Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain
the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in
Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles
and JavaScript.
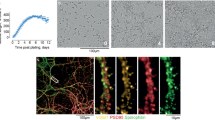
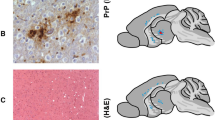
Mammalian prions cause fatal neurodegenerative conditions including Creutzfeldt–Jakob disease in humans and scrapie and bovine spongiform encephalopathy in animals1. Prion infections are typically associated with remarkably prolonged but highly consistent incubation periods followed by a rapid clinical phase. The relationship between prion propagation, generation of neurotoxic species and clinical onset has remained obscure. Prion incubation periods in experimental animals are known to vary inversely with expression level of cellular prion protein. Here we demonstrate that prion propagation in brain proceeds via two distinct phases: a clinically silent exponential phase not rate-limited by prion protein concentration which rapidly reaches a maximal prion titre, followed by a distinct switch to a plateau phase. The latter determines time to clinical onset in a manner inversely proportional to prion protein concentration. These findings demonstrate an uncoupling of infectivity and toxicity. We suggest that prions themselves are not neurotoxic but catalyse the formation of such species from PrPC. Production of neurotoxic species is triggered when prion propagation saturates, leading to a switch from autocatalytic production of infectivity (phase 1) to a toxic (phase 2) pathway.
Gajdusek, D. C. Transmissible and non-transmissible amyloidoses: autocatalytic post-translational conversion of host precursor proteins to β- pleated sheet configurations. J. Neuroimmunol.20, 95–110 (1988)
Come, J. H., Fraser, P. E. & Lansbury, P. T. J. A kinetic model for amyloid formation in the prion diseases: importance of seeding. Proc. Natl Acad. Sci. USA90, 5959–5963 (1993)
Scott, M. et al. Transgenic mice expressing hamster prion protein produce species- specific scrapie infectivity and amyloid plaques. Cell59, 847–857 (1989)
Büeler, H. et al. High prion and PrPSc levels but delayed onset of disease in scrapie-inoculated mice heterozygous for a disrupted PrP gene. Mol. Med.1, 19–30 (1994)
Dickinson, A. G., Meikle, V. M. & Fraser, H. Genetical control of the concentration of ME7 scrapie agent in the brain of mice. J. Comp. Pathol.79, 15–22 (1969)
Kimberlin, R. H. & Walker, C. A. Pathogenesis of mouse scrapie: dynamics of agent replication in spleen, spinal cord and brain after infection by different routes. J. Comp. Pathol.89, 551–562 (1979)
Kimberlin, R. H. & Walker, C. A. Pathogenesis of scrapie (strain 263K) in hamsters infected intracerebrally, intraperitoneally or intraocularly. J. Gen. Virol.67, 255–263 (1986)
Czub, M., Braig, H. R. & Diringer, H. Pathogenesis of scrapie: study of the temporal development of clinical symptoms, of infectivity titres and scrapie-associated fibrils in brains of hamsters infected intraperitoneally. J. Gen. Virol.67, 2005–2009 (1986)
Klohn, P., Stoltze, L., Flechsig, E., Enari, M. & Weissmann, C. A quantitative, highly sensitive cell-based infectivity assay for mouse scrapie prions. Proc. Natl Acad. Sci. USA100, 11666–11671 (2003)
Fischer, M. et al. Prion protein (PrP) with amino-proximal deletions restoring susceptibility of PrP knockout mice to scrapie. EMBO J.15, 1255–1264 (1996)
Cronier, S. et al. Detection and characterization of proteinase K-sensitive disease-related prion protein with thermolysin. Biochem. J.416, 297–305 (2008)
Race, R., Raines, A., Raymond, G. J., Caughey, B. & Chesebro, B. Long-term subclinical carrier state precedes scrapie replication and adaptation in a resistant species: analogies to bovine spongiform encephalopathy and variant Creutzfeldt-Jakob disease in humans. J. Virol.75, 10106–10112 (2001)
Asante, E. et al. BSE prions propagate as either variant CJD-like or sporadic CJD-like prion strains in transgenic mice expressing human prion protein. EMBO J.21, 6358–6366 (2002)
This work was funded by the UK Medical Research Council. We thank J. Wadsworth for critical review of the manuscript, S. Walker for statistical advice, staff of our Biological Services Facility for animal care and technical assistance and R. Young for preparation of figures.
Author information
Authors and Affiliations
MRC Prion Unit and Department of Neurodegenerative Disease, UCL Institute of Neurology, Queen Square, London WC1N 3BG, UK
Malin K. Sandberg, Huda Al-Doujaily, Bernadette Sharps, Anthony R. Clarke & John Collinge
M.K.S., H.A.-D. and B.S. performed the work. M.K.S., A.R.C. and J.C. designed the study and analysed the data. J.C. drafted the manuscript. All authors discussed the results and commented on the manuscript.
Sandberg, M., Al-Doujaily, H., Sharps, B. et al. Prion propagation and toxicity in vivo occur in two distinct mechanistic phases.
Nature470, 540–542 (2011). https://doi.org/10.1038/nature09768
This is another study that clashes with the prion hypothesis, which has dominated the TSE field for almost three decades. Although, surprisingly, the study by Sandberg et al. did not explore pathological markers at tissue or molecular level, which left half of the study, that on studying toxicity, in the shade, this is certainly an elegant and highly valuable study. The problem is with the interpretation of the data in context of the prion hypothesis, which led to a lot of confusion despite obvious intellectual effort by authors to keep the discussion and the conclusions of the study on the rational side; and, that is evident also in the associated editorial piece by Reed Wickner(http://www.nature.com/natur.... We learn, for example, that apparently the 'prions themselves are not neurotoxic'. So, after all, what are the prions if they are not the pathogens causing the TSEs?
As I discussed at length in a series of papers (http://precedings.nature.co... posted in Nature Precedings, the prion hypothesis is flawed. According to a radically new model on the evolutionary origin and function of PrP, this protein is and symbiotic endogenous viral protein that, similar to other proteins implicated in neurodegenerative diseases, including amyloid beta, alpha-synuclein, tau isoforms and huntingtin, is a member of the innate immune system. The malfunction of these proteins leads to a wide range of (innate immune system) autoimmune diseases, including TSEs, Alzheimer's disease, Parkinson's disease, and Huntington's disease. Obviously, in context of this new paradigm on the etiology of these neurodegenerative diseases as autoimmune diseases, the prion hypothesis is misleading; indeed, there are no such things as 'self-replicating protein' or 'prion replication'.
Yervand Karapetyan
Why Tga20 mouse infected with RML dies earlier than the wild type mouse infected with the same strain?
Authors answer to this question is quite simple: Tga20 has more PrP therefore more "Lethal" PrP is generated from it to cause early death of mice. This simplistic view assumes PrP is overexpressed in Tga20 mouse brain equally in all regions of the brain and that PrPSc regional distribution is also identical to that of a wild type mouse.
Not only authors failed to envisage a different possibility but they failed to refer to published data describing in details PrPc, PrPSc distribution in the brains of Tga20 mice infected with several scrapie strains including RML.
PLoS One. 2009 May 29;4(5):e5730. Karapetyan YE et al.Prion strain discrimination based on rapid in vivo amplification and analysis by the cell panel assay.
In my previous comment, I pointed out a major limitation of the study by Sandberg et al., namely, the use of clinical onset as the sole marker for toxicity. Here, I discuss additional problems with the interpretation of the data. The major conclusion of the study was that production of neurotoxic species is triggered when prion propagation saturates, leading to a switch from autocatalytic production of infectivity (phase 1) to a toxic (phase 2) pathway. However, the results of the study do not support such a switch. Specifically, there is no data indicating that toxic species (whatever they are) are not produced during phase 1, or that infectivity (whatever it is) is not produced during phase 2. As I pointed out in my previous comment, the study did not investigate toxicity markers at the tissue, cellular, or molecular level, so there is no way to know if toxic species were produced, or not, in phase 1. However, the results of this study do indicate, although indirectly, that infectivity continues to be produced in phase 2. As shown in the study, infectivity titers undergo rapid clearance; for example, in the Prnp null control mice, the 5.8 logs of LD-50 units used for inoculum were cleared within ten days. Therefore, in order to maintain a steady level (i.e. the plateau level) of infectivity during phase 2, infectivity must be produced at a rate equal to that of clearance. This paper is a revealing example of the limitations imposed by the prion theory, which has been the major working hypothesis in the TSE and associated fields for almost three decades. By portraying prions as novel pathogens that replicate independently of the PrP gene, the prion hypothesis has uncoupled the TSE phenomenon from the function of this gene. Indeed, not a single reference was made in this paper about the physiological function of PrP and on how it might relate to, or explain, the results of the study. As discussed in detail in the series of papers and comments I posted in Nature Precedings (http://precedings.nature.co..., the prion hypothesis has failed to connect some of the most basic scientific aspects concerning PrP and TSE phenomenon, such as: (1) the evolutionary origin of PrP; (2) its physiological function; (3) the TSE-associated pathogenic mechanisms; and (4) TSE-associated transmissibility. The new model addresses and integrates all these aspects into a biological framework that makes evolutionary sense and explains many puzzles associated with this group of devastating diseases. Moreover, unlike the prion hypothesis, this model is consistent with all the experimental data produced during the last half of century, including the results of this study, which are difficult to interpret in context of the prion hypothesis (see my previous comment). According to this new model, the PrP gene is a symbiotic endogenous virus gene. Similar to other proteins implicated in neurodegeneration, such as amyloid beta, alpha-synuclein, tau isoforms, and huntingtin, PrP is a member of the innate immune system. PrP molecules interact with the components of various pathogens (such as viruses) and block their life cycle or, in order to stop the spread of infection, they induce cellular death by various mechanisms such as apoptosis. Similar to other members of the immune system, the PrP gene was selected against pathogenic expression or autoimmune diseases. However, due to genetic mutations or other factors, the PrP molecules inadvertently assemble into oligomeric species or structures that resemble pathogenic components of foreign infectious pathogens. The native PrP molecules, which act as ligands, recognize these species as pathogenic components. Unfortunately, during their immune response, the native PrP molecules assemble into additional pathogenic components, leading to a vicious, autoimmune cycle, which results in cellular death and disease. Therefore, in context of this new model, the concepts of prion replication and prion pathogenesis are obviously flawed. First of all, what has been considered to be the prion replication phenomenon, which has been the famed and highly prized novelty of the prion hypothesis, doesn't exist; in reality, the TSE phenomenon represents a series of cyclic autoimmune reactions. Second, the mechanisms leading to these cyclic autoimmune reactions and to cellular death and disease are intrinsic properties or functions of the native PrP molecules; they are not intrinsic properties or functions of the so called prions. And, obviously, if there is an endogenous virus involved in TSE, then, there is no need for the prion hypothesis. What about TSE infectivity, or transmission phenomena, which are central to this paper and to the prion hypothesis? Because of its viral origin, some of the oligomeric structures produced by PrP during its cyclic autoimmune reactions resemble virus-like structures, a phenomenon that is well documented in the TSE literature although not interpreted this way (see my comment at: http://precedings.nature.co.... These TSE-associated virus-like structures, which resemble the protein-only viral structures or particles produced by many viruses, are highly stable structures and, therefore, they can be passed or transmitted between various cells and tissues, as well as between individuals of the same or different species. However, similar to the protein-only virus particles produced by viruses, which can be transmitted to new individuals and can interfere with the life cycle of the parental or related viruses, or similar to various toxins that can cause pathology and diseases when transmitted to distant cells and tissues, the transmission of TSE associated virus-like structures or particles to new cells and individuals is not a true infectious process, it just mimics one. The TSE strains phenomenon has been at the heart of the controversy regarding the nature of the TSE pathogens for half of a century. Interestingly, this phenomenon has been used for decades as evidence for the viral nature of TSEs and as a pivotal argument against the prion hypothesis. Ironically, this phenomenon is now being increasingly used in support of the prion hypothesis and as evidence for prion-based hereditary information. However, it needs to be realized that the vast majority of TSE cases are not initiated by infectious prions, which means that the prions (whatever they are) initiating these cases are not the progenies of prion replication, nor are they being produced based on prion-encoded hereditary information. So, if there are no such things as prion replication or prion hereditary information, how do we explain the apparent existence of TSE strains? Within the framework of the new model on the etiology of TSEs and other neurodegenerative diseases, the concept of TSE strains is basically flawed. To understand this, we need to consider the criteria used for defining the prion strains, such as the incubation period. Take, for example, the study by Sandberg et al. discussed here. Clearly, the incubation period of the RLM prion strain used in this study varies greatly, from approximately 60 to 260 days, depending on the level of native PrP expression, not on the hereditary properties of the prion strain in the inoculum. Similar to any biochemical or immunological event, if all the factors involved in the TSE autoimmune cyclic reaction, such as the amino acid sequence of the native PrP and its expression level, the genetic and epigenetic background (including factors such as metal ion concentration and pH), and the structural and biochemical specificity and quantity of the inoculum, then, the reaction dynamics and the characteristics of products (i.e. the newly formed PrP auto-immune complexes) will stay relatively constant, mimicking the occurrence of strains. According to the new model on the etiology of TSEs, the PrP cyclic autoimmune reactions lead to the production of a population of diverse PrP oligomeric structures, or immune complexes, with various properties, just as shown in this and other previous studies (see comment above by Biasini, Harris, and Chiesa). Some of these complexes are more toxic or more immunogenic than others and, obviously, the factors (e.g. quinacrine, small RNA molecules) that can influence the formation of PrP immune complexes will have differential effects on their formation, leading to preferential assembly of certain immune complexes versus the others. These processes occur in many biochemical or immunological reaction, but they should not be confused with quasispecies or evolution phenomena seen in infectious pathogens such as viruses. Before concluding this comment, I would like to mention two other issues that are relevant to this study by Sandberg et al. First, similar to the intricate process of folding and assembly of thousands of other proteins into various structures, which uses biological information encoded in other proteins or other molecules that act as partners, templates, chaperons, or just as substrate, the assembly of PrP molecules into new immune complexes uses the biological information encoded the previously formed PrP immune complexes, which represent their substrate. Also, it is very likely that some of the newly formed PrP immune complexes, such as those that are highly immunogenic, are produced at a high rate only when the immune reaction is chaperoned by specific molecules, such as nucleic acids, which might explain some of the results of this and other studies. Second, PrP interact with other related members of the innate immune system, such as amyloid beta, alpha-synuclein, tau isoforms and huntingtin, and with their immune complexes that might resemble pathogenic components; the interactions among this family of proteins run a thin line between protective or pathogenic outcomes, which make understanding the nature of devastating diseases even more complicated. This new model on the etiology of TSEs and other neurodegenerative disorders, including Alzheimer's disease, Parkinson's disease, and Huntington's disease, opens these highly important medical and public health fields to new research avenues and therapeutic approaches. Considering that tens of millions of people are affected by these devastating diseases, any new potential advancement in understanding their etiology deserves serious consideration. Therefore, I encourage the authors of this paper, as well as the community of researchers working in these fields, to engage in a constructive evaluation of these new models and paradigms. And, in that spirit, they should respond to these comments, which challenge this paper and their work at the most fundamental level.
Emiliano Biasini
The study by Sandberg et al. makes the important point that the infectious and toxic forms of PrP are not identical. This is a crucial conclusion, and one that has important implications for understanding the pathogenesis of prion diseases and the development of effective therapies. However, we would like to remind the readers that this concept is not new, and has been extensively investigated and discussed by others in the field. The temporal dissociation between prion infectivity (and PrPSc levels) and clinical or neuropathological manifestation of disease, particularly in Prn-p +/0 and PrP-overexpressing mice, has been pointed out previously (1,2). The Sandberg et al. paper provides a more precise measure of prion titers using the scrapie cell assay, but this dissociation has been known for years, and the hypothesis that infectious and neurotoxic PrPs are distinct molecular species had been put forward before (3). Unfortunately, the Sandberg et al. paper does not provide any new insight into the molecular nature of infectious and toxic PrP. We have investigated this issue in transgenic mice, and provided evidence for subtle differences in the state of aggregation of pathogenic and infectious PrP (4,5). In addition, as we and others have pointed out previously, several lines of evidence suggest that the toxicity of PrPSc is likely to involve corruption of the physiological activity of PrPC (6,7). The possibility that PrPC function can be subverted to generate neurotoxicity has also been highlighted by recent data suggesting that PrPC may function as a receptor for Abeta 1-42 oligomers and thus play a role in Alzheimer's disease (8). In agreement with Dr. Wickner's commentary on this paper (9), we believe that identification of the toxic forms of PrP and how they cause neuronal death will require development of a robust cellular assay capable of measuring prion toxicity in vitro, as well as a deeper understanding of the cellular functions of PrPC.
Emiliano Biasini (1) David A. Harris (1) Roberto Chiesa (2) 1 Department of Biochemistry, Boston University School of Medicine, Boston, MA 02118, USA; 2 Dulbecco Telethon Institute and Mario Negri Institute for Pharmacological Research, Milano, Italy
References. 1. Fischer, M. et al. Prion protein (PrP) with amino-proximal deletions restoring susceptibility of PrP knockout mice to scrapie. EMBO Journal 15, 1255-1264 (1996). 2. Bueler, H. et al. High prion and PrPSc levels but delayed onset of disease in scrapie-inoculated mice heterozygous for a disrupted PrP gene. Molecular Medicine 1, 19-30 (1994). 3. Chiesa, R. & Harris, D. A. Prion diseases: what is the neurotoxic molecule? Neurobiol Dis 8, 743-763. (2001). 4. Chiesa, R. et al. Molecular distinction between pathogenic and infectious properties of the prion protein. J Virol 77, 7611-7622 (2003). 5. Biasini, E., Medrano, A. Z., Thellung, S., Chiesa, R. & Harris, D. A. Multiple biochemical similarities between infectious and non-infectious aggregates of a prion protein carrying an octapeptide insertion. J Neurochem 104, 1293-1308 (2008). 6. Harris, D. A. & True, H. L. New insights into prion structure and toxicity. Neuron 50, 353-357 (2006). 7. Rambold, A. S. et al. Stress-protective signalling of prion protein is corrupted by scrapie prions. EMBO J 27, 1974-1984 (2008). 8. Lauren, J., Gimbel, D. A., Nygaard, H. B., Gilbert, J. W. & Strittmatter, S. M. Cellular prion protein mediates impairment of synaptic plasticity by amyloid-beta oligomers. Nature 457, 1128-1132 (2009). 9. Wickner, R. B. Prion diseases: Infectivity versus toxicity. Nature 470, 470-471, doi:470470a [pii]10.1038/470470a (2011).





Claudiu Bandea
This is another study that clashes with the prion hypothesis, which has dominated the TSE field for almost three decades. Although, surprisingly, the study by Sandberg et al. did not explore pathological markers at tissue or molecular level, which left half of the study, that on studying toxicity, in the shade, this is certainly an elegant and highly valuable study. The problem is with the interpretation of the data in context of the prion hypothesis, which led to a lot of confusion despite obvious intellectual effort by authors to keep the discussion and the conclusions of the study on the rational side; and, that is evident also in the associated editorial piece by Reed Wickner(http://www.nature.com/natur.... We learn, for example, that apparently the 'prions themselves are not neurotoxic'. So, after all, what are the prions if they are not the pathogens causing the TSEs?
As I discussed at length in a series of papers (http://precedings.nature.co... posted in Nature Precedings, the prion hypothesis is flawed. According to a radically new model on the evolutionary origin and function of PrP, this protein is and symbiotic endogenous viral protein that, similar to other proteins implicated in neurodegenerative diseases, including amyloid beta, alpha-synuclein, tau isoforms and huntingtin, is a member of the innate immune system. The malfunction of these proteins leads to a wide range of (innate immune system) autoimmune diseases, including TSEs, Alzheimer's disease, Parkinson's disease, and Huntington's disease. Obviously, in context of this new paradigm on the etiology of these neurodegenerative diseases as autoimmune diseases, the prion hypothesis is misleading; indeed, there are no such things as 'self-replicating protein' or 'prion replication'.
Yervand Karapetyan
Why Tga20 mouse infected with RML dies earlier than the wild type mouse infected with the same strain?
Authors answer to this question is quite simple: Tga20 has more PrP therefore more "Lethal" PrP is generated from it to cause early death of mice. This simplistic view assumes PrP is overexpressed in Tga20 mouse brain equally in all regions of the brain and that PrPSc regional distribution is also identical to that of a wild type mouse.
Not only authors failed to envisage a different possibility but they failed to refer to published data describing in details PrPc, PrPSc distribution in the brains of Tga20 mice infected with several scrapie strains including RML.
PLoS One. 2009 May 29;4(5):e5730.
Karapetyan YE et al.Prion strain discrimination based on rapid in vivo amplification and analysis by the cell panel assay.
http://www.ncbi.nlm.nih.gov...
Claudiu Bandea
In my previous comment, I pointed out a major limitation of the study by Sandberg et al., namely, the use of clinical onset as the sole marker for toxicity. Here, I discuss additional problems with the interpretation of the data.
The major conclusion of the study was that production of neurotoxic species is triggered when prion propagation saturates, leading to a switch from autocatalytic production of infectivity (phase 1) to a toxic (phase 2) pathway. However, the results of the study do not support such a switch. Specifically, there is no data indicating that toxic species (whatever they are) are not produced during phase 1, or that infectivity (whatever it is) is not produced during phase 2. As I pointed out in my previous comment, the study did not investigate toxicity markers at the tissue, cellular, or molecular level, so there is no way to know if toxic species were produced, or not, in phase 1. However, the results of this study do indicate, although indirectly, that infectivity continues to be produced in phase 2. As shown in the study, infectivity titers undergo rapid clearance; for example, in the Prnp null control mice, the 5.8 logs of LD-50 units used for inoculum were cleared within ten days. Therefore, in order to maintain a steady level (i.e. the plateau level) of infectivity during phase 2, infectivity must be produced at a rate equal to that of clearance.
This paper is a revealing example of the limitations imposed by the prion theory, which has been the major working hypothesis in the TSE and associated fields for almost three decades. By portraying prions as novel pathogens that replicate independently of the PrP gene, the prion hypothesis has uncoupled the TSE phenomenon from the function of this gene. Indeed, not a single reference was made in this paper about the physiological function of PrP and on how it might relate to, or explain, the results of the study. As discussed in detail in the series of papers and comments I posted in Nature Precedings (http://precedings.nature.co..., the prion hypothesis has failed to connect some of the most basic scientific aspects concerning PrP and TSE phenomenon, such as: (1) the evolutionary origin of PrP; (2) its physiological function; (3) the TSE-associated pathogenic mechanisms; and (4) TSE-associated transmissibility. The new model addresses and integrates all these aspects into a biological framework that makes evolutionary sense and explains many puzzles associated with this group of devastating diseases. Moreover, unlike the prion hypothesis, this model is consistent with all the experimental data produced during the last half of century, including the results of this study, which are difficult to interpret in context of the prion hypothesis (see my previous comment).
According to this new model, the PrP gene is a symbiotic endogenous virus gene. Similar to other proteins implicated in neurodegeneration, such as amyloid beta, alpha-synuclein, tau isoforms, and huntingtin, PrP is a member of the innate immune system. PrP molecules interact with the components of various pathogens (such as viruses) and block their life cycle or, in order to stop the spread of infection, they induce cellular death by various mechanisms such as apoptosis. Similar to other members of the immune system, the PrP gene was selected against pathogenic expression or autoimmune diseases. However, due to genetic mutations or other factors, the PrP molecules inadvertently assemble into oligomeric species or structures that resemble pathogenic components of foreign infectious pathogens. The native PrP molecules, which act as ligands, recognize these species as pathogenic components. Unfortunately, during their immune response, the native PrP molecules assemble into additional pathogenic components, leading to a vicious, autoimmune cycle, which results in cellular death and disease.
Therefore, in context of this new model, the concepts of prion replication and prion pathogenesis are obviously flawed. First of all, what has been considered to be the prion replication phenomenon, which has been the famed and highly prized novelty of the prion hypothesis, doesn't exist; in reality, the TSE phenomenon represents a series of cyclic autoimmune reactions. Second, the mechanisms leading to these cyclic autoimmune reactions and to cellular death and disease are intrinsic properties or functions of the native PrP molecules; they are not intrinsic properties or functions of the so called prions. And, obviously, if there is an endogenous virus involved in TSE, then, there is no need for the prion hypothesis.
What about TSE infectivity, or transmission phenomena, which are central to this paper and to the prion hypothesis? Because of its viral origin, some of the oligomeric structures produced by PrP during its cyclic autoimmune reactions resemble virus-like structures, a phenomenon that is well documented in the TSE literature although not interpreted this way (see my comment at: http://precedings.nature.co.... These TSE-associated virus-like structures, which resemble the protein-only viral structures or particles produced by many viruses, are highly stable structures and, therefore, they can be passed or transmitted between various cells and tissues, as well as between individuals of the same or different species. However, similar to the protein-only virus particles produced by viruses, which can be transmitted to new individuals and can interfere with the life cycle of the parental or related viruses, or similar to various toxins that can cause pathology and diseases when transmitted to distant cells and tissues, the transmission of TSE associated virus-like structures or particles to new cells and individuals is not a true infectious process, it just mimics one.
The TSE strains phenomenon has been at the heart of the controversy regarding the nature of the TSE pathogens for half of a century. Interestingly, this phenomenon has been used for decades as evidence for the viral nature of TSEs and as a pivotal argument against the prion hypothesis. Ironically, this phenomenon is now being increasingly used in support of the prion hypothesis and as evidence for prion-based hereditary information. However, it needs to be realized that the vast majority of TSE cases are not initiated by infectious prions, which means that the prions (whatever they are) initiating these cases are not the progenies of prion replication, nor are they being produced based on prion-encoded hereditary information. So, if there are no such things as prion replication or prion hereditary information, how do we explain the apparent existence of TSE strains?
Within the framework of the new model on the etiology of TSEs and other neurodegenerative diseases, the concept of TSE strains is basically flawed. To understand this, we need to consider the criteria used for defining the prion strains, such as the incubation period. Take, for example, the study by Sandberg et al. discussed here. Clearly, the incubation period of the RLM prion strain used in this study varies greatly, from approximately 60 to 260 days, depending on the level of native PrP expression, not on the hereditary properties of the prion strain in the inoculum. Similar to any biochemical or immunological event, if all the factors involved in the TSE autoimmune cyclic reaction, such as the amino acid sequence of the native PrP and its expression level, the genetic and epigenetic background (including factors such as metal ion concentration and pH), and the structural and biochemical specificity and quantity of the inoculum, then, the reaction dynamics and the characteristics of products (i.e. the newly formed PrP auto-immune complexes) will stay relatively constant, mimicking the occurrence of strains.
According to the new model on the etiology of TSEs, the PrP cyclic autoimmune reactions lead to the production of a population of diverse PrP oligomeric structures, or immune complexes, with various properties, just as shown in this and other previous studies (see comment above by Biasini, Harris, and Chiesa). Some of these complexes are more toxic or more immunogenic than others and, obviously, the factors (e.g. quinacrine, small RNA molecules) that can influence the formation of PrP immune complexes will have differential effects on their formation, leading to preferential assembly of certain immune complexes versus the others. These processes occur in many biochemical or immunological reaction, but they should not be confused with quasispecies or evolution phenomena seen in infectious pathogens such as viruses.
Before concluding this comment, I would like to mention two other issues that are relevant to this study by Sandberg et al. First, similar to the intricate process of folding and assembly of thousands of other proteins into various structures, which uses biological information encoded in other proteins or other molecules that act as partners, templates, chaperons, or just as substrate, the assembly of PrP molecules into new immune complexes uses the biological information encoded the previously formed PrP immune complexes, which represent their substrate. Also, it is very likely that some of the newly formed PrP immune complexes, such as those that are highly immunogenic, are produced at a high rate only when the immune reaction is chaperoned by specific molecules, such as nucleic acids, which might explain some of the results of this and other studies. Second, PrP interact with other related members of the innate immune system, such as amyloid beta, alpha-synuclein, tau isoforms and huntingtin, and with their immune complexes that might resemble pathogenic components; the interactions among this family of proteins run a thin line between protective or pathogenic outcomes, which make understanding the nature of devastating diseases even more complicated.
This new model on the etiology of TSEs and other neurodegenerative disorders, including Alzheimer's disease, Parkinson's disease, and Huntington's disease, opens these highly important medical and public health fields to new research avenues and therapeutic approaches. Considering that tens of millions of people are affected by these devastating diseases, any new potential advancement in understanding their etiology deserves serious consideration. Therefore, I encourage the authors of this paper, as well as the community of researchers working in these fields, to engage in a constructive evaluation of these new models and paradigms. And, in that spirit, they should respond to these comments, which challenge this paper and their work at the most fundamental level.
Emiliano Biasini
The study by Sandberg et al. makes the important point that the infectious and toxic forms of PrP are not identical. This is a crucial conclusion, and one that has important implications for understanding the pathogenesis of prion diseases and the development of effective therapies. However, we would like to remind the readers that this concept is not new, and has been extensively investigated and discussed by others in the field.
The temporal dissociation between prion infectivity (and PrPSc levels) and clinical or neuropathological manifestation of disease, particularly in Prn-p +/0 and PrP-overexpressing mice, has been pointed out previously (1,2). The Sandberg et al. paper provides a more precise measure of prion titers using the scrapie cell assay, but this dissociation has been known for years, and the hypothesis that infectious and neurotoxic PrPs are distinct molecular species had been put forward before (3). Unfortunately, the Sandberg et al. paper does not provide any new insight into the molecular nature of infectious and toxic PrP. We have investigated this issue in transgenic mice, and provided evidence for subtle differences in the state of aggregation of pathogenic and infectious PrP (4,5). In addition, as we and others have pointed out previously, several lines of evidence suggest that the toxicity of PrPSc is likely to involve corruption of the physiological activity of PrPC (6,7). The possibility that PrPC function can be subverted to generate neurotoxicity has also been highlighted by recent data suggesting that PrPC may function as a receptor for Abeta 1-42 oligomers and thus play a role in Alzheimer's disease (8).
In agreement with Dr. Wickner's commentary on this paper (9), we believe that identification of the toxic forms of PrP and how they cause neuronal death will require development of a robust cellular assay capable of measuring prion toxicity in vitro, as well as a deeper understanding of the cellular functions of PrPC.
Emiliano Biasini (1)
David A. Harris (1)
Roberto Chiesa (2)
1 Department of Biochemistry, Boston University School of Medicine, Boston, MA 02118, USA;
2 Dulbecco Telethon Institute and Mario Negri Institute for Pharmacological Research, Milano, Italy
References.
1. Fischer, M. et al. Prion protein (PrP) with amino-proximal deletions restoring susceptibility of PrP knockout mice to scrapie. EMBO Journal 15, 1255-1264 (1996).
2. Bueler, H. et al. High prion and PrPSc levels but delayed onset of disease in scrapie-inoculated mice heterozygous for a disrupted PrP gene. Molecular Medicine 1, 19-30 (1994).
3. Chiesa, R. & Harris, D. A. Prion diseases: what is the neurotoxic molecule? Neurobiol Dis 8, 743-763. (2001).
4. Chiesa, R. et al. Molecular distinction between pathogenic and infectious properties of the prion protein. J Virol 77, 7611-7622 (2003).
5. Biasini, E., Medrano, A. Z., Thellung, S., Chiesa, R. & Harris, D. A. Multiple biochemical similarities between infectious and non-infectious aggregates of a prion protein carrying an octapeptide insertion. J Neurochem 104, 1293-1308 (2008).
6. Harris, D. A. & True, H. L. New insights into prion structure and toxicity. Neuron 50, 353-357 (2006).
7. Rambold, A. S. et al. Stress-protective signalling of prion protein is corrupted by scrapie prions. EMBO J 27, 1974-1984 (2008).
8. Lauren, J., Gimbel, D. A., Nygaard, H. B., Gilbert, J. W. & Strittmatter, S. M. Cellular prion protein mediates impairment of synaptic plasticity by amyloid-beta oligomers. Nature 457, 1128-1132 (2009).
9. Wickner, R. B. Prion diseases: Infectivity versus toxicity. Nature 470, 470-471, doi:470470a [pii]10.1038/470470a (2011).