
Abstract
Li Fraumeni syndrome (LFS) is a rare familial cancer predisposition syndrome with autosomal-dominant inheritance, occurring as frequently as one in 5,000–20,000 individuals. However, no LFS case has been reported from mainland China although it constitutes one quarter of population on earth. In this study, we identified, to our best knowledge, the first Li Fraumeni syndrome family in China. Six family members were affected with various tumors. A TP53 mutation (c.730G > A; p.G244S) co-segregated with the tumor phenotype within this family. Functional analysis indicated that G244S mutation disrupted the transactivity, DNA-binding and cell growth inhibition activity of p53 protein. Two available tumor samples (medulloblastoma and choroid plexus papilloma) underwent large rearrangement in the chromosomes and loss of wild-type TP53. Our data warranted further studies on the prevalence of germline TP53 mutation in various tumor patients in China.
Similar content being viewed by others

Introduction
Li Fraumeni syndrome (LFS), first introduced by Li and Fraumeni in 1969, is a rare familial cancer predisposition syndrome with autosomal-dominant inheritance1,2. The clinical definition of LFS includes a proband with a sarcoma under the age of 45 years, a first-degree relative with a cancer prior to the age of 45 years and a first- or second-degree relative with any cancer before 45 years or a sarcoma at any age3. The major tumors in LFS include soft tissue and bone sarcomas, breast cancer, brain tumors, adrenocortical carcinomas and acute leukemia, et al.4,5,6,7.
The risk of individuals with LFS to develop any invasive cancer is ~50% by age 30 and is 90% by age 708. Females with this syndrome have almost a 100% lifetime risk of developing cancer, whereas the risk for affected males is 73%9. Individuals with LFS are also prone to develop second, third, even fourth malignant neoplasms, with the highest risk of additional cancers being in those diagnosed with their first cancer during childhood.
Approximately 70% of families with LFS have a mutation in the TP53 gene10. TP53 encodes the p53 tumor suppressor, a critical transcription factor that promotes cell-cycle arrest, apoptosis and DNA repair in response to cellular stresses such as exposure to ionizing radiation11,12,13,14,15,16,17,18,19,20. Mutations that interfere with the transcriptional activity of p53 reduce its growth suppressive functions. Moreover, mutant p53 proteins may also acquire oncogenic gain of function21,22. For instance, mice with one mutant allele equivalent to human R175H, R273H, or R248Q showed more spontaneous carcinomas, sarcomas and lymphomas than heterozygous or null (TP53+/− or TP53−/−) mice23,24,25,26.
LFS may be as frequent as one in 5,000–20,000 individuals27. Until now, there are more than 500 LFS families have been identified worldwide10. Although it constitutes one quarter of population on earth, no LFS has been reported from mainland China. In this study, we identified a family with multiple types of tumors. A TP53 mutation (c.730G > A; p.G244S) was identified in the patients. This mutation led to functional disruption of p53 protein.
Results and Discussion
Clinical findings of the family
In 2014, the proband (III: 4, a 5-year-old girl) was diagnosed with a tumor located in the left cerebellar hemisphere (Fig. 1A1, 2) in the Department of Neurosurgery at Xiangya Hospital, Central South University of China. The mass was removed surgically and diagnosed as classical medulloblastoma pathologically (Fig. 1B,C). The proband undertook a standard radiotherapy after surgery and no recurrence was detected after 18 months (Fig. 1A3).

The Magnetic Resonance Imaging (MRI) and histology of tumors from the proband (III: 4) and her brother (III: 5).
(A) The MRI of the proband’s tumor (indicated with arrows) located in the left cerebellar hemisphere (A1-A2). No recurrence was detected at 18 months post-surgery (A3). (B) The haematoxylin and eosin (H&E) staining of the proband’s tumor sample. (C) The immunohistochemistry of proband’s tumor sample stained with neuron specific enolase (NSE) antibody. (D) The MRI of the III: 5’s mass (indicated with arrows) in the posterior of left lateral ventricle (D1-D2). No recurrence was detected at 18 months post-surgery (D3). (E) The H&E staining of the III: 5′s tumor sample. (F) The immunohistochemistry of III: 5′s tumor sample stained with pan cytokeratin antibody. Scale bar: B and D: 200 μm; C and E, 100 μm.

The patient information.
(A) The pedigree of this family. Subjects in the family are identified by the Roman and Arabic numerals below the symbol, in which the Roman numerals denote the generations. Open symbols = unaffected; filled symbols = affected; symbols with a diagonal line = deceased subjects; squares = male; circles = female; arrow = the proband; WT = wild-type TP53 allele; Mut = G244S mutant TP53 allele. (B) The patient information including age, tumor type and tumor onset age.
Soon later, the proband’s younger brother (III: 5, a 3-year-old boy), was accidentally diagnosed with a mass in the posterior of left lateral ventricle (Fig. 1D1, 2). After a craniotomy, it was proved to be choroid plexus papilloma (Fig. 1E,F). The mass was removed by surgery and no recurrence was detected after 18 months (Fig. 1D3).
The fact that two individuals in a family carried tumors at very young age caught our attention; we therefore surveyed the tumor history in this family. As shown in Fig. 2A,B, the proband’s mother (II: 4) was diagnosed with ductal carcinoma in situ by breast needle biopsy at the age of 34 y in 2013. No pathogenic mutation was found in genes BRCA1 or BRCA2 by Sanger sequencing. She underwent a radical operation later and was well in the two-year follow-up. The proband’s aunt (II: 2) underwent a modified radical mastectomy and was diagnosed as occult breast cancer in 2010; she died of distant metastasis in 2013 despite of standard chemotherapy. The tumor samples from II: 2 was demonstrated to be negative in ER and PR by immunohistochemistry. The oldest daughter (III: 1) of patient II: 2 underwent two surgical treatments. She was diagnosed with adrenal pheochromocytoma at 3 y and renal cyst at 12 y. Moreover, the proband’s maternal grandfather died with a liver mass at his 40 s. Accordingly, the tumors in this family were aggregated and likely to be Li Fraumeni syndrome.
TP53 mutation in the family
To understand the genetic basis of this family, we performed exome sequencing on the proband and her parents28. As a result, we generated an average of 55.6 million reads that passed the quality assessment and were aligned to the human reference sequence. The mean sequencing depth was 64 and an average of 97.5% sequences was covered by more than 10 times. We identified an average of 40,934 variants and an average of 10,194 NS/SS/Indel (non-synonymous/splice acceptor and donor site/ insertions or deletions) variants was located in the coding regions. After filtering common variants in the databases, we identified 57 gene mutations shared by the proband and her mother, including a TP53 mutation (c.730G > A; p.G244S) (Fig. 3A).

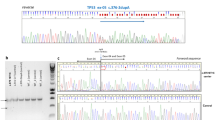
G244S mutation of the TP53 gene.
(A) Sanger sequencing of codons 727–735 of wild-type (WT) and c.730G > A (p.G244S) mutant TP 53 genes. (B) Schematic diagram of TP53 gene and p53 protein. The G244S mutation identified in this study was indicated with an arrow. TAD: transcription-activation domain; PRD: proline-rich domain; DBD: DNA-binding domain; NLS: nuclear localization signal; OD: homo-oligomerization domain. (F) Conservative analysis of the Glycine 244-containing portion in p53 protein. The Glycine 244 was highlighted.
As TP53 is mutated in more than 50% tumors and is the major causative gene for Li Fraumeni syndrome, we analyzed the co-segregation of this variant by polymerase chain reaction (PCR) and direct sequencing in this family. As shown in Fig. 2A,B, G244S TP53 mutation was identified in all available patients. This mutation was also identified in an unaffected individual (III: 3), a 6-year-old boy. Therefore, G244S TP53 mutation co-segregated in this family. Patients (or their parents, if younger than 14 y) were informed consent for the molecular testing.
G244S mutation occurred in the hot spot mutation areas of TP53 gene (Fig. 3B). Glycine 244 was conserved among species from drosophila to human (Fig. 3C). G244S alteration was predicted to be damaging as assayed with functional prediction tools, including PolyPhen-2 (http://genetics.bwh.harvard.edu/pph2/), Mutation Taster (http://www.mutationtaster.org/), Pmut (http://mmb2.pcb.ub.es:8080/PMut/) and SIFT (http://sift.jcvi.org).
In the International Agency for Research on Cancer (IARC) database (http://www.iarc.fr), there is a LFS family from Malaysia carried the G244S TP53 mutation. Nevertheless, there seems no relationship between Malaysia family and the one reported here. Rather, the G244S seems to be a De Novo mutation occurred in patient I: 1, as his parents and siblings survived more than 65 years old with tumor-free.
G244S mutation disrupted p53 function
G244S mutation significantly increased the half-life of p53 protein. As shown in Fig. 4A,B, wild-type (WT) p53 degraded rapidly, with a half-life of ~3 h. However, no significant degradation of G244S p53 was detected in 8 h after cycloheximide treatment. Both WT and G244S p53 proteins were localized in the nuclei (Fig. 4C). To study the functional consequences of G244S mutation, WT or G244S p53-expressing constructs were co-transfected with a luciferase construct under the control of p21 promoter into TP53-deficient HCT116 cells. As shown in Fig. 4D, WT, but not G244S p53 elicited a significant increase in luciferase activity, which was also observed in other cell lines such as SaoS-2 and H1299 cells (data not shown).

G244S mutation disrupted the p53 function.
(A) G244S mutation increased the half-life of p53. TP53-deficient HCT116 cells were transiently transfected with plasmids expressing either the WT or G244S p53. 48 h later, cells were treated with cycloheximide (CHX) for 0, 1, 2, 4, 6 and 8 h. Equal amounts of whole cell lysates were analyzed by Western blot with a p53 antibody. Actin was used as an internal control. (B) The graph illustrates the quantification of WT and G244S p53 by densitometry of triplicate experiments (mean ± SEM). *p < 0.05, **p < 0.01 by post hoc Bonferroni t test. (C) The subcellular localization of WT and G244S p53 proteins as analyzed with immunofluorescence. Scale bar = 10 μm. (D) Transcriptional activities of WT and G244S p53 as determined by luciferase activity assays. The luciferase reporter plasmid p21-Luc were transiently transfected into TP53-deficient HCT116 cells in combination with WT or G244S p53 expressing plasmids. **p < 0.01 by post hoc Dunnett’s t test (E) The dominant negative effect of G244S p53 on WT p53. Increasing amount of G244S p53-expressing plasmid was cotransfected with a fixed amount of WT p53-expressing plasmid and the p21-Luc into TP53-deficient HCT116 cells. The basal level of luciferase was set as 1. Data from all other transfection are presented as fold induction above this level. Each value was the mean ± SEM of three replicates from a single assay. The results shown were representative of three independent experiments. **p < 0.01 by post hoc Bonferroni t test. (F) The induction of p21 after transfection of WT or G244S p53-expressing plasmids in TP53-deficient HCT116 cells. The protein levels of p53, p21 and β-actin was analyzed with immunoblotting. (G) The ability of WT and G244S p53 to bind corresponding DNA as assayed with a chromatin immunoprecipitation (ChIP) assay in TP53-deficient HCT116 cells. (H) The growth curve of SaoS2 cells stably transfected with WT or G244S TP53, as measured with a MTT assay. The cell numbers at day 1 was set to 1. **p < 0.01, post hoc Bonferroni t test.
To see if G244S p53 has dominant negative effect on the WT p53, the p21-luciferase construct was co-transfected with WT p53-expressing plasmid and increased doses of G244S p53-expressing constructs. Although high doses of G244S p53 inhibited the transactivity of WT p53, only minimal inhibition was observed when they were at a 1:1 ratio (Fig. 4E).
The TP53-deficient HCT116 cells were transfected with WT and G244S TP53 gene and the protein levels of p53 and p21 were assayed with Western blot. As shown in Fig. 4F. WT p53 significantly increased the p21 level, whereas G244S p53 showed no effect. As Glycine 244 was located in the DNA-binding domain of p53, we performed a chromatin immunoprecipitation (ChIP) assay in the TP53-deficient HCT116 cells to see if G244S mutation disrupted the DNA-binding activity of p53. As shown in Fig. 4G, the ability of p53 to pulldown p21 promoter was disrupted by G244S mutation.
The p53 protein potently inhibited cell growth. To study the physiological consequence of G244S mutation, WT and G244S TP53 were stably transfected into TP53-deficient SaoS-2 cells and their growth curve was assayed. As shown in Fig. 4H, transfection of WT TP53 significantly suppressed the cell growth, which is absent when G244S TP53 was transfected.
Loss of wild-type TP53 in the tumor samples
Consistent with most Li Fraumeni families, the tumor onset age became younger with successive generations. It has been suggested that the anticipation pattern may result from increased DNA copy-number variations (CNVs) with successive generations29. However, this phenomena was not verified in a whole genome sequencing study30. We performed karyotype as well as CNVs analysis on this family and did not identify tumor-associated chromosome abnormality or CNVs. We did not either see a trend of increase in the CNVs numbers with successive generations (data not shown). We also performed CNVs analysis on the tumor samples from III: 4 and III: 5, there were 95 and 63 CNVs in the tumor samples from III: 4 and III: 5, respectively, covering about 50% of the chromosome (Fig. 5).

The Copy Number Variations (CNVs) identified in the tumor samples from III:4 (A) and III:5 (B).
CNVs were analyzed with an Illumina Human Cyto SNP12, followed by Illumina’s Genome Studio Genotyping Module. The arrows indicated loss-of-heterozygosity (LOH) of chromosome 17, where TP53 gene was located.
About 50% tumors from the LFS patients showed loss-of-heterozygosity (LOH), in which the functional wild-type TP53 was lost. When checking the CNVs from III: 4 and III: 5’s tumor samples, we found that both tumors have two identical copies of chromosome 17 where TP53 gene was located (Fig. 5). We therefore performed a PCR-Sanger sequencing and found the G244S TP53 mutation was homozygous in the tumor sample, in contrast to the heterozygous status in the germline. Quantitative PCR analysis indicated that there were two copies of mutant TP53 gene in both tumor samples.
In summary, we have identified, to our best knowledge, the first Li Fraumeni syndrome family in China. Six family members were affected with various tumors. A TP53 mutation (G244S) co-segregated with the tumor phenotype within this family. Functional analysis indicated that G244G mutation disrupted the transactivity, DNA-binding and cell growth inhibition activity of p53 proteins. The identification of germline TP53 mutation has important clinical implications for this family. Individuals with a deleterious TP53 germline mutation have an approximately 90% lifetime risk of developing cancer and 50% occurring before the age of 40 years. We have informed the patients (or their parents, if younger than 14 y) with consent for the TP53 mutation. The mutation carriers are suggested to performed regular check according to the National Comprehensive Cancer Network (NCCN) guidelines. The carriers were also strongly recommended to perform pre-implantation genetic diagnosis if they get pregnancy in the future.
Considering the large population in China and the high risk of TP53 germline carrier to develop cancer, the prevalence of germline TP53 mutation in various tumor patients should be surveyed and the possibility to perform TP53 mutation test in high-risk individuals should be considered in China. Indeed, Cao et al. recently identified eight TP53 mutations in non-LFS/non-LFL breast cancer families from China. Five of them are located in introns and three of them are in the protein encoding region (L188P, R72P and S215_Y220del)31.
Methods
Patients
The proband was identified in the Department of Neurosurgery at Xiangya Hospital, Central South University of China. Nine subjects in her family were included in this study. Genomic DNA was extracted from peripheral blood leukocytes and genomic DNA of tumors from the proband and her younger brother was also extracted. The study was approved by the Ethics Committee of Xiangya Hopsptal and signed informed consent was obtained from each of the subjects (or their parents if younger than 14 y). All methods were performed in accordance with approved guidelines.
Exome sequencing
For each sample, 3 μg of genomic DNA was sonicated into fragments and used to construct a paired-end sequencing library with the Agilent SureSelect Target Enrichment System. Exome capture was performed with the Agilent SureSelect Human All Exon kit. Each sample was sequenced on an Illumina HiSeq2000 instrument. The sequenced reads were aligned to the human genome reference (UCSC hg 19 version) using Burrows-Wheeler Aligner (BWA). Reads qualities were recalibrated using Genome Analysis Toolkit (GATK). Picard 1.14 was used to flag duplicate reads. GATK IndelRealigner was used to realign reads around insertion/deletion sites. The Single Nucleotide Variants (SNVs) and small insertions and deletions (InDels) were generated with GATK Unified Genotyper and in parallel with the SAMtools pipeline. The called SNVs and Indels were annotated with ANNOVAR32,33,34,35,36,37.
Plasmids and antibodies
The coding region of human TP53 was amplified using polymerase chain reaction (PCR) and cloned into pcDNA3.1. The p21-luciferase reporter plasmid (p21-Luc) was a kind gift from Professor Peter Kaiser (University of California at Irvine)38. The G244S mutation was engineered using the QuikChange mutagenesis kit (Stratagene, La Jolla, CA) according to the manufacturer’s instructions. All the constructs were confirmed by Sanger sequencing. Antibodies against p53 and p21 were purchased from Cell Signaling Technology.
Immunofluorescence staining
TP53-deficient HCT116 cells (a gift from Dr. Bert Vogelstein) were plated on coverslips and transfected with wild-type (WT) or G244S p53-expressing plasmids. At 36 h after transfection, cells were fixed in 4% paraformaldehyde for 30 min, followed by treatment with 1% Triton X-100 for 1 h at room temperature. Cells were then incubated with blocking solution (PBS, 3% bovine serum albumin, 5% goat serum) for 1 h. Incubation with p53 mouse antibody (1:400 dilution) was performed at 4 °C overnight. After being washed with PBS, fluorescence-labeled secondary anti-mouse antibodies (1:400 dilution; Invitrogen, Carlsbad, CA) were incubated for 2 h at room temperature in a dark room. After incubation with 4,6-diamino-2-phenylindole (DAPI; Invitrogen, Carlsbad, CA) for 5 min, cells were mounted in Fluoromount medium (Sigma, St Louis, WA) and fluorescence images were examined with a laser scanning confocal system installed on a Carl Zeiss microscope (Zeiss, Gottingen, Germany) with a 63× oil immersion objective.
Luciferase reporter assay
TP53-deficient HCT116 cells were transfected with 5 ng of p21-luciferase reporter plasmid and 20 ng of expression constructs for wild-type (WT) or G244S p53. At 48 h after transfection, cells were washed with PBS and lysed with Reporter Lysis Buffer (Promega, Madison, WI). The extracts were assayed for luciferase activity using the Luciferase Assay System (Promega, Madison, WI) according to the manufacturer’s protocol. All reporter assays were conducted at least three times and performed in triplicate on different days using different batches of cells.
Western blot analysis
TP53-deficient HCT116 cells were transfected with 200 ng of WT or G244S p53-expressing plasmids in 12-well plates. At 48 h after transfection, cells were lysed in 2XSDS loading buffer, containing 1 mM phenylmethanesulfonyl fluoride (PMSF) (Sigma, St Louis, WA) and 0.2 mM β-mercaptoethanol supplemented with protease inhibitor cocktail (Sigma, St Louis, WA). The lysate containing the same total protein was separated on 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis, transferred onto a polyvinylidene fluoride membrane and subjected to immunoblotting with p53 mouse antibody and p21 rabbit antibody. β-actin acts as a loading control.
To analyze the stability of WT and G244S p53, cycloheximide (Sigma, St Louis, WA, USA) was added to a final concentration of 100 μg/ml at 48 h after transfection. The cells were lysed at 0, 1, 2, 4, 6, 8 h after cycloheximide treatment and the proteins were detected with Western blot.
Real-time quantitative PCR
50 ng genomic DNA was used for real-time quantitative PCR using 1XSYBR Green PCR master mix (Takara Shuzo Co., Kyoto, Japan). The relative TP53 gene copy number in comparison with the cordon-bleu WH2 repeat protein gene (COBL) was calculated using the comparative cycles of threshold method.
Cell proliferation assay
Human osteosarcoma SaoS-2 cells stably expressing WT or G244S p53 were plated onto 96-well plates and the plates were harvested at day 1, 2, 3, 4 and 5. 20 μl of 5 mg/ml MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) were added to each well and carefully removed without disturbing the cells after incubation for 4 h at 37 °C. 150 μl dimethyl sulfoxide (DMSO) was added into each well and the plate was shaken at room temperature for 15 min. Absorbance was read at 590 nm with a reference filter of 620 nm.
Chromatin immunoprecipitation assay
Chromatin immunoprecipitation (ChIP) assays were performed using Chromatin Immunoprecipitation Assay Kit (Millipore, Bedford, MA, USA) according to the manufacturer’s instructions. Chromatin was sonicated into 200–1000 bp fragments and precipitated with 2 μg of p53 mouse antibody or normal mouse IgG. Purified DNA was subjected to PCR amplification using primers spanning the p53-binding site on p21 promoter (forward primer, 5′-AGCAGGCTGTGGCTCTGATT-3′; reverse primer, 5′-CAAAATAGCCACCAGCCTCTTCT-3′).
Statistical analyses
A repeated measures ANOVA followed by unpaired t test was used to analyze the data for differences. All statistical analyses were performed using Prism 6 (GraphPad Software, San Diego, CA).
Additional Information
How to cite this article: Hu, H. et al. Genetic and functional analysis of a Li Fraumeni syndrome family in China. Sci. Rep. 6, 20221; doi: 10.1038/srep20221 (2016).
References
Li, F. P. & Fraumeni, J. F. Rhabdomyosarcoma in children: epidemiologic study and identification of a familial cancer syndrome. J Natl Cancer Inst 43, 1365–1373 (1969).
Li, F. P. & Fraumeni, J. F. Soft-tissue sarcomas, breast cancer and other neoplasms. A familial syndrome? Ann Intern Med 71, 747–752 (1969).
Li, F. P. et al. A cancer family syndrome in twenty-four kindreds. Cancer Res 48, 5358–5362 (1988).
Masciari, S. et al. Gastric cancer in individuals with Li-Fraumeni syndrome. Genet Med 13, 651–657, 10.1097/GIM.0b013e31821628b6 (2011).
Ruijs, M. W. et al. TP53 germline mutation testing in 180 families suspected of Li-Fraumeni syndrome: mutation detection rate and relative frequency of cancers in different familial phenotypes. J Med Genet 47, 421–428, 10.1136/jmg.2009.073429 (2010).
Wong, P. et al. Prevalence of early onset colorectal cancer in 397 patients with classic Li-Fraumeni syndrome. Gastroenterology 130, 73–79, 10.1053/j.gastro.2005.10.014 (2006).
Nichols, K. E., Malkin, D., Garber, J. E., Fraumeni, J. F. & Li, F. P. Germ-line p53 mutations predispose to a wide spectrum of early-onset cancers. Cancer Epidemiol Biomarkers Prev 10, 83–87 (2001).
Malkin, D. et al. Germ line p53 mutations in a familial syndrome of breast cancer, sarcomas and other neoplasms. Science 250, 1233–1238 (1990).
Chompret, A. et al. P53 germline mutations in childhood cancers and cancer risk for carrier individuals. Br J Cancer 82, 1932–1937, 10.1054/bjoc.2000.1167 (2000).
Nichols, K. E. & Malkin, D. Genotype Versus Phenotype: The Yin and Yang of Germline TP53 Mutations in Li-Fraumeni Syndrome. J Clin Oncol 33, 2331–2333, 10.1200/JCO.2015.61.5757 (2015).
Green, D. R. & Kroemer, G. Cytoplasmic functions of the tumour suppressor p53. Nature 458, 1127–1130, 10.1038/nature07986 (2009).
Riley, T., Sontag, E., Chen, P. & Levine, A. Transcriptional control of human p53-regulated genes. Nat Rev Mol Cell Biol 9, 402–412, 10.1038/nrm2395 (2008).
Sablina, A. A. et al. The antioxidant function of the p53 tumor suppressor. Nat Med 11, 1306–1313, 10.1038/nm1320 (2005).
Vousden, K. H. & Prives, C. Blinded by the Light: The Growing Complexity of p53. Cell 137, 413–431, 10.1016/j.cell.2009.04.037 (2009).
Zhao, S. et al. Regulation of cellular metabolism by protein lysine acetylation. Science 327, 1000–1004, 10.1126/science.1179689 (2010).
Slee, E. A. & Lu, X. Requirement for phosphorylation of P53 at Ser312 in suppression of chemical carcinogenesis. Sci Rep 3, 3105, 10.1038/srep03105 (2013).
Xu, J. et al. RhoGAPs attenuate cell proliferation by direct interaction with p53 tetramerization domain. Cell Rep 3, 1526–1538, 10.1016/j.celrep.2013.04.017 (2013).
Yang, P. M. et al. Zebularine inhibits tumorigenesis and stemness of colorectal cancer via p53-dependent endoplasmic reticulum stress. Sci Rep 3, 3219, 10.1038/srep03219 (2013).
Maddocks, O. D. et al. Serine starvation induces stress and p53-dependent metabolic remodelling in cancer cells. Nature 493, 542–546, 10.1038/nature11743 (2013).
Guo, L. et al. Ionizing radiation induces a dramatic persistence of p53 protein accumulation and DNA damage signaling in mutant p53 zebrafish. Oncogene 32, 4009–4016, 10.1038/onc.2012.409 (2013).
Dittmer, D. et al. Gain of function mutations in p53. Nat Genet 4, 42–46, 10.1038/ng0593-42 (1993).
Lang, G. A. et al. Gain of function of a p53 hot spot mutation in a mouse model of Li-Fraumeni syndrome. Cell 119, 861–872, 10.1016/j.cell.2004.11.006 (2004).
Song, H., Hollstein, M. & Xu, Y. p53 gain-of-function cancer mutants induce genetic instability by inactivating ATM. Nat Cell Biol 9, 573–580, 10.1038/ncb1571 (2007).
Joerger, A. C. & Fersht, A. R. The tumor suppressor p53: from structures to drug discovery. Cold Spring Harb Perspect Biol 2, a000919, 10.1101/cshperspect.a000919 (2010).
Olive, K. P. et al. Mutant p53 gain of function in two mouse models of Li-Fraumeni syndrome. Cell 119, 847–860, 10.1016/j.cell.2004.11.004 (2004).
Liu, D. P., Song, H. & Xu, Y. A common gain of function of p53 cancer mutants in inducing genetic instability. Oncogene 29, 949–956, 10.1038/onc.2009.376 (2010).
Lalloo, F. et al. Prediction of pathogenic mutations in patients with early-onset breast cancer by family history. Lancet 361, 1101–1102, 10.1016/S0140-6736(03)12856-5 (2003).
Choi, M. et al. Genetic diagnosis by whole exome capture and massively parallel DNA sequencing. Proc Natl Acad Sci USA 106, 19096–19101, 10.1073/pnas.0910672106 (2009).
Shlien, A. et al. Excessive genomic DNA copy number variation in the Li-Fraumeni cancer predisposition syndrome. Proc Natl Acad Sci USA 105, 11264–11269, 10.1073/pnas.0802970105 (2008).
Ariffin, H. et al. Whole-genome sequencing analysis of phenotypic heterogeneity and anticipation in Li-Fraumeni cancer predisposition syndrome. Proc Natl Acad Sci USA 111, 15497–15501, 10.1073/pnas.1417322111 (2014).
Cao, A. Y. et al. Identification and characterization of two novel germ line p53 mutations in the non-LFS/non-LFL breast cancer families in Chinese population. Breast Cancer Res Treat 119, 295–303, 10.1007/s10549-009-0349-6 (2010).
DePristo, M. A. et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat Genet 43, 491–498, 10.1038/ng.806 (2011).
Koboldt, D. C. et al. VarScan 2: somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res 22, 568–576, 10.1101/gr.129684.111 (2012).
Li, H. & Durbin, R. Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760 (2009).
Sherry, S. T. et al. dbSNP: the NCBI database of genetic variation. Nucleic Acids Res 29, 308–311 (2001).
Abecasis, G. R. et al. A map of human genome variation from population-scale sequencing. Nature 467, 1061–1073, 10.1038/nature09534 (2010).
Wang, K., Li, M. & Hakonarson, H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res 38, e164, 10.1093/nar/gkq603 (2010).
Wassman, C. D. et al. Computational identification of a transiently open L1/S3 pocket for reactivation of mutant p53. Nat Commun 4, 1407, 10.1038/ncomms2361 (2013).
Acknowledgements
We thank Drs. Peter Kaiser and Bert Vogelstein for kindly providing materials. This work was supported by the National Basic Research Program of China (2012CB517904), the National Natural Science Foundation of China (31371187), the Lotus Scholar Professorship Funds from Hunan Province Government and a research grant provided by the Central South University of China.
Author information
Authors and Affiliations
Contributions
X.L., J.L. and J.-D. L. initiated this project and designed the study; X.L. and J.L. recruited the patients and collected the clinical data; H.H., S.Z., M.L., H.L. and R.L. performed the experiments and analyzed the data; X.L., Z.X., W.L., G.Q. and J.-D.L. analyzed the data; H.H. and J.-D.L. wrote the paper.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Hu, H., Liu, J., Liao, X. et al. Genetic and functional analysis of a Li Fraumeni syndrome family in China. Sci Rep 6, 20221 (2016). https://doi.org/10.1038/srep20221
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep20221
This article is cited by
-
First case of endometrial cancer after yolk sac tumor in a patient with Li-Fraumeni syndrome
BMC Women's Health (2023)
-
Synchronous choroid plexus papilloma and Wilms tumor in a girl, disclosing a Li-Fraumeni syndrome
Hereditary Cancer in Clinical Practice (2021)
