
Abstract
Persistent bacterial lung infections in children lead to significant morbidity and mortality due to antibiotic resistance. In this paper, we describe how phage therapy has shown remarkable efficacy in preclinical and clinical studies, demonstrating significant therapeutic benefits through various administration routes. Ongoing trials are evaluating its safety and effectiveness against different pathogens. Advancing phage therapy through systematic studies and international collaboration could provide a viable alternative to traditional antibiotics for persistent infections.
Similar content being viewed by others


Introduction
Persistent bacterial lung infections, including pneumonia, are the primary cause for hospitalization, morbidity, and mortality globally, accounting for over 40 million cases reported annually, and over 650,000 fatalities in children1. Persistent bacterial lung infections in children are often associated with underlying diseases including cystic fibrosis (CF), immune deficiencies, congenital pulmonary or cardiac anomalies, primary ciliary dyskinesia, and secondary lung damage from severe pneumonia, food or foreign bodies aspiration and chronic obstructive pulmonary diseases, which can hinder pathogens removal from the airways2. The hallmarks of persistent lung infections are intervals of stability interspersed with acute exacerbations of lung infection caused by bacterial pathogens including Pseudomonas aeruginosa, Acinetobacter baumanii, Klebsiella pneumoniae, Escherichia coli, Burkholderia cenocepacia, Achromobacter xylosoxidans, Staphylococcus aureus, and Mycobacterium abscessus complex respectively3,4,5,6,7,8,9,10. Because of the increasing selection for antibiotic resistance after prolonged exposure to broad-spectrum antibiotics, these pathogens display an important clinical issue of bacterial persistence even under antimicrobial therapy3. Furthermore, the microbes causing persistent lung infections are reported to develop in airways as microcolonies that aggregate into biofilms, which makes them more tolerant to antibiotics3. Although systemic or localized inhaled antibiotic therapy can reduce the pulmonary bacterial burden, insufficient antibiotic penetration to deeper lung areas and the bacterial capabilities to grow slowly or remain dormant inside lung epithelial and immune cells additionally contribute to the establishment of persistent lung infections3,11. Meanwhile, children with compromised immunity frequently experience persistent lung infections due to limited immune-mediated bacterial clearance which allows bacteria to persist even in the presence of antibiotic12. The combination of these adaptive modifications exhibited by pulmonary pathogens makes antibiotic therapy unsuccessful in overcoming persistent lung infections in children, leading to progressive, irreversible lung damage with impaired pulmonary functions13. This emphasizes the importance of finding a viable alternative therapy that will maximize the effectiveness of infection management in these pediatric patients. Owing to the growing issue of antibiotic resistance and limited possibilities for discovering novel antibiotics, bacteriophage therapy is increasingly reported as a promising alternative therapy for conquering persistent bacterial lung infections in children9,14.
Bacteriophage (phage)
Bacteriophages (phages) are viruses that specifically target bacteria by binding to their unique surface receptors, injecting their genomes to replicate within the bacteria, and inducing bacterial lysis to release new virions which infect neighboring bacteria, thereby initiating their new lytic cycles15,16,17,18,19,20. Because these lytic phages preferentially kill their hosts while replicating, they represent tremendous potential as effective and targeted alternative therapy to antibiotics in combating bacterial infections, especially those triggered by antibiotic-resistant bacteria involved in persistent bacterial lung infections15,16. Meanwhile, temperate phages enter lysogenic cycles by integrating as prophages into bacterial genomes, allowing a significant proportion of bacteria to withstand phage-induced lysis15,16. Temperate phages can be genetically modified by deleting their repressor genes, which prevents them from entering lysogenic cycles and allows them to be used therapeutically to induce target bacterial lysis while avoiding the risks of lysogeny or transfer of virulent and antibiotic-resistant genes15,16. Currently, preclinical studies have been conducted to assess the safety and effectiveness of phage therapy using the clinically relevant animal models of acute and chronic lung infections, with the goal of validating the clinical applications of phage therapy as targeted therapy in overcoming bacterial lung infections.
Preclinical studies of phage therapy against Gram-negative bacteria
Phages-PAK-P1 and PAK-P3 demonstrated significant potential for treating and preventing acute multidrug-resistant (MDR) P. aeruginosa lung infections in mice, according to previous studies21,22 (Table 1). The same bioluminescent P. aeruginosa strain was subsequently utilized in another study to develop acute murine lung infection and assessed activities of 9 distinct phages in vitro and in vivo23 (Table 1). Although seven phages (PAK_P1, PAK_P2, PAK_P3, PAK_P4, PAK_P5, LBL3, LUZ19) demonstrated an excellent correlation between in vitro and in vivo activities for improving survival with a decrease in bacterial load signal of luminescence in treated animals, two phages (PhiKZ, CHA_P1) were insufficiently active in vivo, emphasizing the importance of isolating phages directly on its targeted host to clinically apply phage as personalized therapy for achieving its therapeutic significance in pulmonary infections23 (Table 1). Meanwhile, intranasal phage-YH6 treatment was demonstrated to successfully decrease the bacterial load in internal organs including lungs, and rescued mice from death while treating murine hemorrhagic pneumonia triggered by MDR P. aeruginosa. Moreover, there were no inflammation or other pathological alterations in lungs of the mice following YH6 treatment24 (Table 1). When treating MDR P. aeruginosa-associated lung infections, either immediately or 12-h post- infection, an intranasal phage-MMI-Ps1 treatment resulted in significantly longer survival in treated animals with a lower bacterial burden in their lungs and blood25 (Table 1). Intranasal Bϕ-R656 or Bϕ-R1836 treatments improved animal survival and bacterial clearance in extensively drug-resistant (XDR) P. aeruginosa-induced murine pneumonia, with minimal histologic damages and no adverse consequences26. The phage-treated group showed higher levels of TNF-α and IL-6 compared to the negative control group, but still lower than the untreated group26 (Table 1).
According to an earlier study, intratracheal administration of PEV31 significantly reduced bacterial load and proinflammatory cytokines in the lungs of mice having P. aeruginosa-associated lung infection27 (Table 1). When mice with acute pneumonia caused by P. aeruginosa-PA01 were given intratracheal aerosolized phage-vB_PaeP_PA01EW, their lung tissues showed significantly lower bacterial burden, fewer inflammatory cell infiltration, and minimal pulmonary congestion as compared the control group28 (Table 1). In acute lung infection caused by P. aeruginosa-FADDI-PA001, intratracheal inhalation of phage-PEV31 at three distinct dosages (7.5 × 104, 5 × 106, and 5 × 108 PFU per mouse) effectively decreased the pulmonary bacterial burden, regardless of phage dosage and triggered a dose-dependent reduction of pulmonary inflammatory cytokines29 (Table 1). The bactericidal activities of PEV20 in a dry-powder formulation were also significantly effective when sprayed directly into the trachea of mice with MDR P. aeruginosa-induced lung infection, implying that pulmonary delivery of dry-powder formulation is an additional feasible therapeutic option for acute P. aeruginosa-associated lung infections30 (Table 1). When the efficacy of the phage-derived lysin was further tested in the treatment of acute murine P. aeruginosa pneumonia, PlyPa9 given both intratracheally and intranasally resulted in higher animal survival rates (70%) than its intranasal therapy (20%), emphasizing the therapeutic significance of phage’s product-lysin and its administration route for improving effectiveness in acute bacterial lung infections31 (Table 1). In mice with P. aeruginosa-associated acute and chronic pneumonia, both intravenous and intratracheal administration of phages-MYY9 or HX1, resulted in a significant reduction in bacterial burden, with fewer inflammatory responses and pathological damage to the lungs32 (Table 1). In murine chronic lung infection caused by MDR P. aeruginosa-LESB65 wild type and its adapted strain-NP22_2, intranasal administration of phage-PELP20 up to 7 days post-infection was reported to exhibit significant therapeutic efficacy to reduce pulmonary bacterial burden33 (Table 1). The findings of these studies revealed the significant efficacy and safety of phage therapy, irrespective of the administration route or formulation, in treating both acute and chronic P. aeruginosa-associated lung infections.
Importantly, P. aeruginosa in the murine lung was successfully eradicated by a combination of myovirus (φNH-4) and podovirus (φMR299-2) administered intranasally, indicating the therapeutic efficacy of intranasal phage cocktail treatment for P. aeruginosa-associated lung infections34 (Table 1). Intranasal phage cocktail therapy delivered simultaneously, 24 h-post infection and prophylactically (48 h prior to infection), was also observed to be efficient in eradicating P. aeruginosa-PAO1 in acute murine lung infection, and lowering inflammation, resulting in preventing the systemic dissemination of pathogen irrespective of when it was administered35 (Table 1). In addition to displaying in vitro lytic activities against P. aeruginosa in both planktonic and biofilms, the cocktail containing six phages (PYO2, DEV, E215, E217, PAK_P1, and PAK_P4) demonstrated dose-dependent curative effects for reducing the respiratory bacterial burden in acute pneumonia, suggesting a potent synergistic effect of the combined phage cocktail in pulmonary infections36 (Table 1). Meanwhile, intraperitoneal phage cocktail treatment for acute pulmonary infection of P. aeruginosa failed to protect mice from dying, despite a notable reduction in the bacterial burden in the lungs and spleen37. However, when a phage cocktail (PaAH2ΦP, PaBAP5Φ2, and PaΦ134) was given both intraperitoneally and intratracheally at 3 h after bacterial challenge, 100% of the mice revealed no mortality along with a significant decrease in bacterial load and pulmonary damage37. Remarkably, intratracheal phage cocktail monotherapy effectively prevented all mice from dying, and reduced lung bacterial burden and pathologic abnormalities even it was provided 6 h after bacterial challenge37 (Table 1). When a phage cocktail (PP1450, PP1777, and PP1902) was administered via nebulization during mechanical ventilation to treat porcine ventilator-associated pneumonia (VAP) caused by P. aeruginosa, the bacterial load was rapidly and significantly decreased, and the phages were distributed uniformly throughout the treated animals’ lungs38 (Table 1). These studies suggest that inhaled phage cocktail therapy offers therapeutic benefits for both non-ventilated and ventilated patients with P. aeruginosa-related lung infections.
In a prior study, phage therapy was discovered to improve the efficiency of complement-mediated pathogen elimination in vitro, underscoring the significance of a functioning complement system for complete eradication of P. aeruginosa during phage therapy25 (Table 1). When the effects of host immunity on phage therapy in acute P. aeruginosa-induced pneumonia were investigated by comparing treatment responses in healthy immunocompetent, MyD88-deficient, lymphocyte-deficient, and neutrophil-depleted mice, the neutrophil-phage synergy was crucial for providing the curative and prophylactic efficacies of intranasal phage therapy in respiratory infection39 (Table 1).
Furthermore, phage therapy was observed to modify the bacteria’s antimicrobial susceptibility profiles in vitro, making them more susceptible to tobramycin, ciprofloxacin, and amikacin, but less susceptible to colistin, which may in part be related to the mechanism of action of colistin (outer membrane) and their interaction with the phage receptors27 (Table 1). Because epithelial proliferation in alveoli and immune cell infiltration in alveolar walls and bronchi were identified despite phage therapy, the prior study proposed combining phages with antibiotics for improving clinical outcomes28 (Table 1). Interestingly, delivering an intraperitoneal phage cocktail in conjunction with meropenem at sub-inhibitory dosage subcutaneously improved animal survival (>50%), suggesting the potential additive outcome of a phage-antibiotic combination in treating murine pulmonary infections37 (Table 1). Following nebulization with air-jet or vibrating mesh nebulizers, aerosolization of the PEV20-ciprofloxacin combination was observed to retain synergistic antibacterial efficacy against MDR P. aeruginosa isolates in vitro40. While mice treated with phage or ciprofloxacin monotherapy showed no reduction in bacterial burden or inflammation, intratracheal aerosolization of a dry powder formulation containing PEV20-ciprofloxacin combination dramatically decreased pulmonary bacterial load and inflammatory responses in acute lung infection41 (Table 1). Intranasal inhalation of a triple combination therapy containing phage-KKP10, ceftazidime/avibactam, and CaEDTA was recently discovered to be significantly effective in managing chronic P. aeruginosa lung infections, resulting in 100% survival with complete pulmonary bacterial clearance in treated animals, and reducing bacterial pathogenicity and virulence19 (Table 1). These studies demonstrate the significant therapeutic potential of phage-antibiotic synergy in overcoming acute and chronic P. aeruginosa-associated lung infections.
A prior study revealed that higher phage doses were linked to a greater development of phage resistance in a P. aeruginosa subpopulation recovered from the lungs of neutropenic mice with acute pulmonary infection29. The phage-resistant P. aeruginosa displayed modified bacterial pathogenicity in vitro, such as increased ciprofloxacin susceptibility, decreased twitching motility, and reduced synthesis of blue-green pigment29. According to a prior study, P. aeruginosa-PAO1 developed phage resistance in vitro associated with an increase in pyomelanin synthesis, a greater susceptibility, and a faster rate of killing by peptides-LL-37 and colistin, after being exposed to phages-AM.P2, Mat, and Kat42. In human blood, serum, and a mouse pneumonia model, these phage-resistant mutants with genomic deletions showed significantly lower survival than its wild type P. aeruginosa-PAO1, suggesting the evolutionary trade-offs from phage resistance that reduce bacterial pathogenicity and increase susceptibility to immune-mediated clearance42. However, the emergence of phage-resistant variants in the presence of immune signaling deficiency was documented in a prior study, highlighting the importance of complementing phage therapy to maximize its effectiveness in immunodeficient individuals with P. aeruginosa-associated lung infections39.
Previous studies on mice with A. baumannii-induced pneumonia found that administering phages-Bϕ-C62 or Bϕ-R2096 intranasally at 30 min after infection resulted in 100% animal survival, complete bacterial clearance, a decrease in pulmonary proinflammatory cytokines, and an improvement in histological damage without causing adverse effects, revealing the therapeutic potential of phage therapy for overcoming pneumonia caused by carbapenemase-producing A. baumannii43,44 (Table 1). In a prior investigation, mice with lung infections were given time-dependent phage SH-Ab15519 treatment intranasally at 1 or 2 h after carbapenem-resistant A. baumannii (CRAB) challenge45. Intranasal phage therapy effectively lowered proinflammatory cytokines and bacteria in the lungs while also protecting mice from fatal infections45. The promising efficacy of intranasal phage SH-Ab15519 therapy in resolving CRAB-associated lung infection was further evidenced histopathologically by reduced pulmonary congestion and inflammatory cell invasion45. Notably, mice that received early phage therapy shortly after infection survived longer than mice that underwent delayed treatment, indicating that phage therapy requires prompt intervention to be successful45 (Table 1). Intranasal delivery of phage-vB_AbaM-IME-AB2 demonstrated dose- and time-dependent therapeutic effectiveness, with the highest beneficial effects at MOI-10 given 1-h after infection, in preventing 100% of mice against fatal A. baumannii-associated pneumonia46. Mice treated with phage revealed no inflammatory cell infiltration or pathologic abnormalities in their lungs during histology or microcomputed tomography studies. Collectively, intranasal vB_AbaM-IME-AB2 therapy was revealed to be a highly efficacious therapeutic option for treating pneumonia caused by MDR A. baumannii46 (Table 1). The effectiveness of phage-vB_AbaM_Acibel004 against MDR A. baumannii-induced pneumonia in mice was assessed using intratracheal aerosolization 12-h after the bacterial challenge47. The animals treated with phage showed a lower lung bacterial burden as a result of phage replication at the infection site, as well as milder signs of pneumonia47. The severity of pulmonary histopathologic lesions in phage-treated mice was dramatically diminished, with decreased inflammation and cytokine production, and no phage-related adverse effects47 (Table 1). Learning from investigating into the effects of intranasal phage cocktail (PBAB08, PBAB25, PBAB68, PBAB80, and PBAB93) treatment in MDR A. baumannii-associated acute lung infections, this phage cocktail demonstrated remarkable therapeutic efficacy for enhancing animal survival and reducing pulmonary bacterial burden48. Interestingly, phage cocktail administration via intraperitoneal, intranasal, and oral routes was found to be safe, with minimal inflammatory responses, as phage-treated mice showed no significant increase in serum IgE, inflammatory cytokines, or histamine levels48 (Table 1). The findings of different studies indicate the potential benefit of utilizing phage in clinical settings for managing MDR A. baumannii lung infections.
A prior study discovered that an intraperitoneal phage-SS therapy administered both immediately after and 3 h before an intranasal bacterial challenge was significantly efficient in rescuing mice from K. pneumoniae-mediated pneumonia49. Significantly, the phage treatment became ineffective when it was administered 6 h after the infection was induced49 (Table 1). In murine acute pneumonia produced by MDR K. pneumoniae-KP1513, mice were protected from mortality by a single intranasal dosage of phage151350. Furthermore, phage-treated mice showed a reduced burden of K. pneumoniae in their lungs with an improvement in lung damage and lower inflammatory cytokine in lungs as compared to the control group50 (Table 1). In an earlier investigation, intranasal application of phage-VTCCBPA43 at 2-h post-infection with virulent K. pneumoniae resulted in a significant reduction in bacterial load and an improvement in the severity of the pathologic lesions in the murine pneumonic lung51 (Table 1). Significantly, liposome-entrapped phages had been demonstrated to improve the effectiveness of intraperitoneal phage therapy, resulting in complete clearance of K. pneumoniae-induced lobar pneumonia even though the therapy was administered 72 h after and 48 h before the mice developed pneumonia52. Meanwhile, non-liposomal phage administration resulted in a lower bacterial burden when given 24 h after and 6 h before infection, highlighting the superiority of liposomal delivery in IP phage administration for both therapeutic and prophylactic treatment of K. pneumoniae-induced lobar pneumonia52 (Table 1). A phage cocktail containing pKp11 and pKp383 outperformed its monotherapy in terms of animal survival, bacterial load, inflammatory cytokines, and lung damage in early stage of murine pneumonia caused by refractory ST11 and ST383 K. pneumoniae, suggesting the promising potential of phages in addressing difficult-to-treat K. pneumoniae lung infections53 (Table 1). Furthermore, phage-derived endolysin-LysCA and LysG24 exhibited remarkable antibacterial activity in vitro and therapeutic effects in vivo, drastically lowering the bacterial load while leaving no pathologic lesions in the lungs of treated mice with K. pneumoniae-associated pneumonia. These endolysins demonstrated excellent environmental adaptability and safety, suggesting the possibility of endolysin as an alternative therapy for the treatment of K. pneumoniae-associated lung infections54 (Table 1).
When treating pneumonia caused by highly virulent E. coli 536-Lux strain, intranasal phage-536_P1 therapy was proven to be as successful as ceftriaxone therapy in animal survival and lowering bacterial load55. Phage-536_P7 treatment, meanwhile, did not significantly reduce mortality in VAP mice infected with a virulent strain of E. coli-PDP30255. Adapting phage-536_P7_PDP302 against E. coli-PDP302 in vitro improved its therapeutic efficacy against E. coli-associated pneumonia in clinical settings, as evidenced by a significant increase in animals’ survival55 (Table 1). Irrespective of the virulence and antimicrobial resistance profiles of the targeted E. coli strains-536 and LM33, intranasal treatment with specific phages (536_P1 and LM33_P1) resulted in a greater reduction of the bacterial load, a quicker correction of abnormal blood cell counts, and lower inflammatory responses than intraperitoneal antibiotic treatments in mice with acute E. coli-associated pneumonia56 (Table 1). These findings highlight the potential significance of phage therapy in the treatment of E. coli-associated lung infections in clinical settings.
Interestingly, intraperitoneal phage therapy was found to be more efficient than intranasal inhalation for lowering bacterial load and inflammatory cytokines in acute murine B. cepacia lung infection, implying that phage could be more readily accessible to these pulmonary pathogens through systemic administration in this study57 (Table 1). In contrast, aerosol inhalation of B. cenocepacia-specific phages-KS12, KS5, KS14 and DC1 via NOID (nose-only inhalation device) reduced bacterial load more significantly than intraperitoneal delivery in a murine lung infection model, suggesting that aerosol phage delivery is an effective administration route for treating antibiotic-resistant B. cepacia complex-induced respiratory infections58 (Table 1). The discrepancies in treatment responses via different administration routes observed between studies could be explained by differences in the phage species, immunological condition of the mice, or bacterial strains used in those studies.
In prior studies, lytic phages exhibiting potential antibacterial efficacy against MDR A. xylosoxidans were isolated and characterized, including a detailed genomic study59,60,61. These findings highlight the significance of utilizing phage therapy as a potent alternative therapy to address the growing issue of antibiotic resistance in A. xylosoxidans-associated infections59,60,61.
Each of these preclinical studies involved isolating phages from the environment that displayed target-specific lytic activities against clinical pathogens that are highly resistant to conventional antibiotics. Importantly, different phage therapies have been investigated, primarily in preclinical scenarios, for the treatment of animal lung infections caused by a single pathogen. There is limited evidence on the efficacy of phage therapy against polymicrobial respiratory infections. Furthermore, these studies documented that prompt administration of phage therapy immediately after development of infection is necessary for successful therapeutic outcomes in animals. However, phage therapy is not expected to be administered to patients shortly after the initial onset of lung infection in clinical settings, which could frequently lead to the development of chronic pulmonary infections. Further preclinical research is needed to better understand the potential of phage therapy as an alternative treatment for acute and chronic lung infections involving polymicrobial scenarios. Despite promising preclinical results, there are still important issues to be addressed about the toxicity, safety, and effectiveness of phage therapy in clinical settings. Moreover, preclinical findings from mice should be extrapolated with caution to humans due to the significant immunological differences between humans and mice62. Therefore, clinical evaluations, including case studies and clinical trials, are required to validate the efficacy and safety of phage therapy in addressing bacterial lung infections in children.
Preclinical studies of phage therapy against Gram-positive bacteria
When given intranasally, both phage-AB-SA01 and its prototype products were equally effective as vancomycin in reducing lung bacterial burden in acute pneumonia, despite using animals with different immunological conditions, genetic backgrounds, and S. aureus strains with varying methicillin susceptibility profiles63 (Table 1). In VAP with methicillin-resistant S. aureus (MRSA) in rats, intravenous therapy of a phage cocktail (2003, 2002, 3A, and K) was comparably efficacious as teicoplanin in improving animal survival and reducing bacterial burden in lungs with improved histopathological outcomes64. The phage cocktail-teicoplanin combination however did not increase survival in treated rats64 (Table 1). The same phage cocktail also failed to outperform its monotherapy in improving animal survival or lowering bacterial burdens in the lungs or spleen when administered via nebulization in combination with IV daptomycin in a rat model of VAP caused by MRSA, implying the limited effectiveness of this phage cocktail-daptomycin combination for individuals with MRSA pneumonia65 (Table 1). When the therapeutic outcomes of different delivery routes for the same phage cocktail were evaluated in rats with MRSA-induced VAP, the combination of intravenous and nebulized delivery of aerosolized phage cocktail demonstrated significant synergistic potential in rescuing animals from death when compared to giving phage cocktail via an intravenous or nebulized route. However, the synergistic effects failed to occur when linezolid was given in combination with nebulized phage cocktail66 (Table 1). Meanwhile, when the phage isolated from sewage was administered intravenously in murine MDR S. aureus-induced pneumonia, phage monotherapy significantly reduced bacterial load and caused less pulmonary histologic damage than clindamycin monotherapy or phage- clindamycin combination treatment67 (Table 1). In a previous study, intraperitoneal phage therapy was demonstrated to be therapeutically effective in saving mice from death and reducing the severity of infection in lung-derived septicemia triggered by S. aureus, as evidenced by significantly higher rates of animal survival and lower levels of inflammatory cytokines and bacterial burden in phage-treated mice compared to control mice68 (Table 1).
Although the antimycobacterial phages isolated using M. smegmatis have been demonstrated to be effective against Mycobacterium species, limited preclinical study has been undertaken on their lytic activities against M. abscessus69. A recent study discovered that the combination of antimycobacterial phage-Muddy and the conventional antibiotic-rifabutin, was significantly improved M. abscessus-GD01 clearance and enhanced treated animals’ survival in CF transmembrane conductance regulator-depleted zebrafish model, revealing the appealing potential of phage-antibiotic combination in resolving M. abscessus-associated infections70. Importantly, functional innate immunity was necessary for phage-Muddy to be efficacious, as evidenced by the lack of efficacy in macrophage-ablated larvae70.
Clinical case reports of phage therapy in bacterial lung infection of children
An earlier study discovered that high and repeated dosages of phages (3–5 ml) instilled into the pleural cavity or around abscesses, experienced no adverse consequences during treatment of suppurative lung diseases in neonates and infants71. The author therefore hypothesized that applying phages directly into the site of infection by lung puncture would avoid the necessity for additional surgical operations71 (Table 2). A previous case study used three rounds of nebulized phage therapy to treat pulmonary S. aureus and P. aeruginosa infections in a 5-year-old cystic fibrosis child, and the patient’s overall well-being improved as a result of microbial eradication from the phage treatment72 (Table 2). Both P. aeruginosa and S. aureus were found to have chronically colonized in the lungs of a 7-year-old cystic fibrosis girl, and the patient required broad-spectrum antibiotic treatment for several years despite the absence of inhibitory effects on bacterial colonization. In this patient, delivering the Pyophage cocktail and Sb-1 phage by nebulization, significantly reduced both pathogens during therapy and follow-up, resulting in improved clinical conditions, pathogen eradication and less antibiotic needs with no phage-related adverse consequences73 (Table 2). Using a personalized phage cocktail inhalation, a 6-year-old cystic fibrosis patient with end-stage lung diseases positive for MDR P. aeruginosa demonstrated symptomatic improvement and well-tolerance to phage treatment without inducing bronchospasm or compromising liver functions74 (Table 2). When phage therapy was given to treat MDR P. aeruginosa-associated sternal wound abscess in a 13-year-old bilateral lung-transplanted patient, phage treatment led to complete wound healing and microbiological eradication, highlighting the potential benefits of phage therapy for antibiotic-resistant bacterial infections following cardiothoracic surgery75 (Table 2).
A recent cohort study of patients with antibiotic-resistant mycobacterial infections, comprising 13 adult and 7 pediatric patients, revealed that 11 individuals—6 of whom were pediatric patients—showed favorable clinical and microbiological responses76. Administration of different phages via IV or aerosolized delivery was well-tolerated, and no patient experienced any serious phage-related side effects76. Although neutralizing antibodies against the phage were discovered in serum after the initial administration of intravenous phage therapy in eight patients, and may have contributed to treatment failure in four of these cases, they were not consistently associated with unsuccessful outcomes in the other cases76. Notably, none of the bacteria isolate from the 11 patients who received phage monotherapy developed phage resistance76. The effectiveness of phages administered orally and locally via inhalation was studied in 93 patients between 1981 and 1986 who had lung abscess, bronchitis, and pneumonia caused by P. aeruginosa, K. pneumoniae, E. coli, S. pyogenes, and Salmonella. Of these, 83 patients experienced successful therapeutic outcomes, 8 patients showed a temporary improvement, and only 2 had no therapeutic impact77.
Due to persistent lung infection with pandrug-resistant (PDR) A. xylosoxidans, a 12-year-old lung-transplanted patient with cystic fibrosis underwent two rounds of 2 phage cocktails given via nebulization alone (first phage cocktail-APC 1.1) and direct instillation followed by nebulization (second phage cocktail-APC 2.1), resulting in clinical tolerance with improvement of patient’s respiratory functions78. Although the low microbial density of A. xylosoxidans persisted in airways for months, this colonization eventually went negative and did not reoccur more than two years after phage therapy78 (Table 2). Periodic phage cocktail therapy, given orally and by inhalation, effectively healed a 17-year-old cystic fibrosis patient’s persistent lung infection caused by A. xylosoxidans, improving the patient’s general well-being and lung functions while decreasing the need for antibiotics and hospitalization79 (Table 2). According to a prior report, a 10-year-old cystic fibrosis patient with PDR Achromobacter spp. was successfully treated with intravenous phage-cefiderocol-meropenem/vaborbactam triple combination therapy80. This treatment regimen was observed to be safe, well-tolerated and efficacious for clearing infection from the lungs while improving patient’s pulmonary functions80 (Table 2).
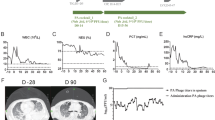
Meanwhile, a 15-year-old cystic fibrosis patient with a disseminated M. abscessus infection following bilateral lung transplants received an engineered three-phage cocktail intravenously with good tolerance, resulting in microbial clearance, clinical improvement with no occurrences of adverse reactions81 (Table 2).
Clinical trials of phage therapy in children
Among the clinical trials conducted on pulmonary infections, most of them assessed the safety and effectiveness of phage therapy in adult patients with cystic fibrosis who had chronic P. aeruginosa-induced pulmonary infections. The safety and efficacy of nebulized YPT-01 phage treatment for reducing the bacterial load in sputum of cystic fibrosis patients with P. aeruginosa was investigated in a phase II clinical trial (NCT04684641)82 (Table 3). The trials-SWARM-Pa (NCT04596319) and Tailwind (NCT0561622) used the same inhaled phage therapy-AP-PA02 in patients who had different clinical presentations83,84. The SWARM-Pa trial, a phase 1b/2a study that assessed the safety, tolerability, and phage recovery profile in CF patients with P. aeruginosa-induced lung infections completed in 202283 (Table 3). Meanwhile, the Tailwind phase 2 trial continues to enroll non-CF subjects with P. aeruginosa-induced bronchiectasis and persistent lung infection to evaluate the safety, phage kinetics, and effectiveness of inhaled AP-PA02 phage therapy84 (Table 3). Patients with CF and persistent P. aeruginosa-induced lung infection continue to recruit for a trial (NCT05010577) investigating the safety and tolerability of nebulized phage, BX004-A85 (Table 3). Concurrently, a multicenter randomized double-blind study (NCT05453578) has begun enrolling CF patients to evaluate the safety and antibacterial activity of intravenous phage therapy-WRAIR-PAM-CF1 against P. aeruginosa86 (Table 3).
A clinical trial involving 6–24-month-old children with acute diarrhea who received oral phage therapy demonstrated the safety of phage therapy by achieving safe phage transit with no alterations to their intestinal microbiome, while there was no discernible improvement in intestinal phage replication or diarrhea outcomes87. A single-arm, non-randomized, open-labeled trial with the Clinical Trial Registry-ACTRN12622000767707 will begin recruiting children with CF who tested positive for P. aeruginosa even after eradication therapy, with the objective of investigating the safety and tolerability of intrabronchial and nebulized phage treatment. In this trial, the selected phage will be administered endo-bronchially under direct vision, followed by twice-daily nebulization for 7 days while receiving standard CF therapy. Following phage therapy, the patients’ progress will be evaluated using clinical assessments which involve spirometry and follow-up sputum cultures at 3, 6, 9, and 12 months88.
Clinical benefits of phage therapy
Phage therapy offers several clinical benefits as an alternative or adjunct to traditional antibiotic treatments. Its specificity to target pathogenic bacteria without affecting the beneficial microbiota, ability to disrupt biofilms, and potential to address antibiotic-resistant infections are among its key advantages. These benefits can significantly improve patient outcomes and provide new avenues for combating bacterial infections.
Ease of isolating target-specific potent phages from environment
With their approximate population of 1032, phages are the most prevalent naturally existing lifeform which influence the microbial equilibrium in human and animal environments89,90. According to fecal metagenomic studies, phages comprise a vital component of the microbiome and are more abundant than eukaryotic viruses, having 1015 versus 108–109/g of feces. When compared to developing novel antibiotics, such ubiquitous characteristics make it easier and less expensive to isolate highly efficient target-specific phages from a variety of environments89,90. This simple and affordable procedure of isolating phages, particularly in low-resource regions of the world, is an appealing reason for investing in phage production to establish a phage bank for future use as an alternative antimicrobial therapy in persistent bacterial lung infections, and it additionally may render importing phages from abroad less necessary89,90. The discovery of therapeutic phages in the specific regions is also beneficial due to the co-evolution of phage and bacteria under the same selective pressures in a shared geographic region, leading to retaining phage-bacteria sensitivity, which is not typically observed in phages obtained from geographically distinct environments89,90.
Reduced selection of antibiotic resistance
The widespread use of broad-spectrum antibiotics puts the pathogens at the site of infection and commensal microbes in the respiratory tract under selection pressure89,91. This eventually leads to the emergence of difficult-to-treat antibiotic-resistant organisms, generating substantial public health concerns and demanding immediate global action due to the extremely limited treatment options in the affected population89,91. In contrast, phage therapy does not pose the risk of triggering antibiotic selection pressure, making it a fascinating possibility for implementation in the global attempt to mitigate antibiotic resistance89,91. By employing phage as an alternative therapy, patients will be exposed to fewer antibiotics, preserving the effectiveness of currently available antibiotics while minimizing the incidence of antibiotic-induced adverse effects and establishment of antibiotic resistance89,91. Moreover, a synergistic impact can be achieved by using specific phages in combination with antibiotics to re-sensitize pathogens to antibiotics to which they were previously resistant89,91.
Reversal of antibiotic resistance by phage-antibiotic synergy (PAS)
When phages were added to pre-existing antibiotic regimens, the combined effect exhibited stronger antibacterial effects than the individual effects, a phenomenon known as phage-antibiotic synergy (PAS)92,93,94. During PAS, target pathogens, even those resistant to the paired antibiotic, were eliminated because of the accelerated multiplication and larger plaque size of combined phages which were facilitated by antibiotic-enhanced phage production in the presence of subinhibitory antibiotic doses92,93,94. Bacterial filamentation caused by β-lactam-related PBPs inhibition, or SOS-response-facilitated survival mechanisms when exposed to sublethal concentrations of β-lactam or DNA-disrupting antibiotics and lethal concentrations of protein or RNA synthesis inhibitors, have also been reported to trigger the PAS response in several studies92,93,94. In addition to increasing bacterial size and surface area, filamentation enables bacterial genomic expression to continue without cell division, allowing phages’ replication for the PAS-associated lysis achievable92,93,94. Furthermore, antibiotic-induced bacterial filamentation improved phage adsorption through enhancing the density of phage receptors on bacterial cell walls, consequently accelerating the phage-induced lysis for PAS92,93,94. However, filamentation mediated by protein or RNA synthesis inhibitors usually results in shorter bacterial cells than those seen with β-lactams or fluoroquinolones, while the specific mechanisms remain unclear92,93,94. The greater probability of producing PAS with β-lactams and fluoroquinolones compared to other antibiotics therefore could be attributed to these distinct variations in bacterial cell surface area modifications92,93,94. Importantly, if the antibiotic dose is insufficient to elicit a subinhibitory response to the bacteria, or if the bacteria are highly resistant to the prescribed drugs, filamentation is unlikely to initiate, and PAS may not occur92,93,94. Therefore, selecting the appropriate antibiotic class and dose for a given phage is crucial for achieving the PAS response92,93,94. Antibiotic-induced bacterial membrane destabilization could shorten the phages’ latent time to invade and destroy bacteria, resulting in the production of PAS and accelerated bacterial lysis92,93,94. Because of a shorter latent interval and faster bacterial lysis, phages can infect nearby bacteria quickly for initiating new lytic cycles, potentially increasing PAS92,93,94. Not only has rapid lysis been linked to PAS, but delayed cell lysis also allows for a longer period for phage synthesis, maturation, and assembly, resulting in greater burst size and phage plaque enlargement, which contributes to PAS92,93,94.
Antibiofilm effects
Biofilms are composed of bacterial microcolonies that aggregate inside a matrix predominantly containing exopolysaccharides, extracellular DNA, secreted proteins, and lipids and have been commonly reported in persistent lung infections95. During phage infection, lytic phages and host bacteria interact by phage adsorption, followed by the release of new progenies through bacteriolysis95. However, phage invasion may be physically impeded by the existence of biofilm matrix95. To overcome this barrier, phages use depolymerase enzymes associated with their tails, which cause the progressive cleavage of polymer bonds, culminating in the degradation of not only the exopolysaccharide matrix of biofilm, but also host bacteria’s polysaccharide capsules95. Consequently, depolymerases facilitate the infiltration and distribution of phages through biofilms while simultaneously eradicating the target bacteria present in biofilm, hence eliminating the pre-existing biofilms and preventing new biofilms’ development95. Reduced bacterial virulence resulting from bacterial capsules degradation by phage-encoded polymerase is an additional therapeutic benefit of phage therapy95. Concurrently, phages generate lysin or virion-associated lysins, which are the hydrolytic enzymes present in phage tails. They serve as receptor recognition proteins, allowing phage genomic material entry into host bacterial cells via bacterial cell wall penetration95. Additionally, they assist to release virions by cleaving the bacterial cell wall from within95. Lysins, can also degrade the extracellular matrix of biofilms and trigger the lysis of bacteria that reside within them95. These phage-related enzymes outperform broad-spectrum antibiotics in clinical settings with their abilities to selectively target pathogens without harming the normal flora, improve their effectiveness through genetic engineering, produce strong synergistic effects when combined with other antibiotics, and rapidly lyse host bacteria with a lower risk of resistance development95. Phages exhibit excellent antibiofilm effects in the presence of these enzymes and phage-induced bacteriolysis, implying that phage or phage-related enzymes, either alone or in conjunction with antibiotics, could ultimately result in greater clinical cure and shorter therapeutic duration while combating biofilm-associated lung infections in children95.
No cross-resistance with antibiotics
Previous studies have revealed that cross-resistance was frequently observed in phages that employed the same target receptors while producing phage-mediated bacteriolysis against their target bacterial infections96,97. However, antibiotic and phage use completely distinct mechanisms for achieving their antibacterial effects, whereas phages infect to replicate within specific bacteria for bacterial lysis and death92,93,98, and antibiotics target cellular mechanisms to disrupt bacterial growth and metabolism99,100. Interestingly, there is no cross-resistance between antibiotics and phages, because the specific mechanisms generating antibiotic resistance such as enzymatic inactivation or modification of antibiotics, reduced permeability of antibiotics, target site protection or modification, target bypass, increased efflux and decreased influx of antibiotics101, cannot be translated into phages. Additionally, while bacteria develop resistance to a specific phage under its selection pressure, this resistance fails to confer cross-resistance to antibiotics as these bacteria employ distinct anti-phage defense mechanisms to generate phage resistance, such as modifying host surface receptors to prevent phage adsorption, restricting phage’s genome entry into the host, minimizing secondary phage infection, activating the restriction-modification and CRISPR-Cas systems, and initiating an abortive infection system, which cannot be translated into antibiotics102,103,104. Among the different phage-resistant mechanisms, the alteration in host surface receptors such as lipopolysaccharide, capsule polysaccharide, pili, outer membrane porins, and efflux pumps are the most significant mechanisms involved in bacterial resistance to phage102,103,104. The receptors alteration can potentially affect bacterial fitness because these surface structures are essential for various cellular functions, bacterial virulence, and antibiotic resistance90,104,105,106. Therefore, the presence of phage resistance frequently triggers several trade-offs in bacterial pathogens, including decreased pathogenicity, delayed bacterial growth, and increased antibiotic susceptibilities, which eventually cause phage-resistant bacteria regaining their susceptibility to antibiotics105,106,107. Phage-induced restoration of antibiotic sensitivity is reported in several studies to positively impact antibiotic resistance in clinical settings, providing an important clinical benefit for repurposing the conventional antibiotics in phage combination therapy to combat drug-resistant respiratory pathogens90,91,92,93,104,105,106,107. However, pleiotropic alterations in bacterial susceptibilities to antibiotics was observed when phage resistance evolved from a variety of mutations in distinct bacterial target structures that antibiotics employed to demonstrate their antibacterial effects108,109,110. It was reported in a prior study whereas resistance to both phages-T6, U115, and albicidin has observed in E. coli through mutations in Tsx, an outer membrane porin that functions as a receptor for these phages, or a porin for an uptake of albicidin, an antibiotic inhibiting DNA gyrase109. A recent study found that phage-U136B employed the antibiotic efflux pump-TolC and lipopolysaccharide (LPS) as its target receptors to induce bacteriolysis in E. coli. Mutations in the tolC or LPS synthesis gene of phage-resistant E. coli in their study resulted in lower resistance to tetracycline and colistin110. Interestingly, a subgroup of their LPS mutants developed a greater resistance to tetracycline while reducing colistin resistance, demonstrating that these LPS mutants displayed a pleiotropic impact on tetracycline susceptibilities110. These findings highlighted not only the development of trade-offs, but also the pleiotropic evolution of trade-ups could be generated by specific phage-resistant mutations, emphasizing the necessity of screening to identify phages capable of pleiotropically selecting against drug-resistance genes in target pathogens.
Self-replication at the site of infection
During the phage infection cycle, the first infection of host bacteria for phage-induced lysis is primary infection, and the subsequent replication in nearby bacteria by virions released from lysed cells is referred to secondary infection90,111. Phage therapy for bacterial lung infections frequently results in a secondary infection of phage replication in the lungs90,111,112. Phage replication additionally allows phages to multiply in deeper lung infection areas that are more difficult to access but have a high bacterial load90,111,112. Because they can only exist while their target pathogenic bacteria are present at the infection site, their replication is however self-limiting90,111,112. When treating persistent bacterial lung infections, this self-replicating ability offers the benefit of auto-dosing, whereas high-dosage of phage administrations will be less needed to achieve the optimal therapeutic outcomes89,90. Auto-dosing additionally contributes the benefit of safety with few to no side effects as its antibacterial impact is confined to its infection site at lower dosage89,90.
Coevolution of phages and bacteria
Because phages and bacteria coexist in diverse environments, bacteria are constantly adapting to survive phage attacks by generating anti-phage defense mechanisms as described previously102,103,104. Meanwhile, phages also adopt a variety of coevolutionary strategies to keep attacking bacteria that resist its infection106,113,114,115. One of these is enhanced phage’s adsorption and entry through the identification of new or altered host surface receptors and modification of receptor-binding regions to compensate for alterations in its bacterial surface receptors106,113,114,115. Other strategies also assist phages to overcome restriction modification-mediated phage resistance by modifying the restriction sites within genomes, methylating genomes, and eliminating the cofactors required for restriction-modification systems106,113,114,115. Phages effectively circumvent CRISPR-Cas-mediated resistance by generating anti-CRISPR proteins and introducing point mutations in the protospacer regions or CRISPR-targeted sequences106,113,114,115. Phages generate antitoxin proteins and mutations in their genomes to terminate abortive infection-mediated phage resistance106,113,114,115. Because of these coevolutionary interactions between phages and bacteria, there is a lower risk of phage and/or antibiotic-resistant bacterial escape, which is a significant therapeutic benefit of utilizing phages over antibiotics106,113,114,115.
Minimal to no toxicity
Phages can be regarded as non-toxic because their main components are single or double-stranded nucleic acids wrapped in protein capsids116,117. Due to their receptor-mediated host specificity, phages pose no threat to mammalian eukaryotic cells as these cells lack the required surface receptors to allow for phage infection116,117. These characteristics provide phage therapy tolerable, resulting in minimal to no toxicity when compared to antibiotics116,117.
Minimal risks of dysbiosis
While broad-spectrum antibiotics produce deleterious impacts on the airway microbiome due to their non-specific antibacterial effects, target-specific phage therapy offers a significant therapeutic benefit with minimal dysbiosis risks118,119. Phage therapy has been reported in human and animal studies to have no impact on the microbiome, preventing the eradication of potentially beneficial microbes, secondary pathogens proliferation, and the emergence of resistant microorganisms118,119.
Clinical challenges of phage therapy
Bacteriophages, used to treat persistent bacterial lung infections, present several clinical challenges despite their potential as an alternative to antibiotics. These challenges span various aspects, including therapeutic efficacy, safety, and logistical considerations. The complexity of phage-bacteria interactions, the body’s immune response to phages, and the stability of phage preparations are key factors that influence the clinical success of this therapeutic approach. Understanding and addressing these challenges is crucial for the effective implementation of phage therapy in medical practice.
Therapeutic failure from strains variation and phage resistance
The most notable obstacle related to phage’s host specificity is the failure of phage-induced bacteriolysis among bacterial isolates belonging to same species120,121. Because there is a significant diversity across bacterial strains within a species, this strain variation contributes to phage non-susceptibility120,121. The use of phage cocktails, which comprise various clinically efficacious phages belonging to different families with diverse mechanisms of action, and the administration of phages with broad-spectrum host ranges against diverse set of clinical strains belonging to the major lineages of target pathogens species, are promising approaches to address this issue120,121.
Another unavoidable and frequent issue encountered during phage therapy is the emergence of phage-insensitive mutations, which ultimately leads to inadequate bacterial clearance and phage therapy failure122,123. Sequential or simultaneous administration of a phage cocktail comprising different phages with distinct modes of action, provided encouraging results in the fight against the development of bacterial phage resistance122,123. Meanwhile, the use of genetically engineered phages can bring the advantages of increasing the therapeutic efficacy and safety of phages and limiting the development of phage resistance in the clinical settings122,123.
Antagonistic phage–phage interactions
Antagonistic phage-phage interaction occurs when phages hinder or oppose the growth, replication, or functionality of other phages while infecting the same host bacterium124,125. It occurs through different mechanisms, including superinfection and competitive exclusion by blocking host surface receptors and decreasing cellular resources as a result of phage-phage competition, phage-mediated defense triggered by restriction modification systems that identify and degrade competing phage’s DNA, and the production of anti-phage proteins like bacteriocins or phage-encoded abortive infection systems that target and limit the growth or replication of other phages124,125. The effectiveness of phage therapy may therefore be adversely affected by antagonistic phage-phage interactions124,125. Undoubtedly, administering different phages sequentially or individually could lower the possibility of undesirable phage-phage interactions when compared to giving combined phage cocktails124,125. Because minimal research has been conducted on these detrimental consequences of antagonistic interaction during phage therapy, it can be challenging to predict how these phenomenon would impact different treatment modalities124,125. It is therefore imperative to continue researching the mechanisms underlying antagonistic phage-phage interactions to develop alternative strategies to increase the efficacy of phage-based therapies in combating persistent lung infections124,125.
Safety concerns
Currently, the potential clinical risk linked with phage therapy is that phages, particularly temperate phages, could trigger safety challenges by dissemination of new genetic characteristics among bacterial populations116,126. During lysogenic cycles, temperate phages integrate into host bacterial genomes as stable prophages and do not induce host bacterial lysis116,126. However, they can facilitate lysogenic conversion of host bacteria, resulting in the emergence of pathogenic strains in their hosts via phage-mediated transduction of new genetic characteristics, such as genes involved in bacterial virulence, toxin production, and antibiotic resistance116,126. Risks assessment therefore needs to involve an evaluation of the complete phage genomes, along with transduction capacity and host range studies for a particular phage to address safety concerns related to phage therapy116,126.
Immunity to phages
Due to their antigenic nature, prolonged exposure to phages and their byproducts can trigger the mammalian immune system to generate phage-specific T and B cells immune responses in vitro and in vivo127,128. Previous studies discovered that phage survival was significantly longer in immunocompromised animals and humans as compared to immunocompetent ones, highlighting the critical role of adaptive immunity in phage clearance127,129,130. The phage titer remained unchanged in B-cell-deficient mice, suggesting that anti-phage antibody production was significantly responsible for the phage clearance131. Although IgM accounts for the majority of anti-phage neutralizing antibodies, IgG and IgA are also produced132. It has been revealed that both environmental and genetically engineered phages can trigger the development of anti-phage antibodies132. Although it has not been further investigated in controlled trials, administering phages at the site of infection was reported to generate stronger antibody responses than oral administration in patients133. Individuals who have been given structurally comparable phages could possess phage-specific antibodies, which could mitigate the effectiveness of subsequent phage treatments127,132. Interestingly, enrolled human subjects were discovered to have preexisting anti-phage antibodies with significant reduction of phage’s activity, even though none of them had previously received phage therapy134,135. Nevertheless, the impact of anti-phage antibodies on phage therapy outcomes remains uncorrelated since there was no discernible relationship between the patient’s treatment responses and the development of phage-specific humoral immunity133,136. The absence of controlled trials using standardized phage products, administration protocols, and treatment durations makes it extremely difficult to effectively evaluate how patients’ development of anti-phage antibodies influences therapeutic outcomes127,132. It will also be necessary to explore whether the existence of neutralizing antibodies to a specific phage limits the future therapeutic application of that particular phage or other phage families.
Stability and pharmacokinetic issues
Under diverse environmental conditions, phages containing nucleic acids encased in proteinaceous capsids have the same potential to denature as proteins137,138,139. Phage manufacturing, transportation, and storage are therefore challenging due to its limited stability137,138,139. Phage instability can be triggered by fluctuations in temperature, pH, and mechanical stresses during manufacturing, resulting in a reduction in phage therapeutic effectiveness137,138,139. Therefore, it is preferable to implement minimal steps to ensure the stability and potency of phages throughout manufacturing137,138. Moreover, the stability of phages in different formulations has been documented to be phage-specific, and variations in stabilities between phages may jeopardize the process of developing phage cocktails in diverse formulations137,138,139.
Furthermore, phages possess different pharmacokinetic profiles from conventional antibiotics whereas their ability for self-replication is associated with eradication of pathogens137,138,139. The effectiveness of phage therapy is significantly influenced by the presence of target pathogens, the relative quantity of phages to bacteria, and the ideal timing for phage delivery137,138,139. The pharmacokinetics of phages, including their distribution, metabolism, and excretion in vivo, need to be investigated for optimizing treatment regimens137,138,139. However, the limited knowledge about phage pharmacokinetics renders manufacturing and developing a standardized protocol for phage therapy challenging.
Future research directions and conclusion
The use of phage therapy has demonstrated remarkable therapeutic potential in preclinical and clinical settings when convectional antibiotics failed to control persistent bacterial lung infections in children. Moving forward, research endeavors in phage therapy for bacterial lung infections in children should focus on several key areas to advance its clinical applicability and effectiveness. First and foremost, standardizing phage formulations is crucial to ensure consistent therapeutic outcomes and streamline the regulatory approval process. This involves refining phage production techniques, enhancing the stability of phage formulations for extended storage, and determining the optimal phage-to-bacteria ratios for maximizing its treatment efficacy. Understanding the pharmacokinetics of phages is pivotal for optimizing treatment regimens. Investigating the distribution, metabolism, and excretion of phages in vivo will provide insights into their bioavailability, persistence, and overall therapeutic profile. Additionally, it’s imperative to delve deeper into the mechanisms governing antagonistic interactions between phages. Uncovering these interactions will pave the way for developing strategies to mitigate their adverse effects, thereby boosting phage treatment’s efficacy.
Long-term studies are also essential to ascertain the safety and efficacy of phage therapy across diverse patient populations and geographical locations. These studies should focus on monitoring potential adverse effects, tracking the emergence of phage and antibiotic resistance, and assessing long-term patients’ outcomes. Exploring the synergistic potential of combining phage therapy with other therapeutic modalities, such as antibiotics or immunomodulatory agents, could offer a multifaceted approach to enhancing treatment outcomes. Lastly, the establishment of comprehensive regulatory frameworks and ethical guidelines is paramount to ensure patient’s safety and foster broader acceptance of phage therapy. This includes the development of standardized protocols, rigorous safety assessments, and clear guidelines for informed consent and patients’ monitoring.
In conclusion, advancing our understanding of phage therapy through systematic research and addressing these pivotal research directions will be instrumental in realizing its full therapeutic potential for combating persistent bacterial lung infections in children.
References
Collaro, A. J. et al. The effect of early childhood respiratory infections and pneumonia on lifelong lung function: a systematic review. The Lancet Child & Adolescent Health (2023).
Couriel, J. Assessment of the child with recurrent chest infections. Br. Med. Bull. 61, 115–132 (2002).
Pragman, A. A., Berger, J. P. & Williams, B. J. Understanding persistent bacterial lung infections: clinical implications informed by the biology of the microbiota and biofilms. Clin. Pulm. Med. 23, 57–66 (2016).
Blanchard, A. C. & Waters, V. J. Opportunistic pathogens in cystic fibrosis: epidemiology and pathogenesis of lung infection. J. Pediatr. Infect. Dis. Soc. 11, S3–S12 (2022).
Delfino, E. et al. First report of chronic pulmonary infection by KPC-3-producing and colistin-resistant Klebsiella pneumoniae sequence type 258 (ST258) in an adult patient with cystic fibrosis. J. Clin. Microbiol. 53, 1442–1444 (2015).
Jenkinson, A. C. et al. Acinetobacter baumannii Infection in a child with cystic fibrosis. J. Paediatr. Child Health 57, 1148–1149 (2021).
Debnath, S. K., Debnath, M. & Srivastava, R. Opportunistic etiological agents causing lung infections: emerging need to transform lung-targeted delivery. Heliyon 8, e12620 (2022).
Degiacomi, G. et al. Mycobacterium abscessus, an emerging and worrisome pathogen among cystic fibrosis patients. Int. J. Mol. Sci. 20, 5868 (2019).
Shein, A. M. S. et al. Current and novel therapies for management of Acinetobacter baumannii-associated pneumonia. Crit. Rev. Microbiol. 1–22 (2024).
Chirabhundhu, N. et al. Occurrence and mechanisms of tigecycline resistance in carbapenem- and colistin-resistant Klebsiella pneumoniae in Thailand. Sci. Rep. 14, 5215 (2024).
Akkerman-Nijland, A. M. et al. The pharmacokinetics of antibiotics in cystic fibrosis. Expert Opin. Drug Metab. Toxicol. 17, 53–68 (2021).
AlKhater, S. A. Approach to the child with recurrent infections. J. Fam. Community Med. 16, 77–82 (2009).
La Rosa, R., Johansen, H. K. & Molin, S. Persistent bacterial infections, antibiotic treatment failure, and microbial adaptive evolution. Antibiotics 11, 419 (2022).
Reardon, S. Phage therapy gets revitalized: the rise of antibiotic resistance rekindles interest in a century-old virus treatment. Nature 510, 15–17 (2014).
Ioannou, P., Baliou, S. & Samonis, G. Bacteriophages in infectious diseases and beyond—a narrative review. Antibiotics 12, 1012 (2023).
Ranveer, S. A. et al. Positive and negative aspects of bacteriophages and their immense role in the food chain. npj Sci. Food 8, 1 (2024).
Cui, L., Veeranarayanan, S., Thitiananpakorn, K. & Wannigama, D. L. Bacteriophage bioengineering: a transformative approach for targeted drug discovery and beyond. Pathogens 12, 1179 (2023).
Nguyen, H. M. et al. RNA and single-stranded DNA phages: unveiling the promise from the underexplored world of viruses. Int J. Mol. Sci. 24, 17029 (2023).
Shein, A. M. S. et al. Novel intranasal phage-CaEDTA-ceftazidime/avibactam triple combination therapy demonstrates remarkable efficacy in treating Pseudomonas aeruginosa lung infection. Biomed. Pharmacother. 168, 115793 (2023).
Sutnu, N. et al. Bacteriophages isolated from mouse feces attenuates pneumonia mice caused by Pseudomonas aeruginosa. PLOS ONE 19, e0307079 (2024).
Debarbieux, L. et al. Bacteriophages can treat and prevent Pseudomonas aeruginosa lung infections. J. Infect. Dis. 201, 1096–1104 (2010).
Morello, E. et al. Pulmonary bacteriophage therapy on Pseudomonas aeruginosa cystic fibrosis strains: first steps towards treatment and prevention. PloS One 6, e16963 (2011).
Henry, M., Lavigne, R. & Debarbieux, L. Predicting in vivo efficacy of therapeutic bacteriophages used to treat pulmonary infections. Antimicrob. Agents Chemother. 57, 5961–5968 (2013).
Yang, M. et al. Therapeutic effect of the YH6 phage in a murine hemorrhagic pneumonia model. Res. Microbiol. 166, 633–643 (2015).
Abd El-Aziz, A. M., Elgaml, A. & Ali, Y. M. Bacteriophage therapy increases complement-mediated lysis of bacteria and enhances bacterial clearance after acute lung infection with multidrug-resistant Pseudomonas aeruginosa. J. Infect. Dis. 219, 1439–1447 (2019).
Jeon, J. & Yong, D. Two novel bacteriophages improve survival in Galleria mellonella infection and mouse acute pneumonia models infected with extensively drug-resistant Pseudomonas aeruginosa. Appl. Environ. Microbiol. 85, e02900–e02918 (2019).
Chow, M. Y. et al. Pharmacokinetics and time-kill study of inhaled antipseudomonal bacteriophage therapy in mice. Antimicrob. Agents Chemother. 65, https://doi.org/10.1128/aac.01470-01420 (2020).
Zhang, Y. et al. Evaluation of phage therapy for pulmonary infection of mouse by liquid aerosol-exposure Pseudomonas aeruginosa. Infect. Drug Resist. 14, 4457–4469 (2021).
Chang, R. Y. K. et al. The effects of different doses of inhaled bacteriophage therapy for Pseudomonas aeruginosa pulmonary infections in mice. Clin. Microbiol. Infect. 28, 983–989 (2022).
Chang, R. Y. K. et al. Proof-of-principle study in a murine lung infection model of antipseudomonal activity of phage PEV20 in a dry-powder formulation. Antimicrob. Agents Chemother. 62, https://doi.org/10.1128/aac.01714-01717 (2018).
Raz, A., Serrano, A., Hernandez, A., Euler, C. W. & Fischetti, V. A. Isolation of phage lysins that effectively kill Pseudomonas aeruginosa in mouse models of lung and skin infection. Antimicrob. Agents Chemother. 63, https://doi.org/10.1128/aac.00024-00019 (2019).
Chen, F. et al. Novel lytic phages protect cells and mice against Pseudomonas aeruginosa infection. J. Virol. 95, https://doi.org/10.1128/jvi.01832-01820 (2021).
Waters, E. M. et al. Phage therapy is highly effective against chronic lung infections with Pseudomonas aeruginosa. Thorax 72, 666–667 (2017).
Alemayehu, D. et al. Bacteriophages ϕMR299-2 and ϕNH-4 can eliminate Pseudomonas aeruginosa in the murine lung and on cystic fibrosis lung airway cells. MBio 3, https://doi.org/10.1128/mbio.00029-00012 (2012).
Pabary, R. et al. Antipseudomonal bacteriophage reduces infective burden and inflammatory response in murine lung. Antimicrob. Agents Chemother. 60, 744–751 (2016).
Forti, F. et al. Design of a broad-range bacteriophage cocktail that reduces Pseudomonas aeruginosa biofilms and treats acute infections in two animal models. Antimicrob. Agents Chemother. 62, https://doi.org/10.1128/aac.02573-02517 (2018).
Duplessis, C., Warawa, J. M., Lawrenz, M. B., Henry, M. & Biswas, B. Successful intratracheal treatment of phage and antibiotic combination therapy of a multi-drug resistant Pseudomonas aeruginosa murine model. Antibiotics 10, 946 (2021).
Guillon, A. et al. Inhaled bacteriophage therapy in a porcine model of pneumonia caused by Pseudomonas aeruginosa during mechanical ventilation. Br. J. Pharmacol. 178, 3829–3842 (2021).
Roach, D. R. et al. Synergy between the host immune system and bacteriophage is essential for successful phage therapy against an acute respiratory pathogen. Cell Host Microbe 22, 38–47. e34 (2017).
Lin, Y. et al. Synergy of nebulized phage PEV20 and ciprofloxacin combination against Pseudomonas aeruginosa. Int. J. Pharm. 551, 158–165 (2018).
Lin, Y. et al. Synergistic activity of phage PEV20-ciprofloxacin combination powder formulation—a proof-of-principle study in a P. aeruginosa lung infection model. Eur. J. Pharm. Biopharm. 158, 166–171 (2021).
Menon, N. D. et al. Increased innate immune susceptibility in hyperpigmented bacteriophage-resistant mutants of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 66, e00239–00222 (2022).
Jeon, J. et al. In vivo application of bacteriophage as a potential therapeutic agent to control OXA-66-like carbapenemase-producing Acinetobacter baumannii strains belonging to sequence type 357. Appl. Environ. Microbiol. 82, 4200–4208 (2016).
Jeon, J., Park, J.-H. & Yong, D. Efficacy of bacteriophage treatment against carbapenem-resistant Acinetobacter baumannii in Galleria mellonella larvae and a mouse model of acute pneumonia. BMC Microbiol. 19, 1–14 (2019).
Hua, Y. et al. Phage therapy as a promising new treatment for lung infection caused by carbapenem-resistant Acinetobacter baumannii in mice. Front. Microbiol. 8, 318476 (2018).
Wang, Y. et al. Intranasal treatment with bacteriophage rescues mice from Acinetobacter baumannii-mediated pneumonia. Future Microbiol. 11, 631–641 (2016).
Wienhold, S.-M. et al. Preclinical assessment of bacteriophage therapy against experimental Acinetobacter baumannii lung infection. Viruses 14, 33 (2021).
Cha, K., Oh, H. K. & Myung, H. Characterization of two novel bacteriophages infecting multidrug-resistant (MDR) Acinetobacter baumannii and evaluation of their therapeutic efficacy in vivo. Front Microbiol 9, 324473 (2018).
Chhibber, S., Kaur, S. & Kumari, S. Therapeutic potential of bacteriophage in treating Klebsiella pneumoniae B5055-mediated lobar pneumonia in mice. J. Med. Microbiol. 57, 1508–1513 (2008).
Cao, F. et al. Evaluation of the efficacy of a bacteriophage in the treatment of pneumonia induced by multidrug resistance Klebsiella pneumoniae in mice. BioMed. Res. Int. 2015, 752930(2015).
Anand, T. et al. Phage therapy for treatment of virulent Klebsiella pneumoniae infection in a mouse model. J. Glob. Antimicrob. Resist. 21, 34–41 (2020).
Singla, S., Harjai, K., Katare, O. P. & Chhibber, S. Bacteriophage-loaded nanostructured lipid carrier: improved pharmacokinetics mediates effective resolution of Klebsiella pneumoniae–induced lobar pneumonia. J. Infect. Dis. 212, 325–334 (2015).
Gan, L. et al. Bacteriophage effectively rescues pneumonia caused by prevalent multidrug-resistant Klebsiella pneumoniae in the early stage. Microbiol. Spectr. 10, e02358–02322 (2022).
Lu, B. et al. Isolation of Klebsiella pneumoniae phage vB_KpnS_MK54 and pathological assessment of endolysin in the treatment of pneumonia mice model. Front. Microbiol. 13, 854908 (2022).
Dufour, N., Debarbieux, L., Fromentin, M. & Ricard, J.-D. Treatment of highly virulent extraintestinal pathogenic Escherichia coli pneumonia with bacteriophages. Crit. Care Med. 43, e190–e198 (2015).
Dufour, N., Delattre, R., Chevallereau, A., Ricard, J.-D. & Debarbieux, L. Phage therapy of pneumonia is not associated with an overstimulation of the inflammatory response compared to antibiotic treatment in mice. Antimicrob. Agents Chemother. 63, https://doi.org/10.1128/aac.00379-00319 (2019).
Carmody, L. A. et al. Efficacy of bacteriophage therapy in a model of Burkholderia cenocepacia pulmonary infection. J. Infect. Dis. 201, 264–271 (2010).
Semler, D. D., Goudie, A. D., Finlay, W. H. & Dennis, J. J. Aerosol phage therapy efficacy in Burkholderia cepacia complex respiratory infections. Antimicrob. Agents Chemother. 58, 4005–4013 (2014).
Ma, Y. et al. Isolation and molecular characterisation of Achromobacter phage phiAxp-3, an N4-like bacteriophage. Sci. Rep. 6, 24776 (2016).
Li, E. et al. Characterization of a novel Achromobacter xylosoxidans specific siphoviruse: phiAxp-1. Sci. Rep. 6, 21943 (2016).
Wittmann, J., Dreiseikelmann, B., Rohde, C., Rohde, M. & Sikorski, J. Isolation and characterization of numerous novel phages targeting diverse strains of the ubiquitous and opportunistic pathogen Achromobacter xylosoxidans. PLoS One 9, e86935 (2014).
Mestas, J. & Hughes, C. C. Of mice and not men: differences between mouse and human immunology. J. Immunol. 172, 2731–2738 (2004).
Lehman, S. M. et al. Design and preclinical development of a phage product for the treatment of antibiotic-resistant Staphylococcus aureus infections. Viruses 11, 88 (2019).
Prazak, J. et al. Bacteriophages improve outcomes in experimental Staphylococcus aureus ventilator-associated pneumonia. Am. J. Respir. Crit. Care Med. 200, 1126–1133 (2019).
Valente, L. G. et al. Searching for synergy: combining systemic daptomycin treatment with localised phage therapy for the treatment of experimental pneumonia due to MRSA. BMC Res. Notes 14, 1–4 (2021).
Prazak, J. et al. Benefits of aerosolized phages for the treatment of pneumonia due to methicillin-resistant Staphylococcus aureus: an experimental study in rats. J. Infect. Dis. 225, 1452–1459 (2022).
Ochieng’Oduor, J. M., Onkoba, N., Nyachieo, A. & Maloba, F. Experimental phage therapy against haematogenous multi-drug resistant Staphylococcus aureus pneumonia in mice. Afr. J. Lab. Med. 5, 1–7 (2016).
Takemura-Uchiyama, I. et al. Experimental phage therapy against lethal lung-derived septicemia caused by Staphylococcus aureus in mice. Microbes Infect. 16, 512–517 (2014).
Hashemi Shahraki, A. & Mirsaeidi, M. Phage therapy for Mycobacterium abscessus and strategies to improve outcomes. Microorganisms 9, 596 (2021).
Johansen, M. D. et al. Mycobacteriophage–antibiotic therapy promotes enhanced clearance of drug-resistant Mycobacterium abscessus. Dis. Models Mech. 14, dmm049159 (2021).
Shishenko, S. F. To the issue of application of D’Herelle’s bacteriophage in children surgery. Sel. Art. Azerbaijan Inst. Micro Epidem. 6, 107–114 (1938).
Kutateladze, A. & Adamia, R. Phage therapy experience at the Eliava Institute. Méd. Maladies Infect. 38, 426–430 (2008).
Kvachadze, L. et al. Evaluation of lytic activity of staphylococcal bacteriophage Sb‐1 against freshly isolated clinical pathogens. Microb. Biotechnol. 4, 643–650 (2011).
Hahn, A. et al. Bacteriophage therapy for pan-drug-resistant Pseudomonas aeruginosa in two persons with cystic fibrosis. J. Investig. Med. High Impact Case Rep. 11, 23247096231188243 (2023).
Rubalskii, E. et al. Bacteriophage therapy for critical infections related to cardiothoracic surgery. Antibiotics 9, 232 (2020).
Dedrick, R. M. et al. Phage therapy of Mycobacterium infections: compassionate use of phages in 20 patients with drug-resistant mycobacterial disease. Clin. Infect. Dis. 76, 103–112 (2023).
Slopek, S., Weber-Dabrowska, B., Dabrowski, M. & Kucharewicz-Krukowska, A. Results of bacteriophage treatment of suppurative bacterial infections in the years 1981-1986. Arch. Immunol. Ther. Exp. 35, 569–583 (1987).
Lebeaux, D. et al. A case of phage therapy against pandrug-resistant Achromobacter xylosoxidans in a 12-year-old lung-transplanted cystic fibrosis patient. Viruses 13, 60 (2021).
Hoyle, N. et al. Phage therapy against Achromobacter xylosoxidans lung infection in a patient with cystic fibrosis: a case report. Res. Microbiol. 169, 540–542 (2018).
Gainey, A. B. et al. Combining bacteriophages with cefiderocol and meropenem/vaborbactam to treat a pan‐drug resistant Achromobacter species infection in a pediatric cystic fibrosis patient. Pediatr. Pulmonol. 55, 2990–2994 (2020).
Dedrick, R. M. et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 25, 730–733 (2019).
Koff J., Yale University CYstic Fibrosis bacterioPHage Study at Yale (CYPHY) [(accessed 16 March 2024)]; Available online: https://clinicaltrials.gov/ct2/show/NCT04684641.
Cystic Fibrosis Foundation Ph 1/2 study evaluating safety and tolerability of inhaled AP-PA02 in subjects with chronic Pseudomonas aeruginosa lung infections and cystic fibrosis (SWARM-Pa) [(accessed 18 March 2024)]; Available online: https://clinicaltrials.gov/ct2/show/NCT04596319.
Armata Pharmaceuticals, Inc. Study to evaluate the safety, phage kinetics, and efficacy of inhaled AP-PA02 in subjects with non-cystic fibrosis bronchiectasis and chronic pulmonary pseudomonas aeruginosa infection (Tailwind) [(accessed 12 March 2024)]; Available online: https://clinicaltrials.gov/ct2/show/NCT05616221.
BiomX, Inc. Nebulized bacteriophage therapy in cystic fibrosis patients with chronic pseudomonas aeruginosa pulmonary infection. [(accessed 15 March 2024)]; Available online: https://clinicaltrials.gov/ct2/show/NCT05010577.
National Institute of Allergy and Infectious Diseases (NIAID) A phase 1b/2 trial of the safety and microbiological activity of bacteriophage therapy in cystic fibrosis subjects colonized with Pseudomonas aeruginosa [(accessed 15 March 2024)]; Available online: https://clinicaltrials.gov/ct2/show/NCT05453578.
Sarker, S. A. et al. Oral phage therapy of acute bacterial diarrhea with two coliphage preparations: a randomized trial in children from Bangladesh. EBioMedicine 4, 124–137 (2016).
Singh, J. et al. Single-arm, open-labelled, safety and tolerability of intrabronchial and nebulised bacteriophage treatment in children with cystic fibrosis and Pseudomonas aeruginosa. BMJ Open Respir. Res. 10, e001360 (2023).
Rahimi-Midani, A., Lee, S.-W. & Choi, T.-J. Potential solutions using bacteriophages against antimicrobial resistant bacteria. Antibiotics 10, 1496 (2021).
Loc-Carrillo, C. & Abedon, S. T. Pros and cons of phage therapy. Bacteriophage 1, 111–114 (2011).
Azam, A. H. et al. Selective bacteriophages reduce the emergence of resistant bacteria in bacteriophage-antibiotic combination therapy. Microbiol. Spectrum, e00427–00423 (2024).
Liu, C., Hong, Q., Chang, R. Y. K., Kwok, P. C. L. & Chan, H.-K. Phage–antibiotic therapy as a promising strategy to combat multidrug-resistant infections and to enhance antimicrobial efficiency. Antibiotics 11, 570 (2022).
Gu Liu, C. et al. Phage-antibiotic synergy is driven by a unique combination of antibacterial mechanism of action and stoichiometry. MBio 11, https://doi.org/10.1128/mbio.01462-01420 (2020).
Gurney, J. et al. Phage steering of antibiotic-resistance evolution in the bacterial pathogen, Pseudomonas aeruginosa. Evol. Med. Public Health 2020, 148–157 (2020).
Chang, C., Yu, X., Guo, W., Guo, C. & Zhu, Y. Bacteriophage-mediated control of biofilm: a promising new dawn for the future. Front. Microbiol. 13, 825828 (2022).
Wright, R. C., Friman, V.-P., Smith, M. C. & Brockhurst, M. A. Cross-resistance is modular in bacteria–phage interactions. PLoS Biol. 16, e2006057 (2018).
Adler, B. A. et al. The genetic basis of phage susceptibility, cross-resistance and host-range in Salmonella. Microbiology 167, 001126 (2021).
Łusiak-Szelachowska, M. et al. Bacteriophages and antibiotic interactions in clinical practice: what we have learned so far. J. Biomed. Sci. 29, 23 (2022).
Halawa, E. M. et al. Antibiotic action and resistance: updated review of mechanisms, spread, influencing factors, and alternative approaches for combating resistance. Front. Pharmacol. 14, 1305294 (2024).
Stokes, J. M., Lopatkin, A. J., Lobritz, M. A. & Collins, J. J. Bacterial metabolism and antibiotic efficacy. Cell Metab. 30, 251–259 (2019).
Darby, E. M. et al. Molecular mechanisms of antibiotic resistance revisited. Nat. Rev. Microbiol. 21, 280–295 (2023).
Labrie, S. J., Samson, J. E. & Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 8, 317–327 (2010).
Egido, J. E., Costa, A. R., Aparicio-Maldonado, C., Haas, P.-J. & Brouns, S. J. Mechanisms and clinical importance of bacteriophage resistance. FEMS Microbiol. Rev. 46, fuab048 (2022).
Koskella, B. & Brockhurst, M. A. Bacteria–phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol. Rev. 38, 916–931 (2014).
Mangalea, M. R. & Duerkop, B. A. Fitness trade-offs resulting from bacteriophage resistance potentiate synergistic antibacterial strategies. Infect. Immunity 88, https://doi.org/10.1128/iai.00926-00919 (2020).
Hasan, M. & Ahn, J. Evolutionary dynamics between phages and bacteria as a possible approach for designing effective phage therapies against antibiotic-resistant bacteria. Antibiotics 11, 915 (2022).
McGee, L. W., Barhoush, Y., Shima, R. & Hennessy, M. Phage‐resistant mutations impact bacteria susceptibility to future phage infections and antibiotic response. Ecol. Evol. 13, e9712 (2023).
Burmeister, A. R. & Turner, P. E. Trading-off and trading-up in the world of bacteria–phage evolution. Curr. Biol. 30, R1120–R1124 (2020).
Kortright, K. E., Doss-Gollin, S., Chan, B. K. & Turner, P. E. Evolution of bacterial cross-resistance to lytic phages and albicidin antibiotic. Front. Microbiol. 12, 658374 (2021).
Burmeister, A. R. et al. Pleiotropy complicates a trade-off between phage resistance and antibiotic resistance. Proc. Natl. Acad. Sci. 117, 11207–11216 (2020).
Payne, R. J. & Jansen, V. A. Pharmacokinetic principles of bacteriophage therapy. Clin. Pharmacokinet. 42, 315–325 (2003).
Abedon, S. T. Phage therapy of pulmonary infections. Bacteriophage 5, e1020260 (2015).
Gao, Z. & Feng, Y. Bacteriophage strategies for overcoming host antiviral immunity. Front. Microbiol. 14, 1211793 (2023).
Borin, J. M., Avrani, S., Barrick, J. E., Petrie, K. L. & Meyer, J. R. Coevolutionary phage training leads to greater bacterial suppression and delays the evolution of phage resistance. Proc. Natl Acad. Sci. 118, e2104592118 (2021).
Wang, M. et al. Coevolutionary phage training and Joint application delays the emergence of phage resistance in Pseudomonas aeruginosa. Virus Evol. 9, vead067 (2023).
Liu, D. et al. The safety and toxicity of phage therapy: a review of animal and clinical studies. Viruses 13, 1268 (2021).
Chung, K. M., Nang, S. C. & Tang, S. S. The safety of bacteriophages in treatment of diseases caused by multidrug-resistant bacteria. Pharmaceuticals 16, 1347 (2023).
Emencheta, S. C. et al. The role of bacteriophages in the gut microbiota: implications for human health. Pharmaceutics 15, 2416 (2023).
Kirk, D. et al. Bacteriophages, gut bacteria, and microbial pathways interplay in cardiometabolic health. Cell Rep. 43 (2024).
Bleriot, I. et al. Improving phage therapy by evasion of phage resistance mechanisms. JAC-Antimicrob. Resist. 6, dlae017 (2024).
Ly-Chatain, M. H. The factors affecting effectiveness of treatment in phages therapy. Front. Microbiol. 5, 51 (2014).
Yoo, S. et al. Designing phage cocktails to combat the emergence of bacteriophage-resistant mutants in multidrug-resistant Klebsiella pneumoniae. Microbiol. Spectr. 12, e01258–01223 (2024).
Chan, B. K., Abedon, S. T. & Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 8, 769–783 (2013).
Molina, F., Menor-Flores, M., Fernández, L., Vega-Rodríguez, M. A. & García, P. Systematic analysis of putative phage-phage interactions on minimum-sized phage cocktails. Sci. Rep. 12, 2458 (2022).
Niu, Y. D. et al. Efficacy of individual bacteriophages does not predict efficacy of bacteriophage cocktails for control of Escherichia coli O157. Front. Microbiol. 12, 616712 (2021).
Nale, J. Y. & Clokie, M. R. Preclinical data and safety assessment of phage therapy in humans. Curr. Opin. Biotechnol. 68, 310–317 (2021).
Popescu, M., Van Belleghem, J. D., Khosravi, A. & Bollyky, P. L. Bacteriophages and the immune system. Annu. Rev. Virol. 8, 415–435 (2021).
Górski, A., Międzybrodzki, R., Jończyk-Matysiak, E., Żaczek, M. & Borysowski, J. Phage-Specific Diverse Effects of Bacterial Viruses on the Immune System. Future Microbiol. 14, 1171–1174 (2019).
Ochs, H. D., Davis, S. D. & Wedgwood, R. J. Immunologic responses to bacteriophage ϕX 174 in immunodeficiency diseases. J. Clin. Investig. 50, 2559–2568 (1971).
Sokoloff, A. V., Zhang, G., Sebestyén, M. G., Wolff, J. A. & Bock, I. The interactions of peptides with the innate immune system studied with use of T7 phage peptide display. Mol. Ther. 2, 131–139 (2000).
Srivastava, A. S., Kaido, T. & Carrier, E. Immunological factors that affect the in vivo fate of T7 phage in the mouse. J. Virol. Methods 115, 99–104 (2004).
Gembara, K. & Dąbrowska, K. Phage-specific antibodies. Curr. Opin. Biotechnol. 68, 186–192 (2021).
Łusiak-Szelachowska, M. et al. Phage neutralization by sera of patients receiving phage therapy. Viral Immunol. 27, 295–304 (2014).
Dąbrowska, K. et al. Immunogenicity studies of proteins forming the T4 phage head surface. J. Virol. 88, 12551–12557 (2014).
Rubinstein, A. et al. Progressive specific immune attrition after primary, secondary and tertiary immunizations with bacteriophage φX174 in asymptomatic HIV-1 infected patients. AIDS 14, F55–F62 (2000).
Łusiak-Szelachowska, M. et al. Antiphage activity of sera during phage therapy in relation to its outcome. Future Microbiol. 12, 109–117 (2017).
Malik, D. J. et al. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv. Colloid Interface Sci. 249, 100–133 (2017).
Międzybrodzki, R. et al. Pharmacokinetic and pharmacodynamic obstacles for phage therapy from the perspective of clinical practice. Clin. Infect. Dis. 77, S395–S400 (2023).
Dąbrowska, K. Phage therapy: what factors shape phage pharmacokinetics and bioavailability? Systematic and critical review. Med. Res. Rev. 39, 2000–2025 (2019).
Acknowledgements
No specific funding was granted for this study. Aye Mya Sithu Shein was supported by Chulalongkorn University (Second Century Fund- C2F Fellowship). Dhammika Leshan Wannigama was supported by Chulalongkorn University (Second Century Fund- C2F Fellowship), the University of Western Australia (Overseas Research Experience Fellowship), and Yamagata Prefectural Central Hospital, Yamagata, Japan (Clinical Residency Fellowship). Anthony Kicic is a Rothwell Family Fellow. The sponsor(s) had no role in the study design; in the collection, analysis, and interpretation of data; in the writing of the report; or in the decision to submit the article for publication. We, the authors of this paper, embrace inclusive, diverse, and equitable conduct of research. Our team comprises individuals who self-identify as underrepresented ethnic minorities, gender minorities, members of the LGBTQIA+ community, and individuals living with disabilities. We actively promote gender balance in our reference list while maintaining scientific relevance.
Author information
Authors and Affiliations
Contributions
D.L.W., S.A, P.G.H. and A.Ki. conceived the idea and formatted the structure of the manuscript. A.M.S.S and P.H. wrote the main body, A.K., P.P., K.M., L.C., K.S., M.A., P.M, and T.C. reviewed and added to the manuscript. The authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
A.M.S.S., P.H., A.K., P.P., K.M., L.C., K.S., M.A., P.M, A.Ki., T.C., P.G.H., and S.A. declare no financial or non-financial competing interests. D.L.W. serves as an editorial board member of this journal and had no role in the peer-review or decision to publish this manuscript. D.L.W. declares no financial competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Sithu Shein, A.M., Hongsing, P., Khatib, A. et al. Phage therapy could be key to conquering persistent bacterial lung infections in children. npj Antimicrob Resist 2, 31 (2024). https://doi.org/10.1038/s44259-024-00045-4
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s44259-024-00045-4
This article is cited by
-
R-pyocins as targeted antimicrobials against Pseudomonas aeruginosa
npj Antimicrobials and Resistance (2025)
-
Intranasal phage therapy overcomes antibody neutralization challenges in pulmonary Pseudomonas aeruginosa infections
Archives of Microbiology (2025)
-
Phage cocktail amikacin combination as a potential therapy for bacteremia associated with carbapenemase producing colistin resistant Klebsiella pneumoniae
Scientific Reports (2024)
