
Abstract
Chronic diabetic wounds represent a major clinical challenge, compounded by persistent inflammation, microbial invasion, and deficient angiogenesis. To address these intertwined pathophysiological features, we developed a copper-ion coordinated andrographolide-loaded hydrogel (ASFH), significantly enhancing andrographolide solubility and promoting wound healing dynamics. In vitro assessments demonstrated superior antimicrobial activity, optimal mechanical strength, self-healing ability, and cytocompatibility. In diabetic mice, ASFH notably accelerated wound closure, stimulated collagen maturation and re-epithelialization, dynamically shifted macrophages toward an anti-inflammatory phenotype, and markedly enhanced angiogenesis. Mechanistic studies integrating network pharmacology, molecular docking, dynamics simulations, and SPR validation pinpointed the Rac1/JNK1/Jun/Fos signaling cascade as a primary mediator of these regenerative effects. This work presents ASFH as a translationally relevant dressing system, simultaneously addressing critical limitations in diabetic wound management through targeted molecular therapeutic intervention.

Similar content being viewed by others
Introduction
Diabetic wounds (DWs)—particularly diabetic foot ulcers—pose severe complications in diabetes mellitus, with global prevalence exceeding 25% among diabetic patients and accounting for >80% of non-traumatic lower-limb amputations1. These recalcitrant wounds arise from a triad of hyperglycemia-driven microvascular dysfunction, chronic inflammation, and peripheral neuropathy, which collectively impair tissue repair mechanisms2,3. Neuropathy-induced insensitivity delays injury detection, enabling minor trauma to escalate into non-healing ulcers4,5,6. While physiological wound healing progresses through four orchestrated phases—hemostasis, inflammatory resolution, proliferative remodeling, and extracellular matrix (ECM) maturation—DWs pathophysiology disrupts this cascade via persistent oxidative stress, dysregulated immune responses, and microbial colonization7.
Hyperglycemia exacerbates impaired angiogenesis and fibroblast senescence, while skewed macrophage polarization (M1/M2 imbalance) sustains pro-inflammatory cytokine storms (e.g., interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α)) and ECM degradation8,9. Current interventions—debridement, hyperbaric oxygen, and moisture-retentive dressings—fail to address the multifactorial etiology, yielding 40% recurrence rates within one year10,11. Emerging biomaterial strategies now prioritize multifunctional platforms capable of synchronizing antimicrobial action, immunomodulation, and angiogenic stimulation12.
Hydrogels, with their ECM-mimetic 3D architectures, tunable porosity, and stimuli-responsive hydration, have emerged as ideal candidates for dynamic wound management13. They facilitate moisture retention, promote cell proliferation, and can be functionalized with bioactive agents for enhanced wound healing14. Gelatin (Gel), a denatured collagen derivative, offers intrinsic cell-adhesive motifs and biocompatibility but suffers from rapid enzymatic degradation and mechanical fragility15. Though Gel’s hydrophilic characteristics help absorb wound exudates and maintain moisture16, its limited adhesion and mechanical strength necessitate modification or combination with other biomaterials17. Carboxymethyl chitosan (CMCS), a polyampholyte with pH-dependent charge states, enhances antimicrobial activity and hydrogel stability through carboxymethylation-driven solubility18. However, suboptimal adhesion and limited functionalization potential necessitate combinatorial strategies with bioactive agents or metal-ion crosslinkers19.
Andrographolide (Andro, C20H30O5), a labdane diterpenoid from Andrographis paniculata (Burm.f.) Nees, exhibits a remarkable pharmacological profile, including antimicrobial, anti-inflammatory, antioxidant, and hypoglycemic activities20,21. Previous studies have demonstrated Andro’s efficacy in attenuating various diabetes-associated complications through modulation of oxidative stress pathways and inflammatory mediators22,23,24. In cutaneous biology, Andro protects keratinocytes and fibroblasts from apoptosis and senescence25,26,27, while effectively ameliorating inflammatory dermatoses through inhibition of NF-κB signaling and pro-inflammatory cytokine production28,29,30. Its favorable physicochemical profile—molecular weight (350.45 Da), hydrolytic stability (melting point 229–232 °C), and moderate lipophilicity—enables dermal penetration, but its poor aqueous solubility (31.74 μg/mL at 25 °C) limits therapeutic delivery.
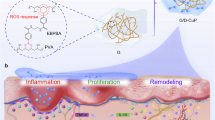
In this study, we developed an andrographolide-integrated self-assembled flexible hydrogel (ASFH) utilizing Cu2+-mediated coordination chemistry between CMCS, gelatin, and Andro. The design leverages dynamic coordination bonds between Cu2+ and functional groups (–COOH in CMCS, –NH2 in Gel, and lactone moieties in Andro) to achieve self-healing rheology, sustained Andro release19,31. Direct Cu2+–Andro coordination was confirmed through Raman spectroscopy, showing the characteristic Cu–O vibrational mode at 585 cm-1, distinguishing true coordination from simple polymer encapsulation. This approach simultaneously enhances Andro solubility while conferring self-healing properties and mechanical adaptability crucial for wound application32,33,34.
We hypothesized that ASFH would accelerate DWs healing by simultaneously addressing three key pathophysiological barriers: microbial burden, chronic inflammation, and impaired angiogenesis. Through comprehensive in vitro and in vivo investigations, we demonstrate that ASFH promotes fibroblast migration, neovascularization, and resolution of inflammation via modulation of the Rac1/JNK1/Jun/Fos signaling axis. This mechanistically driven approach represents a significant advancement in DWs management, offering a cost-effective therapeutic platform with translational potential for this challenging clinical condition.
Results
Physicochemical properties
Raman spectroscopy was employed to analyze the composition of Andro, as well as its mixtures with NaCl and CuCl2. As shown in Fig. 1A, the Raman spectra of pure Andro and the Andro + NaCl mixture exhibited a characteristic signal at 1725 cm−1, which is attributed to the C=O stretching vibration of the lactone ring in Andro. The molecular structure of Andro contains several double bonds, and the characteristic peaks observed at 1650 cm−1 and 1675 cm−1 correspond to the stretching vibrations of the endocyclic and exocyclic C=C bonds, respectively, reflecting vibrational coupling between these groups. Multiple peaks appearing in the region of ~1350–1480 cm−1 (band ①) are primarily associated with the bending vibrations of –CH2 and –CH3 groups in Andro, while those in the ~1000–1300 cm−1 range (band ②) are mainly ascribed to the asymmetric stretching vibrations of C–O–C groups.

A Raman spectra of Andro (black), Andro + NaCl (red), and Andro + CuCl2 (blue). The Andro + CuCl2 trace exhibits a new Cu–O coordination band at ~585 cm-1 and attenuation of the 1725 cm-1 (C=O) and 1675 cm-1 (C=C) features; Andro + NaCl overlaps with pure Andro, confirming the specificity of Cu2+ coordination. B FTIR spectra of Matrix, ASFH-L, and ASFH-H hydrogels showing coordination-induced shifts. C–E Angular frequency scanning curves of matrix, ASFH-L, and ASFH-H hydrogels at 25 °C, showing storage modulus G’ and loss modulus G” as functions of angular frequency. F–H Temperature scanning curves of matrix, ASFH-L, and ASFH-H hydrogels.
Notably, no changes were observed in the Raman spectrum of the Andro + NaCl sample compared to that of pure Andro, indicating that the addition of NaCl did not induce any specific chemical alterations. In contrast, the Andro + CuCl2 sample showed a clear decrease in peak intensities at 1725 cm−1 and 1675 cm−1 relative to the control groups (Andro and Andro + NaCl). This reduction can be explained by the coordination of Cu2+ ions with Andro, which likely weakens the signal intensities of the C=O and C=C bonds. More importantly, a new peak emerged at 585 cm−1 in the Andro + CuCl2 spectrum, which is assigned to the vibrational mode of the Cu–O coordination bond. This result confirms the direct coordination between Cu2+ and andrographolide, rather than a simple physical mixture or encapsulation.
Fourier transform infrared (FTIR) spectroscopy confirmed successful Andro incorporation within the hydrogel matrix (Fig. 1B). While the matrix hydrogel showed no characteristic lactone signals, ASFH-L (low-Andro) and ASFH-H (high-Andro) displayed distinctive absorption peaks at 1636.56 cm−1 and 1635.75 cm−1, respectively, corresponding to the conjugated five-membered lactone ring of Andro. These values are comparable to the previously reported absorption peak of Andro’s five-membered lactone ring at 1646.98 cm−1 35. The slight discrepancy is within acceptable limits, considering that absorption peak positions can vary depending on the compound’s physical state.
Rheological analysis revealed viscoelastic properties suitable for wound applications, with storage modulus (G') consistently exceeding loss modulus (G" across all formulations. Frequency sweep measurements (Fig. 1C–E) demonstrated that ASFH-L exhibited superior structural integrity (G' = 207.6 Pa at 1 rad/s) compared to ASFH-H (G' = 187.9 Pa). The frequency-independent G' plateau observed from 10 to 100 rad/s (Fig. 1D) confirms that ASFH-L behaves as a viscoelastic solid capable of maintaining structural integrity under physiological shear conditions (e.g., joint movement, exudate flow). Temperature-dependent rheology (Fig. 1F–H) confirmed ASFH-L maintained higher mechanical stability at physiological temperature (G' = 160.4 Pa at 37 °C) vs ASFH-H (G' = 114.5 Pa), with corresponding viscosities of 18.1 Pa·s and 13.6 Pa·s, respectively. The thermal stability of G' from 25 to 37 °C ensures consistent mechanical performance under body temperature, which is critical for in vivo depot function and sustained drug release.
The modest loss tangent (tan δ ≈ 0.2–0.3) indicates appropriate energy dissipation that facilitates conformability to irregular wound surfaces without compromising network cohesion. The optimal balance between mechanical strength and viscosity achieved in ASFH-L is critical for withstanding physiological stresses (e.g., tissue movement, exudate flow) while maintaining structural integrity to function as a protective barrier and sustained drug-release depot in the wound environment.
Furthermore, the amplitude-sweep recovery experiments (Fig. 2J) demonstrated that transient high strain—mimicking compression during dressing application or patient movement—does not permanently disrupt the network, enabling the gel to self-repair and maintain both barrier and drug-release functions. Together, these rheological features predict robust in vivo stability: the high G' and low degradation rate (Fig. 2G) enable the gel to resist erosion by wound exudate, the modest tan δ allows conformability to tissue contours, and the self-healing capacity ensures recovery from mechanical insults, thereby sustaining a protective microenvironment and controlled Andro release over the 7–12 day healing window.

A Fluidity assay. B SEM representative micrographs of lyophilized hydrogels; note that ASFH-H displays smaller pores but heterogeneous microstructure due to undissolved Andro particulates, whereas ASFH-L exhibits larger, uniformly interconnected pores; Scale bars: 500 µm (top, 150×), 100 µm (middle, 1000×), 50 µm (bottom, 2000×). Accelerating voltage 5.0 kV, working distance 10.0 mm. C–E Statistical graphs of matrix, ASFH-L, and ASFH-H hydrogels indicating pore size distributions (n = 4). F–G Statistical chart of swelling and degradation data reflects that homogeneity, not pore size alone, governs network stability (n = 3). H Macroscopic bioadhesion assay on porcine skin. ASFH-L adhered to the skin, which was immersed in water for 30 s (simulating wet wound conditions), then subjected to bending and twisting; the gel remained intact, demonstrating adhesion under hydration and mechanical stress. I In vitro drug release profiles showing sustained Andro release. J Consecutive amplitude sweep tests (three cycles with 60 s recovery intervals) demonstrate rheological self-healing via overlapping G' curves. K Macroscopic self-healing visualization by dye-labeled rejoining. Note: all bioadhesion (H) and self-healing (J, K) experiments used the ASFH-L formulation.
Interestingly, we observed that ASFH-H resulted in somewhat weaker viscoelastic behavior at room temperature, likely attributable to altered polymer chain mobility and looser network densities caused by the heterogeneous microstructure resulting from excess undissolved Andro particulates36.
Flow behavior assessment (Fig. 2A and Supplementary Video 1) corroborated rheological findings, with both Andro-containing formulations demonstrating reduced flow compared to the matrix hydrogel. Scanning electron microscope (SEM) micrographs (Fig. 2B) revealed an interconnected porous architecture with ASFH-L exhibiting optimized pore morphology (160.1 ± 60.3 μm) compared to matrix and ASFH-H variants (Fig. 2C–E), facilitating nutrient diffusion and cellular infiltration while maintaining structural integrity. Specifically, ASFH-L demonstrated larger and more regular pore structures beneficial for fluid absorption and exudate management, important considerations for DWs characterized by elevated exudation levels.
Functional characterization demonstrated controlled swelling behavior (Fig. 2F) across all formulations (<125%). Interestingly, while SEM analysis revealed ASFH-H possessed a comparatively smaller average pore size than ASFH-L, it exhibited a higher degradation rate and lower mechanical strength (Fig. 2G). This apparent contradiction can be explained by the heterogeneous microstructure of ASFH-H. The incorporation of Andro beyond its solubility limit (0.2 g in ASFH-H vs 0.05 g in ASFH-L) led to the formation of insoluble particulate domains. These domains, while potentially reducing average pore size, act as structural defects that compromise the hydrogel’s integrity. Consequently, the matrix becomes more susceptible to enzymatic penetration and chain scission, leading to accelerated degradation and diminished mechanical properties despite the smaller pore size. In contrast, ASFH-L, with its fully dissolved and molecularly integrated Andro, forms a more homogeneous and coherent network, resulting in superior structural stability as reflected in its lower degradation rate (74.2 ± 2.4% mass retention for ASFH-H vs a higher value for ASFH-L) and more restrained swelling (113.8 ± 9.0% for ASFH-H).
The ex vivo bioadhesion was designed to mimic the hydrated environment of a wound bed. Immersion in water for 30 s challenged the gel–tissue interface with a burst of hydration representative of exudate conditions. ASFH-L maintained robust adhesion to porcine skin through multiple mechanical cycles (Fig. 2H and Supplementary Video 2), indicating that the Cu2+-mediated coordination network forms sufficiently strong interfacial interactions with tissue proteins (collagen, elastin) to withstand both hydration and motion, key requirements for a functional wound dressing.
Both ASFH-L and ASFH-H exhibited sustained Andro release over 12 h under sink conditions (Fig. 2I). Notably, ASFH-H did not display a higher early cumulative release fraction than ASFH-L, indicating that the bioavailable Andro is governed primarily by the dissolved/complexed fraction rather than by excess undissolved particulates. These findings align with the physical-state rationale and in vivo efficacy plateau for ASFH-H vs ASFH-L.
While Andro is the primary therapeutic agent, the role of the Cu2+ cross-linker warrants consideration. The sustained release of Andro was confirmed experimentally. For Cu2+, a theoretical maximum release calculation indicates that the concentration in our highest-dose cellular extract (2.0 mg/mL hydrogel-equivalent) would be approximately 4.8 nM. This level is one to two orders of magnitude below the typical background copper concentration in standard cell culture media (0.1–1 µM, per USP < 1043 >). As this released amount is negligible and falls within the safe physiological range, the observed in vitro bioactivity can be confidently attributed to the effects of Andro, with Cu2+ functioning primarily as a structural cross-linker to control the material’s properties and Andro’s release kinetics. This dual functionality—Cu2+ as both a structural scaffold and a regulator of Andro bioavailability—underpins the rational design of the ASFH system.
The self-healing property of the ASFH-L hydrogel was evaluated through rheological measurements and macroscopic observation. Consecutive amplitude sweep tests (Fig. 2J) directly assessed self-healing rheology: ASFH-L exhibited high storage modulus (G' > G") in the low-strain linear viscoelastic region, indicating solid-like behavior. Upon increasing strain amplitude beyond ~10%, G' decreased sharply, reflecting network breakdown and transition to a fluid-like state (G" > G'). Critically, after each destructive cycle, G' recovered substantially during the 60 s rest interval at 0.1% strain. The overlapping G' curves across three consecutive cycles demonstrate reproducible mechanical recovery, with recovery efficiencies of ≥85% relative to the initial G'. The slight variations between scans are attributed to the progressively enhanced reformation of the dynamic cross-linked network during the short recovery intervals.
This rheological behavior confirms that the dynamic Cu2+-coordination bonds enable reversible crosslinking and self-healing, which was further corroborated by the macroscopic dye-tracking experiment (Fig. 2K and Supplementary Video 3), where a completely severed hydrogel rapidly re-integrated and restored its structural integrity within 120 s. This effective self-healing capability ensures that the hydrogel can maintain a continuous barrier, providing reliable physical protection in dynamic wound environments.
In conclusion, ASFH-L’s pore diameter, swelling kinetics, controlled degradation, and self-healing capacity collectively provided superior characteristics suited for dynamic DW environments.
Cytocompatibility and migration enhancement
Cell viability assessment using CCK-8 assays demonstrated dose-dependent cytocompatibility in both keratinocytes (HaCaT) and fibroblasts (HFF-1) (Fig. 3A, B). At concentrations between 0.25 and 1 mg/mL, all hydrogel extracts maintained >95% viability, with ASFH-L significantly enhancing cellular proliferation compared to matrix controls at 6–24 h (P < 0.01). Notably, HaCaT exhibited moderate cytotoxicity at 2 mg/mL (94.61 ± 1.66% viability, P < 0.001), establishing 1 mg/mL as the optimal concentration for subsequent experiments.

A, B Viability of HaCaT keratinocytes (A) and HFF-1 fibroblasts (B) exposed to hydrogel extracts for 6-24 h (n = 3). C, D Representative graphs of cell scratch assay of hydrogels acting on glycosylated HaCaT cells and HFF-1 cells. Scale Bar: 50 μm. E, F Statistical graphs of migration rates of glycosylated HaCaT cells and HFF-1 cells (n = 6). *P < 0.05, **P < 0.01, ***P < 0.001, vs Control group, #P < 0.05, ##P < 0.01, ###P < 0.001, vs Matrix group. G Representative inhibition zones graphs of hydrogel against E. coli and S. aureus (n = 3). Scale Bar: 1 cm. ***P < 0.001, vs Blank group, #P < 0.05, vs Matrix group. H, I Quantification of bacteriostatic activity against E. coli (H) and S. aureus (I) by plate colony counting assay (n = 3). J, K Quantification of inhibition zone diameters for E. coli (J) and S. aureus (K) (n = 3). The bar charts present mean ± SD. ***P < 0.001, vs Matrix group, ##P < 0.01, ###P < 0.001, vs Matrix group.
In hyperglycemic conditions (40 mM glucose) that recapitulate DW microenvironments37, scratch wound assays revealed significant migration enhancement by all hydrogel formulations (P < 0.01 vs high glucose (HG) control) (Fig. 3C, D). It is important to note that the 40 mM glucose condition employed in these assays does not stimulate cell migration or proliferation; rather, HG is well-documented to inhibit both processes. Multiple studies have demonstrated that hyperglycemia impairs endothelial and fibroblast migration through mechanisms including increased oxidative stress, mitochondrial dysfunction, impaired VEGF and growth factor signaling, and activation of apoptotic pathways38,39,40. The use of 0.5% FBS further minimized proliferative contributions to wound closure. Therefore, the observed enhancement of migration by ASFH treatments reflects genuine pro-migratory effects that counteract the inhibitory hyperglycemic environment, rather than indirect effects via increased proliferation. ASFH-L demonstrated superior efficacy, accelerating closure by an additional 10.9% compared to matrix hydrogel at 24 h in both cell types (Fig. 3E, F), suggesting Andro directly counteracts hyperglycemia-induced migratory impairment.
Antimicrobial efficacy
Given the polymicrobial colonization prevalent in DWs, we evaluated antibacterial activity against representative gram-positive (S. aureus) and gram-negative (E. coli) pathogens (Fig. 3G).
Colony formation assays with standardized inocula (103 CFU/mL) demonstrated significant bacterial inhibition by all hydrogel formulations, with matrix hydrogel substantially reducing viable counts compared to untreated controls (P < 0.001). ASFH variants exhibited dose-dependent enhancement of antimicrobial activity, with both ASFH-L and ASFH-H significantly outperforming the matrix formulation (P < 0.05) (Fig. 3H, I).
Zone of inhibition assays corroborated these findings, with ASFH-H generating significantly larger inhibition zones against S. aureus (20.3 ± 0.2 mm) and E. coli (17.1 ± 0.2 mm) compared to ASFH-L (P < 0.01, Fig. 3J, K). This concentration-dependent antimicrobial effect highlights the synergistic activity between Andro’s bioactive properties and the intrinsic antibacterial characteristics of the Cu2+-CMCS matrix, providing broad-spectrum protection against common wound pathogens.
Network pharmacology elucidates Andro’s mechanism in DWs
Integration of PharmMapper-predicted Andro targets (289 genes) with DWs-associated genes from GeneCards/OMIM (3219 genes) revealed 163 overlapping candidates (Fig. 4A). Protein–protein interaction (PPI) network construction and topology analysis (Fig. 4B) identified critical nodes including MAPK8 (JNK1; degree = 42), GSTP1 (degree = 38), SRC (degree = 36), AKT1 (degree = 34), HSP90AA1 (degree = 32), and JUN (degree = 30). MAPK8 exhibited the highest betweenness centrality (Table S2), indicating its pivotal role in mediating Andro’s effects on DW healing.

A Venny plots of Andro target genes vs DWs causative genes. B PPI network highlighting key signaling proteins modulated by Andro treatment. Nodes represent individual proteins (colored by pathway/function; see legend); edges represent validated interactions. Major pathway clusters are annotated (e.g., JNK1/MAPK, Rac1/cell migration). C GO functional enrichment analysis of intersecting targets, showing significantly enriched biological processes (e.g., inflammatory response, cell migration, angiogenesis) ranked by gene count or significance. D KEGG pathway enrichment analysis ranked by gene count and significance. E Top 20 KEGG pathway enrichment analysis (sorted by enrichment score). F Representative docking pose of Andro within the proteins binding pocket, showing key noncovalent interactions.
Gene Ontology (GO) enrichment analysis revealed that Andro likely modulates biological processes critical for wound healing, including MAPK cascade regulation, cell migration, protein phosphorylation, and receptor tyrosine kinase signaling (Fig. 4C). Molecular function analysis highlighted protein kinase activity, MAP kinase activity, and transcription factor binding. KEGG pathway analysis identified significant enrichment in VEGF signaling, MAPK signaling, PI3K-Akt signaling, and Ras signaling pathways (Fig. 4D, E and Table S3). These findings directed our focus to the Rac1/JNK1/Jun/Fos signaling axis as a putative mechanism for Andro’s pro-healing effects.
Molecular docking validates Andro-target interactions
Docking simulations predicted high-affinity binding between Andro and Rac1 mediated by hydrogen bonds at Glu31 and Leu160 residues and VDW interactions with Tyr32, Phe28, and Cys18 (Fig. 4F). All tested targets (Rac1, JNK1, Jun, Fos) showed binding energies below −5.5 kcal/mol (Table 1), confirming stable ligand-receptor interactions.
Molecular dynamics (MD) simulations reveal dynamic binding behavior
To investigate the intracellular binding mechanisms of Andro to Rac1, JNK1, Jun, and Fos—independent of Cu2+-mediated solubilization in the hydrogel—we performed molecular docking followed by 100 ns all-atom MD simulations in GROMACS 2022. Building on our prior MD work in protein–DNA41, protein–protein42, and protein–drug43 interactions, we modeled copper-free aqueous complexes to reflect post-release cellular conditions and to assess binding stability and conformational dynamics.
Focusing on the Andro–Rac1 complex, the root mean square deviation (RMSD)—a metric quantifying the average displacement of atoms relative to a reference structure—revealed a transient destabilization event at approximately 50 ns (Fig. 5A). Following this transient, the RMSD stabilized and fluctuated within a narrow range (~0.3–0.4 nm), indicating that the system had reached equilibrium. Concurrent fluctuations were observed in the radius of gyration (Rg), which reflects the overall compactness of the complex (Fig. 5B), and in the distance between the centers of mass of Rac1 and Andro (Fig. 5C); both metrics remained stable throughout the 60–100 ns window, further confirming structural equilibration. The free energy landscape (FEL), constructed to visualize the conformational stability and predominant low-energy states, confirmed that the system predominantly sampled a single low-energy basin (indicated by the dark blue region in Fig. 5D), a hallmark of a stable, equilibrated bound state. Further analysis of the buried solvent-accessible surface area (SASA)—an indicator of interface stability and interaction strength—showed a reduction near 50 ns, consistent with the transient weakening of binding interactions during structural rearrangement (Fig. 5E), but recovered and stabilized thereafter.

A RMSD of complex, protein, and Ligand, reflecting structural deviation and conformational stability over time. B Rg of the complex, indicating overall compactness and structural tightness. C Rac1 and Andro binding site distance, monitoring binding stability and positional consistency. D FEL showing a single dominant basin, indicating conformational stability without large-scale reorganization. E Buried area between Andro and Rac1, representing the extent of buried surface and interaction strength. F Simulated conformational superposition, illustrating structural flexibility and binding site variations. G Modeling trajectory change, displaying dynamic shifts in binding site and molecular orientation. H PCA and protein structure, highlighting dominant motion modes and conformational flexibility. I Andro and Rac1 binding energies VDW and ELE, showing contributions from van der Waals and electrostatic interactions. J Hydrogen bond number, reflecting the stability and strength of polar interactions. K Hydrogen bond frequency between Andro and Rac1, indicating persistence and key residues involved in H-bond formation.
Trajectory superposition and snapshot analysis (Fig. 5F, G) identified three distinct binding pockets on Rac1, between which Andro migrated in response to local conformational dynamics. This binding-site hopping is consistent with the RMSD transient at 50 ns and reflects the flexible nature of the Rac1 surface rather than dissociation. Principal component analysis (PCA), employed to reduce the dimensionality of conformational sampling and highlight essential motions, indicated that Andro itself maintained considerable conformational stability throughout the simulation (Fig. 5H), suggesting that Andro adapts its binding mode without large intramolecular distortions. The electrostatic (ELE) and van der Waals (VDW) interaction energies exhibited initial variability, consistent with binding-site adjustment, before stabilizing in the latter half of the simulation at approximately –150 kJ/mol (VDW) and –50 kJ/mol (ELE) (Fig. 5I), demonstrating strong and stable non-covalent interactions. Hydrogen bond analysis revealed intermittent formation, with the number of simultaneous hydrogen bonds ranging from 0 to 2 (Fig. 5J, K), indicating moderate but not dominant contributions from polar interactions; the binding is primarily driven by hydrophobic and electrostatic complementarity.
Molecular Mechanics Poisson-Boltzmann Surface Area (MM-PBSA) binding free energy calculations (Table 2) demonstrated that VDW interactions (ΔEvdw = –167.5 kJ/mol) constituted the dominant favorable contribution to binding, supplemented by electrostatic (ΔEele = –45.9 kJ/mol) and non-polar solvation (ΔEnonpol = –19.3 kJ/mol) terms. The net binding energy (ΔEMM-PBSA) of –35.7 ± 4.2 kJ/mol signifies a stable and pharmacologically relevant binding interaction. In summary, while Andro undergoes transient positional shifts on the Rac1 surface, it ultimately forms a stable complex driven primarily by VDW and electrostatic forces, providing a structural basis for Rac1 activation in DW healing.
ASFH accelerates wound healing in DW mice
To assess ASFH’s wound-healing capacity, a full-thickness excisional model was implemented in diabetic mice, and wound area, as well as perfusion, were monitored (Fig. 6A, B, D). By Day 3 post-injury, matrix, ASFH-L, and ASFH-H treatments each significantly facilitated wound closure compared with Controls (P < 0.001). Notably, ASFH-L surpassed matrix hydrogel by ~13% at Days 3 and 7 (P < 0.001), whereas ASFH-H and matrix treatments showed no significant difference at these early time points (Fig. 6C).

A Schematic timeline of the in vivo wound healing experiment. Full-thickness skin wounds with a diameter of 0.8 cm were created on both sides of the dorsal midline. The day of surgery was designated as day 0. Treatments were initiated from day 1 onwards. The schematic indicates the time points for RFLSI monitoring, photographic documentation, and sample collection throughout the 12-day study period. B Representative images of wounds in different treatment groups on days 0, 3, 7, and 12. Grid marks at the bottom indicate 1 mm divisions. C Wound closure rates for each group over 12 days. The graph visually represents changes in wound area over time. Scale Bar: 2 mm. D Wound healing rate. E Representative RFLSI perfusion maps of wound regions at days 3, 7, and 10, with red and blue colors indicating high and low blood flow, respectively. Scale Bar: 5 mm. F Quantitative analysis of perfusion signal intensity within the wound area over time. The bar charts present mean ± SD (n = 6 per group). *P < 0.05, **P < 0.01, ***P < 0.001, vs Control group, #P < 0.05, ###P < 0.001, vs Matrix group.
Regional perfusion, measured by RFLSI (Fig. 6D), was likewise improved in ASFH-L–treated wounds relative to matrix at Days 3 and 7 (P < 0.05). By Day 10, ASFH-L treatment remained significantly superior to the Control group in perfusion enhancement (1394.49 ± 162.84 Units, P < 0.05). (Fig. 6E). Collectively, these findings highlight ASFH-L as the most effective formulation for DW healing in vivo.
ASFH enhances wound microarchitecture and tissue regeneration
Epithelialization and ECM remodeling
Histological assessments revealed that ASFH significantly improved wound architecture. H&E staining demonstrated increased hair follicle density in ASFH-L and ASFH-H groups compared to matrix controls (Fig. 7A, B), indicating enhanced epithelial stem cell activation. By day 7, ASFH-L-treated wounds exhibited complete re-epithelialization with stratified epidermis and organized dermal architecture.

A–C Representative graphs and statistics of H&E and Masson’s trichrome staining of wound tissues at days 3 and 7 post-injury for each treatment group, with corresponding quantitative analyses (n = 6). H&E images highlight epidermal thickness, granulation tissue formation, and inflammatory cell infiltration. Note: Blue arrows represent hair follicles, red arrows represent sebaceous glands, black arrows represent lymphatic vessels, and green arrows represent collagen fibers. Masson’s trichrome staining specifically highlights collagen fibers in blue. Scale Bar: 50 μm. D–F Immunohistochemical analysis and quantification of collagen deposition. Representative images (D) and statistical data show the expression of Col I (E) and Col III (F) in wound tissues at days 3 and 7 post-wounding (n = 3). Brown deposits indicate positive immunoreactivity for the respective collagen types. Scale bar: 50 μm. The bar charts present mean ± SD. *P < 0.05, ***P < 0.001, vs Control group, #P < 0.05, ##P < 0.01, ###P < 0.001, vs Matrix group.
Masson’s trichrome staining quantified accelerated collagen deposition in ASFH-treated wounds (Fig. 7A, C). By day 3, ASFH-L increased collagen content to 41.50 ± 2.21% vs 24.37 ± 3.40% in controls (P < 0.001). This advantage persisted at day 7, where ASFH-L significantly outperformed both control and matrix hydrogel groups (P < 0.01).
Immunohistochemical analysis of collagen subtypes44 revealed that ASFH-L selectively enhanced type I collagen (mature collagen) expression at both timepoints (P < 0.05 vs control) (Fig. 7D, E). Type III collagen (immature, pro-regenerative) showed increased deposition at day 3 in ASFH-L-treated wounds (0.171 ± 0.010 vs 0.124 ± 0.007 intensity units in controls, P < 0.05), with sustained elevation through day 7 (0.187 ± 0.006 vs 0.149 ± 0.005 intensity units in matrix group, P < 0.01) (Fig. 7D, F). This collagen profile indicates progression from provisional matrix formation to mature scar organization, essential for tensile strength recovery.
Inflammatory resolution
Immunofluorescence staining for F4/80, CD86, and CD206 was performed on skin samples collected at days 3 and 7 (Fig. 8A, B, E, F). At day 3, ASFH-L significantly reduced pro-inflammatory M1 macrophages (CD86⁺, 47.12 ± 5.13% vs 72.85 ± 11.53% in matrix group, P < 0.001) while increasing pro-resolving M2 macrophages (CD206⁺, 248.59 ± 25.96% vs 178.12 ± 10.94% in matrix group, P < 0.001) (Fig. 8C, D). This immunomodulatory effect was further enhanced by day 7, during which ASFH-L treatment significantly reduced M1 macrophage prevalence (32.98 ± 2.50% vs 79.28 ± 6.78%, P < 0.001). Meanwhile, M2 macrophage levels in ASFH-L-treated wounds increased to 240.13 ± 30.35% of baseline, nearly doubling those observed in matrix controls (121.91 ± 21.64%, P < 0.001) (Fig. 8G, H). This dual action—suppressing the M1 phenotype while actively promoting the M2 phenotype—provides strong evidence for a true M1‑to‑M2 polarization switch, rather than just a passive reduction of inflammation. Mediated by HG, the pro-inflammatory factors IL-1β and TNF-α increase glucose consumption and mitochondrial superoxide production, causing cellular inflammation and oxidative damage45. Gene expression analysis corroborated these findings, showing that ASFH-L and ASFH-H significantly downregulated pro-inflammatory cytokines Il1b and Tnf compared to matrix controls at days 3 and 7 (Fig. 8I–L).

A, B, E, F Representative immunofluorescence images of wound sections at days 3 (A, B) and 7 (E, F) stained for macrophage markers. F4/80 (green) labels total macrophages, CD86 (red) marks pro-inflammatory M1 macrophages, and CD206 (red) marks pro-resolving M2 macrophages; nuclei are counterstained with DAPI (blue). Scale bars: 50 µm. C, D, G, H Quantification of CD86⁺ (M1) and CD206⁺ (M2) macrophages at days 3 and 7, demonstrating a coordinated decrease in M1 and increase in M2 markers, indicative of a phenotypic switch. Mean ± SD (n = 6). I–L Relative mRNA expression levels of pro-inflammatory cytokines Il1b and Tnf in wound tissues at days 3 and 7 (n = 3). Data are mean ± SD. **P < 0.01, ***P < 0.001, vs Control group, ##P < 0.01, ###P < 0.001, vs Matrix group.
Furthermore, Treatment with the ASFH-L hydrogel reversed the pro-inflammatory phenotype induced by lipopolysaccharide (LPS) in RAW264.7 macrophages. The hydrogel significantly reduced the surface expression of the M1 marker CD86 (mean fluorescence intensity [MFI] 37.44 ± 1.90 vs 58.24 ± 1.58 for the LPS group, P < 0.01) while simultaneously increasing the expression of the M2 marker CD206 (MFI 12.36 ± 2.03 vs 7.09 ± 0.77 for the LPS group, P < 0.01) (Fig. 9A–D).

A, B Representative immunofluorescence images of CD86 (A, M1 marker) and CD206 (B, M2 marker) in RAW264.7 cells under different treatments: untreated control, LPS, matrix hydrogel extract (Matrix), and Andro‑loaded hydrogel extracts (ASFH‑L, ASFH‑H). Staining shows CD86/CD206 (green) and DAPI (blue). Scale bar: 100 μm. C, D Quantification of CD86 (C) and CD206 (D) fluorescence intensities, normalized to the LPS group. E–G ELISA quantification of IL‑6 (E), TNF‑α (F), and IL‑10 (G) in culture supernatants from LPS‑primed RAW264.7 cells treated with the indicated hydrogel extracts. Data in (C–G) are presented as mean ± SD (n = 3). *P < 0.05, **P < 0.01, ***P < 0.001, vs LPS group, #P < 0.05, ##P < 0.01, vs Matrix group.
Consistent with this polarization shift, the ASFH-L hydrogel modulated cytokine secretion. Compared to the LPS-treated group, the hydrogel reduced the release of the pro-inflammatory cytokines IL-6 (0.77-fold) and TNF-α (0.83-fold) and enhanced the secretion of the anti-inflammatory cytokine IL-10 (1.46-fold) (Fig. 9E–G).
This inflammation-resolution profile is particularly relevant for DW management, where chronic inflammation perpetuates tissue damage and impairs healing46,47. The observed shift from M1 to M2 macrophage polarization, characterized by reduced pro-inflammatory cytokine production and enhanced anti-inflammatory mediator release, recapitulates the physiological transition from the inflammatory to proliferative phase of wound repair. These findings indicate that ASFH can effectively dampen the exaggerated inflammatory response characteristic of DW, promoting a transition from the pro-inflammatory to the proliferative phase of healing.
Vascular regeneration
Microvascular restoration, critical for nutrient delivery in DW, was assessed through CD31 and α-SMA immunofluorescence. ASFH-L significantly enhanced CD31⁺ vessel density at days 3 and 7 (163.81 ± 16.02 and 157.02 ± 14.42 intensity/μm2, respectively, P < 0.01 vs matrix) (Fig. 10A, B). Pericyte recruitment, essential for vessel maturation, was evaluated through α-SMA expression, which reached 171.28 ± 14.31 intensity/μm2 in ASFH-L-treated wounds by day 7 (P < 0.05 vs controls) (Fig. 10C, D). Supporting these vascular changes, ASFH upregulated Egfr expression at both timepoints (P < 0.05) (Fig. 10E, F), a key mediator of keratinocyte-fibroblast crosstalk during angiogenesis.

A, B Immunofluorescence staining and quantification of CD31-positive blood vessels in wound sections at days 3 and 7 (n = 6). Tissue sections were stained for CD31 (red, marking vascular endothelial cells) and DAPI (blue, marking nuclei). Scale bar: 100 μm. C, D Immunofluorescence analysis and quantification of α-SMA expression in wound tissues at days 3 and 7 (n = 6). α-SMA (red) marks myofibroblasts and vascular smooth muscle cells, reflecting wound contraction and vessel maturation; DAPI (blue) indicates nuclei. Scale bar: 100 μm. E, F Relative mRNA expression levels of Egfr in wound tissues at days 3 and 7 (n = 3). The bar charts present mean ± SD. **P < 0.01, ***P < 0.001, vs Control group, #P < 0.05, ##P < 0.01, ###P < 0.001, vs Matrix group.
ASFH Activates the Rac1/JNK1/Jun/Fos signaling axis
Rac1 activation plays a critical role in tissue repair through activation of JNK1 and downstream transcription factors Jun and Fos, collectively influencing cell proliferation, migration, and angiogenesis48,49. Molecular pathway analyses revealed that ASFH modulates the Rac1/JNK1/Jun/Fos signaling cascade. Quantitative reverse transcription polymerase chain reaction (RT-qPCR) demonstrated that ASFH-L significantly upregulated Rac1 (1.54-fold), Jnk1 (1.35-fold), and Fos (1.11-fold) transcripts by day 3 compared to matrix controls (P < 0.05) (Fig. 11C). Surface plasmon resonance (SPR) assays quantified moderate binding affinity between Andro and Rac1 (KD = 3.82 × 10−6 M, Fig. 11D), aligning with computational predictions (Figs. 4F and 5). ASFH-H selectively enhanced Jun expression (1.54-fold, P < 0.05). This coordinated transcriptional response persisted through day 7, with both ASFH formulations maintaining elevated pathway component expression (Fig. 11E).

A, B Representative western blots (A) and quantification (B) of Rac1, JNK1, Jun, and Fos protein levels in wound tissues at day 3 for each treatment group. C Relative mRNA expression levels of Rac1, Jnk1, Jun, and Fos in day-3 DW tissues. D SPR analysis confirming direct binding of Andro to Rac1 with a dissociation constant (KD) of 3.82 µM. E Relative mRNA expression levels of Rac1, Jnk1, Jun, and Fos in day-7 wound tissues. F, G Representative western blots (F) and quantification (G) of Rac1, JNK1, Jun, and Fos protein levels in day-7 wound tissues. The bar charts present mean ± SD (n = 3 per group). *P < 0.05, **P < 0.01, ***P < 0.001, vs Control group, #P < 0.05, ##P < 0.01, ###P < 0.001, vs Matrix group. H Schematic model summarizing the proposed mechanism by which ASFH-delivered Andro treats DWs. Andro directly binds Rac1, activates the Rac1/JNK1/Jun/Fos signaling axis, promotes M1-to-M2 macrophage polarization, modulates cytokine profiles, enhances angiogenesis and collagen remodeling, and collectively accelerates wound closure.
Western blot validation confirmed these findings at the protein level. By day 3, ASFH-L significantly increased Rac1 (1.59-fold), JNK1 (1.29-fold), Jun (1.34-fold), and Fos (1.48-fold) protein abundance compared to matrix controls (P < 0.05) (Fig. 11A, B). ASFH-H showed comparable but less pronounced effects. By day 7, ASFH-H demonstrated robust upregulation of Rac1 (2.73-fold), JNK1 (1.52-fold), Jun (1.21-fold), and Fos (1.32-fold) (P < 0.05) (Fig. 11F, G). Collectively, these findings establish that ASFH accelerates DW healing through coordinated activation of the Rac1/JNK1/Jun/Fos signaling axis, culminating in enhanced cellular migration, inflammation resolution, and vascular restoration.
Discussion
DWs remain a substantial clinical challenge, owing to their multifaceted pathological environment, which includes persistent inflammation, chronic microbial colonization, and impaired angiogenesis driven by prolonged hyperglycemia. Andro demonstrable efficacy in mitigating cutaneous inflammation and oxidative stress26,27, its limited aqueous solubility (<0.1 mg/mL) hindered prior translational efforts. In the present study, we addressed these limitations by engineering a hydrogel (ASFH-L) utilizing copper-ion (Cu2+)-mediated coordination chemistry. This strategy significantly enhanced Andro solubility (to approximately 2.5 mg/mL), improving dermal bioavailability and efficacy. Beyond chemical and biological functionality, the wet-adhesion performance of ASFH-L addresses a critical clinical challenge in DWs management. DWs are characteristically exudate-rich, and conventional dressings frequently fail under persistent moisture, leading to dressing displacement, loss of barrier function, and increased risk of secondary infection. The Cu2+-coordinated network architecture of ASFH-L maintains robust interfacial bonding with tissue even after repeated hydration challenges, as demonstrated by our immersion-based bioadhesion assay. This moisture-resistant adhesion ensures continuous contact between the therapeutic hydrogel and the wound bed, sustaining localized Andro delivery and mechanical protection throughout the healing process, thereby reducing the frequency of dressing changes and minimizing trauma to fragile regenerating tissue.
Our findings establish that exceeding the drug coordination/solubility threshold compromises network homogeneity and functional integrity, despite superficially reducing pore size. This principle has translational implications: formulation optimization should target full molecular integration of the active pharmaceutical ingredient within the coordination network, rather than maximizing nominal loading, to ensure robust mechanical properties, controlled degradation, and sustained bioavailability.
Our network pharmacology screen identified hub proteins with high centrality and established clinical druggability—including MAPK8/JNK1, AKT1, SRC, and HSP90AA1—with five of six top hubs possessing approved drugs or clinical candidates. Critically, the predicted MAPK, inflammatory regulation, and wound-repair pathways showed strong concordance with our experimental outcomes: Rac1/JNK1 activation in both in vitro and in vivo assays (Figs. 5, 11), M1-to-M2 macrophage polarization with reduced IL-6/TNF-α and elevated IL-10 (Figs. 8 and 9), enhanced angiogenesis (Figs. 6, 10), and accelerated tissue closure with improved collagen architecture (Figs. 6 and 7). Notably, the robust M1-to-M2 shift—evidenced by simultaneous CD86 downregulation and CD206 upregulation alongside a near-doubling of the M2/M1 ratio—indicates that ASFH actively reprograms macrophage phenotypes rather than passively attenuating inflammation. This bidirectional polarization likely contributes to the accelerated transition from inflammatory to proliferative healing phases, representing a key mechanistic link between Andro-mediated Rac1/JNK1 activation and the resolution of chronic wound inflammation.
This pathway-to-phenotype agreement validates the network pharmacology approach for natural product mechanism elucidation and establishes the Rac1/JNK1 axis—confirmed through orthogonal computational (MD) and experimental (SPR, cellular signaling) methods—as a key mechanistic convergence point linking Andro–target binding to downstream wound-healing responses. Notably, our SPR analysis provided the first quantitative biophysical measurement, to our knowledge, of direct Andro–Rac1 binding affinity (KD = 3.82 µM). While published SPR-derived KD values for other Rac1 inhibitors remain scarce in the literature, this moderate micromolar affinity is characteristic of natural product modulators and validates Rac1 as a direct molecular target of Andro rather than an indirect downstream effector. PCA and FEL calculations revealed that Andro binding stabilizes a single dominant conformational basin in Rac1 with localized interfacial plasticity, suggesting an interface-stabilization or allosteric mechanism rather than wholesale conformational switching. The distinct hydrogen-bonding fingerprint of Andro relative to synthetic inhibitors (e.g., NSC23766) reflects its unique lactone-diol chemotype and underscores the value of natural product scaffolds in expanding the chemical space for Rac1 modulation. These multi-target effects contrast with single-agent therapies and rationalize Andro’s broad efficacy across the complex DWs microenvironment.
Practically, our dose-escalation analysis demonstrates that exceeding Andro’s coordination capacity (ASFH-H) yields no additional therapeutic benefit while compromising network integrity, positioning ASFH-L as the optimized, resource-efficient formulation for clinical translation. The druggable hub architecture suggests promising combinatorial strategies: ASFH may synergize with selective kinase inhibitors (e.g., low-dose AKT inhibitors) or HSP90 modulators to amplify regenerative responses while minimizing systemic toxicity—a hypothesis testable in future preclinical models. Immediate translational priorities include prospective randomized controlled trials in human diabetic foot ulcers, manufacturing scale-up under aseptic conditions to enable direct-contact applications (currently limited by sterilization-induced structural alterations), and pharmacokinetic profiling of sustained Andro release in relation to wound biomarkers. Beyond Andro, this coordination-driven solubilization platform holds potential for delivering diverse hydrophobic wound therapeutics (e.g., curcumin, resveratrol analogs) and warrants systematic structure–activity relationship studies to optimize Cu2+-ligand pairing for maximal network stability and bioactivity.
Methods
Preparation of hydrogels
ASFH was synthesized using a one-step self-assembly method. Briefly, 20 mL of deionized water was added to a beaker and heated with stirring at 450 rpm. Subsequently, 0.5 g of Gel, 0.05 g or 0.2 g of Andro, and 12.5 mg of copper nitrate trihydrate were dissolved in the mixture. Once fully dissolved and heated to 80 °C, the stirring speed was increased to 1000 rpm. Then, 0.5 g of CMCS was rapidly added and stirred thoroughly for 15 min. The resultant mixture was dispensed and subjected to three freeze-thaw cycles to obtain the ASFH. A matrix hydrogel without Andro was prepared using identical steps. Hydrogels containing 0.05 g of Andro were designated as ASFH-L, representing the maximum solubility of Andro facilitated by Cu²⁺ coordination, resulting in a homogeneous system with 100% drug complexation. Hydrogels containing 0.2 g of ASFH-H, intentionally exceeding the solubility limit 4-fold, represent a heterogeneous biphasic system containing both complexed drug and free particulate Andro. This design enables systematic investigation of how drug physical state influences material properties and biological performance.
Rheological properties
Rheological properties were assessed using two complementary protocols50,51. Basic viscoelastic characterization was performed on a DHR-1 rotational rheometer (Waters, USA) with parallel-plate geometry. Hydrogel specimens were prepared as circular disks (60 mm diameter, 3 mm thickness) and gently trimmed to match the plate. Unless otherwise stated, the gap was set at 1.0 mm, and edges were lightly coated with low-viscosity mineral oil to minimize evaporation. Frequency sweeps were performed at 25 °C with a fixed strain amplitude of 1% (within the linear viscoelastic regime), over an angular frequency range of 0.1–100 rad/s. Temperature ramps were conducted from 3 to 45 °C at a heating rate of 2 °C/min, with strain amplitude fixed at 1% and oscillation frequency at 10 rad/s.
For self-healing assessment, consecutive amplitude sweep tests were performed on a HAAKE MARS40 rotational rheometer (Thermo Fisher Scientific, USA) equipped with parallel-plate geometry (20 mm diameter, 1 mm gap) at 25 °C. ASFH-L specimens were subjected to three successive amplitude sweeps at a constant oscillation frequency of 1 Hz, scanning strain amplitude from 0.1% to 1000% in each sweep. Between consecutive sweeps, a 60 s recovery interval was applied during which the hydrogel was maintained at 0.1% strain amplitude and 1 Hz frequency to allow network reformation. Throughout all experiments, the G', G"), and complex viscosity (η) were continuously recorded. Recovery efficiency was calculated as the ratio of G' in the linear viscoelastic region (at 1% strain) after each recovery interval to the initial G' value in the first sweep.
In vitro release study
The drug release profiles of the hydrogels were investigated using the dialysis bag method. Hydrogel samples (1.00 g; ASFH-L and ASFH-H) were loaded into pre-soaked dialysis bags (MWCO 8–14 kDa) and immersed in 250 mL phosphate‑buffered saline (PBS) (pH 7.4) at 37.0 ± 0.5 °C with shaking at 100 rpm (sink conditions maintained by large medium volume relative to Andro solubility). At predetermined intervals (0.5, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 h), 5.0 mL of medium was withdrawn and replaced with equal-volume fresh PBS. Andro concentration was measured by ultraviolet–visible spectrophotometry (UV–Vis) at 232 nm using a 1 cm path length (baseline-corrected PBS blank), with calibration over an appropriate linear range (R² ≥ 0.999) and triplicate technical replicates per time point (n = 3 independent gels per group). The cumulative release percentage was calculated correcting for sample replacement:
Where Qn: cumulative release percentage at the nth time point (%). Cn: drug concentration at the nth time point (μg/mL). V: total volume of the release medium (mL). Ci: Drug concentration at the ith time point (for i = 1 to n − 1) (μg/mL). Vi: sample volume withdrawn at each time point (mL). W: total amount of drug loaded in the dialysis bag (μg).
Microscopic analysis
The microporous structure of the hydrogels was examined using a Quattro S scanning electron microscope (Thermo Fisher Scientific, USA). Lyophilized hydrogel samples (0.8 cm × 0.8 cm × 0.8 cm) were gold-coated using a HITACHI MCI000 ion sputter coater (Tokyo, Japan) for 20 s. SEM imaging was performed at an accelerating voltage of 1.0–5.0 kV, working distance of 5.2–9.3 mm, spot size 3.0, and magnifications ranging from 150× to 2000× as indicated in figure panels, utilizing secondary electron mode. Unlike light microscopy, SEM magnification is controlled electronically by the integrated electron-optical column rather than by interchangeable objective lenses.
Infrared absorption characteristics
FTIR spectra of the hydrogels were obtained using a Thermo Nicolet iS5 spectrometer (Thermo Fisher Scientific, USA). Lyophilized hydrogel samples (1 cm × 1 cm × 1 cm) were scanned with 10–32 scans, a resolution of 4–10 cm−1, and other parameters set as follows: sampling gain 1.0–10.0, moving mirror speed 0.1000–0.9747 cm/min, diaphragm aperture 10.00–100.00%, detector DTGS KBr, beam splitter KBr, and infrared light source. Data analysis was performed using OMNIC and Origin 2021 software.
Raman spectroscopy
Raman spectroscopy was performed on a Thermo Scientific DXR confocal micro-Raman spectrometer. The powdered samples, including pure Andro, Andro/NaCl (1:2 molar ratio) mixture, and Andro/CuCl2 (1:1 molar ratio) mixture, were placed on glass slides and mounted on the sample stage for focused measurement. Spectra were acquired using a 785 nm laser source with an excitation power of 2 mW and a 50× objective. The key instrumental parameters were set as follows: grating at 1200 lines/mm, confocal pinhole aperture of 1.1 µm, and spectral range from 50 to 2000 cm−1. Each measurement was conducted with an acquisition time of 5 s and 20 accumulated scans.
Macroscopic bioadhesion, self-healing, and flowability assays
Bioadhesion assay (wet-tissue simulation)
Fresh porcine skin (dermal side, hair removed, rinsed with PBS) was used as a biologically relevant substrate to mimic human wound tissue. Approximately 2 g of ASFH-L hydrogel (disc format, ~20 mm diameter, 3 mm thickness) was carefully applied to the prepared skin surface and allowed to adhere for 5 min at ambient temperature to establish interfacial bonding. To simulate the hydrated environment of exudate-rich wound beds, the gel–skin construct was then fully immersed in deionized water at room temperature for 30 s, introducing a hydration challenge representative of physiological wound conditions. After immersion, the construct was gently blotted with filter paper to remove excess surface water and immediately subjected to mechanical stress testing. This consisted of repeated bending (90° flexion) and twisting (45° torsion) cycles—three cycles of each motion—to assess adhesive durability under dynamic mechanical stress. The entire immersion and mechanical testing sequence was repeated for an additional three cycles to evaluate long-term adhesion stability. Adhesion performance was qualitatively scored at each cycle (intact/partial detachment/complete detachment), photographed, and visually documented.
Self-healing assay
ASFH-L hydrogel samples were divided into two portions and colored with blue and red pigments, respectively, to enable visual differentiation. Each colored portion was bisected into two equal semicircular halves using a sharp blade. One blue half and one red half were then carefully aligned at the cut interface and brought into contact at room temperature without external pressure. After allowing 120 s for spontaneous healing via reformation of dynamic coordination bonds, the rejoined construct was manually stretched from both ends to assess self-healing efficacy. Successful healing was indicated by the ability of the hydrogel to withstand elongation without separation at the original interface, demonstrating restoration of mechanical integrity.
Flowability assay
Hydrogel flowability was evaluated by placing approximately 5 g of freshly prepared hydrogel in a transparent glass vial (20 mL capacity). The vial was tilted at a fixed angle of 45° from the vertical, and the hydrogel flow behavior was recorded using time-lapse video imaging at room temperature. The duration between the cessation of visible flow and the complete stabilization of the hydrogel was measured. Longer stabilization intervals indicate reduced fluidity and higher structural integrity, whereas shorter intervals suggest more fluid-like behavior. All measurements were performed in triplicate.
Swelling and degradation properties
Swelling properties
Hydrogel cubes (1 cm × 1 cm × 1 cm; initial mass W₀) were immersed in 15 mL of PBS at room temperature. At predetermined intervals, excess surface water was blotted with filter paper, and the samples were weighed (Wt). The swelling process continued until the hydrogel mass remained constant, indicating equilibrium. The swelling ratio was calculated using:
Where W t is the hydrogel mass at time, and W 0 is the initial mass
Degradation characteristics
Hydrogel cubes (1 cm × 1 cm × 1 cm; initial mass W₀) were incubated in 5 mL of PBS containing 1.5 mg/mL lysozyme in a shaking incubator at 37 °C and 40 rpm. At regular intervals, excess surface water was removed with filter paper, and samples were weighed (Wd). The PBS-lysozyme solution was refreshed every 12 h following protocols outlined in previous studies52,53,54. The weight remaining ratio was calculated as:
Where W d is the hydrogel mass at time, and W 0 is the initial mass
In vitro bacteriostatic properties
All antibacterial and cellular assays in this study were performed using the indirect extract method in accordance with ISO 10993-12, which ensures controlled, sterile exposure to hydrogel leachables under standardized conditions. Direct-contact assays were not performed, as the hydrogel formulation is not amenable to terminal sterilization (e.g., autoclaving or gamma irradiation) without compromising its coordination chemistry and structural integrity.
For antibacterial and cell-based assays, sterile hydrogel extracts were prepared as follows. Hydrogel (2.0 g) was placed in a 10 mL sterile centrifuge tube, filled with sterile deionized water (or culture medium as specified), and incubated at 37 °C for 6 h with gentle shaking. The mixture was centrifuged at 10,000 rpm for 5 min at 4 °C, and the supernatant was collected and passed through a 0.22 µm sterile syringe filter twice. This extract, containing leached Andro, Cu2+, and other mobile components, was used immediately or stored at 4 °C for up to 48 h. Hydrogel sample formats and dimensions for each assay are reported in Supplementary Table S1.
The bacteriostatic activity of the hydrogels against E. coli and S. aureus was evaluated using plate colony counting and inhibition zone assays.
Plate colony counting assay
Bacterial suspensions were prepared at a concentration of 108 CFU/mL and serially diluted as required for subsequent assays. For the preparation of sterile hydrogel extract, 2 g of hydrogel was placed in a 10 mL centrifuge tube, which was then filled with sterile water. The mixture was incubated at 37 °C for 6 h in a shaking incubator, followed by centrifugation at 10,000 rpm for 5 min. The supernatant was collected and filtered twice through a 0.22 μm membrane filter to obtain the sterile hydrogel extract. Each 100 µL aliquot of the diluted bacterial solution was mixed with 200 µL of sterile hydrogel extract and immediately plated onto agar media. Plates were incubated at 37 °C for 12 h. Experimental groups included: Control (bacterial solution only), Matrix (base hydrogel without active components), ASFH-L, and ASFH-H.
Inhibition zone assay
Agar plates were uniformly spread with 100 µL of bacterial suspension (108 CFU/mL). Four holes of approximately 8 mm diameter were punched into each plate. Each hole was treated with 150 µL of sterile saline (blank) or corresponding sterile hydrogel extract. Plates were incubated at 37 °C for 12 h, after which inhibition zones were imaged and measured.
Cytotoxicity assessment
Cytotoxicity was assessed by cell counting kit-8 (CCK-8) following the indirect extract protocol established in section 4.1.9. The experimental groups included the control group, the matrix hydrogel group, the ASFH-L group, and the ASFH-H group. Sterile hydrogel extraction solutions were prepared by dissolving 20 mg of the hydrogel in 10 mL of Dulbecco’s modified Eagle medium (DMEM) (2 mg/mL stock), followed by incubation at 4 °C for 24 h. The resulting solution was subjected to sterile filtration through a 0.22 μm membrane filter twice. Subsequently, the filtered solution was serially diluted with DMEM to achieve final concentrations of 2.0, 1.0, 0.5, and 0.25 mg/mL.
Keratinocytes (HaCaT) and fibroblast cells were seeded into 96-well plates at a density of 5 × 103 cells/well. After 12 h of adhesion, cells were serum-starved (0.5% fetal bovine serum, FBS) for 12 h to achieve cell cycle synchronization (G0/G1 arrest) and minimize proliferation-driven wound closure. Subsequently, 100 μL of the hydrogel extract was added to each well and incubated for 24 h. Following incubation, the extract was removed, and 100 μL of 10% CCK-8 medium was added. The cells were incubated for an additional 1.5 h, after which absorbance values were measured using a microplate reader (Thermo Fisher Scientific, USA). Each experiment was performed in triplicate with n = 6 wells per condition.
Cell migration assay
Cell migration was assessed using the scratch-wound assay with hydrogel extracts prepared as described in section 4.1.9. Four groups were designated: High glucose (HG), matrix hydrogel (HG + matrix hydrogel extract), ASFH-L (HG + ASFH-L extract), and ASFH-H (HG + ASFH-H extract).
HaCaT and HFF-1 cells were seeded in 6-well plates at a density of 4 × 105 cells per well and cultured in DMEM supplemented with 10% FBS until reaching 80-90% confluence. The medium was then replaced with 0.5% FBS-containing medium to minimize the confounding effects of cell proliferation on the migration readout55,56,57. After serum starvation for 12 h, a scratch wound was created using a sterile 200 μL pipette tip, and cellular debris was gently removed by washing with PBS. The glucose concentration was maintained at 40 mM to model hyperglycemic conditions, and hydrogel extracts were added at a concentration of 1 mg/mL based on prior cytotoxicity assay results. Cell migration into the denuded area was monitored at 0, 24, 36, and 48 h post-treatment by phase-contrast microscopy, and wound closure was quantified using ImageJ software.
M1/M2 polarization of RAW.264.7 macrophages
The LPS-induced inflammatory model in RAW 264.7 murine macrophages (RAW 264.7) were utilized to simulate the chronic pro-inflammatory microenvironment observed in DWs, which is characterized by aberrant and sustained macrophage activation and elevated levels of pro-inflammatory mediators such as TNF-α, IL-6, and IL-1β58,59. RAW 264.7 murine macrophages (2 × 106 cells per well) were seeded in 6-well plates containing DMEM supplemented with hydrogel extract (1 mg/mL) and cultured for 24 h. To induce inflammatory activation, cells were treated with LPS (1 μg/mL) for the indicated time periods; untreated cells served as controls. Culture supernatants were collected and analyzed for IL-6, TNF-α and IL-10 concentrations using enzyme-linked immunosorbent assay (ELISA) kits according to the manufacturer’s protocols.
For immunofluorescence analysis, RAW 264.7 cells (1 × 106 per well) were cultured on glass coverslips in 24-well plates. Following experimental treatments, cells were fixed with 4% paraformaldehyde for 15 min at room temperature, permeabilised as required, and blocked with 5% bovine serum albumin (BSA) for 2 h at room temperature. Primary antibodies were applied at optimized dilutions and incubated overnight at 4 °C. After washing, cells were incubated with fluorophore-conjugated secondary antibodies for 1 h at room temperature in the dark. Nuclei were counterstained with DAPI for 5 min, and coverslips were mounted using antifade mounting medium. Images were acquired using a fluorescence microscope equipped with appropriate filter sets.
Network pharmacology research
Pharmacophore-target genes of Andro were retrieved using the PharmMapper database (http://www.lilab-ecust.cn/pharmmapper/)60, which employs a pharmacophore mapping approach to identify potential targets. The predicted targets were selected based on normalized fit scores ≥ 0.70 to ensure prediction reliability. Genes associated with DWs were identified through the GeneCards (https://www.genecards.org/) and OMIM (https://www.omim.org/) databases using the search terms “diabetic wound,” “diabetic ulcer,” and “impaired wound healing.” Only genes with a relevance score ≥15.0 in GeneCards were retained for subsequent analysis to minimize false positives. The intersection of pharmacophore-target genes and pathogenic genes was identified to obtain overlapping genes using a Venn diagram analysis, which yielded the candidate therapeutic targets that form the basis of Andro’s mechanism of action in DW healing.
These intersecting genes were imported into the STRING database (http://stitch.embl.de/) to construct a PPI network, with the following parameters: organism set to “Homo sapiens,” minimum required interaction score set at 0.900 (high confidence), and network edges representing both functional and physical protein associations. Disconnected nodes were hidden to focus on core interacting clusters. The network was visualized using Cytoscape 3.9.1. with the following optimizations to ensure clarity and interpretability: (i) node size was scaled proportionally to degree centrality (number of interactions); (ii) node color was mapped to pathway membership or functional clusters identified through MCODE clustering analysis; (iii) edge thickness represented interaction confidence scores from STRING; (iv) a force-directed layout algorithm (prefuse force-directed layout) was applied with manual adjustments to minimize edge crossings and improve readability; and (v) key hub proteins (top 10% by betweenness centrality) were labeled with enlarged, bold font for emphasis.
GO and KEGG pathway enrichment analyses were performed using the Metascape database (www.metascape.org/). Enrichment analysis parameters were set as follows: minimum overlap = 3 genes, P-value cutoff = 0.01, and minimum enrichment factor = 1.5. Visualizations were generated using Wei Sheng Xin (https://www.bioinformatics.com.cn) to display the top 20 significantly enriched GO terms (biological processes, cellular components, and molecular functions) and KEGG pathways, presented as bubble charts with bubble size representing gene count and color intensity indicating −log₁₀ (P-value).
Molecular docking and SPR
The structure of andrographolide was downloaded from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/), while the protein structures of Rac1, JNK1, Jun, and Fos were obtained from the PCSB PDB database (https://www.pdbus.org/). Binding sites were predicted using the DeepSite (https://open.playmolecule.org/) and Proteins Plus (https://proteins.plus/) databases. Docking simulations were performed using AutoDock Vina 1.5.7, and the results were visualized with PyMOL 3.1 and LigPlus+ 2.2.8.
The interaction between Andro and Rac1 was validated using a Biacore T200 instrument (Cytiva, USA) with a CM5 sensor chip (Cytiva, USA). Rac1 protein (MCE, China) was diluted to 50 μg/mL in sodium acetate and immobilized onto the chip’s second channel at a flow rate of 10 μL/min. Gradual concentrations of analytes were introduced in a gradient pattern, with a flow rate of 30 μL/min for 150 s. After each cycle, the chip was regenerated using 10 mM glycine hydrochloride (pH 2.0) for 5 min. Data were analyzed using the Biacore Insight evaluation software, and binding kinetics were fitted to a 1:1 Langmuir binding model to calculate association and dissociation constants.
MD simulation
MDs simulations were performed using Gromacs 2022 with the GAFF force field for small molecules and the TIP3P water model. The ligand molecule (Andro) was parameterized using the General AMBER Force Field (GAFF), and partial charges were derived using the restrained electrostatic potential (RESP) method at the HF/6-31G* level in Gaussian 09. The protein was parameterized with the AMBER ff14SB force field. No metal coordination parameters were applied, as copper ions were not included in the simulation. Our previous studies have utilized Gromacs and AMBER ff14SB to investigate the biophysical properties of biomaterials such as collagen and gold surfaces61.
The structures of Andro and target proteins from the docking studies were used for MD simulations under constant temperature and pressure with periodic boundary conditions (1.0 nm water boundary). It is important to note that the MD simulations modeled only the protein–ligand binding interactions in aqueous solution and did not include copper ions. In the experimental system, Cu2+ coordinates with Andro within the hydrogel to enhance solubility and facilitate sustained release; however, once Andro is released and internalized by cells, it interacts with intracellular target proteins (Rac1, JNK1, Jun, Fos) independently of copper. Therefore, the simulation systems consisted solely of the protein, andrographolide ligand, water molecules, and counterions (Na⁺/Cl⁻) for charge neutralization.
Hydrogen bonds were constrained using the LINCS algorithm with a 2 fs integration step. ELE interactions were calculated via the particle-mesh Ewald method with a 1.2 nm cutoff, while non-bonded interactions had a 1.0 nm cutoff, updated every 10 steps. The simulation temperature was maintained at 298 K using the V-rescale method, and pressure was controlled at 1 bar using the Berendsen method. To justify the simulation length and define convergence, a production duration of 100 ns was selected based on established precedent for probing protein–ligand binding dynamics and conformational stability62,63. Equilibration and convergence were assessed by multiple standard metrics: (i) RMSD and Rg to assess equilibration and structural stability, (ii) buried SASA to quantify interface formation, (iii) FEL to identify thermodynamically favored conformations, (iv) PCA to capture dominant motions, (v) interaction energy decomposition (electrostatic and van der Waals-VDW), (vi) hydrogen bond occupancy, and (vii) MM-PBSA to estimate binding free energy. These metrics collectively provide a comprehensive picture of binding stability, mechanism, and energetics. Systems were considered equilibrated when RMSD fluctuations stabilized within ± 0.3 nm, and FEL analysis revealed sampling of a dominant low-energy basin. Equilibration comprised 100 ps in both NVT and NPT ensembles, followed by 100 ns production runs, with conformations saved every 10 ps. Trajectories were analyzed using VMD and PyMOL 3.1. The binding free energy between proteins and Andro was calculated using the MMPBSA method via the g_mmpbsa program.
Monitoring of wound healing and perfusion in mice
All animal assays were carried out in accordance with the standard of the National Regulation of China for the Care and Use of Laboratory Animals, which complied with the National Research Council’s Guide for the Care and Use of Laboratory Animals. And all animal procedures adhered to protocols approved by the Animal Ethics Committee of Zunyi Medical University (approval no. ZMU21-2107-037). Twenty-four 8-week-old male SPF-grade C57BL/6J mice (22–25 g) were fasted for 12 h and then administered a single intraperitoneal injection of 1% streptozotocin at 125 mg/kg to induce diabetes. Following 3–5 days for blood glucose stabilization, mice with fasting blood glucose levels ≥ 16.7 mmol/L, measured via tail vein sampling, were considered successfully modeled for type I diabetes mellitus. Anesthesia was achieved with 1.25% tribromoethanol at 24 mg/kg via intraperitoneal injection. Full-thickness skin wounds (0.8 cm diameter) were created bilaterally on the dorsum after hair removal64,65. Mice were randomly assigned to four groups: Control (DWs + saline, n = 6), Matrix Hydrogel (DWs + matrix hydrogel, n = 6), ASFH-L (DWs + ASFH-L, n = 6), and ASFH-H (DWs + ASFH-H, n = 6). Hydrogel treatments were administered twice daily, starting the day after surgery (day 0). Wound areas were photographed on days 3, 7, and 12 post-injury, and blood perfusion at the wound site was assessed using a laser speckle flow imaging system (RFLSI, RWD, Shenzhen, China) on days 3, 7, and 10. Euthanasia was performed at the end of the experiment on day 12 by cervical dislocation under deep anesthesia induced with an intraperitoneal injection of an overdose of 1.25% tribromoethanol. The absence of a pedal reflex was verified to confirm deep anesthesia prior to proceeding, and death was confirmed by the cessation of respiration and cardiac activity. All animal experiments in this study were reported in accordance with the ARRIVE 2.0 guidelines.
Histological staining
Wound tissues were harvested on days 3 and 7, then fixed, dehydrated, clarified, paraffin-embedded, and sectioned for hematoxylin and eosin (H&E), Masson’s trichrome, immunohistochemical, and immunofluorescence staining. Collagen density was assessed via immunohistochemistry using COL I and COL III antibodies. Inflammation was evaluated with F4/80, CD86, and CD206 antibodies, while angiogenesis was assessed using CD31 and α-SMA antibodies.
RT-qPCR
RNA from wound tissues collected on days 3 and 7 was extracted using Trizol reagent. Complementary DNA was synthesized using a Takara Reverse Transcription Kit, followed by qPCR amplification with the provided kit. Relative gene expression was calculated using the 2−(ΔΔCT) assay, with β-Actin as the internal control. Primers were synthesized by Sangon Biotech (Shanghai, China) as detailed in Table 3.
Western blotting
Proteins from wound tissues on days 3 and 7 were extracted using a lysis buffer containing RIPA and PMSF (100:1). Protein concentrations were determined using the BCA assay, and samples were diluted in PBS. Proteins were denatured at 100 °C for 10 min, separated by 12% SDS-PAGE, and transferred to 0.45 μm PVDF membranes. Membranes were blocked with 5% skim milk for 1.5 h, incubated overnight at 4 °C with primary antibodies against Rac1, JNK1, Fos, and Jun, and then with anti-rabbit secondary antibodies. Membranes were stripped with regeneration solution and washed three times with TBST buffer between each step.
Statistical analysis
Data are presented as mean ± standard deviation (SD) from at least three independent experiments. Normality was assessed using the Shapiro–Wilk test. For experiments involving multiple treatment groups at individual time points (e.g., wound closure rate, migration assays, biochemical assays), one-way ANOVA followed by Tukey’s post hoc test was employed to compare group means at each time point separately. This approach was chosen because our primary scientific objective was to identify which treatment was most effective at clinically relevant stages of healing, rather than to model the treatment × time interaction. One-way ANOVA at discrete time points is standard in preclinical wound-healing studies and provides maximal statistical power for between-group comparisons at critical endpoints. Statistical significance was set at P < 0.05. All analyses were conducted using GraphPad Prism 9.0 (GraphPad Software, San Diego, CA, USA).
Data availability
All data are available in the main text or the supplementary materials. The raw datasets used and/or analyzed during the current study are available from the corresponding author.
References
Yin, G. N. et al. Latrophilin-2 is a novel receptor of LRG1 that rescues vascular and neurological abnormalities and restores diabetic erectile function. Exp. Mol. Med. 54, 626–638 (2022).
Fabio, F. et al. Prospective study on microangiopathy in type 2 diabetic foot ulcer. Diabetologia 59, 1542–1548 (2016).
Yang, L., Rong, G. C. & Wu, Q. N. Diabetic foot ulcer: challenges and future. World J. Diab. 13, 1014–1034 (2022).
He, W. et al. The cGAS-STING pathway: a therapeutic target in diabetes and its complications. Burns Trauma 12, tkad050 (2024).
Wu, X. et al. Elucidating the dual roles of apoptosis and necroptosis in diabetic wound healing: implications for therapeutic intervention. Burns Trauma 13, tkae061 (2025).
Deng, J.-Y. et al. Targeting DNA methylation and demethylation in diabetic foot ulcers. J. Adv. Res. 54, 119–131 (2023).
Cui, T. et al. Micro-gel ensembles for accelerated healing of chronic wound via pH regulation. Adv. Sci. 9, e2201254 (2022).
Ma, W. J. et al. Polydopamine decorated microneedles with Fe-MSC-derived nanovesicles encapsulation for wound healing. Adv. Sci. 9, e2103317 (2022).
Xu, Z. et al. Nanofiber-mediated sequential photothermal antibacteria and macrophage polarization for healing MRSA-infected diabetic wounds. J. Nanobiotechnol. 19, 404 (2021).
Raghavan, J. V. et al. Immunomodulatory bandage for accelerated healing of diabetic wounds. ACS Bio. Med. Chem. Au 2, 409–418 (2022).
Wu, X. Q. et al. Macrophage polarization in diabetic wound healing. Burns Trauma 10, tkac051 (2022).
Wang, C. et al. Engineering bioactive self-healing antibacterial exosomes hydrogel for promoting chronic diabetic wound healing and complete skin regeneration. Theranostics 9, 65–76 (2019).
Zou, F. et al. A novel bioactive polyurethane with controlled degradation and L-Arg release used as strong adhesive tissue patch for hemostasis and promoting wound healing. Bioact. Mater. 17, 471–487 (2022).
Zhong, G. F. et al. A photo-induced cross-linking enhanced A and B combined multi-functional spray hydrogel instantly protects and promotes of irregular dynamic wound healing. Small 20, e2309568 (2024).
Han, K. et al. Gelatin-based adhesive hydrogel with self-healing, hemostasis, and electrical conductivity. Int J. Biol. Macromol. 183, 2142–2151 (2021).
Zhang, W. et al. Antibacterial coaxial hydro-membranes accelerate diabetic wound healing by tuning surface immunomodulatory functions. Mater. Today Bio. 16, 100395 (2022).
Sharmeen, S. et al. Polyethylene glycol functionalized carbon nanotubes/gelatin-chitosan nanocomposite: an approach for significant drug release. Bioact. Mater. 3, 236–244 (2018).
Chen, X. et al. Efficient drug delivery and anticancer effect of micelles based on vitamin E succinate and chitosan derivatives. Bioact. Mater. 6, 3025–3035 (2021).
Geng, Y. T. et al. Recent advances in carboxymethyl chitosan-based materials for biomedical applications. Carbohydr. Polym. 305, 120555 (2023).
Tran, Q. T. N., Tan, W. S. D., Wong, W. S. F. & Chai, C. L. L. Polypharmacology of andrographolide: beyond one molecule one target. Nat. Prod. Rep. 38, 682–692 (2021).
Dai, Y. et al. Overview of pharmacological activities of Andrographis paniculata and its major compound andrographolide. Crit. Rev. Food Sci. 59, S17–S29 (2019).
Islam, M. T. Andrographolide, a new hope in the prevention and treatment of metabolic syndrome. Front. Pharm. 8, 571 (2017).
Veeresham, C., Swetha, E., Rao, A. R. & Asres, K. In vitro and in vivo aldose reductase inhibitory activity of standardized extracts and the major constituent of andrographis paniculata. Phytother. Res. 27, 412–416 (2012).
Zhang, Z. et al. Hypoglycemic and beta cell protective effects of andrographolide analogue for diabetes treatment. J. Transl. Med. 7, 62 (2009).
Messire, G., Serreau, R. & Berteina-Raboin, S. Antioxidant effects of catechins (EGCG), andrographolide, and curcuminoids compounds for skin protection, cosmetics, and dermatological uses: an update. Antioxidants 12, 1317 (2023).
Mussard, E. et al. Andrographis paniculata and its bioactive diterpenoids against inflammation and oxidative stress in keratinocytes. Antioxidants 9, 530 (2020).
Mussard, E. et al. Andrographis paniculata and its bioactive diterpenoids protect dermal fibroblasts against inflammation and oxidative stress. Antioxidants 9, 432 (2020).
Li, C. X. et al. Andrographolide suppresses thymic stromal lymphopoietin in phorbol myristate acetate/calcium ionophore A23187-activated mast cells and 2,4-dinitrofluorobenzene-induced atopic dermatitis-like mice model. Drug Des. Dev. Ther. 10, 781–791 (2016).
Shao, F. L. et al. Andrographolide alleviates imiquimod-induced psoriasis in mice via inducing autophagic proteolysis of MyD88. Biochem. Pharm. 115, 94–103 (2016).
Zhan, J. Y. X. et al. Andrographolide sodium bisulfate prevents UV-induced skin photoaging through inhibiting oxidative stress and inflammation. Mediat. Inflamm. 2016, 3271451 (2016).
Wahid, F., Wang, H.-S., Zhong, C. & Chu, L.-Q. Facile fabrication of moldable antibacterial carboxymethyl chitosan supramolecular hydrogels cross-linked by metal ions complexation. Carbohydr. Polym. 165, 455–461 (2017).
Haas, K. L. & Franz, K. J. Application of metal coordination chemistry to explore and manipulate cell biology. Chem. Rev. 109, 4921–4960 (2009).
Lee, Y., Choi, K., Kim, J. E., Cha, S. & Nam, J. M. Integrating, validating, and expanding information space in single-molecule surface-enhanced Raman spectroscopy for biomolecules. ACS Nano 18, 25359–25371 (2024).
Raza, A. & Wu, W. Metal-organic frameworks in oral drug delivery. Asian J. Pharm. Sci. 19, 100951 (2024).
Septiani, D. A., Hakim, A., Patech, L. R., Zulhalifah, Z. & Siswadi, S. Isolation and identification of andrographolide compounds from the leaves of sambiloto plant (Andrographis paniculata ness). Acta Chim. Asian. 4, 108–113 (2021).
Clarke, D. E., Pashuck, E. T., Bertazzo, S., Weaver, J. V. M. & Stevens, M. M. Self-healing, self-assembled β-sheet peptide-poly(γ-glutamic acid) hybrid hydrogels. J. Am. Chem. Soc. 139, 7250–7255 (2017).
Liu, Y. et al. Asiaticoside-nitric oxide promoting diabetic wound healing through the miRNA-21-5p/TGF-β1/SMAD7/TIMP3 signaling pathway. J. Ethnopharmacol. 319, 117266 (2024).
Li, S., Ding, X., Yan, X., Qian, J. & Tan, Q. ceAF ameliorates diabetic wound healing by alleviating inflammation and oxidative stress via TLR4/NF- κ B and Nrf2 pathways. J. Diab. Res. 2023, 2422303 (2023).
Lan, C. C., Wu, C. S., Huang, S. M., Wu, I. H. & Chen, G. S. High-glucose environment enhanced oxidative stress and increased interleukin-8 secretion from keratinocytes: new insights into impaired diabetic wound healing. Diabetes 62, 2530–2538 (2013).
Suzuki, K. et al. Hydrogen sulfide replacement therapy protects the vascular endothelium in hyperglycemia by preserving mitochondrial function. Proc. Natl. Acad. Sci. USA 108, 13829–13834 (2011).
Ming, T. et al. Mono-phosphorylation at Ser4 of barrier-to-autointegration factor (Banf1) significantly reduces its DNA binding capability by inducing critical changes in its local conformation and DNA binding surface. Phys. Chem. Chem. Phys. 25, 24657–24677 (2023).
Tang, M. et al. Targeting the COMMD4–H2B protein complex in lung cancer. Br. J. Cancer 129, 2014–2024 (2023).
Young, M. J. et al. Nicotine binds to the transthyretin-thyroxine complex and reduces its uptake by placental trophoblasts. Mol. Cell Endocrinol. 549, 111642 (2022).
Ye, P. et al. SOX family transcription factors as therapeutic targets in wound healing: a comprehensive review. Int J. Biol. Macromol. 253, 127243 (2023).
Busik, J. V., Mohr, S. & Grant, M. B. Hyperglycemia-induced reactive oxygen species toxicity to endothelial cells is dependent on paracrine mediators. Diabetes 57, 1952–1965 (2008).
Wang, R. et al. Targeting oxidative damage in diabetic foot ulcers: integrative strategies involving antioxidant drugs and nanotechnologies. Burns Trauma 13, tkaf020 (2025).
Uberoi, A., McCready-Vangi, A. & Grice, E. A. The wound microbiota: microbial mechanisms of impaired wound healing and infection. Nat. Rev. Microbiol 22, 507–521 (2024).
Brandt, B., Abou-Eladab, E. F., Tiedge, M. & Walzel, H. Role of the JNK/c-Jun/AP-1 signaling pathway in galectin-1-induced T-cell death. Cell Death Dis. 1, e23 (2010).
Xu, H. et al. Integrative single-cell RNA-Seq and ATAC-Seq analysis of peripheral mononuclear cells in patients with ankylosing spondylitis. Front. Immunol. 12, 760381 (2021).
Yang, L. et al. Initial IL-10 production dominates the therapy of mesenchymal stem cell scaffold in spinal cord injury. Theranostics 14, 879–891 (2024).
Li, Z. et al. Self-healing hydrogel bioelectronics. Adv. Mater. 36, e2306350 (2024).
Rocasalbas, G. et al. Laccase-assisted formation of bioactive chitosan/gelatin hydrogel stabilized with plant polyphenols. Carbohydr. Polym. 92, 989–996 (2013).
Osi, A. R. et al. Three-dimensional-printable thermo/photo-cross-linked methacrylated chitosan-gelatin hydrogel composites for tissue engineering. ACS Appl. Mater. Inter. 13, 22902–22913 (2021).
Ressler, A. et al. Injectable chitosan-hydroxyapatite hydrogels promote the osteogenic differentiation of mesenchymal stem cells. Carbohydr. Polym. 197, 469–477 (2018).
Liu, T. et al. PSMC2 promotes the progression of gastric cancer via induction of RPS15A/mTOR pathway. Oncogenesis 11, 12 (2022).
Jonkman, J. E. et al. An introduction to the wound healing assay using live-cell microscopy. Cell Adh. Migr. 8, 440–451 (2014).
Liang, C. C., Park, A. Y. & Guan, J. L. In vitro scratch assay: a convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2, 329–333 (2007).
Jin, E. et al. Lemon-derived nanoparticle-functionalized hydrogels regulate macrophage reprogramming to promote diabetic wound healing. J. Nanobiotechnol. 23, 68 (2025).
Gurtner, M. R. N. K. C. A. B. G. C. Wound healing: a cellular perspective. Physiol. Rev. 99, 665–706 (2019).
Wang, X. et al. PharmMapper 2017 update: a web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 45, W356–W360 (2017).
Tang, M., Gandhi, N. S., Burrage, K. & Gu, Y. Adsorption Of Collagen-like Peptides Onto Gold Nanosurfaces. Langmuir 35, 4435–4444 (2019).
Wu, X. et al. The Gut Microbiota-xanthurenic Acid-aromatic Hydrocarbon Receptor Axis Mediates The Anticolitic Effects Of Trilobatin. Adv. Sci. 12, e2412234 (2025).
Wei, J. et al. Targeting FDX1 by trilobatin to inhibit cuproptosis in doxorubicin-induced cardiotoxicity. Br. J. Pharm. 182, 2409–2425 (2025).
Mu, X. et al. Asiaticoside–nitric oxide synergistically accelerate diabetic wound healing by regulating key metabolites and SRC/STAT3 signaling. Burns Trauma 13, tkaf009 (2025).
Ye, P. et al. ACNO hydrogel enhances diabetic wound healing by modulating the Bcl-2/Bax/Caspase-3/PARP pathway. Int Immunopharmacol. 147, 113997 (2025).
Acknowledgements
This study was funded by the National Natural Science Foundation of China (82460792, 82160770), the Zunyi Science and Technology Talent Platform Carrier Construction Project (ZSKRPT2023-1), the Department of Science and Technology of Guizhou Province (QKHPTRC-CXTD [2023] 024), the Guizhou Key Laboratory of Modern Traditional Chinese Medicine Creation (Qian Ke He Platform ZSYS [2025] 019), and the Zunyi City Municipal and University Joint Science and Technology Funding Project (Zunshi Kehe HZ [2025]251). The funder played no role in study design, data collection, analysis and interpretation of data, or the writing of this manuscript.
Author information
Authors and Affiliations
Contributions
Penghui Ye—original draft, methodology, investigation, formal analysis, and data curation. Yuhe Dai and Qianbo Zhang—investigation and data curation. Junqi Yang, Lele Liu, and Xiuying Guo—software, Formal analysis, and data curation. Rifang Gu, Ming Tang, Min Tan, Huan Zhu, Jitao Chen, and Felicity Han—Writing—review and editing. Xuqiang Nie—Writing—review and editing, supervision, conceptualization, project administration, and funding acquisition.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Ye, P., Dai, Y., Zhang, Q. et al. Novel copper-ion coordinated andrographolide-loaded hydrogel activates Rac1/JNK1 axis for enhancing diabetic wound healing. npj Regen Med 11, 14 (2026). https://doi.org/10.1038/s41536-026-00457-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41536-026-00457-y
