. Categorizing the geometry of animal diel movement patterns with examples from high-resolution barn owl tracking. Mov. Ecol. 11, 15 (2023).
Efrat, E. Man and the Environment in Israel—Geographical Essays (Ramot Publishing, Tel Aviv University, Tel Aviv, Israel: Ramot Publishing, 2004).
Ostrovsky, G., Kozer, R. & Garfinkel, M. Garbage and Recycling in Israel: a Report and Recommendations. (Adam Teva Din, Tel Aviv, 2009).
Yom-Tov, Y., Hatzofe, O. & Geffen, E. Israel’s breeding avifauna: a century of dramatic change. Biol. Conserv. 147, 13–21 (2012).
Anderies, J. M., Katti, M. & Shochat, E. Living in the city: resource availability, predation, and bird population dynamics in urban areas. J. Theor. Biol. 247, 36–49 (2007).
Shochat, E. Credit or debit? Resource input changes population dynamics of city-slicker birds. Oikos 106, 622–626 (2004).
Carbone, C., Cowlishaw, G., Isaac, N. J. B. & Rowcliffe, J. M. How far do animals go? Determinants oF Day Range in Mammals. Am. Nat. 165, 290–297 (2005).
Noonan, M. J. et al. Scale-insensitive estimation of speed and distance traveled from animal tracking data. Mov. Ecol. 7, 35 (2019).
Hedenström, A. & Alerstam, T. Optimal flight speed of birds. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 348, 471–487 (1997).
Hedenström, A. & Alerstam, T. How fast can birds migrate?. J. Avian Biol. 29, 424–432 (1998).
Hall-Karlsson, K. S. S. & Fransson, T. How far do birds fly during one migratory flight stage?. Ringing Migr. 24, 95–100 (2008).
Leshem, Y. & Yom-Tov, Y. Routes of migrating soaring birds. Ibis 140, 41–52 (2008).
Harris, R. J. & Reed, J. M. Behavioral barriers to non-migratory movements of birds. Ann. Zool. Fenn. 39, 275–290 (2002).
Poessel, S. A. et al. Interpreting long-distance movements of non-migratory golden eagles: prospecting and nomadism? Ecosphere 13, e4072 (2022).
Cooper, N. W. & Marra, P. P. Hidden long-distance movements by a migratory bird. Curr. Biol. 30, 4056–4062.e3 (2020).
Kuefler, D., Hudgens, B., Haddad, N. M., Morris, W. F. & Thurgate, N. The conflicting role of matrix habitats as conduits and barriers for dispersal. Ecology 91, 944–950 (2010).
Pomara, L. Y., Ruokolainen, K. & Young, K. R. Avian species composition across the Amazon River: the roles of dispersal limitation and environmental heterogeneity. J. Biogeogr. 41, 784–796 (2014).
Sweaney, N., Lindenmayer, D. B. & Driscoll, D. A. Movement across woodland edges suggests plantations and farmland are barriers to dispersal. Landsc. Ecol. 37, 175–189 (2022).
Munguia-Vega, A., Rodriguez-Estrella, R., Shaw, W. W. & Culver, M. Localized extinction of an arboreal desert lizard caused by habitat fragmentation. Biol. Conserv. 157, 11–20 (2013).
Cloudsley-Thompson, J. Thermal and water relations of desert beetles. Naturwissenschaften 88, 447–460 (2001).
Holm, E. & Edney, E. B. Daily activity of namib desert arthropods in relation to climate. Ecology 54, 45–56 (1973).
Oswald, K. N., Berger-Tal, O. & Roll, U. Small-scale land-use change effects on breeding success in a desert-living social bird. Behav. Ecol. 35, arae023 (2024).
Sorato, E., Gullett, P. R., Griffith, S. C. & Russell, A. F. Effects of predation risk on foraging behaviour and group size: adaptations in a social cooperative species. Anim. Behav. 84, 823–834 (2012).
Riehl, C. Evolutionary routes to non-kin cooperative breeding in birds. Proc. R. Soc. B Biol. Sci. 280, 20132245 (2013).
Ridley, A. R. in Cooperative Breeding in Vertebrates: Studies in Ecology, Evolution and Behaviour (eds Koenig, W. D. & Dickinson, J. L) 115–132 (Cambridge University Press, Cambridge, 2016).
Wright, J. Helping-at-the-nest in Arabian babblers: signalling social status or sensible investment in chicks?. Anim. Behav. 54, 1439–1448 (1997).
Wright, J. Helpers-at-the-nest have the same provisioning rule as parents: experimental evidence from play-backs of chick begging. Behav. Ecol. Sociobiol. 42, 423–429 (1998).
Wright, J. Helping-at-the-Nest and Group Size in the Arabian babbler Turdoides squamiceps. J. Avian Biol. 29, 105–112 (1998).
Brown, J. L. The evolution of diversity in avian territorial systems. Wilson Bull. 76, 160–169 (1964).
Golabek, K., Ridley, A. & Radford, A. Food availability affects strength of seasonal territorial behaviour in a cooperatively breeding bird. Anim. Behav. 83, 613–619 (2012).
Ord, T. J. Costs of territoriality: a review of hypotheses, meta-analysis, and field study. Oecologia 197, 615–631 (2021).
Anava, A., Kam, M., Shkolnik, A. & Degen, A. A. Seasonal field metabolic rate and dietary intake in Arabian Babblers (Turdoides squamiceps) inhabiting extreme deserts. Funct. Ecol. 14, 607–613 (2000).
Chetkiewicz, C.-L. B., St. Clair, C. C. & Boyce, M. S. Corridors for conservation: integrating pattern and process. Annu. Rev. Ecol. Evol. Syst. 37, 317–342 (2006).
Bennett, A. F. Linkages in the Landscape: The Role of Corridors and Connectivity in Wildlife Conservation. (IUCN, Gland, Switzerland and Cambridge, UK, 2003).
Rudnick, D. et al. The role of landscape connectivity in planning and implementing conservation and restoration priorities. Issues Ecol. 16, 1–23 (2012).
Keynan, O. Effect of group size and composition on individual behavior, group dynamics and population regulation in the Arabian babbler (Turdoides squamiceps). PhD Thesis, Macquarie University. See https://figshare.mq.edu.au/articles/thesis/Effect_of_group_size_and_composition_on_individual_behavior_group_dynamics_and_population_regulation_in_the_Arabian_babbler_Turdoides_squamiceps_/19430942 (2022).
Murray, B. G. The ecological consequences of interspecific territorial behavior in birds. Ecology 52, 414–423 (1971).
Foltz, S. L. et al. Get off my lawn: increased aggression in urban song sparrows is related to resource availability. Behav. Ecol. 26, 1548–1557 (2015).
Brown, J. L. Territorial behavior and population regulation in birds: a review and re-evaluation. Wilson Bull. 81, 293–329 (1969).
Wingfield, J. C. Control of territorial aggression in a changing environment. Psychoneuroendocrinology 19, 709–721 (1994).
Beardsworth, C. E. et al. Validating ATLAS: a regional-scale high-throughput tracking system. Methods Ecol. Evol. 13, 1990–2004 (2022).
Nathan, R. et al. Big-data approaches lead to an increased understanding of the ecology of animal movement. Science 375, eabg1780 (2022).
Toledo, S. et al. Vildehaye: a family of versatile, widely-applicable, and field-proven lightweight wildlife tracking and sensing tags. In: 2022 21st ACM/IEEE International Conference on Information Processing in Sensor Networks (IPSN) 1–14 (IEEE, 2022).
Alon, A. editor. Plants and Animals of the Land of Israel: An Illustrated Encyclopedia—Introductions and Indexes. See https://bookgallery.co.il/content/english/bookpageschema.asp?BookPageID=96677 (Tel Aviv, Israel: Ministry of Defense Publishing, 1990).
Zohary, M. Geobotany. 2nd edn. (Hapoalim Ltd., Holon, Israel, 1959)
ESRI. ArcGIS Desktop: Release 10.8.2. (Environmental Systems Research Institute, Redlands, Canada, 2021).
R Core Team. R: A Language and Environment for Statistical Computing (R Core Team, 2022).
RStudioTeam. RStudio: Integrated Development for R. (RStudio, Inc., Boston, MA, 2020).
Gupte, P. R. et al. A guide to pre-processing high-throughput animal tracking data. J. Anim. Ecol. 91, 287–307 (2020).
Tomotani, B. M., Bil, W., van der Jeugd, H. P., Pieters, R. P. M. & Muijres, F. T. Carrying a logger reduces escape flight speed in a passerine bird, but relative logger mass may be a misleading measure of this flight performance detriment. Methods Ecol. Evol. 10, 70–79 (2019).
Getz, W. M. et al. LoCoH: nonparameteric kernel methods for constructing home ranges and utilization distributions. PLoS ONE 2, e207 (2007).
Scull, P., Palmer, M., Frey, F. & Kraly, E. A comparison of two home range modeling methods using Ugandan mountain gorilla data. Int. J. Geogr. Inf. Sci. 26, 2111–2121 (2012).
Lyons, J. E. et al. Population size and stopover duration estimation using mark–resight data and Bayesian analysis of a superpopulation model. Biometrics 72, 262–271 (2016).
Signer, J., Fieberg, J. & Avgar, T. Animal movement tools (amt): R package for managing tracking data and conducting habitat selection analyses. Ecol. Evol. 9, 880–890 (2019).
Wickham, H., Francois, R., Henry, L., Muller, K. & Vaughan D. dplyr: a grammar of data manipulation. R package version 1.1.4. https://CRAN.R-project.org/package=dplyr (2023).
Thieurmel, B. & Elmarhraoui, A. suncalc: compute sun position, sunlight phases, moon position and lunar phase. R package version 0.5.1. https://CRAN.R-project.org/package=suncalc (2022).
Wickham, H. & Grolemund, G. Dates and times made easy with lubridate. J. Stat. Softw. 40, 1–25 (2011).
Margalit, S. toolsForAtlas: useful tools for atlas users. R package version 0.0.0.9004. http://rdrr.io/github/sivanMargalit/toolsForAtlas/ (2021).
Bivand, R., Pebesma, E. & Gomez-Rubio, V. Applied Spatial Data Analysis with R. 2nd edn. (Springer, New York, NY, USA, 2013). See https://asdar-book.org/ (Springer, New York, NY, USA, 2013).
Bivand, R. & Rundel, C. rgeos: Interface to Geometry Engine—Open Source (‘GEOS’). R Package Version 06-4. https://CRAN.R-project.org/package=rgeos (2023).
Calenge, C. & Fortmann-Roe, S. adehabitatHR: Home Range Estimation. R package version 0.4.21, https://CRAN.R-project.org/package=adehabitatHR (2023).
Bivand, R., Keitt, T. & Rowlingson, B. rgdal: Bindings for the ‘Geospatial’ Data Abstraction Library. R package version 1.6-7, https://CRAN.R-project.org/package=rgdal (2023).
Hijmans, R. J. geosphere: Spherical Trigonometry. R package version 1.5-18, https://CRAN.R-project.org/package=geosphere (2022).
Hijmans, R. J. raster: Geographic Data Analysis and Modeling. R package version 3.6-26, https://CRAN.R-project.org/package=raster (2023).
Cheng, J., Schloerke, B., Karambelkar, B. & Xie, Y. leaflet: Create Interactive Web Maps with the JavaScript ‘Leaflet’ Library. R package version 2.2.2, https://CRAN.R-project.org/package=leaflet (2023).
Fox, J. & Weisberg, S. An R Companion to Applied Regression, Third Edition. See https://socialsciences.mcmaster.ca/jfox/Books/Companion/. (Sage, Thousand Oaks, CA, USA, 2019).
Bates, D., Maechler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48 (2015).
Kuznetsova, A., Brockhoff, P. B. & Christensen, R. H. B. Package ‘lmerTest’. J. Stat. Softw. 82, 1–26 (2017).
Wickham, H. ggplot2: Elegant Graphics for Data Analysis. See https://ggplot2.tidyverse.org. (Springer-Verlag, New York, NY, USA, 2016).
Acknowledgements
We thank the people in the region around Kibbutz Sde Boker, Midreshet Ben-Gurion, and Merhav-Am for their cooperation in the project, for allowing us to set up antennas, and for showing interest in the project. We further thank Shmuel Dovrat (Ben-Gurion University) for communicating with Cellcom Israel Ltd. and Phi-Networks Communications Company, who we thank for allowing us to place antennas on their infrastructure. Lastly, we would like to thank the many people who contributed to data collection and helped with analysis and general information: Darren Burns, Emanuelle Lourie, Ofir Altstein, Amir Lewin, Alex Alaman, Marie Guggenberger, Nikola Dragic, Orr Spiegel, Michal Handel, Yael Lenhardt, and Ron Efrat. KNO is supported by a scholarship from the Zuckerman Scholars Programme.
Author information
Authors and Affiliations
Contributions
K.N.O., U.R. & O.B.T.—conceptualised the project and the methodology. K.N.O.—wrote the main manuscript text, collected the data, and did the main analyses. G.O.C. & U.R.—provided additional analyses. T.R., S.T. & R.N.—provided resources, software and methodology. G.O.C., U.R., O.B.T., O.K. & G.O.C.—reviewed and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Oswald, K.N., Rozenberg, T., Keynan, O. et al. The value of human resources changes with season for a social desert passerine bird. npj biodivers 4, 15 (2025). https://doi.org/10.1038/s44185-025-00078-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s44185-025-00078-6
