
Abstract
The area and elevation of the Tibetan Plateau over time has directly affected Asia’s topography, the characteristics of the Asian monsoon and modified global climate, but in ways that are poorly understood. Charting the uplift history is crucial for understanding the mechanisms that link elevation and climate irrespective of time and place. While some palaeoelevation data are available for southern and central Tibet, clues to the uplift history of northern Tibet remain sparse and largely circumstantial. Leaf fossils are extremely rare in Tibet but here we report a newly discovered early Miocene barberry (Berberis) from Wudaoliang in the Hoh-Xil Basin in northern Tibet, at a present altitude of 4611 ± 9 m. Considering the fossil and its nearest living species probably occupied a similar or identical environmental niche, the palaeoelevation of the fossil locality, corrected for Miocene global temperature difference, is estimated to have been between 1395 and 2931 m, which means this basin has been uplifted ~2–3 km in the last 17 million years. Our findings contradict hypotheses that suggest northern Tibet had reached or exceeded its present elevation prior to the Miocene.
Similar content being viewed by others
Introduction
Despite the critical role Tibet plays in understanding the processes linking subcrustal dynamics to climate decades of research have failed to produce a clear view of the plateau’s elevation history1,2. Data evidencing the elevation of different parts of Tibet over time have been mainly divided into two types3. One represents an indirect analysis2,3,4, such as crustal movement models based on geomorphological evidence or rates of sedimentation. The other one constitutes a direct quantitative estimation of Tibet’s palaeoaltimetry, mostly based on δ18O values in carbonate sediments5,6,7,8,9,10, but with two case studies utilizing the climate leaf analysis multivariate program (CLAMP)11,12.
Current opinions consider that south central Tibet reached or exceeded its current elevation by the Miocene and since then there has been minimal uplift, or possibly even a 1 km deflation5,6,12. By 11 Ma (late Miocene) the Thakkhola Graben appears to have reached a height of 4500 ± 430 m to 6300 ± 330 m or 3800 ± 480 m to 5900 ± 350 m6, between 8 Ma and 2 Ma (the late Miocene to Pliocene) Gyirong was elevated to 5850 + 1410/−730 m7 and the Zanda Basin rose to between 4000–4500 m since 9.2–1 Ma8,13. In central Tibet, the Lunpola Basin reached about 4000 m by 35 ± 5 Ma9 and the Nima Basin had an elevation of 4500–5000 m at 26 Ma10.
Interpreting the history of northern Tibet Plateau’s uplift generates controversy due to the paucity of palaeoaltimetric data and a lack of consensus. For example quantitative estimations of palaeoaltimetry for the northern Tibetan Plateau, correspondingly, are also divided with estimates for the height of the Hoh-Xil Basin ranging from ~2000 m to 4000 m at 39–36 Ma14,15,16, while modelling studies suggest that either there has been a significant uplift of northern Tibet since the Miocene (~15 Ma)2,4, or that it was near or above its modern elevation at 20 Ma17.
Different methods used for estimating the palaeo-elevations of the Tibetan Plateau have their own advantages and disadvantages, which means cross validation is needed in order to overcome their innate shortcomings.
Palaeoelevation estimations based on biological evidence differ from those using geophysical methods in that the discoveries of biological evidence (fossil materials) are usually chance occurrences and rare. Nevertheless, the unique advantage of biological evidence is that fossils may be clearly identified to a particular taxon and if that taxon is extant its unique ecological niche can be used to infer past conditions, including palaeoelevation.
Here we report a fossil Berberis leaf from the early-middle Miocene sediments of the Wudaoliang Basin in northern Tibet, recovered from a present day altitude of 4600 m, whereas its living modern counterpart (B. asiatica) is confined to altitudes of 914–2450 m. It provides an opportunity to estimate the palaeoaltimetry of the Wudaoliang Basin during the early Miocene and to evaluate previous hypotheses regarding the uplift of northern Tibet.
Material
Geological context
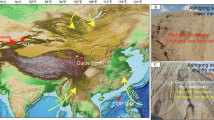
The solitary fossil leaf specimen that we can identify accurately was collected from a natural outcrop of lacustrine sediments of the Wudaoliang Group near Wudaoliang Town, Qinghai Province, northern Tibetan Plateau (Fig. 1A,B, 35°13’56.90“N, 93° 05'10.50“E, 4611 ± 9 m). The fossil leaf is preserved as an impression on a matrix of pale-yellow, fine-grained marl and is now housed in the Institute of Botany, Chinese Academy of Sciences in Beijing (specimen no. WDL 2010-001).

Map showing the fossil locality (A, B) and the stratigraphical section of the Wudaoliang Group (C).
A showing the Tibetan Plateau and quantitative estimations of palaeoaltimetry for northern Tibet. B showing the locations of our fossil (35°13’56.90“N, 93° 05'10.50“E, 4611 ± 9 m) and core ZK1 (35°14’18.40“N, 93° 05'22.80“E, 4666 m). C showing the stratigraphical section of core ZK118, including the ages at top and bottom of the Wudaoliang Group, which are in accordance with previous data from dating of the Wudaoliang volcanics20,21,22. The maps are created by authors using “ArcGIS 10” and “CorelDraw 14” software. The stratigraphical section is drawn by authors using “CorelDraw 14” software based on the data from refs. 18, 20, 21, 22.
The same block contains another specimen (see: SI, Figure S2) and numerous other specimens occur in the adjoining layers (see: SI, Figure S3). Most specimens were not well enough preserved to allow us to pin-point their systematic position but one fossil leaf of Berberis was sufficiently well preserved to allow its determination to species level and provide a palaeoelevation datum.
Age estimates for the fossil
In the study area, the ZK1 drill hole (see SI for core description), 720 m away from the fossil locality, penetrated the Wudaoliang Group to a depth of 154 m18. The dating of core ZK1 (Fig. 1) is calibrated using climatostratigraphic methods, based on a comparison between the palaeoclimatic cycles recorded by carbon and oxygen isotope changes within lacustrine deposits18 and the palaeoclimatic cycles recorded by the deep-sea oxygen isotope curve19. The ages at both top and bottom of the Wudaoliang Group calibrated in this way are in accordance with those from dating of Wudaoliang volcanics20,21,22 (see more details in SI). Based on carbon and oxygen isotope studies, the age of these lacustrine sediments is regarded as early to middle Miocene (24.1–14.5 Ma)18. Comparisons of the coordinates and altitude of our fossil layer with those of core ZK1, enabled us to correlate the fossil layer to a depth of 55 m in the ZK1 drill hole, hence an age of ca. 17 Ma. (Fig. 1C, see more details in SI).
Result
Systematic treatment
The detailed fossil description and taxonomic discussions are included in SI. The specific combination of leaf characters in the fossil such as its dentate margin with 4-5 fine spinose or setaceous teeth on each side, pinnate festooned brochidodromous venation, the presence of inter-secondary veins and random reticulate tertiary veins falls within the circumscription of the genus Berberis of the Berberidaceae23.
By eliminating the obviously different species in the genus (see the detailed elimination in SI) we were left with only 4 most similar species. The supplementary tables and keys (see SI, Table S1 – S2, Key S1 – S2) clearly show that two, or more than two, morphological character differences exist between our fossil specimen and the other 3 extant species except B. asiatica. A survey of herbarium material of Berberis revealed that these leaf architectural characters are stable at the species level and provide a robust basis for identification at the generic and species levels24.
The detailed comparison with fossil and extant species of Berberis (see SI for comparison, Fig. 2) indicates that the fossil cannot be distinguished from extant B. asiatica, for the leaf architectural characters of the fossil are identical to those of B. asiatica except that no veinlet are preserved in the fossil. If it were not for the fact that the specimen is a fossil, it would be unequivocally assigned to B. asiatica (Fig. 2C). However, considering its fossil state it is referred to B. cf. asiatica.

Leaf architecture of extant (C–F) and fossil (A, B, G–L) Berberis.
A, B. Fossil from Wudaoliang, C. B. asiatica, D. B. chitria, E. B. taronensis, F. B. phanera, G. B. huziokai, H. B. teutonica, I. B. lycium, J. B. poblana, K. B. longipetiolata, L. B. ahuehuetensis A, B, D, E, F, scale bar = 0.5 cm. C, G, H, I, J, K, L, scale bar = 1 cm. The photographs of fossil specimen (A) and extant Berberis (C–F) are taken by authors., the leaf architecture drawing (B) is drawn by authors using “CorelDraw 14” software and drawings (G–L) are cited from Ref. 23.
Modern B. asiatica grows on the southern slope of the Himalayas at altitudes ranging from 914 to 2450 m (Fig. 3 green range)25,26,27. The NLR (Nearest Living Relative) concept28, assumes that the fossil B. cf. asiatica and its nearest living species B. asiatica occupied similar or identical niches and lived at correspondingly similar altitudes. Normally such an assumption could not be justified but extensive analysis of fossil occurrences in Eurasia spanning the Miocene to present using the Co-Existence Approach29 indicate that the genus Berberis does not occur as an outlier (Utescher, T., personal communication August 2014, see the e-mail permission messages in SI) and thus can be regarded as a conservative genus. On this basis the altitudinal range of B. cf. asiatica is inferred to be similar to or the same as that of B. asiatica.

The uplift of fossil locality.
Green range: altitudinal range of B. asiatica, Pink range: altitudinal range of B. cf. asiatica. ΔHmin: the minimum value of uplift, ΔHmax: the maximum value of uplift. This figure is drawn by authors using “CorelDraw 14” software.
Discussion
Altitude of northern Tibet during the early Miocene
To correct for secular climate differences between the middle Miocene and the present we used both modelled lapse rates and enthalpy30. Assuming that the mid-Miocene was 2.89 °C warmer than now20,30,31 and taking the lapse rate to be 6.01 °C/1000 m11,30, the maximum altitude at which Berberis asiatica could have grown in the early Miocene was 481 m higher than present due solely to this climatic difference. Therefore, when B. cf. asiatica was flourishing in the early Miocene the palaeoaltitude of the Wudaoliang Basin could have been no more than 1395–2931 m (Fig. 3 pink range). It therefore follows that because our fossil was found at a present day elevation of 4611 m the Wudaoliang Basin must have been uplifted by 1680–3216 m (Fig. 3) in the last 17 Ma.
The uplift of northern Tibet
In recent years, both geophysical modelling and palaeoaltimetry studies have generated different hypotheses concerning the uplift history of the northern Tibetan Plateau. Some studies suggest northern Tibet attained today’s elevation in the Miocene17 while the “far field deformation” hypothesis advocates northern Tibetan uplift preceding that of southern Tibet as early as the middle to late Eocene32. Other views such as the ‘stepwise model’ propose that northern Tibet only achieved today’s elevation after the Miocene2,33.
Our estimate for the Wudaoliang Basin suggests a palaeoelevation of 1395–2931 m during the early Miocene, which is close to an estimation for the same area based on δ18O values in late Eocene carbonate sediments14. This shows that the elevation of northern Tibet probably remained stable from the late Eocene to early Miocene but since then there has been a considerable uplift (~2000 m).
Our finding does not support earlier views that northern Tibet had reached or even exceeded its modern elevation before the Miocene17,32, but seems to be close to the predictions of stepwise uplift models, suggesting a significant uplift of northern Tibet since Miocene time2,33. Here the palaeoaltitude of northern Tibet estimated by using new independent biological evidence provides cross validation of results from geophysical models and geochemical evidence.
Methods
The fossil leaf architecture characters were exposed by dégagement and examined under a stereomicroscope and an environmental scanning electron microscope. The leaf architecture is described following the terminology of the Leaf Architecture Working Group24. The comparative extant Berberis material came from the PE Herbarium of the Institute of Botany, Chinese Academy of Sciences and the G.B. Pant Institute of Himalayan Environment and Development, India. It was cleared in a 10% aqueous solution of NaOH.
Palaeoaltimetry correction
Because the global climate has undergone a secular change in temperature of about 2.89 °C since the early Miocene20,30,31, a correction factor based on the palaeotemperature has to be applied when estimating palaeoaltitude. Assuming the Miocene lapse rate to be 6.01 °C/1000 m30, the corrected altitude is derived as follows:

(Where ΔHT is the correction in palaeoaltitude and ΔT is the temperature difference between the Miocene and today).
Additional Information
How to cite this article: Sun, B. et al. Early Miocene elevation in northern Tibet estimated by palaeobotanical evidence. Sci. Rep. 5, 10379; doi: 10.1038/srep10379 (2015).
References
Dewey, J. Extensional collapse of orogens. Tectonics 7, 1123–1139 (1988).
Tapponnier, P. et al. Oblique stepwise rise and growth of the Tibet Plateau. Science 294, 1671–1677 (2001).
Molnar, P., Boos, W. R. & Battisti, D. S. Orographic controls on climate and palaeoclimate of Asia: thermal and mechanical roles for the Tibetan Plateau. Annu. Rev. Earth Planet. Sci. 38, 77–102 (2010).
Feldl, N. & Bilham, R. Great Himalayan earthquakes and the Tibetan Plateau. Nature 444, 165–170 (2006).
Currie, B. S., Rowley, D. B. & Tabor, N. J. Middle Miocene palaeoaltimetry of southern Tibet: implications for the role of mantle thickening and delamination in the Himalayan orogen. Geology 33, 181 (2005).
Garzione, C. N., Quade, J., DeCelles, P. G. & English, N. B. Predicting palaeoelevation of Tibet and the Himalaya from δ18 O vs. altitude gradients in meteoric water across the Nepal Himalaya. Earth Planet. Sci. Lett. 183, 215–229 (2000).
Rowley, D. B., Pierrehumbert, R. T. & Currie, B. S. A new approach to stable isotope-based palaeoaltimetry: implications for palaeoaltimetry and palaeohypsometry of the High Himalaya since the late Miocene. Earth Planet. Sci. Lett. 188, 253–268 (2001).
Saylor, J. et al. The late Miocene through present palaeoelevation history of southwestern Tibet. Am. J. Sci. 309, 1–42 (2009).
Rowley, D. B. & Currie, B. S. Palaeo-altimetry of the late Eocene to Miocene Lunpola Basin, central Tibet. Nature 439, 677–681 (2006).
DeCelles, P. G. et al. High and dry in central Tibet during the late Oligocene. Earth Planet. Sci. Lett. 253, 389–401 (2007).
Spicer, R. A. et al. Constant elevation of southern Tibet over the past 15 million years. Nature 421, 622–624 (2003).
Khan, M. A. et al. Miocene to Pleistocene floras and climate of the eastern Himalayan Siwaliks and new palaeoelevation estimates for the Namling–Oiyug Basin, Tibet. Global and Planetary Change 113, 1–10 (2014).
Deng, T. et al. Locomotive implication of a Pliocene three-toed horse skeleton from Tibet and its palaeo-altimetry significance. Proc. Natl. Acad. Sci. 109, 7374–7378 (2012).
Cyr, A. J., Currie, B. S. & Rowley, D. B. Geochemical evaluation of Fenghuoshan Group lacustrine carbonates, north-central Tibet: Implications for the palaeoaltimetry of the Eocene Tibetan Plateau. J. Geol. 113, 517–533 (2005).
Quade, J., Breecker, D. O., Daëron, M. & Eiler, J. The palaeoaltimetry of Tibet: An isotopic perspective. Am. J. Sci. 311, 77–115 (2011).
Polissar, P. J., Freeman, K. H., Rowley, D. B., McInerney, F. A. & Currie, B. S. Palaeoaltimetry of the Tibetan Plateau from D/H ratios of lipid biomarkers. Earth Planet. Sci. Lett. 287, 64–76 (2009).
Wang, C. S. et al. Constraints on the early uplift history of the Tibetan Plateau. Proc. Natl. Acad. Sci. 105, 4987–4992 (2008).
Wu, Z. H. et al. Carbon and oxygen isotope changes and palaeoclimate cycles recorded by lacustrine deposits of Miocene Wudailiang Group in northern Tibetan Plateau. Geol. China. 36, 966–975 (In Chinese with English abstract) (2009).
Zachos, J., Pagani, M., Sloan, L., Thomas, E. & Billups, K. Trends, rhythms and aberrations in global climate 65 Ma to present. Science 292, 686–693 (2001).
Wu, Z. H. et al. Oligocene thrust systems in central Tibetan Plateau. Geol. China. 38, 522–536 (In Chinese with English abstract) (2009).
Li, J. D. et al. Ages of volcanic rocks and planation surface in the Canmei Mountain area, northern Tibet. Regional Geology of China 7, 005 (2004).
Blisniuk, P. M., Hacker, B. R., Glodny, Johannes et al. Normal faulting in central Tibet since at least 13.5 Myr ago. Nature, 412, 628–632 (2001).
Li, Y. L. et al. The fossil record of Berberis (Berberidaceae) from the Palaeocene of NE China and interpretations of the evolution and phytogeography of the genus. Rev. Palaeobot. Palynol. 160, 10–31 (2010).
Ellis, B. et al. in Manual of Leaf Architecture. (Cornell University Press, New York, 2009)
Hooker, J. D. in The Flora of British India: Volume 4(L. Reeve & Co., London, 1885)
Andola, H. C., Gaira, K. S., Rawal, R. S., Rawat, M. S. & Bhatt, I. D. Habitat-dependent variations in berberine content of Berberis asiatica Roxb. ex DC. in Kumaon, Western Himalaya. Chemistry & Biodiversity 7, 415–420 (2010).
Andola, H. C., Gaira, K. S., Rawal, R. S., Rawat, M. S. & Bhatt, I. D. Influence of environmental factors on production of berberine Content in Berberis asiatica Roxb. ex DC in Kumaun West Himalaya, India. Journal of Herbs, Spices & Medicinal Plants. 17, 329–338 (2011).
Mosbrugger, V. Nearest-living-relative method. In Encyclopedia of Palaeoclimatology and Ancient Environments 607–609 (Springer Netherlands, 2009).
Mosbrugger, V., Utescher, T. The coexistence approach—a method for quantitative reconstructions of Tertiary terrestrial palaeoclimate data using plant fossils. Palaeogeography, Palaeoclimatology, Palaeoecology. 134, 61–86 (1997).
Song, X. Y., Spicer, R. A., Yang, J., Yao, Y. F. & Li, C. S. Pollen evidence for an Eocene to Miocene elevation of central southern Tibet predating the rise of the High Himalaya. Palaeogeography, Palaeoclimatology, Palaeoecology. 297, 159–168 (2010).
Billups, K. & Schrag, D. P. Palaeotemperatures and ice volume of the past 27 Myr revisited with paired Mg/Ca and 18O/16O measurements on benthic foraminifera. Palaeoceanography 17, 1003 (2002).
Dayem, K. E., Molnar, P., Clark, M. K. & Houseman, G. A. Far-field lithospheric deformation in Tibet during continental collision. Tectonics 28, TC6005 (2009).
Mulch, A. & Chamberlain, C. P. Earth science: The rise and growth of Tibet. Nature 439, 670–671 (2006).
Acknowledgements
We thank Dr. Mukund Sharma, Birbal Sahni Institute of Palaeobotany, Lucknow, India for reading and improving the first version of the manuscript. This research was supported by China National Key Basic Research Program (2014CB954201), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB03020104), the State Key Laboratory of Loess and Quaternary Geology, Institute of Earth Environment, Chinese Academy of Sciences (SKLLQG1333) and the National Natural Science Foundation of China (Nos. 30990241; 41072022, 41210001, 31370254, 31300186). Prof. David K. Ferguson and Prof. Robert A. Spicer were supported by the Chinese Academy of Sciences Visiting Professorship for Senior International Scientists at the Institute of Botany (Nos. 90004F1005 and 2009S1-20 respectively).
Author information
Authors and Affiliations
Contributions
Y.F.W. and C.S.L. conceived the ideas; M.Z., S.Q.W. and T.D. collected the fossils; B.S., Y.L.L. and Y.F.W. identified the fossil and analyzed the data; B.S. and Y.F.W. wrote the first draft of this manuscript; C.S.L., J.Y., J.F.L., T.D. and R.C.M. revised the draft versions. R.A.S. and D.K.F. rewrote some of the discussion and corrected the final manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Sun, B., Wang, YF., Li, CS. et al. Early Miocene elevation in northern Tibet estimated by palaeobotanical evidence. Sci Rep 5, 10379 (2015). https://doi.org/10.1038/srep10379
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep10379
This article is cited by
-
Mid-Miocene sea level altitude of the Qaidam Basin, northern Tibetan Plateau
Communications Earth & Environment (2023)
-
Cenozoic plants from Tibet: An extraordinary decade of discovery, understanding and implications
Science China Earth Sciences (2023)
-
Integrative Neogene stratigraphy, biotas and paleogeographic evolution of the Tibetan Plateau and its surrounding area
Science China Earth Sciences (2023)
-
Timing and mechanisms of Tibetan Plateau uplift
Nature Reviews Earth & Environment (2022)
-
Fossil climbing perch and associated plant megafossils indicate a warm and wet central Tibet during the late Oligocene
Scientific Reports (2017)

Thomas Denk
Comment to Sun et al. ?Early Miocene elevation in northern Tibet estimated by palaeobotanical evidence?
Thomas Denk, Swedish Museum of Natural History, Department of Palaeobiology, Box 50007, 10405 Stockholm, Sweden
Email: thomas.denk@nrm.se
In their recent contribution, Sun et al.<sup>1</sup> used a fossil leaf to infer the elevation of northern Tibet during the early Miocene (ca 17 Ma). The authors compared the fossil leaf to a particular species of barberries, Berberis asiatica Roxb. ex DC., which today occurs in Nepal and the Tibetan Plateau^2^ at elevations between 914 and 2,450 m^1^ or 1,000 to 2,700 m a.s.l. <sup>2</sup>. Sun and colleagues assumed that the fossil taxon and its potential modern counterpart (its ?nearest living relative?, NLR) occupied similar or identical niches and, hence, must have lived at correspondingly similar altitudes. Applying a palaeoaltimetric correction they added 481 m to the modern elevation of B. asiatica and inferred a palaeoelevation of 1,395?2,931 m for northern Tibet at 17 Ma. Since the fossil was collected at an elevation of 4,611 m in the Wudaoling Basin, the authors concluded that the basin must have been uplifted by 1,680?3,216 m during the last 17 Myrs.
There is great risk in ascribing a single fossil leaf impression to any particular modern species and this procedure risks inducing first-level errors if the chosen NLR turns out to be inappropriate. Sun and colleagues chose four modern species out of about 500 living Berberis species as potential closest extant relatives and further narrowed down their selection of NLR to a single modern species.
There are considerable problems with the study of Sun et al. related to the choice of the NLR, the assumption of niche conservatism in Berberis, and consequently, the inference of palaeoelevation of the Tibet Plateau.
It is unclear how exactly the selection of the single NLR was made by Sun et al. There is no reference to the single monographic study of Berberis s. l. by Ahrendt^3^ and it is unclear how a previous study^4^ contributes to the selection of the NLR by Sun et al. The criteria used by Sun et al. to discriminate species suitable as NLR for the fossil leaf not only apply to the four species that they selected but also to a range of other species. Leaves virtually identical to the fossil are encountered in the following species^2, 3, 5^ ( Fig. 1 ): Berberis aristata DC., B. concinna Hook.f, B. jaeschkiana C.K.Schneid., B. koehneana C.K.Schneid., B. orthobotrys Bienert ex Aitch., and to a lesser degree in B. everestiana Ahrendt and B. asiatica Roxb. ex DC. This has immediate consequences for the inference of palaeoelevation using the methodology of Sun et al. ( Table 1 ). Notably, most estimates using this broader selection of taxa exceed or equal the current elevation of the Wudaoling Basin but at the same time do not rule out uplift of more than 3 km (based on the lowest occurrence of B. asiatica) during the last 17 Myrs.
Since the validity of the conclusions presented by Sun et al. depends solely on how well the systematic affinity of the fossil leaf is resolved, it comes as a surprise that no attempt was made to place the fossil taxon and the potential modern analogue(s) used as NLR into a general phylogenetic context. For example, a recent molecular phylogeny of Berberis<sup>6</sup> identified B. asiatica as part of a group of species growing in the Himalayas and Tibet ( Fig. 2, clade 7.2 ). The mean crown age of this clade is 13.1 Ma based on a time-calibrated phylogeny^7^. Thus, the fossil from the Wudaoling Basin may represent the lineage leading to the modern members of clade 7.2, which includes species growing today at very high elevations. Other species qualifying as a potential modern analogue (B. aristata, B. concinna, B. koehneana and B. orthobotrys var. rubicunda) of the fossil taxon occur in a sister clade that also comprises Himalayan and Tibetan species. According to the time-calibrated phylogeny^7^ all modern species mentioned here diverged after the late Miocene to Pliocene. It is clear from this that the appropriate NLR for the fossil taxon from the Wudaoling Basin cannot be a single species but a group of species representing an entire clade.
Despite these problems, there may be some reality in the palaeoelevation inferred by Sun et al. Considering only clade 7.2, the early Miocene fossil perhaps belongs to the lineage leading to the modern members of this clade and may have had ecological/distributional properties similar to B. asiatica. At the same time, there is no justification for disregarding the ecological/distributional properties of other species in this clade. The first appropriate step in such a study would have been for the authors to demonstrate that there is a general trend in this sublineage of Berberis to extend the distributional range into higher elevations. This would permit the argument that the lowest-thriving species is a better modern analogue for a putative ancestral member of the clade than its high-montane relatives and morphologically similar forms from the sister clade.
Contrary to the statement by Sun et al. Berberis is not a ?conservative genus? but evolved into a wide range of biomes^2, 3, 6, 7^. Adaptation to current high elevation habitats may have happened in parallel with the uplift of the Himalayas and is not restricted to one particular clade in the phylogeny of Berberis<sup>6</sup>. It is impossible, based on the available data, to predict at which altitude a fossil Berberis species thrived or to know when and where elevational shifts occurred in such a diverse and opportunistic group.
Other proxy data have recently suggested moderate uplift between 23 and 18?16 Ma based on excellently preserved fish fossils^8^. Cyprinids in Oligocene deposits of the Tibet Plateau have significantly fewer vertebrae than fossils from early Miocene deposits. Since there appears to be a functional relationship between number of vertebrae and water temperature, fossils cyprinids appear to be a more reliable proxy than the single leaf fossil reported by Sun et al.
(1) Sun, B., Wang, Y.-F., Li, C.-S., Yang, J., et al. 2015. Early Miocene elevation in northern Tibet estimated by palaeobotanical evidence. Scientific Rep. 5: 10379. DOI: 10.1038/sreo10379.
(2) Adhikari, B., Pendry, C. A., Pennington, R. T., Milne, R. I. 2012. A revision of Berberis s.s. (Berberidaceae) in Nepal. Edinburgh J. Bot. 69: 447?522.
(3) Ahrendt, L. W. A. 1961. Berberis and Mahonia. Journal of the Linnean Society, Botany 57: 1?369.
(4) Li, Y.-L., Kva?ek, Z., Ferguson, D. K., Wang, Y.-F., Li, C.-S., et al. 2010. The fossil record of Berberis (Berberidaceae) from the Palaeocene of NE China and interpretations of the evolution and phytogeography of the genus. Rev. Palaeobot. Palynol. 160: 10?31.
(5) Royal Botanic Garden Edinburgh. 2015. Herbarium Catalogue. "http://elmer.rbge.org.uk/bg...": http://elmer.rbge.org.uk/bg.... Accessed May 26, 2015.
(6) Adhikari, B., Milne, R., Pennington, R. T., Särkinen, T. et al. 2015. Systematics and biogeography of Berberis s.l. inferred from nuclear ITS and chloroplast ndhF gene sequences. Taxon 64: 39?48.
(7) Adhikari, B. 2010. Systematics and Phylogeographic studies of Berberis L. (Berberidaceae) in the Nepal Himalaya. Doctoral thesis, The University of Edinburgh, Royal Botanical Garden Edinburgh.
(8) Wang, N., Wu, F., 2015. New Oligocene cyprinid in the central Tibetan Plateau documents the pre-uplift tropical lowlands. Ichthyol. Res. 62: 274?285.