
Abstract
Improving rice productivity remains essential under land constraints and rising food demand. However, spatial yield variation of super hybrid rice across agroecosystems and the mechanisms driving it is not fully resolved. We evaluated how thermal regimes, nitrogen management and genotype jointly shaped yield differences of super hybrid rice across two ecological regions. Field experiments (2021–2022) used three super-hybrid cultivars — Liangyoupeijiu (LYPJ), Y-liangyou-1 (YLY1) and Y-liangyou-900 (YLY900) — under four N rates (0, 150, 240, and 330 kg ha−1) at Longhui and Changsha. Averaged across varieties and years, grain yield in Longhui exceeded Changsha by 16.8% (2021) and 26.7% (2022). These site differences were associated with higher temperatures in Changsha during panicle initiation and grain filling, which were accompanied by reductions in spikelets per panicle (~ 5.6%), total spikelets (~ 7.7%) and seed-setting rate (~ 10.6%). Longhui also exhibited greater leaf area index, dry-matter accumulation, and crop growth rate, supporting superior sink formation and grain filling. Partial least squares path modeling indicated that crop growth rate, total dry weight, and seed-setting rate mediated much of the observed yield gap. Nitrogen at 240 and 330 kg ha−1 narrowed inter-site yield differences by improving yield components and growth traits. Among cultivars, YLY900 achieved the highest yield, while YLY1 showed the greatest cross-site stability. Under the tested conditions, these results suggest that aligning N management with genotype selection relative to local thermal regimes can help reduce temperature-driven yield losses in super hybrid rice.
Similar content being viewed by others


Introduction
Rice (Oryza sativa L.) is a principal staple for more than half of the world’s population, and yield stability is crucial for global food security1. China remains both the largest producer and consumer of rice, contributing about 29% of global production while accounting for only 19% of the world’s rice area2. To meet the growing food demand under climate change, coordinated efforts in breeding and precision agronomy have been pursued to raise yield potential and resilience.
Since the 1980s, China has made remarkable progress in breeding super-high-yielding rice. The national Super Rice Breeding Program, launched in 1996, combined ideotype selection with heterosis-based breeding. Across four breeding phases, yield targets rose from 10.5 t ha−1 (circa 2000) to 15 t ha−1 by 20143,4. Today, super hybrid rice occupies roughly 30% of China’s rice area and plays a vital role in national food security5.
Compared with conventional hybrids, super hybrid rice typically exhibits improved canopy architecture, greater biomass production and enhanced resource use efficiency. Traits such as thicker, more erect leaves, a well-developed upper canopy, vigorous root systems, efficient post-heading dry matter translocation, and superior N uptake contribute to improved source–sink coordination and high yield potential6,7. Nevertheless, yield performance of these cultivars can be unstable when exposed to diverse environments and interannual climatic variability.
Temperature during the reproductive phase strongly affects spikelet development, grain filling and final yield8,9. Longhui, a high-yield rice zone in southwest Hunan, is characterized by cooler and more stable temperatures during panicle initiation and grain filling, whereas Changsha in the central basin of Hunan experiences higher temperatures and more frequent heat stress in the same stages10. This contrast provides an opportunity to examine how thermal regimes influence super hybrid rice performance and whether genotype selection and nitrogen (N) management can mitigate adverse thermal effects11.
Nitrogen is a key regulator of rice growth and productivity: optimal N enhances yield but excessive application raises environmental risks such as soil acidification and eutrophication12. While studies have explored N use in super hybrid rice13,14, how N interacts with genotype and environment to affect yield stability across ecological regions remains inadequately resolved. Genotype-specific responses to N inputs are especially underexplored, despite evidence that N can improve canopy structure, reproductive sink strength, and stress tolerance15,16. The phenotypic plasticity observed in large-scale super hybrid rice cultivation — driven by spatial variability and microclimate differences — complicates extrapolation of single-site results.
Given the growing complexity of genotype × environment × management (G × E × M) interactions under climate change, there is a need for integrative, multi-site evaluations. Few studies have simultaneously examined temperature variability, gradients of N inputs and contrasting breeding-phase genotypes under multi-year conditions. To fill this gap, we conducted a two-year, two-site experiment using three super hybrid cultivars and four N levels to (i) quantify temperature-driven yield variation across ecological regions, (ii) assess the capacity of N management to buffer thermal stress, and (iii) identify genotype traits associated with yield stability. We further applied random forest and PLS-PM analyses to disentangle direct and indirect effects of environmental, physiological, and yield component traits, providing mechanistic insight for stabilizing super hybrid rice productivity.
Materials and methods
Experimental sites
Field experiments were conducted during 2021 and 2022 at Longhui (N27°28′29″, E110°56′18″) and Changsha (N28°12′14″, E113°16′59″), Hunan Province, China. These two sites were selected to represent ecological conditions in South China: Longhui as a high-yield mountainous rice zone with stable and cooler temperatures, and Changsha as a general-yielding lowland basin prone to thermal stress. This ecological contrast provides an ideal framework to assess temperature-induced yield responses in super hybrid rice. The average soil properties of the upper 20-cm layer over two years indicated that the Changsha sites had a clay loam texture, with a pH of 5.70, organic matter content (SOM) of 23.29 g kg−1, total nitrogen content (TN) of 1.41 g kg−1, Olsen P (AP) of 5.97 mg kg−1, and available potassium (AK) content of 198.17 mg kg−1 (Table 1). Similarly, the Longhui field also exhibited a clay loam texture, with a pH of 5.83, organic matter content of 33.97 g kg−1, total nitrogen of 1.76 g kg−1, Olsen P of 17.82 mg kg−1, and available potassium of 214.00 mg kg−1. Thus, these site characterizations provide a scientifically sound ecological contrast while ensuring experimental comparability.
Plant materials
Liangyoupeijiu (LYPJ), Y-liangyou-1 (YLY1), and Yliangyou-900 (YLY900), these three varieties, certified as super rice by the Ministry of Agriculture of China in 2005, 2006, and 2017, respectively, were used in the experiments. These cultivars represent three distinct breeding phases of the Chinese Super Rice Breeding Program17. LYPJ first met the Phase I yield target, embodying an early ideotype with large panicles and moderate biomass, YLY1 achieved the Phase II target, offering yield stability, and YLY900 set a Phase IV yield record, exhibiting high radiation use efficiency, rapid post-heading growth, and strong sink capacity18. Detailed varietal traits are provided in Table S1.
Experimental design and field management
The experiments were conducted using a randomized complete block design with four nitrogen (N) treatments: N0 (0 kg N ha−1), N1 (150 kg N ha−1), N2 (240 kg N ha−1), and N3 (330 kg N ha−1), each with three replicates. These N application rates were selected to represent a range from no input to typical farmer practices, as well as extended experimental levels commonly used in super hybrid rice cultivation. Previous studies have reported that super hybrid rice can achieve high grain yield (11.42 t ha−1) at approximately 245.9 kg N ha−119. Other research indicated that 300 kg N ha−1 provides a favorable balance between yield and nitrogen use efficiency13. Therefore, we designed a nitrogen gradient covering zero, moderate, optimal, and high input levels to comprehensively assess nitrogen × environment × genotype interactions under different ecological conditions.
Individual plot size was 30 m2, and adjacent plots were separated by 50 cm deep plastic barriers to ensure independent irrigation and nutrient management. Nitrogen was applied as urea (46% N) in three splits: 50% as basal fertilizer (2 days before transplanting), 20% at the tillering stage (7 days after transplanting), and 30% during panicle initiation (Stage II of young panicle differentiation). Phosphorus at a rate of 40 kg P2O5 ha− 1 as single superphosphate was applied as a basal fertilizer, and K as potassium oxide at 100 kg K2O ha− 1 was split equally between the basal and panicle initiation. Phosphorus (superphosphate) and potassium (potassium oxide) were applied at N: P and N: K ratios of 1:0.6 and 1:1.1, respectively, with phosphorus fully applied as basal fertilizer and potassium split into basal (40%), tillering (20%), and panicle initiation (40%) applications.
To synchronize heading dates among varieties with differing growth durations, staggered sowing was implemented. In Changsha, YLY900, YLY1, and LYPJ were sown on 10 May (first batch), while YLY1 and LYPJ received a second sowing on 13 May. In Longhui, YLY900, YLY1, and LYPJ were sown on 10 April (first batch), followed by YLY1 on 18 April (second batch) and LYPJ on 20 April (third batch). Seeds were soaked, disinfected, and germinated before being sown in wet nursery. Twenty-five-day-old seedlings were transplanted at a density of 16.7 cm × 30.0 cm (hill spacing) with two seedlings per hill. Post-transplanting water management included maintaining a 3–5 cm water layer until tillering, followed by intermittent wetting until mid-tillering. Drainage and field drying were applied at peak tillering to suppress ineffective tillers, with re-flooding during panicle initiation and subsequent wet-dry cycles until 10 days before maturity. Insects, weeds and diseases were intensively controlled by chemicals to avoid biomass and yield loss20,21.
Sampling and measurements
Phenological stages (sowing, transplanting, panicle initiation, heading, and maturity) and meteorological parameters (daily rainfall, solar radiation, and temperature) measured by an on-site weather station (CR800, Campbell Scientific Inc., USA) were recorded. At panicle initiation, heading, and maturity stage, 12 representative plants per plot were sampled for growth analysis. the plant samples were separated into leaves, stems and panicles. The green leaf area was measured by a leaf area meter (LI-3000, LI − COR Inc., Lincoln, NE, USA) to determine the leaf area index (LAI). The dry weights of all the plant parts were determined after oven-dried at 105 °C for 30 min and 80 °C to a constant weight, and then the aboveground total dry weight (TDW) was calculated. And crop growth rate (CGR) were calculated using the following equation:
where W1 and W2 represent the total dry biomass measured at two consecutive sampling times, and t1 and t2 denote the corresponding measurement dates.
At maturity, the number of panicles from the 12 plants were counted before hand-threshed, and then the filled grains were separated from the unfilled grains by submerging them into tap water. The empty grains were separated from the half-filled grains by winnowing. Three sub-samples of the filled (30 g), the half-filled (whole) and the empty (3 g) grains were taken to count the grain number. Then the yield components including panicles m− 2, spikelets per panicle, seed-setting rate, 1000-grain weight) were calculated. The grain yield was determined from a 5m2 area in the center of each plot and was adjusted to 13.5% moisture content. The grain moisture content was determined by a digital moisture tester (DMC-700, Seedburo, Chicago, IL, USA). Grain-leaf ratios were calculated as:
Statistical analysis
Data were processed in Excel (Microsoft Office 2019), with results expressed as mean ± standard error (SE). Statistical analyses included ANOVA with LSD post-hoc tests (R 4.1.2), random forest modeling (Python 3.6) to rank trait contributions to yield, and partial least squares path modeling (PLS-PM; SmartPLS 3) for causal inference. PLS-PM validity was assessed by R² for endogenous variables and the Goodness-of-Fit (GoF) index22, with GoF > 0.7 indicating strong model reliability. To isolate environmental effects from genotypic variability, all traits were normalized as relative values (actual value / multi-year, multi-site mean). Yield stability was evaluated using two standard indices: the coefficient of variation (CV), reflecting overall yield fluctuation across environments, and Finlay–Wilkinson regression coefficient (bi), indicating yield responsiveness to environmental quality. These indices were calculated across sites and years to support genotype-specific interpretations.
Results
Meteorological conditions and growth duration
Significant climatic differences were observed between the two sites, particularly in thermal conditions (Fig. S1a–d). Changsha consistently exhibited higher air temperatures across both growing seasons, especially during the panicle initiation and grain-filling stages (Table S2). Daily maximum air temperatures in Changsha frequently exceeded 35 °C, which is known to suppress leaf expansion and biomass accumulation in addition to impairing reproductive development23, whereas Longhui maintained air temperatures within the optimal range for spikelet fertility (25–32°C;24. In addition, Changsha experienced greater variability in solar radiation (Fig. S1 i–l) and more frequent high-intensity rainfall events (Figure S1m–p). In contrast, Longhui maintained relatively stable relative humidity and light conditions during key reproductive periods (Table S2).
Grain yield and yield components
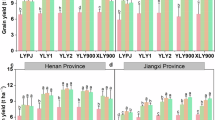
Grain yield and its components were significantly influenced by ecological site, nitrogen level, and genotype (Fig. 1). Longhui consistently outperformed Changsha in grain yield across all varieties and nitrogen treatments. Averaged over varieties and nitrogen levels, yields in Longhui were 16.8% and 26.7% higher than in Changsha in 2021 and 2022, respectively. Correspondingly, Longhui exhibited higher values for key yield components, with increases of 1.8% in panicle number, 5.6% in spikelets per panicle, 7.7% in total spikelets, 10.6% in seed-setting rate, and 2.0% in 1,000-grain weight over the two-year period (Fig. 2).

Grain yield of three super hybrid rice cultivars (LYPJ, YLY1, YLY900) under four nitrogen levels (N0–N3) at Longhui (high-yielding) and Changsha (general-yielding) sites in 2021 and 2022. Error bars denote SE (n = 3). Bars sharing the same lowercase letter within a subplot indicate no significant difference (LSD, p < 0.05).

Yield components (panicles per m2, spikelets panicle− 1, total spikelets, seed-setting rate, 1000-grain weight) of three super hybrid rice cultivars (LYPJ, YLY1, YLY900) under four nitrogen levels (N0–N3) at Longhui (high-yielding) and Changsha (general-yielding) sites in 2021 and 2022. Error bars denote SE (n = 3). Bars sharing the same lowercase letter within a subplot indicate no significant difference (LSD, p < 0.05).
Nitrogen application gradually narrowed the inter-site yield difference. Yield differences between Longhui and Changsha declined from 43.6% under N0 to 13.8%-15.3% under N1-N3, respectively. Nitrogen inputs effectively mitigated the temperature-induced reduction in spikelet number and seed-setting. Specifically, inter-site differences in spikelets per panicle and seed-setting rate declined from 8.1% to 18.5% under N0 to 4.8% and 8.0% on average under nitrogen fertilization (Figure S2). In contrast, inter-site differences in panicle number and 1,000-grain weight remained relatively stable across nitrogen treatments.
Genotypic differences significantly modulated yield stability. Among the three varieties, YLY900 consistently produced the highest grain yield across all conditions, followed by YLY1 and LYPJ. However, YLY1 exhibited the highest yield stability across ecological regions (Table S3). Averaged over two years, the inter-site yield differences were 27.1%, 14.3%, and 23.8% for LYPJ, YLY1, and YLY900, respectively. Notably, under nitrogen application, the yield differences for YLY1 between Longhui and Changsha were only 4.0% in 2021 and 7.7% in 2022, which is much lower than the yield difference between locations under N0 conditions.
Leaf area index and grain–leaf ratio
Leaf area index (LAI) and grain–leaf ratio (GLR) differ notably across ecological regions and nitrogen levels (Fig. 3). LAI values in Longhui were consistently higher than those in Changsha, with average increases of 113.5% in 2021 and 35.3% in 2022. Nitrogen application significantly increased LAI for all varieties at both sites and effectively reduced the inter-site differences. Compared to N0 (139.6%), the average LAI gap between Longhui and Changsha under nitrogen treatments (N1, N2, N3) was reduced to 54.7%, 49.8%, and 51.4%, respectively. Despite the difference in magnitude between years, the pattern of higher LAI in Longhui was consistent.

Leaf area index at heading stage (LAI_HD) and grain-leaf ratio (GLR) of three super hybrid rice cultivars (LYPJ, YLY1, YLY900) under four nitrogen levels (N0–N3) at Longhui (high-yielding) and Changsha (general-yielding) sites in 2021 and 2022. Error bars = SE (n = 3). Bars with shared lowercase letters within a panel are statistically similar (LSD, p < 0.05).
In contrast, GLR exhibited an opposite trend, with generally higher values observed in Changsha than in Longhui. Overall, as nitrogen application rates increased, GLR values showed a declining trend, accompanied by a progressive reduction in inter-site variation. Specifically, the relative differences in GLR between the two sites under the N0, N1, N2, and N3 treatments were 39.4%, 26.8%, 26.0%, and 23.8%, respectively.
Dry matter accumulation, crop growth rate and harvest index
Dry matter accumulation (DMA) and crop growth rate (CGR) varied significantly across sites, nitrogen levels, and varieties. Longhui consistently showed higher DMA at both the heading (HD) and maturity (MA) stages, as well as higher CGR from HD to MA, compared to Changsha (Fig. 4). On average, DMA at HD in Longhui was 14.5% and 6.1% higher than in Changsha in 2021 and 2022, respectively. The differences became more pronounced at the MA stage, reaching 15.6% and 26.9% in the two years. Similarly, CGR in Longhui exceeded that in Changsha by 73.1% in 2021 and 14.6% in 2022.

Dry matter accumulation (DMA) at heading (HD) and maturity (MA) stages and crop growth rate (CGR) from HD to MA of three super hybrid rice cultivars (LYPJ, YLY1, YLY900) under four nitrogen levels (N0–N3) at Longhui (high-yielding) and Changsha (general-yielding) sites in 2021 and 2022. Error bars = SE (n = 3). Bars with common lowercase letters within a subplot are not significantly different (LSD, p < 0.05).
Nitrogen application significantly enhanced DMA in both ecological regions. It also effectively narrowed the inter-site differences. Under N0, the differences in DMA at HD and MA, as well as in CGR, between Longhui and Changsha reached 22.5%, 41.2%, and 87.7%, respectively. With nitrogen application (N1–N3), these differences were reduced to averages of 6.2%, 14.6%, and 29.2%, respectively.
Interestingly, YLY900 generally exhibited greater dry matter accumulation and CGR than LYPJ and YLY1 across all treatments. However, YLY1 showed the smallest inter-site differences in DMA and CGR. The average inter-site differences in DMA at HD and MA, and CGR for YLY1 were 8.6%, 17.0%, and 29.2%, respectively, notably lower than those for LYPJ (11.3%, 20.5%, and 66.2%) and YLY900 (12.8%, 23.7%, and 42.3%).
In contrast, harvest index (HI) remained relatively stable across sites and years, ranging from 54.0% to 55.4%(Figure S3). Although HI tended to decline slightly with increasing nitrogen input, this trend was statistically non-significant.
Factors affecting yield formation
Random Forest and Partial Least Squares Path Modeling (PLS-PM) were used to identify key environmental, agronomic, and yield component variables associated with yield differences between ecological regions (Fig. 5). According to the random forest results, accumulated temperature during the grain filling stage (AT_GFS), daily mean temperature during grain filling stage (DMT_GFS), and maximum temperature during panicle initiation (Tmax_PI) were the top three environmental drivers, collectively accounting for over 50% of the total relative importance (Fig. 5a). Among agronomic traits, total dry weight at maturity (TDW_MA), leaf area index at heading (LAI_HD), and crop growth rate from heading to maturity (CGR_HD–MA) each contributed more than 20% to the variation in yield differences (Fig. 5b), indicating that post-heading biomass accumulation and canopy development are key factors. Regarding yield components, total spikelets number (24.12%), seed-setting rate (SSR, 21.64%), and 1,000-grain weight (GW, 21.44%) were identified as the most influential factors associated with inter-site yield variation (Fig. 5c).

Mechanistic drivers of yield variations in super hybrid rice across contrasting ecological zones. (a–c) Relative importance of environmental variables (a), agronomic traits (b), and yield components (c) in explaining yield differences, as quantified by Random Forest modeling. (d) Direct and indirect effects of meteorological factors, yield components, and agronomic traits on yield variations under Partial Least Squares Path Modeling (PLS-PM). Path coefficients represent standardized beta values (p < 0.05), with arrow thickness proportional to effect magnitude. AT: Accumulated temperature; DMT: Daily mean temperature; SR: Solar radiation; Tmax: Maximum temperature; Tmin: Minimum temperature; GW/LA: Grain weight per leaf area; FG/LA: Filled grains per leaf area; HI: Harvest Index; TDW: Total dry weight; CGR: Crop growth rate; GW: Grain weight; SPP: Spikelets per panicle; SSR: Seed setting rate; HD: Heading stage; GFS: Grain filling stage; PI: Panicle initiation stage; VGS: Vegetative growth stage; MA: Maturity stage.
The PLS-PM analysis further exposed the causal pathways underlying inter-site yield differences (Fig. 5d). The PLS-PM model demonstrated strong high explanatory power (R² = 0.97 for yield difference) and high overall validity (GoF = 0.88), indicating a good model fit and robust reliability. SSR, total spikelets, and GW had significant direct effects on the yield difference between sites. Additionally, CGR_HD–MA and TDW_MA positively affected SSR and GW, respectively, with CGR having direct positive effects on both traits. The impact of environmental factors on yield differences between locations can either directly affect yield components or indirectly affect agronomic traits, thereby affecting yield differences. Notably, Tmax_PI had negative effects on both SSR and total spikelets, while DMT_GFS suppressed SSR and GW and reduced CGR_HD–MA and TDW_MA.
Discussion
Ecological differences in temperature influence spikelet development and fertility, driving yield gaps
Temperature is one of the key environmental factors affecting rice yield, with the reproductive phase being particularly sensitive to thermal temperature. Understanding the response of spikelet development to ecological temperature variation is important for improving yield stability in super hybrid rice. Previous studies have shown that elevated temperatures during panicle initiation and grain filling can adversely affect reproductive development, including disrupted anther development, reduced pollen viability and germination which in turn result in lower seed-setting rate and yield8,25. In addition, higher temperature often shortens the grain-filling period and limits the availability of assimilates, further constraining final grain weight and yield26.
In the present study, rice yield was consistently higher in Longhui compared to Changsha across two years, while divergent thermal regimes between the two geographical locations were identified as a key environmental driver. Changsha experienced higher daily maximum and mean temperatures during the panicle initiation and grain-filling stages (Table S2). Moreover, previous syntheses suggest that vapor pressure deficit (VPD) exceeding ~ 2 kPa is widely used as an indicator of “atmospheric drought” or strong evaporative demand27, which can intensify crop transpiration, alter canopy temperature, and potentially reduce yield. In our study, the cumulative hours with VPD > 2 kPa reached 167.82 and 262 h·kPa in Longhui and Changsha in 2021, and 279.25 and 470.5 h·kPa in 2022(Table S4), further demonstrating the greater atmospheric stress in Changsha. The greater yield variation observed in 2022 aligns with a sharper thermal contrast between sites that year. These thermal conditions, characterized by frequent exposure to maximum temperatures exceeding 35 °C during panicle initiation (Figure S1), were closely associated with reductions in spikelets per panicle and seed-setting rate. Both the random forest and PLS-PM approaches indicated that temperature-induced changes in spikelet traits contributed significantly to the yield difference between sites (Figure S4; Fig. 5). Daily maximum temperatures in Changsha frequently exceeded 35 °C during the reproductive phase (Figure S1), which appeared to limit sink formation and reduce source–sink coordination. These findings highlight that temperatures during panicle initiation and grain filling is primarily responsible for sink formation, ultimately leading to pronounced yield variations across ecological regions.
Ecological variation affects leaf area and biomass accumulation, modulating yield formation
Vegetative growth traits such as leaf area index (LAI), dry matter accumulation (DMA), and crop growth rate (CGR) determine source capacity and serve for reproductive development in rice. Favorable environments enhance canopy structure and photosynthetic efficiency, facilitating larger LAI and greater biomass accumulation, which positively correlated with spikelet development and grain filling28,29,30. However, higher temperatures often limits vegetative growth, constraining source capacity and thereby limiting yield potential10.
In the study, Longhui exhibited significantly greater LAI and biomass accumulation than Changsha, particularly at heading and maturity. This advantage was closely associated with a consistently longer field growth duration (from transplanting to maturity, TP–MA), which was primarily driven by an extended period from transplanting to panicle initiation (TP–PI) (Table S5). The cooler vegetative-phase environment in Longhui, where the mean daily maximum temperature from transplanting to heading was 28.8 °C compared with 32.5 °C in Changsha, likely prolonged and enhanced the efficiency of photosynthetic accumulation and sink formation, thereby contributing to the yield advantage observed at this site. Furthermore, the observed interannual variation in LAI underscores the dynamic response of canopy development to environmental conditions. The more favorable canopy development in Longhui during 2021, for example, also can be attributed to that year’s superior solar radiation and early-season thermal accumulation—highlighting the acute environmental sensitivity of super hybrid rice’s vegetative growth. These advantages translated into higher CGR and greater source support during grain filling. In contrast, the reduced LAI and DMA observed in Changsha suggest a source-limited condition, which likely hindered grain growth and contributed to lower yields. Interestingly, although Changsha exhibited a higher grain–leaf ratio (GLR), this did not result in increased yield. This observation indicates that GLR, in this case, reflects a source deficiency rather than improved assimilate use efficiency. Similar patterns have been observed under heat stress, where source constraints prevent full sink realization despite favorable GLR values31. Therefore, in super hybrid rice, ecological conditions modulate yield through source-sink relationships, with temperature influences on canopy structure and biomass accumulation constituting a critical pathway for environmental impacts on yield formation.
Nitrogen management and genotypic traits alleviate temperature-induced yield losses by enhancing yield components and agronomic performance
Rice yield formation depends on the coordination between genetic traits, crop management practices and environmental conditions. Differences in panicle structure, biomass partitioning, and physiological traits among genotypes interact with nitrogen (N) input to regulate both source and sink capacity. A key genotypic trait influencing this interaction is varietal growth duration. In our study, the field growth duration significantly varied among genotypes, ranking as LYPJ < YLY1 < YLY900, a trend that was strongly and positively correlated with final grain yield (r = 0.78, p < 0.05). This underscores the intrinsic link between a cultivar’s maturity period and its yield potential. Previous studies have reported that super hybrid rice cultivars show considerable variation in growth response and nitrogen use efficiency across environments7,32. Genotypes characterized with large panicles, enhanced post-heading biomass accumulation, and efficient assimilates translocation typically demonstrate superior performance under suboptimal temperature conditions. Furthermore, nitrogen application significantly prolonged the growth duration of all three varieties, particularly during the heading to maturity (HD-MA) stage. This N-induced phenological shift likely extended the grain-filling period, enhancing the accumulation of photoassimilates and thereby mitigating yield losses under stressful conditions, such as the higher temperatures in Changsha. Meanwhile, appropriate N application improves spikelet numbers and sustains grain filling by maintaining physiological activity during reproductive development6,18.
In this study, nitrogen application contributed significantly to narrowing the yield gap between ecological regions by reducing differences in multiple yield-related traits. Under relatively higher temperatures in Changsha, nitrogen application at N2 and N3 levels effectively improved agronomic and reproductive characteristics, including LAI, DMA, CGR, spikelets per panicle and seed-setting rate. These enhancements reduced the spatial disparity in source–sink coordination, which played a central role in stabilizing yield across sites. Genotypic performance further modulated the extent of inter-site variation. Although YLY900 maintained the highest grain yield, it exhibited larger differences in traits such as CGR, DMA, and seed-setting rate between Longhui and Changsha. In contrast, YLY1 exhibited the most stable yield across different nitrogen levels and ecological zones, as evidenced by its lowest coefficient of variation (16.2%) and the lowest bi value across environments (Table S3). It maintained relatively stable DMA and CGR. These findings indicate that YLY1 possesses stronger environmental adaptability due to its stable trait expression under varying thermal conditions. Such genotypic consistency is essential for securing yield stability across ecological regions.
Although molecular or biochemical analyses were not conducted in this study, previous research has shown that nitrogen can enhance sink strength and stress resilience through hormonal and antioxidant pathways. For example, appropriate N supply promotes cytokinin activity, delays senescence, and improves spikelet formation33. It also enhances antioxidant defense under heat stress, which may explain the higher seed-setting rate observed in Changsha under N input34. Future biochemical and gene-level validation would be valuable.
Our findings emphasize that genotype × nitrogen interactions play a critical role in stabilizing yield across thermal environments. The interplay between varietal duration and N management is a critical component of these interactions. Longer-duration varieties like YLY900 can maximize yield in favorable, cooler environments (e.g., Longhui) but may suffer greater yield penalties under heat stress, whereas medium-duration varieties like YLY1 offer greater resilience. For instance, YLY1 maintained high stability under moderate N, while YLY900 achieved maximum yield under high N in optimal zones, suggesting that climate-smart, genotype-specific N strategies can reduce N input by 20–30% without yield penalties35. Although our two sites represent distinct agroclimatic zones in South China, broader multi-site validation is needed to generalize these findings across more diverse environments.
Conclusions
This study highlights the influence of temperature differences between ecological regions on yield formation in super hybrid rice. Elevated temperatures during panicle initiation and grain filling in Changsha reduced spikelet formation and seed-setting rate by 5.6–10.6%, driving a 16.8–26.7% yield gap compared to the cooler ultra-high-yield region Longhui. Higher leaf area index, dry matter accumulation, and crop growth rate in Longhui supported greater yield stability under favorable thermal conditions.
Nitrogen application at N2 and N3 levels reduced inter-site yield differences by up to 30%, primarily by narrowing the gaps in spikelet number, seed-setting rate, LAI, GLR, DMA, and CGR between regions. These improvements helped stabilize yield formation under varying temperature conditions. Notably, genotypic differences further influenced yield stability, super hybrid rice genotypes YLY1 maintained higher crop growth rate (CGR), dry matter accumulation under variable thermal conditions, exhibiting greater yield stability. Together, these results underscore the interactive roles of ecological temperature, nitrogen management, and genotype in regulating rice yield, and demonstrate the potential of genotype specific nitrogen strategies to improve rice yield stability and support climate-resilient rice production in regions with similar ecological conditions.
Data availability
All data generated or analyzed during this study are included in this published article and its supplementary information files.
References
Godfray, H. C. J. et al. Food security: the challenge of feeding 9 billion people. Science 327(5967), 812–818. https://doi.org/10.1126/science.1185383 (2010).
Maclean, J. L. Rice Almanac: Source Book for the Most Important Economic Activity on Earth (International Rice Research Institute, 2002).
Yuan, L. Breeding of super hybrid rice. In Rice Research for Food Security and Poverty Alleviation (eds Peng, S. & Hardy, B.) 143–149 (International Rice Research Institute, 2001).
Wang, F. & Peng, S. Yield potential and nitrogen use efficiency of china’s super rice. J. Integr. Agric. 16(5), 1000–1008. https://doi.org/10.1016/S2095-3119(16)615561-7 (2017).
Yuan, L. Research and development of hybrid rice in China. Sci. Technol. Rev. 34(9), 62–64 (2016). (In Chinese).
Zhang, Y. et al. Yield potential and radiation use efficiency of super hybrid rice grown under subtropical conditions. Field Crops Res. 114(1), 91–98. https://doi.org/10.1016/j.fcr.2009.07.008 (2009).
Bai, P., Bai, R. & Jin, Y. Characteristics and coordination of source-sink relationships in super hybrid rice. Open. Life Sci. 11(1), 470–475. https://doi.org/10.1515/biol-2016-0061 (2016).
Jagadish, S. V. K., Craufurd, P. Q. & Wheeler, T. R. High temperature stress and spikelet fertility in rice (Oryza sativa L). J. Exp. Bot. 58(7), 1627–1635. https://doi.org/10.1093/jxb/erm003 (2007).
Peng, S. et al. Rice yields decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. U.S.A. 101(27), 9971–9975. https://doi.org/10.1073/pnas.0403720101 (2004).
Wang, D. et al. Geographic variation in the yield formation of single-season high-yielding hybrid rice in Southern China. J. Integr. Agric. 20(2), 438–449. https://doi.org/10.1016/S2095-3119(20)63360-3 (2021).
Yu, X. et al. Predicting potential cultivation region and paddy area for Ratoon rice production in China using maxent model. Field Crops Res. 275, 108372. https://doi.org/10.1016/j.fcr.2021.108372 (2022).
Peng, S., Tang, Q. & Zou, Y. Current status and challenges of rice production in China. Plant. Prod. Sci. 12(1), 3–8. https://doi.org/10.1626/pps.12.3 (2009).
Wei, H., Zhang, H., Blumwald, E., Liu, J. & Li, Q. Different characteristics of high yield formation between inbred Japonica super rice and inter-sub-specific hybrid super rice. Field Crops Res. 198, 179–187. https://doi.org/10.1016/j.fcr.2016.09.009 (2016).
Deng, J. et al. Integrated crop management practices improve grain yield and resource use efficiency of super hybrid rice. Front. Plant Sci. 13, 851562. https://doi.org/10.3389/fpls.2022.851562 (2022).
Zhao, X. Effect of amount of N-fertilizer on yield of rice. J. Anhui Agricultural Sci. 37(19), 8937–8938 (2009). (In Chinese).
Pan, J. et al. Optimized nitrogen management enhances lodging resistance of rice and its morpho-anatomical, mechanical, and molecular mechanisms. Sci. Rep. 9, 20274. https://doi.org/10.1038/s41598-019-56620-7 (2019).
Deng, J. et al. The tradeoff between increasing productivity and environmental sustainability in super hybrid rice breeding. Npj Sustainable Agric. 3, 17. https://doi.org/10.1038/s44264-025-00059-z (2025).
Liu, K. et al. High radiation use efficiency improves yield in the recently developed elite hybrid rice Y-liangyou 900. Field Crops Res. 253, 107804. https://doi.org/10.1016/j.fcr.2020.107804 (2020).
Xue, Y. et al. An improved crop management increases grain yield and nitrogen and water use efficiency in rice. Crop Sci. 53(1), 271–284. https://doi.org/10.2135/cropsci2012.06.0360 (2013).
Zhang, Y. et al. Integrated management approaches enabling sustainable rice production under alternate wetting and drying irrigation. Agric. Water Manage. 281, 108265. https://doi.org/10.1016/j.agwat.2023.108265 (2023).
Yao, F. et al. Agronomic performance of high-yielding rice variety grown under alternate wetting and drying irrigation. Field Crops Res. 126, 16–22. https://doi.org/10.1016/j.fcr.2011.09.018 (2012).
Esposito Vinzi, V., Chin, W. W., Henseler, J. & Wang, H. Handbook of Partial Least Squares: Concepts, Methods and Applications (Springer, 2010). https://doi.org/10.1007/978-3-540-32827-8
Xie, X. et al. Effect of photosynthetic characteristic and dry matter accumulation of rice under high temperature at heading stage. Afr. J. Agric. Res. 6(7), 1931–1940. https://doi.org/10.5897/AJAR11.087 (2011).
Matsui, T., Omasa, K. & Horie, T. The difference in sterility due to high temperatures during the flowering period among japonica-rice varieties. Plant. Prod. Sci. 4(2), 90–93. https://doi.org/10.1626/pps.4.90 (2001).
Khan, S. et al. Mechanisms and adaptation strategies to improve heat tolerance in rice: A review. Plants 8(12), 508. https://doi.org/10.3390/plants8110508 (2019).
Zhu, G. et al. Genetic improvements in rice yield and concomitant increases in radiation- and nitrogen-use efficiency in middle reaches of Yangtze river. Sci. Rep. 6, 21049. https://doi.org/10.1038/srep21049 (2016).
Fukai, S. & Mitchell, J. Role of canopy temperature depression in rice. Crop Environ. 1(3), 198–213. https://doi.org/10.1016/j.crope.2022.09.001 (2022).
Acevedo-Siaca, L. G. et al. Variation in photosynthetic induction between rice accessions and its potential for improving productivity. New Phytol. 227(4), 1097–1102. https://doi.org/10.1111/nph.16454 (2020).
Gu, J. et al. Canopy light and nitrogen distributions are related to grain yield and nitrogen use efficiency in rice. Field Crops Res. 206, 74–85. https://doi.org/10.1016/j.fcr.2017.02.021 (2017).
Jiang, P. et al. Comparisons of yield performance and nitrogen response between hybrid and inbred rice under different ecological conditions in Southern China. J. Integr. Agric. 14(7), 1283–1294. https://doi.org/10.1016/S2095-3119(14)60929-1 (2015).
Huang, M. et al. Improvement in super hybrid rice: A comparative study between super hybrid and inbred varieties. Res. Crops. 13(1), 1–10 (2012).
Li, M., Zhang, H., Yang, X., Ge, M. & Ding, Y. Accumulation and utilization of nitrogen, phosphorus and potassium of irrigated rice cultivars with high productivities and high N use efficiencies. Field Crops Res. 161, 55–63. https://doi.org/10.1016/j.fcr.2014.02.007 (2014).
Liu, Y. Q., Li, W. W., Liu, X. Y. & Chu, C. C. Molecular mechanism of tillering response to nitrogen in rice. Hereditas (Beijing). 45(4), 367–378. https://doi.org/10.16288/j.yczz.23-084 (2023).
Zhao, Y. et al. Nitrogen regulated reactive oxygen species metabolism of leaf and grain under elevated temperature during the grain-filling stage to stabilize rice substance accumulation. Environ. Exp. Bot. 229, 106037. https://doi.org/10.1016/j.envexpbot.2024.106037 (2025).
Ju, X. T. et al. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. U.S.A. 106(9), 3041–3046. https://doi.org/10.1073/pnas.0813417106 (2009).
Funding
This research was funded by the National Key Research and Development Program of China (Grant No. 2023YFD2301400) and the Changsha Municipal Science and Technology Project (Grant No. kq2106082).
Author information
Authors and Affiliations
Contributions
Conceptualization, Juan Yang; Data curation, Jianwu Li, Zhiqiang Guo, Juan Yang and Yuhao Jin; Formal analysis, Xinzhen Zhang; Funding acquisition, Yuzhu Zhang; Investigation, Zhiqiang Guo, Juan Yang and Yuhao Jin; Methodology, Juan Yang; Project administration, Yuzhu Zhang; Supervision, Yuzhu Zhang; Validation, Xinzhen Zhang; Writing – original draft, Jianwu Li and Xinzhen Zhang; Writing – review & editing, Yuzhu Zhang.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, J., Zhang, X., Guo, Z. et al. Temperature-driven yield variation of super hybrid rice across ecological regions: mitigation by nitrogen management and genotype selection. Sci Rep 16, 7671 (2026). https://doi.org/10.1038/s41598-026-35957-w
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-026-35957-w
