
Abstract
Infancy marks a critical period of developing brain-behavior relationships that might influence the emergence of emotional and behavioral problems and psychopathology later in childhood and adolescence. In this review, we describe infant MRI studies that examined the development of neural networks, their associations with emerging emotional reactivity and regulation, and the relationships of caregiver factors, such as psychopathology, parenting behaviors and socioeconomic status (SES), with these developmental trajectories. We highlight the potential of utilizing infant MRI methodologies to identify key neural network structural and functional substrates of current and future emotional reactivity and regulation. Such an approach could identify early objective neural markers of dysregulation problems that are precursors of emotional and behavioral disorders, help monitor the effectiveness of existing interventions, and ultimately guide the development of new interventions for at-risk infants.
Similar content being viewed by others

Introduction
The rapid development of the human brain in the first years of life [1,2,3,4] determines critical brain-behavior relationships that set the stage for future clinical and functional outcomes. In particular, a parallel, growing literature highlights the critical role of the development of large-scale neural networks supporting early emotional reactivity and emerging emotional regulation (ER) in infancy [5,6,7]. Given that emotional dysregulation is a transdiagnostic risk factor for future child emotional problems and psychopathology [8,9,10,11,12], elucidating neural substrates/mapping brain markers early in development could aid identification of infants at risk before onset of disorder or manifestation of symptoms. However, these brain-behavior relationships also need to be considered in the context of caregiver characteristics such as the presence of postpartum psychopathology, the availability of responsive parenting, social and economic resources (SES) [13,14,15,16], each of which is known to play a significant role in shaping the infant’s developing capacity for ER. Given the enormous neuroplasticity of the infant brain during the first year of life [17,18,19,20,21], intervening during this developmental window might have the potential to adaptively alter ER systems and decrease the likelihood of later psychiatric disorders [22]. Along these lines, understanding the contribution of modifiable caregiving factors to the development of infant neural networks supporting ER could also inform family-based intervention targets to improve the health and well-being of at-risk infants.
The goal of this review is to describe the current literature regarding associations among the development in infancy of large-scale neural networks and infant emotional reactivity and emerging ER, and the relationships of caregiver factors with these developing trajectories. We first summarize evidence showing that emotional dysregulation is a transdiagnostic risk for future psychopathology. We then describe the roles of large-scale neural networks in emotional reactivity and ER, and how these neural networks develop in the first years of life. We next present findings from studies that examined relationships among the development of these large-scale neural networks and emerging ER in infancy, and relationships among the development of these large-scale neural networks in infancy and future psychopathology later in childhood. Following this, we highlight how caregiver factors are associated with these developing brain-behavior relationships in infancy, and the roles of social contextual factors and the sex of the child in moderating these relationships. We conclude with a summary of these findings and limitations, future research directions and clinical implications.
Disrupted ER in infancy is a transdiagnostic risk for future psychopathology
ER involves the integration of emotion and reward evaluative, attentional, self-referential monitoring, self-regulation/inhibition, and executive function processes that modulate or maintain the intensity and valence of emotional experiences [23, 24]. The Emotion Dynamics Model [25] provides a framework for operationalizing ER along key dimensions (e.g., intensity of emotional reactivity; latency from initial arousal to peak arousal intensity; duration of response; time to recovery). Thus, when the intensity, duration and/or rapidity of the emotional experience (e.g., frustration, joy) is ‘too much’ or ‘too little’ to function appropriately within a social environment, then ER is needed [24,25,26,27]. Positive and negative forms of emotional reactivity, i.e., positive and negative emotionality, can be measured reliably in human infants within the first months of life [28,29,30,31]. Infants displaying high levels of negative emotionality (NE) cry frequently and intensely in response to novelty and limitations and are difficult to soothe [32, 33]. These constitutionally-based behaviors are consistent across settings [34], and tend to be more common in males than females [35, 36]. Although there is modest rank-order stability in NE across the first years of life [37,38,39], this is superimposed on mean-level increases in NE during this period of development [28, 40,41,42]. High NE in infancy is also a robust predictor of behavioral and emotional problems later in childhood [33, 43,44,45,46] and even into adolescence and adulthood [47]. When assessed in the childhood years, NE predicts the development of depression and anxiety, suicidal behavior, behavior problems and substance abuse [48,49,50,51,52,53,54,55]. Thus, high infant NE represents an important early vulnerability factor for a broad range of later emerging functional impairments [56].
Displays of positive emotionality (PE) such as smiling, laughter and high intensity pleasure, may be orthogonal to NE [57, 58], and can also be assessed reliably in the first months of life [59, 60]. PE shows significant change across the first year [61]. Although less research has focused on the predictive utility of PE, low PE in infancy may predict behavioral inhibition in early childhood [8], whereas by middle childhood, low PE is associated with later depression [9,10,11, 62,63,64]. Alternatively, some research suggests that it is the ratio of low PE relative to high NE that represents the greatest risk for later depressive symptoms [9].
Infant efforts to self-regulate (e.g. self-soothing when distressed) are evident as early as the first few months of life [65]. Although developmental advances in the ability to regulate emotional responding may continue into adulthood [66], the most dramatic gains in ER capacities occur in the first few years of life [67]. Normative increases in ER capacities contribute to other development progressions, e.g., decreases in aggression and increases in compliance, and effective cognitive and social engagement [68, 69]. Conversely, deficits in ER in infancy are implicated in the etiology of childhood psychopathology [70,71,72], including aggression [12, 73, 74] and other disruptive behavior disorders [69, 72, 75, 76].
In adulthood, large-scale neural networks play important roles in emotional reactivity and ER
Neural networks contributing importantly to ER include: the medial prefrontal cortical (mPFC)-posterior cingulate/posterior parietal cortical (precuneus)-centered default mode network (DMN) [77, 78]; the dorsal and rostral anterior cingulate cortical (d/rACC)-anterior insula-centered salience network (SN) [79]; and the dorsolateral prefrontal cortex (dlPFC) and lateral posterior parietal cortex (lPPC)-centered central executive network (CEN)/frontoparietal network (FPN) [79, 80].
The DMN has long been thought to support self-referential processing [81, 82]. More recently, however, the DMN has emerged as a key neural network in which different DMN nodes, including mPFC and posterior cingulate cortex (PCC), integrate self reference, social cognition, episodic memory, language and semantic memory to develop “frames of thought” within an internal narrative that are suppressed during cognitive task performance, but which can lead to mind wandering during rest or cognitively-undemanding contexts [83]. The internal narrative shaped by these cognitive processes is key to the development of a ”sense of self” that in turn is a prerequisite for the capacity for ER and other adaptive regulatory processes [84].
Critically, the DMN is very closely connected with other neural networks, which facilitates the switching between the internally-driven processes subserved by the DMN nodes described above and externally-driven cognitive processes that are subserved by these other neural networks. A key neural network thought to control such switching is the SN, which detects the most contextually important information to guide behavior and social cognition [82, 85]. To facilitate this, the SN, and the anterior insula node in the SN in particular [86, 87], is thought to suppress the DMN and engage neural networks such as the CEN [88]. Here, the CEN dlPFC integrates information from multiple sources to support rule-based decision-making, the lPPC, attentional processes [89, 90], and the CEN as a whole, planning and top-down inhibitory control processes [91]. This Triple Network Model is supported by an increasing literature indicating inverse connectivity between the DMN and CEN during cognitive task performance [92,93,94], with the nodes within the DMN that subserve different internally-driven cognitive processes having specific patterns of connectivity with neural networks important for externally-driven cognitive task performance [95]. Furthermore, difficulty in disengaging the DMN, reflected in aberrant connectivity between the DMN and the CEN during cognitive task performance and rest, is evident in individuals with various psychiatric disorders, including mood and anxiety disorders [92, 93, 96,97,98,99]. These findings highlight the importance of the DMN and its context-dependent interactions with other neural networks in ER capacity [100,101,102]. It should be noted, however, that DMN nodes can show increased connectivity during task performance with cortical regions that are important for the organization of goal-directed behavior (multiple demand cortical regions centered on CEN dlPFC and dACC and SN anterior insula) [103] when it is important to combine cognitive processes subserved by the DMN (e.g., prior knowledge about the self) with the goals of the current task [104]. This aligns with the idea that the DMN is a central neural network for integrating external and internally-focused (i.e., self-related) information [105], which further underscores the importance of the DMN and its connectivity with other neural networks in ER and self-regulation processes in general.
The SN and CEN also have specific roles in ER. The SN is critical for attention to emotionally salient information, as well as switching between DMN-centered internally-focused and CEN-centered externally-focused processing, as described above. Other large-scale neural networks that are closely aligned and often equated with the SN include the cinguloopercular network (CO), centered on more dorsal regions of the ACC than the SN and the opercular region of the ventrolateral prefrontal cortex, and the ventral attention network (VAN), centered on the right ventrolateral prefrontal cortex and temporoparietal junction [106, 107]. Together, these three closely aligned neural networks are important for attending to and integrating sensory, self-referential and emotionally-salient information with cognitive processes in order to guide complex behaviors such as social behavior that include ER [108]. Contributing to the specific role of the SN in ER is the amygdala, a key region supporting emotion processing [109] and a component of the SN [79, 82], which allows the SN to integrate emotional perceptual and motivational processes. Within the SN, the medial PFC, in particular the rostral ACC (rACC), is thought to modulate amygdala activity in response to emotionally salient stimuli [110], and has strong connectivity with CEN and SN prefrontal regions [111]. For example, more resilient individuals show greater mPFC-amygdala connectivity [112, 113], which is supported by a large rodent literature indicating greater mPFC (rACC and dACC homologs) projections to the amygdala in more resilient/ dominant animals [114,115,116]. The CEN has a critical role in ER via the roles of the dlPFC in integrating information from multiple sources and supporting rule-based decision-making and planning, the lPPC in attentional processes [89, 90], and the CEN as a whole, in planning and top-down inhibitory control processes [91], and reappraisal of negative emotional contexts [102, 117].
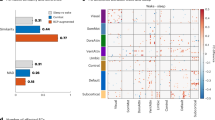
Major white matter (WM) tracts within and among these networks are the cingulum bundle (CB), connecting CEN parietal and prefrontal cortical regions [118]; the anterior corpus callosum (including the forceps minor; FM), providing lateral and medial prefrontal cortical interhemispheric connectivity [119] across the CEN and DMN; and the uncinate fasciculus (UF), which connects the temporal pole with the inferior frontal lobe and posterior orbitofrontal cortices (OFC) [120, 121], regions important for monitoring and learning the emotional (especially reward) value of stimuli [122] (Fig. 1).

[mPFC (Medial Prefrontal Cortex), dlPFC (Dorsolateral Prefrontal Cortex), dACC (Dorsal Anterior Cingulate Cortex)/MCC (Midcingulate Cortex), PCC (Posterior Cingulate Cortex), OFC (Orbitofrontal Cortex), DMN (Default Mode Network), CEN (Central Executive Network), SN (Salience Network)].
The following sections focus on MRI studies that have advanced understanding of the development of these neural networks, and the relationships with developing ER capacity, in infancy.
Feasibility of MRI in infants
Recent and safe pediatric MRI developments have made it possible to conduct MRI studies in infants. For example, the majority of infant MRI studies that are included in this review (except for Ball et al. [123] who used sedation) [124] used the feed-and-bundle approach in order to ensure imaging quality by minimizing excessive movement without sedation. Additionally, pediatric imaging-optimized head coils [125] and acquisition sequences [126] have been implemented to enhance scanning quality while ensuring safety through low specific absorption rate (SAR) and fast acquisition time. Furthermore, manual inspection remains a standard quality control approach to ensure data integrity for subsequent processing and analysis. These infant MRI methodological developments have advanced understanding of the following early neurodevelopmental processes.
Early development of large-scale neural networks
It is well-established that large-scale neural networks associated with emotional reactivity and ER are developing during the first years of life [6, 127,128,129] and continue throughout childhood and adulthood [130,131,132,133,134]. While sensorimotor, auditory and visual networks develop early [135,136,137], the DMN, SN and CEN, and amygdala connectivity with these large-scale networks, continue to develop during early childhood [138,139,140], including large increases in gray matter (GM) volume in cortical and subcortical regions in these networks in the first year [19, 129, 141] and into childhood [142,143,144,145]. In parallel, resting-state functional connectivity (rsFC) among these networks undergoes substantial developmental changes during infancy, initially strengthening within individual networks before increasing inter-network integration [146, 147]. Both intra- and inter-network changes predominantly reflect logarithmic patterns of growth in connectivity [148].
The DMN is initially weaker in neonates, but in infancy becomes more synchronized, particularly between mPFC and PCC regions [147, 149]. Overall adult-like DMN topology is reached around one year old, with continued maturity in the second year of life, mirroring the emergence of early self-referential and social cognitive processing [147, 149]. However, the DMN is still immature and/or sparsely connected until ages 7–9 and continues strengthening into early adolescence [150, 151]. The DMN is the fastest developing among the higher order networks, followed by the right CEN (frontoparietal), SN and the left CEN (frontoparietal) [6] with the CEN still being immature at the end of the first year [152]. The amygdala’s connections with these networks also continue to develop throughout infancy and childhood, reflecting increasing functional specialization in affective and cognitive processing [104, 121].
The functional integration and roles of these networks evolve across development. Specifically, over the first two years of life, decreasing network-level connectivity occurs between DMN-SN and CEN-SN, reflecting the emergence of distinct specializations, while increasing connectivity occurs between right CEN-DMN and left and right CEN [6]. In childhood through emerging adulthood (ages 7–20), inter-network connectivity between higher-order large-scale networks undergoes much development [153], with children displaying weaker DMN-SN and CEN-SN connectivity compared with adults [82]. In particular, the flexibility of switching between distinct inter-network dynamics is weaker in childhood, with adults having rapid-switching of more transient connections between DMN, CEN and SN [154].
WM tracts in these networks develop throughout infancy [155,156,157]. Several tracts, including those described above, show significant increases in fractional anisotropy (FA, the ratio of longitudinal versus transverse water diffusivity, reflecting the degree of longitudinal fiber alignment/collinearity of fibers), and decreases in radial diffusivity (RD, the extent of transverse water diffusivity in WM tracts, thought to reflect non-collinearity of fibers and/or damage to myelin) during the first two years of life [158, 159]. Increases in FA in the majority of WM tracts, especially those connecting visual and subcortical regions implicated in emotional reactivity and ER, continue across the early school years [160,161,162,163]. These microstructural changes reflect the progressive assembly of pathways critical for integrating ER and executive function [18, 164].
The UF demonstrates rapid early development in infancy [18, 165]; maturation of this tract is particularly relevant for emotion-processing circuits, with aberrant UF microstructure linked to less ER capacity [166]. Similarly, the CB develops rapidly over the first year of life [18], and has a long developmental trajectory into adulthood [161] facilitating cognitive control and attentional flexibility [167]. The FM also undergoes significant maturation during infancy [18, 168], contributing to large-scale network integration. This tract plays an important role in the development of higher-order cognitive functions, including ER with patients with emotional dysregulation disorders displaying lower FM structural integrity [169]. These changes in WM microstructure correspond with increasing rsFC within and among the DMN, SN, and CEN [5, 170], with evidence suggesting that infants exhibiting stronger intra- and inter-network connectivity patterns demonstrate more stable regulatory behaviors later in infancy [170, 171].
Relationships among the development of these large-scale neural networks and ER in infancy
A small but increasing number of studies have examined relationships among the development of these large-scale neural networks and ER in infancy [123, 170, 172,173,174]. We reported associations among greater amygdala-DMN and -SN, and lower amygdala-CEN, FC and lower infant positive emotionality (PE), which we replicated in an independent infant sample [94]. These findings parallel those from other infant studies that reported significant relationships between greater amygdala-SN and -DMN FC and greater NE [171, 175, 176]. One study, however, reported an association between greater DMN internetwork FC in early infancy and lower NE later in infancy [177]; and another study reported inverse associations among DMN, SN and CEN and novelty-evoked distress in 4-month infants [178]. Our group also showed associations between greater CB volume, potentially reflecting reduced synaptic pruning [179], and dampened concurrent PE [93]; and reported in 3-month infants that lower UF fiber collinearity predicted greater NE at 9-months [180]. Similarly, another study reported an association between lower early infant fiber collinearity in the inferior stria terminalis, a white matter (WM) tract connecting the amygdala and anterior hypothalamus, and greater infant fear later in infancy [181]. Using multimodal mediation, we further showed that lower rsFC between CEN and DMN structures suppressed the otherwise negative relationship between greater CB volume and dampened concurrent PE [93]. These findings highlight the complex interplay between infant emotionality, brain structure, and functional connectivity in large-scale networks during early development. Further, infant emotionality and rsFC share a bidirectional relationship, where early emotional behaviors can shape neural connectivity patterns in ER networks, and this dynamic interaction underscores the importance of neural plasticity in early emotional development [182].
In a recent study, we used Neurite Orientation Dispersion and Density Imaging (NODDI) to estimate the WM microstructural integrity and myelination of prefrontal cortical regions, using the neurite density index (NDI), and dispersion, using the orientation dispersion index (ODI). These indices of WM microstructure have potential to more accurately evaluate microstructural alterations in the developing brain [183, 184]. We examined relationships in infants among these measures and NE and PE, and reported positive associations among 3-month rACC ODI and cACC NDI and concurrent NE, and 3-month lateral orbitofrontal cortex (lOFC), a region important for decision-making about potential reward value [185, 186], ODI and prospective NE [187]. We also reported a negative association between 3-month dlPFC ODI and concurrent PE. These findings parallel those from our earlier studies of WM microstructure-NE and PE relationships in infants, as they suggest that greater NDI and ODI, reflecting greater microstructural complexity and, likely, more diffuse patterns of connectivity, among prefrontal cortical regions supporting salience perception (rACC), decision-making (lOFC), action selection (cACC), and attentional processes (dlPFC) might result in greater integration of these prefrontal cortical regions with other neural networks, greater attention to salient negative external cues, and thus higher NE and/or lower PE. Furthermore, we have recently shown that in infancy larger increases in ODI in major WM tracts interconnecting neural networks supporting ER, in particular the UF, FM and CB, are associated with disrupted developmental changes, i.e., smaller increases or larger decreases, in PE and ER [188].
We also reported that greater 3-month DMN medial superior frontal cortical volume was associated with higher infant 3-month NE, and that greater 3-month SN and/or ventral attention network (VAN) ventrolateral prefrontal cortical volume predicted lower infant 9-month PE, even after controlling for 3-month PE, highlighting the importance of these latter neural measures in explaining PE change. These findings were replicated in an independent sample [189]. These results add to our previous findings by indicating that greater GM volume in prefrontal cortical regions important for salience perception and attention also predispose to higher levels of infant NE and lower levels of infant PE with increasing age. Other findings indicate positive and inverse associations among subcortical GM volumes and indices of NE in infants [190].
Overall, findings from the above studies suggest that higher levels of NE and lower levels of PE in infancy are associated with: 1. greater levels of endogenous functional connectivity, measured by rsFC, among the amygdala and regions within the DMN and SN, and among regions within the CEN and DMN; 2. lower collinearity (i.e., more diffuse structural connectivity) of fibers in WM tracts among the CEN and DMN (e.g., the CB) and WM tracts connecting the amygdala with the hypothalamus, with worsening ER associated with increasing magnitude of these alterations in WM microstructure during infancy; and 3. greater GM volume in prefrontal cortical regions important for salience perception and attention. One interpretation of these findings is that these patterns of connectivity, WM and GM might result in greater interference by interoceptive and salience processing on executive processing, resulting in greater attention to potentially threatening/worrying emotional stimuli and expression of higher levels of NE and lower levels of PE. Interestingly, greater UF volume was significantly associated with lower concurrent infant NE; but lower orbitofrontal cortex (OFC)-amygdala rsFC, suppressed this otherwise negative relationship, while greater OFC-CEN rsFC mediated this relationship [93]. This finding likely reflects the more specific role of the UF in ER, as it connects prefrontal cortical regions implicated in the evaluation of emotional value of stimuli with the amygdala [191, 192].
Relationships among development of these large-scale neural networks and psychopathology risk
A small number of studies report prospective relationships among infant development of large-scale neural networks, and amygdala rsFC with these networks, and psychopathology risk in early childhood [193,194,195]. Other findings indicate inverse relationships among rsFC among regions in the DMN and SN in infancy and behavioral inhibition at 2 years [170]. Furthermore, there are positive associations among infant amygdala-SN FC and internalizing behaviors in the first 2 years, while inverse associations were found with infant striatal-DMN FC [176, 196, 197]. Additionally, greater levels of atypical development of infant amygdala-DMN FC were associated with greater anxiety at 4 years [198]. Other work has reported prospective associations with other aspects of neurodevelopment. For example, greater amygdala volume in early infancy predicted poorer working memory, a key executive function process, in girls at three years of age [199]. Regarding WM, smaller corpus callosum length measured by cranial ultrasound in 7-week-old infants was associated with greater executive functioning deficits at 4 years of age [200]. Yet, few studies examined associations among large-scale neural networks in infancy and indices of future psychopathology later in childhood. Determining these relationships is a critically important, yet understudied, step toward identifying neural markers of pathophysiological processes in infancy that predispose to future mental health problems in childhood.
Associations between caregiver depression and anxiety and infant neural network-ER relationships
To a large extent, adaptive development of ER systems in infancy is shaped by the quality of interactions with primary caregivers [201, 202]. Developmental studies are consistent in documenting the adverse effects of caregiver (typically maternal) depression and anxiety in the first postpartum year on infant ER [203, 204], and associations with offspring social, emotional and behavioral problems later in childhood [205, 206].
In the field of infant MRI, several studies have reported associations between caregiver depression and anxiety, and other indices of caregiver distress, and the development of GM and WM in neural networks supporting ER in infancy [16, 190, 207,208,209,210,211,212]. In parallel, prenatal maternal depression is associated with greater infant amygdala connectivity with various SN and DMN nodes above and beyond any influence of postnatal depression [213]. These amygdala connectivity patterns are recapitulated later in life among those with major depressive disorder [214, 215]. Furthermore, sensitive parenting has been shown to affect infant ER-related neurodevelopment. For example, maternal sensitivity (timely and appropriate responsiveness to infant signals) was associated with 5-month-old infant DMN rsFC, indicating that infants of more sensitive mothers displayed greater intra-DMN connectivity [14]. Interestingly, greater maternal sensitivity was also associated with greater 6-month hippocampus (a less-well characterized zone of the DMN [216]) connectivity with CEN regions, as well as with lower amygdala connectivity with DMN-related regions [13].
Critically, an emerging literature is exploring how these caregiver factors may help shape developing neural network-ER relationships in infancy. Here, studies reported significant relationships among exposure to postpartum depression, infant WM microstructure and infant NE [217]. Our own work, supported by others [218], shows that more severe postpartum depression and anxiety are associated with greater amygdala-DMN and amygdala-SN, and lower amygdala-CEN, rsFC in 3-month-olds, which in turn are associated with lower levels of infant PE (even after accounting for parenting behaviors) [94], suggesting that caregiver depression and anxiety associate with large-scale neural networks supporting emotional reactivity in early infancy.
Indirect evidence for the effects of caregiver stress, distress or challenges to parenting on infant neural network-ER relationships are suggested by studies that have reported associations between early adversity and infant neural network-ER relationships. Here, maternal childhood and adolescence adverse experiences are associated with smaller neonatal amygdala volume and greater negative emotionality in 6-month old infants. Furthermore, childhood emotional neglect in the mother relates to stronger neonatal rsFC among the amygdala and prefrontal cortical regions in large-scale neural networks including the CEN and SN [219]. Financial strain in the family and/or low SES is also known to affect infant neurodevelopment in regions with broad implications for attention, emotional reactivity, ER and psychopathology, including temporal pole, inferior frontal and anterior cingulate regions [15, 220]. Additionally, prenatal exposure to neighborhood crime was associated with weaker neonatal thalamic-DMN and amygdala-hippocampus rsFC [221].
In addition to the potential modifying effects of environmental factors such as caregiver psychopathology, parenting quality and the broader social context (e.g., financial strain, neighborhood stressors), genetic factors are also likely to play a role. For example, there is a strong genetic component to brain structure and function as evidenced by twin and family studies, with heritability estimates for brain volumes and cortical measures typically ranging from 60–80% [222, 223]. Structural and functional connectivity also show substantial heritability varying by brain white matter tract and network; heritable patterns of functional connectivity were found within the DMN and CEN, with network subregions also showing heritable intra-network connectivity [223, 224]. About half of the inter- and intra-network connectivity of canonical resting-state networks show up to 53% heritability, with stable genetic influences during adolescence [225]. A multi-generational longitudinal study found that individuals at high familial risk for depression show increased DMN connectivity and decreased DMN-CEN negative connectivity, indicating potential biomarkers for depression risk [226]. Despite increasing interest in elucidating environmental and genetic risk factors that influence the development of large-scale neural networks subserving ER in infancy through childhood, many gaps remain. In particular, work is urgently needed to understand the ways in which caregiver resilience, positive well-being and social support could serve as buffers in contexts of stress to support adaptive infant ER development.
While these findings highlight the importance of caregiver emotional reactivity and the caregiving environment on developing infant neural network-ER relationships, it is noteworthy that relationships exist between infant emotional reactivity and ER and caregiver neural activity, where infant emotional behavior can activate caregiver neural networks that support parenting behaviors [227,228,229]. Furthermore, there are well-documented effects of infant emotional reactivity on caregiving affective responses and parenting behaviors [230, 231]. Together, these findings therefore highlight the presence of bidirectional relationships among caregiver affective responses and associated caregiver neural activity, and infant neural network activity and ER. These bidirectional relationships should be the focus of future studies.
Sex of the infant effects on the development of neural network-ER relationships
There are effects of offspring sex on neurodevelopment that vary across studies [16, 199, 212, 232,233,234,235]. For example, we reported stronger positive relationships between postpartum depression and anxiety and infant amygdala-SN, and amygdala-CEN rsFC, and between the latter infant rsFC measures and lower infant PE, in male relative to female infants [94]. Other research showed stronger relationships between lower amygdala-CEN WM FA and externalizing behaviors in boys than girls [233], while other studies showed stronger effects of early financial strain and maternal depression on neurodevelopment in girls than in boys [212, 235]. Further examination of contextual factors and offspring sex on neural indices of ER in infancy through early childhood is clearly warranted.
A framework for understanding neurodevelopmental trajectories underlying emerging NE, PE and ER in infancy, and for identifying neural indices of future psychopathology risk
Emotional dysregulation, evidenced by high NE and low PE in infancy, is a robust predictor of behavioral and emotional problems later in childhood [8, 33, 45, 46] and even into adolescence and adulthood [47], and thus represents an early transdiagnostic marker of psychopathology [70,71,72]. Findings from MRI studies of infants indicate that large-scale networks, including the DMN, SN and CEN, are critical substrates of emotional expressivity and ER [100,101,102], along with the major WM tracts forming the connections within and between each network, including the CB, FM and UF. These large-scale neural networks develop and increase in synchrony by 1 year, with increasing network-level integration [6, 149] and continued maturation beyond the first postnatal year. Findings suggest that higher levels of NE and lower levels of PE in infancy are associated with greater functional connectivity among the amygdala and regions within the DMN and SN, and among regions within the CEN and DMN. This is accompanied by lower collinearity of fibers in WM tracts among the CEN and DMN and greater GM volume in prefrontal cortical regions important for salience perception and attention. Such integration of structure and function facilitates greater maladaptive CEN-DMN connectivity and integration of prefrontal cortical regions with other neural networks, yielding greater attention to salient negative external cues, and thus higher NE and/or lower PE (Fig. 2; Table 1).

While this review focuses on the influence of caregiver factors on infant neural network emotional behavior relationships, it is also acknowledged that bidirectional relationships exist among these factors. [DMN - default mode network, CEN - central executive network, NE - negative emotional reactivity, PE - positive emotional reactivity].
Critically, these indices of neural network structure and function have been shown to be linked with future affective outcomes and psychopathology risk. For example, aberrant infant DMN-SN and amygdala-DMN rsFC were associated with greater childhood behavioral inhibition [170] and anxiety [198], respectively. Furthermore, caregiver factors (e.g., symptoms of psychopathology, parenting behaviors) and limited resources (e.g., low SES) also influence ER-related infant neurodevelopment in ways that recapitulate the infant neural patterns described above, including greater amygdala-large-scale network rsFC [13, 94, 219], and in some studies, indicating that these patterns contribute to high NE and/or low PE [94].
Limitations and future directions
Limitations of the research to date include, in some cases, small sample sizes and/or samples with unbalanced male/female offspring ratios, as well as inherent sampling bias in infant and caregiver participant recruitment and the exclusion of infants who were unable to remain still during scans. Furthermore, some studies examining relationships among social contextual factors and indices of large-scale network structure and function in infancy do not then extend neural findings to examining relationships with measures of infant emotional reactivity or ER. Moreover, future work in larger samples, and studies utilizing advances of integrated multimodal imaging to examine global structure-function relationships will have a beneficial effect on the field, increasing our understanding of neural targets for intervention or prevention of deficient ER in infancy. Additionally, as dysregulated emotional and physiological response to stress in infancy is linked to later negative affective and behavioral outcomes [236,237,238,239], more research is necessary to understand contributions of proximal stress-control neural structures [240, 241] in infancy to other stress response systems and the interactions between infant stress reactivity and ER-related processes.
Conclusion and future clinical implications
Together, findings to date highlight the promise of infant MRI as a valuable approach that can identify key neural network structural and functional correlates of current, and predictors of future, NE, PE and ER (Fig. 2; Table 1). This approach has great potential to provide neural markers to guide early emotional and behavioral disorder risk identification, help monitor the effectiveness of interventions to ameliorate aberrant NE, PE and ER in infancy, and ultimately help to develop new, targeted interventions that are based on an understanding of underlying neural network abnormalities in infancy.
References
Uda S, Matsui M, Tanaka C, Uematsu A, Miura K, Kawana I, et al. Normal development of human brain white matter from infancy to early adulthood: a diffusion tensor imaging study. Dev Neurosci-basel. 2015;37:182–94.
Dubois J, Dehaene-Lambertz G, Kulikova S, Poupon C, Hüppi PS, Hertz-Pannier L. The early development of brain white matter: a review of imaging studies in fetuses, newborns and infants. Neuroscience. 2014;276:48–71.
Deoni SCL, Dean DC, O’Muircheartaigh J, Dirks H, Jerskey BA. Investigating white matter development in infancy and early childhood using myelin water faction and relaxation time mapping. Neuroimage. 2012;63:1038–53.
Vasung L, Abaci Turk E, Ferradal SL, Sutin J, Stout JN, Ahtam B, et al. Exploring early human brain development with structural and physiological neuroimaging. NeuroImage. 2019;187:226–54.
Gao W, Lin W, Grewen K, Gilmore JH. Functional connectivity of the infant human brain. Neuroscientist. 2017;23:169–84.
Gao W, Alcauter S, Smith JK, Gilmore JH, Lin W. Development of human brain cortical network architecture during infancy. Brain Structure Funct. 2015;220:1173–86.
Fransson P, Åden U, Blennow M, Lagercrantz H. The functional architecture of the infant brain as revealed by resting-state fMRI. Cereb Cortex. 2010;21:145–54.
Park S-Y, Belsky J, Putnam S, Crnic K. Infant emotionality, parenting, and 3-year inhibition: exploring stability and lawful discontinuity in a male sample. Developmental Psychol. 1997;33:218.
Dougherty LR, Klein DN, Durbin CE, Hayden EP, Olino TM. Temperamental positive and negative emotionality and children’s depressive symptoms: a longitudinal prospective study from age three to age ten. J Soc Clin Psychol. 2010;29:462–88.
Oldehinkel AJ, Hartman CA, De Winter AF, Veenstra R, Ormel J. Temperament profiles associated with internalizing and externalizing problems in preadolescence. Dev Psychopathology. 2004;16:421–40.
De Pauw SSW, Mervielde I. The role of temperament and personality in problem behaviors of children with ADHD. J Abnorm Child Psychol. 2011;39:277–91.
Crockenberg SC, Leerkes EM, Jó PSB. Predicting aggressive behavior in the third year from infant reactivity and regulation as moderated by maternal behavior. Dev Psychopathology. 2008;20:37–54.
Rifkin-Graboi A, Kong L, Sim LW, Sanmugam S, Broekman BFP, Chen H, et al. Maternal sensitivity, infant limbic structure volume and functional connectivity: a preliminary study. Transl Psychiatry. 2015;5:e668–e668.
Chajes JR, Stern JA, Kelsey CM, Grossmann T. Examining the role of socioeconomic status and maternal sensitivity in predicting functional brain network connectivity in 5-month-old infants. Front Neurosci-switz. 2022;16:892482.
Spann MN, Bansal R, Hao X, Rosen TS, Peterson BS. Prenatal socioeconomic status and social support are associated with neonatal brain morphology, toddler language and psychiatric symptoms. Child Neuropsychol. 2020;26:170–88.
Dean I, Douglas C, Planalp EM, Wooten W, Kecskemeti SR, Adluru N, et al. Association of prenatal maternal depression and anxiety symptoms with infant white matter microstructure. JAMA Pediatrics. 2018;172:973–9.
Gilmore JH, Knickmeyer RC, Gao W. Imaging structural and functional brain development in early childhood. Nat Rev Neurosci. 2018;19:123–37.
Stephens RL, Langworthy BW, Short SJ, Girault JB, Styner MA, Gilmore JH. White matter development from birth to 6 years of age: a longitudinal study. Cereb Cortex. 2020;30:6152–68.
Gilmore JH, Shi F, Woolson SL, Knickmeyer RC, Short SJ, Lin W, et al. Longitudinal development of cortical and subcortical gray matter from birth to 2 years. Cereb Cortex. 2012;22:2478–85.
Deoni SCL, Mercure E, Blasi A, Gasston D, Thomson A, Johnson M, et al. Mapping infant brain myelination with magnetic resonance imaging. J Neurosci. 2011;31:784–91.
Holland D, Chang L, Ernst TM, Curran M, Buchthal SD, Alicata D, et al. Structural growth trajectories and rates of change in the first 3 months of infant brain development. JAMA Neurol. 2014;71:1266–74.
Wakschlag LS, Roberts MY, Flynn RM, Smith JD, Krogh-Jespersen S, Kaat AJ, et al. Future directions for early childhood prevention of mental disorders: a road map to mental health, earlier. J Clin Child Adolesc Psychol. 2019;48:539–54.
Calkins SD, Hill A. Caregiver influences on emerging emotion regulation: biological and environmental transactions in early development. Handbook of emotion regulation. US: New York, NY: Guilford Press; 2007. p. 229–48.
Cole PM, Martin SE, Dennis TA. Emotion regulation as a scientific construct: Methodological challenges and directions for child development research. Child Dev. 2004;75:317–33.
Thompson RA. Emotion regulation: a theme in search of definition. Monogr Soc Res Child Dev. 1994;59:25–52.
Denham SA. Emotional development in young children. New York, NY: Guilford Press; 1998.
Saarni C. The development of emotional competence. New York, NY: Guilford press; 1999.
Gartstein MA, Rothbart MK. Studying infant temperament via the revised infant behavior questionnaire. Infant Behav Dev. 2003;26:64–86.
Pauli-Pott U, Mertesacker B, Beckmann D. Predicting the development of infant emotionality from maternal characteristics. Dev Psychopathology. 2004;16:19–42.
Worobey J, Blajda VM. Temperament ratings at 2 weeks, 2 months, and 1 year: differential stablity of activity and emotionality. Developmental Psychol. 1989;25:257.
Rothbart MK, Chew KH, Gartstein MA. Assessment of temperament in early development. Biobehavioral assessment of the infant. US: New York, NY: Guilford Press; 2001. p. 190–208.
Henderson HA, Wachs TD. Temperament theory and the study of cognition-emotion interactions across development. Developmental Rev. 2007;27:396–427.
Sanson A, Hemphill SA, Smart D. Connections between temperament and social development: a review. Soc Dev. 2004;13:142–70.
Smith AK, Rhee SH, Corley RP, Friedman NP, Hewitt JK, Robinson JL. The magnitude of genetic and environmental influences on parental and observational measures of behavioral inhibition and shyness in toddlerhood. Behav Genet. 2012;42:764–77.
Davis M, Emory E. Sex differences in neonatal stress reactivity. Child Dev. 1995;66:14–27.
Weinberg M, Tronick EZ, Cohn JF, Olson KL. Gender differences in emotional expressivity and self-regulation during early infancy. Developmental Psychol. 1999;35:175–88.
Putnam SP, Gartstein MA, Rothbart MK. Measurement of fine-grained aspects of toddler temperament: the early childhood behavior questionnaire. Infant Behav Dev. 2006;29:386–401.
Putnam SP, Rothbart MK, Gartstein MA. Homotypic and heterotypic continuity of fine-grained temperament during infancy, toddlerhood, and early childhood. Infant Child Dev. 2008;17:387–405.
Rothbart MK, Derryberry D, Hershey K. Stability of temperament in childhood: laboratory infant assessment to parent report at seven years. Temperament and personality development across the life span. p. 85–119 (eds Molfese VJ, Molfese DL) Mahwah, NJ: Lawrence Erlbaum Associates; 2000.
Arterberry ME, Bornstein MH. Development in infancy: a contemporary introduction. New York: Psychology Press; 2014.
Carranza Carnicero JA, Pérez-López J, Del Carmen GS, Martínez-Fuentes MT. A longitudinal study of temperament in infancy: stability and convergence of measures. Eur J Personality. 2000;14:21–37.
Partridge T, Lerner JV. A latent growth-curve approach to difficult temperament. Infant Child Dev. 2007;16:255–65.
Halligan SL, Cooper PJ, Fearon P, Wheeler SL, Crosby M, Murray L. The longitudinal development of emotion regulation capacities in children at risk for externalizing disorders. Dev Psychopathology. 2013;25:391–406.
Gartstein MA, Putnam SP, Rothbart MK. Etiology of preschool behavior problems: contributions of temperament attributes in early childhood. Infant Ment Health J. 2012;33:197–211.
Burgess KB, Marshall PJ, Rubin KH, Fox NA. Infant attachment and temperament as predictors of subsequent externalizing problems and cardiac physiology. J Child Psychol Psychiatry. 2003;44:819–31.
DiCorcia JA, Tronick E. Quotidian resilience: exploring mechanisms that drive resilience from a perspective of everyday stress and coping. Neurosci Biobehav Rev. 2011;35:1593–602.
Causadias JM, Salvatore JE, Sroufe LA. Early patterns of self-regulation as risk and promotive factors in development: a longitudinal study from childhood to adulthood in a high-risk sample. Int J Behav Dev. 2012;36:293–302.
Allen KJ, Gabbay FH. The amphetamine response moderates the relationship between negative emotionality and alcohol use. Alcoholism: Clin Exp Res. 2013;37:348–60.
Eisenberg N, Valiente C, Spinrad TL, Cumberland A, Liew J, Reiser M, et al. Longitudinal relations of children’s effortful control, impulsivity, and negative emotionality to their externalizing, internalizing, and co-occurring behavior problems. Developmental Psychol. 2009;45:988–1008.
Glenn CR, Bagge CL, Osman A. Unique associations between borderline personality disorder features and suicide ideation and attempts in adolescents. J Personality Disord. 2013;27:604–16.
Lonigan CJ, Phillips BM, Hooe ES. Relations of positive and negative affectivity to anxiety and depression in children: evidence from a latent variable longitudinal study. J Consulting Clin Psychol. 2003;71:465–81.
Masse LC, Tremblay RE. Behavior of boys in kindergarten and the onset of substance use during adolescence. Arch Gen Psychiatry. 1997;54:62–68.
Oliva EM, Keyes M, Iacono WG, McGue M. Adolescent substance use groups: antecedent and concurrent personality differences in a longitudinal study. J Personality. 2012;80:769–93.
Slutske WS, Moffitt TE, Poulton R, Caspi A. Undercontrolled temperament at age 3 predicts disordered gambling at age 32: a longitudinal study of a complete birth cohort. Psychological Sci. 2012;23:510–6.
Vitaro F, Barker ED, Boivin M, Brendgen M, Tremblay RE. Do early difficult temperament and harsh parenting differentially predict reactive and proactive aggression? J Abnorm Child Psychol. 2006;34:685–95.
Krueger RF, Markon KE. Reinterpreting comorbidity: a model-based approach to understanding and classifying psychopathology. Annu Rev Clin Psychol. 2006;2:111–33.
Tellegen A Structures of mood and personality and their relevance to assessing anxiety, with an emphasis on self-report. In: Maser AHTJD (ed). Anxiety and the anxiety disorders. Hillsdale, NJ, England: Lawrence Erlbaum Associates, Inc; 1985, pp 681-706.
Watson D, Wiese D, Vaidya J, Tellegen A. The two general activation systems of affect: Structural findings, evolutionary considerations, and psychobiological evidence. J Personality Soc Psychol. 1999;76:820–38.
Bolzani LH, Messinger DS, Yale M, Dondi M Smiling in infancy An empirical reflection on the smile US: Lewiston, NY: Edwin Mellen Press; 2002, pp 111-36.
Bolzani Dinehart LH, Messinger DS, Acosta SI, Cassel T, Ambadar Z, Cohn J. Adult perceptions of positive and negative infant emotional expressions. Infancy. 2005;8:279–303.
Belsky J, Fish M, Isabella RA. Continuity and discontinuity in infant negative and positive emotionality: family antecedents and attachment consequences. Developmental Psychol. 1991;27:421–31.
Anderson E, Hope DA. A review of the tripartite model for understanding the link between anxiety and depression in youth. Clin Psychol Rev. 2008;28:275–87.
Shankman SA, Tenke CE, Bruder GE, Durbin C, Hayden EP, Klein DN. Low positive emotionality in young children: association with EEG asymmetry. Dev Psychopathology. 2005;17:85–98.
Hayden EP, Klein DN, Durbin CE, Olino TM. Positive emotionality at age 3 predicts cognitive styles in 7-year-old children. Dev Psychopathology. 2006;18:409–23.
Crockenberg SC, Leerkes EM. Infant and maternal behaviors regulate infant reactivity to novelty at 6 months. Developmental Psychol. 2004;40:1123–32.
Charles ST, Carstensen LL. Emotion regulation and aging. Handb Emot Regul. 2007;307:327.
Kopp CB. Antecedents of self-regulation: a developmental perspective. Developmental Psychol. 1982;18:199.
Gilliom M, Shaw DS, Beck JE, Schonberg MA, Lukon JL. Anger regulation in disadvantaged preschool boys: strategies, antecedents, and the development of self-control. Developmental Psychol. 2002;38:222.
Kochanska G, Knaack A. Effortful control as a personality characteristic of young children: antecedents, correlates, and consequences. J personality. 2003;71:1087–112.
Kim J, Cicchetti D. Longitudinal pathways linking child maltreatment, emotion regulation, peer relations, and psychopathology. J Child Psychol Psychiatry. 2010;51:706–16.
Cicchetti D, Ackerman BP, Izard CE. Emotions and emotion regulation in developmental psychopathology. Dev Psychopathology. 1995;7:1–10.
Kochanska G, Tjebkes JL, Fortnan DR. Children’s emerging regulation of conduct: Restraint, compliance, and internalization from infancy to the second year. Child Dev. 1998;69:1378–89.
Melnick SM, Hinshaw SP. Emotion regulation and parenting in AD/HD and comparison boys: Linkages with social behaviors and peer preference. J Abnorm Child Psychol. 2000;28:73–86.
Marsee MA, Frick PJ. Exploring the cognitive and emotional correlates to proactive and reactive aggression in a sample of detained girls. J Abnorm Child Psychol. 2007;35:969–81.
Eisenberg N, Smith C, Sadovsky A, Spinrad T, Baumeister R, Vohs K. Handbook of self-regulation: research, theory, and applications. New York, NY: Guilford Press; 2004.
Hinshaw SP, Lee SS. Conduct and oppositional defiant disorders. Child Psychopathology. 2003;2:144–98.
Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proc Natl Acad Sci USA. 2001;98:676–82.
Gusnard DA, Akbudak E, Shulman GL, Raichle ME. Medial prefrontal cortex and self-referential mental activity: relation to a default mode of brain function. Proc Natl Acad Sci USA. 2001;98:4259–64.
Seeley WW, Menon V, Schatzberg AF, Keller J, Glover GH, Kenna H, et al. Dissociable intrinsic connectivity networks for salience processing and executive control. J Neuroscience: Off J Soc Neurosci. 2007;27:2349–56.
Liston C, Matalon S, Hare TA, Davidson MC, Casey B. Anterior cingulate and posterior parietal cortices are sensitive to dissociable forms of conflict in a task-switching paradigm. Neuron. 2006;50:643–53.
Spreng RN, Mar RA, Kim AS. The common neural basis of autobiographical memory, prospection, navigation, theory of mind, and the default mode: a quantitative meta-analysis. J Cognit Neurosci. 2009;21:489–510.
Menon V. Developmental pathways to functional brain networks: emerging principles. Trends Cognit Sci. 2013;17:627–40.
Menon V. 20 years of the default mode network: a review and synthesis. Neuron. 2023;111:2469–87.
Winsler AE, Fernyhough CE, Montero IE Private speech, executive functioning, and the development of verbal self-regulation. Cambridge: Cambridge University Press 2009
Menon V, Uddin LQ. Saliency, switching, attention and control: a network model of insula function. Brain Structure Funct. 2010;214:655–67.
Molnar-Szakacs I, Uddin LQ. Anterior insula as a gatekeeper of executive control. Neurosci Biobehav Rev. 2022;139:104736.
Diveica V, Koldewyn K, Binney RJ. Establishing a role of the semantic control network in social cognitive processing: a meta-analysis of functional neuroimaging studies. NeuroImage. 2021;245:118702.
Menon V. The triple network model, insight, and large-scale brain organization in autism. Biol Psychiatry. 2018;84:236–8.
Reep RL, Corwin JV. Posterior parietal cortex as part of a neural network for directed attention in rats. Neurobiol Learn Mem. 2009;91:104–13.
Malhotra P, Coulthard EJ, Husain M. Role of right posterior parietal cortex in maintaining attention to spatial locations over time. Brain. 2009;132:645–60.
Dosenbach NU, Fair DA, Cohen AL, Schlaggar BL, Petersen SE. A dual-networks architecture of top-down control. Trends Cognit Sci. 2008;12:99–105.
Fournier JC, Bertocci M, Ladouceur CD, Bonar L, Monk K, Abdul-Waalee H, et al. Neural function during emotion regulation and future depressive symptoms in youth at risk for affective disorders. Neuropsychopharmacology. 2021;46:1340–7.
Banihashemi L, Schmithorst VJ, Bertocci MA, Samolyk A, Zhang Y, Lima Santos JP, et al. Neural network functional interactions mediate or suppress white matter–emotional behavior relationships in infants. Biol Psychiatry. 2023;94:57–67.
Phillips ML, Schmithorst VJ, Banihashemi L, Taylor M, Samolyk A, Northrup JB, et al. Patterns of infant amygdala connectivity mediate the impact of high caregiver affect on reducing infant smiling: discovery and replication. Biol Psychiatry. 2021;90:342–52.
Gordon EM, Laumann TO, Marek S, Raut RV, Gratton C, Newbold DJ, et al. Default-mode network streams for coupling to language and control systems. Proc Natl Acad Sci. 2020;117:17308–19.
Bertocci MA, Afriyie-Agyemang Y, Rozovsky R, Iyengar S, Stiffler R, Aslam HA, et al. Altered patterns of central executive, default mode and salience network activity and connectivity are associated with current and future depression risk in two independent young adult samples. Mol Psychiatry. 2023;28:1046–56.
Borserio BJ, Sharpley CF, Bitsika V, Sarmukadam K, Fourie PJ, Agnew LL. Default mode network activity in depression subtypes. Rev Neurosci. 2021;32:597–613.
Ronde M, van der Zee EA, Kas MJH. Default mode network dynamics: an integrated neurocircuitry perspective on social dysfunction in human brain disorders. Neurosci Biobehav Rev. 2024;164:105839.
Yoon S, Kim TD, Kim J, Lyoo IK. Altered functional activity in bipolar disorder: a comprehensive review from a large-scale network perspective. Brain Behav. 2021;11:e01953.
Beauchaine TP, Zisner A. Motivation, emotion regulation, and the latent structure of psychopathology: an integrative and convergent historical perspective. Int J Psychophysiol. 2017;119:108–18.
Nieh EH, Kim SY, Namburi P, Tye KM. Optogenetic dissection of neural circuits underlying emotional valence and motivated behaviors. Brain Res. 2013;1511:73–92.
Phillips ML, Ladouceur CD, Drevets WC. A neural model of voluntary and automatic emotion regulation: implications for understanding the pathophysiology and neurodevelopment of bipolar disorder. Mol Psychiatry. 2008;13:833–57.
Crittenden BM, Mitchell DJ, Duncan J. Task encoding across the multiple demand cortex is consistent with a frontoparietal and cingulo-opercular dual networks distinction. J Neurosci. 2016;36:6147.
Smallwood J, Bernhardt BC, Leech R, Bzdok D, Jefferies E, Margulies DS. The default mode network in cognition: a topographical perspective. Nat Rev Neurosci. 2021;22:503–13.
Yeshurun Y, Nguyen M, Hasson U. The default mode network: where the idiosyncratic self meets the shared social world. Nat Rev Neurosci. 2021;22:181–92.
Sestieri C, Corbetta M, Spadone S, Romani GL, Shulman GL. Domain-general signals in the cingulo-opercular network for visuospatial attention and episodic memory. J Cognit Neurosci. 2014;26:551–68.
Farrant K, Uddin LQ. Asymmetric development of dorsal and ventral attention networks in the human brain. Developmental Cognit Neurosci. 2015;12:165–74.
Menon V Salience network. In: Toga AW (ed). Brain mapping: an encyclopedic reference. Academic Press 2015, vol. 2, pp 597-611.
Phelps EA, LeDoux JE. Contributions of the amygdala to emotion processing: from animal models to human behavior. Neuron. 2005;48:175–87.
Bissière S, Plachta N, Hoyer D, McAllister KH, Olpe H-R, Grace AA, et al. The rostral anterior cingulate cortex modulates the efficiency of amygdala-dependent fear learning. Biol Psychiatry. 2008;63:821–31.
Tang W, Jbabdi S, Zhu Z, Cottaar M, Grisot G, Lehman JF, et al. A connectional hub in the rostral anterior cingulate cortex links areas of emotion and cognitive control. Elife. 2019;8:e43761.
Moreno-López L, Ioannidis K, Askelund AD, Smith AJ, Schueler K, van Harmelen A-L. The resilient emotional brain: a scoping review of the medial prefrontal cortex and limbic structure and function in resilient adults with a history of childhood maltreatment. Biol Psychiatry: Cognit Neurosci Neuroimaging. 2020;5:392–402.
McIntosh RC, Hoshi RA, Nomi J, Goodman Z, Kornfeld S, Vidot DC. I know why the caged bird sings: distress tolerant individuals show greater resting state connectivity between ventromedial prefrontal cortex and right amygdala as a function of higher vagal tone. Int J Psychophysiol. 2024;196:112274.
Dulka BN, Bress KS, Grizzell JA, Cooper MA. Social dominance modulates stress-induced neural activity in medial prefrontal cortex projections to the basolateral amygdala. Neuroscience. 2018;388:274–83.
Lamanna J, Isotti F, Ferro M, Spadini S, Racchetti G, Musazzi L, et al. Occlusion of dopamine-dependent synaptic plasticity in the prefrontal cortex mediates the expression of depressive-like behavior and is modulated by ketamine. Sci Rep. 2022;12:11055.
Maier SF. Behavioral control blunts reactions to contemporaneous and future adverse events: medial prefrontal cortex plasticity and a corticostriatal network. Neurobiol Stress. 2015;1:12–22.
Uchida M, Biederman J, Gabrieli JDE, Micco J, de Los Angeles C, Brown A, et al. Emotion regulation ability varies in relation to intrinsic functional brain architecture. Soc Cognit Affect Neurosci. 2015;10:1738–48.
Heilbronner SR, Haber SN. Frontal cortical and subcortical projections provide a basis for segmenting the cingulum bundle: implications for neuroimaging and psychiatric disorders. J Neurosci. 2014;34:10041–54.
Tsolaki E, Sheth SA, Pouratian N. Variability of white matter anatomy in the subcallosal cingulate area. Hum brain Mapp. 2021;42:2005–17.
Nozais V, Forkel SJ, Petit L, Talozzi L, Corbetta M, Thiebaut de Schotten M, et al. Atlasing white matter and grey matter joint contributions to resting-state networks in the human brain. Commun Biol. 2023;6:726.
Liakos F, Komaitis S, Drosos E, Neromyliotis E, Skandalakis GP, Gerogiannis AI, et al. The Topography of the frontal terminations of the uncinate fasciculus revisited through focused fiber dissections: shedding light on a current controversy and introducing the insular apex as a key anatomoclinical area. World Neurosurg. 2021;152:e625–e634.
Rolls ET. Emotion, motivation, decision-making, the orbitofrontal cortex, anterior cingulate cortex, and the amygdala. Brain Structure Funct. 2023;228:1201–57.
Ball G, Pazderova L, Chew A, Tusor N, Merchant N, Arichi T, et al. Thalamocortical connectivity predicts cognition in children born preterm. Cereb Cortex. 2015;25:4310–8.
Haney B, Reavey D, Atchison L, Poull J, Dryer L, Anderson B, et al. Magnetic resonance imaging studies without sedation in the neonatal intensive care unit: safe and efficient. J Perinat Neonatal Nurs. 2010;24:256–66.
Keil B, Alagappan V, Mareyam A, McNab JA, Fujimoto K, Tountcheva V, et al. Size-optimized 32-channel brain arrays for 3 T pediatric imaging. Magnetic Reson Med. 2011;66:1777–87.
Williams L-A, DeVito TJ, Winter JD, Orr TN, Thompson RT, Gelman N. Optimization of 3D MP-RAGE for neonatal brain imaging at 3.0 T. Magnetic Reson Imaging. 2007;25:1162–70.
Gao W, Lin W, Grewen K, Gilmore JH. Functional connectivity of the infant human brain plastic and modifiable. Neurosci. 2016;32:169–84.
Eggebrecht AT, Elison JT, Feczko E, Todorov A, Wolff JJ, Kandala S, et al. Joint attention and brain functional connectivity in infants and toddlers. Cereb Cortex. 2017;27:1709–20.
Croteau-Chonka EC, Dean III, DC, Remer J, Dirks H, O’Muircheartaigh J, et al. Examining the relationships between cortical maturation and white matter myelination throughout early childhood. Neuroimage. 2016;125:413–21.
Petrican R, Taylor MJ, Grady CL. Trajectories of brain system maturation from childhood to older adulthood: implications for lifespan cognitive functioning. Neuroimage. 2017;163:125–49.
Sheridan M, Kharitonova M, Martin RE, Chatterjee A, Gabrieli JD. Neural substrates of the development of cognitive control in children ages 5–10 years. J Cognit Neurosci. 2014;26:1840–50.
Todd RM, Lee W, Evans JW, Lewis MD, Taylor MJ. Withholding response in the face of a smile: age-related differences in prefrontal sensitivity to Nogo cues following happy and angry faces. Developmental Cognit Neurosci. 2012;2:340–50.
Zielinski BA, Gennatas ED, Zhou J, Seeley WW. Network-level structural covariance in the developing brain. Proc Natl Acad Sci. 2010;107:18191–6.
Wu K, Taki Y, Sato K, Hashizume H, Sassa Y, Takeuchi H, et al. Topological organization of functional brain networks in healthy children: differences in relation to age, sex, and intelligence. PLoS One. 2013;8:e55347.
Lin W, Zhu Q, Gao W, Chen Y, Toh CH, Styner M, et al. Functional connectivity MR imaging reveals cortical functional connectivity in the developing brain. AJNR Am J Neuroradiol. 2008;29:1883–9.
Liu W-C, Flax JF, Guise KG, Sukul V, Benasich AA. Functional connectivity of the sensorimotor area in naturally sleeping infants. Brain Res. 2008;1223:42–49.
Deoni SC, Dean III, DC, Remer J, Dirks H, O’Muircheartaigh J. Cortical maturation and myelination in healthy toddlers and young children. Neuroimage. 2015;115:147–61.
Muetzel RL, Blanken LM, Thijssen S, van der Lugt A, Jaddoe VW, Verhulst FC, et al. Resting-state networks in 6-to-10 year old children. Hum brain Mapp. 2016;37:4286–4300.
Gabard-Durnam L, O’Muircheartaigh J, Dirks H, Dean IIID, Tottenham N, Deoni S. Human amygdala functional network development: a cross-sectional study from 3 months to 5 years of age. Developmental Cognit Neurosci. 2018;34:63–74.
Richardson H, Lisandrelli G, Riobueno-Naylor A, Saxe R. Development of the social brain from age three to twelve years. Nat Commun. 2018;9:1–12.
Knickmeyer RC, Gouttard S, Kang C, Evans D, Wilber K, Smith JK, et al. A structural MRI study of human brain development from birth to 2 years. J Neurosci : Off J Soc Neurosci. 2008;28:12176–82.
Gilmore J, Langworthy B, Girault JB, Fine J, Jha SC, Kim SH, et al. Individual variation of human cortical structure is established in the first year of life. Biol Psychiatry: Cognit Neurosci Neuroimaging. 2020;5:971–80.
Lynch KM, Shi Y, Toga AW, Clark KA, Pediatric Imaging N, Study G. Hippocampal shape maturation in childhood and adolescence. Cereb Cortex. 2019;29:3651–65.
Nie J, Li G, Shen D. Development of cortical anatomical properties from early childhood to early adulthood. Neuroimage. 2013;76:216–24.
Fjell AM, Walhovd KB, Brown TT, Kuperman JM, Chung Y, Hagler DJ, et al. Multimodal imaging of the self-regulating developing brain. Proc Natl Acad Sci. 2012;109:19620–5.
Argyropoulou MI, Xydis VG, Astrakas LG. Functional connectivity of the pediatric brain. Neuroradiology. 2024;66:2071–82.
Zhang H, Shen D, Lin W. Resting-state functional MRI studies on infant brains: a decade of gap-filling efforts. NeuroImage. 2019;185:664–84.
Liu J, Chen H, Cornea E, Gilmore JH, Gao W. Longitudinal developmental trajectories of functional connectivity reveal regional distribution of distinct age effects in infancy. Cereb Cortex. 2023;33:10367–79.
Gao W, Gilmore JH, Shen D, Smith JK, Zhu H, Lin W. The synchronization within and interaction between the default and dorsal attention networks in early infancy. Cereb Cortex. 2012;23:594–603.
Fair DA, Cohen AL, Dosenbach NUF, Church JA, Miezin FM, Barch DM, et al. The maturing architecture of the brain’s default network. Proc Natl Acad Sci. 2008;105:4028–32.
Rebello K, Moura LM, Pinaya WHL, Rohde LA, Sato JR. Default mode network maturation and environmental adversities during childhood. Chronic Stress. 2018;2:2470547018808295.
Gao W, Lin W, Grewen K, Gilmore JH. Functional connectivity of the infant human brain: plastic and modifiable. Neuroscientist. 2016;23:169–84.
Uddin LQ, Supekar KS, Ryali S, Menon V. Dynamic reconfiguration of structural and functional connectivity across core neurocognitive brain networks with development. J Neurosci. 2011;31:18578–89.
Ryali S, Supekar K, Chen T, Kochalka J, Cai W, Nicholas J, et al. Temporal dynamics and developmental maturation of salience, default and central-executive network interactions revealed by variational bayes hidden markov modeling. PLoS Comput Biol. 2016;12:e1005138.
Cascio CJ, Gerig G, Piven J. Diffusion tensor imaging: application to the study of the developing brain. J Am Acad Child Adolesc Psychiatry. 2007;46:213–23.
Gao W, Lin W, Chen Y, Gerig G, Smith JK, Jewells V, et al. Temporal and spatial development of axonal maturation and myelination of white matter in the developing brain. AJNR Am J Neuroradiol. 2009;30:290–6.
Provenzale JM, Liang L, DeLong D, White LE. Diffusion tensor imaging assessment of brain white matter maturation during the first postnatal year. Am J Roentgenology. 2007;189:476–86.
Geng X, Gouttard S, Sharma A, Gu H, Styner M, Lin W, et al. Quantitative tract-based white matter development from birth to age 2 years. Neuroimage. 2012;61:542–57.
Akazawa K, Chang L, Yamakawa R, Hayama S, Buchthal S, Alicata D, et al. Probabilistic maps of the white matter tracts with known associated functions on the neonatal brain atlas: application to evaluate longitudinal developmental trajectories in term-born and preterm-born infants. Neuroimage. 2016;128:167–79.
Reynolds JE, Grohs MN, Dewey D, Lebel C. Global and regional white matter development in early childhood. Neuroimage. 2019;196:49–58.
Lebel C, Treit S, Beaulieu C. A review of diffusion MRI of typical white matter development from early childhood to young adulthood. NMR Biomed. 2019;32:e3778.
Haynes L, Ip A, Cho IY, Dimond D, Rohr CS, Bagshawe M, et al. Grey and white matter volumes in early childhood: A comparison of voxel-based morphometry pipelines. Developmental Cognit Neurosci. 2020;46:100875.
Barnea-Goraly N, Menon V, Eckert M, Tamm L, Bammer R, Karchemskiy A, et al. White matter development during childhood and adolescence: a cross-sectional diffusion tensor imaging study. Cereb Cortex. 2005;15:1848–54.
Lebel C, Gee M, Camicioli R, Wieler M, Martin W, Beaulieu C. Diffusion tensor imaging of white matter tract evolution over the lifespan. Neuroimage. 2012;60:340–52.
Grotheer M, Rosenke M, Wu H, Kular H, Querdasi FR, Natu VS, et al. White matter myelination during early infancy is linked to spatial gradients and myelin content at birth. Nat Commun. 2022;13:997.
Pines AR, Sacchet MD, Kullar M, Ma J, Williams LM. Multi-unit relations among neural, self-report, and behavioral correlates of emotion regulation in comorbid depression and obesity. Sci Rep. 2018;8:14032.
Bathelt J, Johnson A, Zhang M, Astle DE. The cingulum as a marker of individual differences in neurocognitive development. Sci Rep. 2019;9:2281.
Conte S, Zimmerman D, Richards JE. White matter trajectories over the lifespan. PLOS ONE. 2024;19:e0301520.
Gill BK, Balodis IM, Minuzzi L, Shah N, Boylan K. Neural correlates of emotion dysregulation in adolescents: a systematic review. Discov Psychol. 2024;4:31.
Sylvester CM, Smyser CD, Smyser T, Kenley J, Ackerman JJ, Shimony JS, et al. Cortical functional connectivity evident after birth and behavioral inhibition at age 2. Am J Psychiatry. 2017;175:180–7.
Graham AM, Buss C, Rasmussen JM, Rudolph MD, Demeter DV, Gilmore JH, et al. Implications of newborn amygdala connectivity for fear and cognitive development at 6-months-of-age. Developmental Cognit Neurosci. 2016;18:12–25.
Alcauter S, Lin W, Smith JK, Short SJ, Goldman BD, Reznick JS, et al. Development of thalamocortical connectivity during infancy and its cognitive correlations. J Neurosci. 2014;34:9067–75.
Short SJ, Elison JT, Goldman BD, Styner M, Gu H, Connelly M, et al. Associations between white matter microstructure and infants’ working memory. Neuroimage. 2013;64:156–66.
Ramenghi LA, Rutherford M, Fumagalli M, Bassi L, Messner H, Counsell S, et al. Neonatal neuroimaging: going beyond the pictures. Early Hum Dev. 2009;85:S75–77.
Filippi CA, Ravi S, Bracy M, Winkler A, Sylvester CM, Pine DS, et al. Amygdala functional connectivity and negative reactive temperament at age 4 months. J Am Acad Child Adolesc Psychiatry. 2021;60:1137–46.
Thomas E, Buss C, Rasmussen JM, Entringer S, Ramirez JS, Marr M, et al. Newborn amygdala connectivity and early emerging fear. Developmental Cognit Neurosci. 2019;37:100604.
Ravi S, Catalina Camacho M, Fleming B, Scudder MR, Humphreys KL. Concurrent and prospective associations between infant frontoparietal and default mode network connectivity and negative affectivity. Biol Psychol. 2023;184:108717.
Filippi CA, Winkler AM, Kanel D, Elison JT, Hardiman H, Sylvester C, et al. Neural correlates of novelty-evoked distress in 4-month-old infants: a Synthetic Cohort Study. Biol Psychiatry: Cognit Neurosci Neuroimaging. 2024;9:905–14.
Kim H, Cho M, Shim W, Kim J, Jeon E, Kim D, et al. Deficient autophagy in microglia impairs synaptic pruning and causes social behavioral defects. Mol Psychiatry. 2017;22:1576–84.
Banihashemi L, Bertocci MA, Alkhars HM, Versace A, Northrup JB, Lee VK, et al. Limbic white matter structural integrity at 3 months prospectively predicts negative emotionality in 9-month-old infants: a preliminary study. J Affect Disord. 2020;273:538–41.
Planalp EM, Dowe KN, Alexander AL, Goldsmith HH, Davidson RJ, Dean DC. White matter microstructure predicts individual differences in infant fear (But not anger and sadness). Developmental Sci. 2023;26:e13340.
Stark EA, Cabral J, Riem MME, Van Ijzendoorn MH, Stein A, Kringelbach ML. The power of smiling: the adult brain networks underlying learned infant emotionality. Cereb Cortex. 2020;30:2019–29.
Zhao X, Shi J, Dai F, Wei L, Zhang B, Yu X, et al. Brain development from newborn to adolescence: evaluation by neurite orientation dispersion and density imaging. Front Hum Neurosci. 2021;15:616132.
DiPiero M, Rodrigues PG, Gromala A, Dean DC. Applications of advanced diffusion MRI in early brain development: a comprehensive review. Brain Structure Funct. 2023;228:367–92.
Peters J, Büchel C. Neural representations of subjective reward value. Behavioural Brain Res. 2010;213:135–41.
Hocker DL, Brody CD, Savin C, Constantinople CM. Subpopulations of neurons in lOFC encode previous and current rewards at time of choice. eLife. 2021;10:e70129.
Zhang Y, Banihashemi L, Versace A, Samolyk A, Taylor M, English G, et al. Early infant prefrontal cortical microstructure predicts present and future emotionality. Biol Psychiatry. 2024;96:959–70.
Zhang Y, Banihashemi L, Versace A, Samolyk A, Abdelkader M, Taylor M, et al. Associations among white matter microstructural changes and the development of emotional reactivity and regulation in infancy. Mol Psychiatry. 2025;30:3168–3174.
Zhang Y, Banihashemi L, Samolyk A, Taylor M, English G, Schmithorst VJ, et al. Early infant prefrontal gray matter volume is associated with concurrent and future infant emotionality. Transl Psychiatry. 2023;13:125.
Bezanson S, Nichols ES, Duerden EG. Postnatal maternal distress, infant subcortical brain macrostructure and emotional regulation. Psychiatry Res Neuroimaging. 2023;328:111577.
Eden AS, Schreiber J, Anwander A, Keuper K, Laeger I, Zwanzger P, et al. Emotion regulation and trait anxiety are predicted by the microstructure of fibers between amygdala and prefrontal cortex. J Neurosci. 2015;35:6020.
d’Arbeloff TC, Kim MJ, Knodt AR, Radtke SR, Brigidi BD, Hariri AR. Microstructural integrity of a pathway connecting the prefrontal cortex and amygdala moderates the association between cognitive reappraisal and negative emotions. Emotion. 2018;18:912.
Doyle CM, Lasch C, Elison JT. Emerging evidence for putative neural networks and antecedents of pediatric anxiety in the fetal, neonatal, and infant periods. Biol Psychiatry. 2021;89:672–80.
Smith CL, Diaz A, Day KL, Bell MA. Infant frontal electroencephalogram asymmetry and negative emotional reactivity as predictors of toddlerhood effortful control. J Exp Child Psychol. 2016;142:262–73.
Wee CY, Tuan TA, Broekman BF, Ong MY, Chong YS, Kwek K, et al. Neonatal neural networks predict children behavioral profiles later in life. Hum Brain Mapp. 2017;38:1362–73.
Rogers CE, Sylvester CM, Mintz C, Kenley JK, Shimony JS, Barch DM, et al. Neonatal amygdala functional connectivity at rest in healthy and preterm infants and early internalizing symptoms. J Am Acad Child Adolesc Psychiatry. 2017;56:157–66.
Ramphal B, Whalen DJ, Kenley JK, Yu Q, Smyser CD, Rogers CE, et al. Brain connectivity and socioeconomic status at birth and externalizing symptoms at age 2 years. Developmental Cognit Neurosci. 2020;45:100811.
Salzwedel AP, Stephens RL, Goldman BD, Lin W, Gilmore JH, Gao W. Development of amygdala functional connectivity during infancy and its relationship with 4-year behavioral outcomes. Biol Psychiatry: Cognit Neurosci Neuroimaging. 2019;4:62–71.
Nolvi S, Tuulari JJ, Pelto J, Bridgett DJ, Eskola E, Lehtola SJ, et al. Neonatal amygdala volumes and the development of self-regulation from early infancy to toddlerhood. Neuropsychology. 2021;35:285.
Ghassabian A, Herba CM, Roza SJ, Govaert P, Schenk JJ, Jaddoe VW, et al. Infant brain structures, executive function, and attention deficit/hyperactivity problems at preschool age. A prospective study. J Child Psychol Psychiatry. 2013;54:96–104.
Calkins SD, Dollar JM, Wideman L. Temperamental vulnerability to emotion dysregulation and risk for mental and physical health challenges. Dev Psychopathology. 2019;31:957–70.
Meaney MJ. Perinatal maternal depressive symptoms as an issue for population health. Am J Psychiatry. 2018;175:1084–93.
Maughan A, Cicchetti D, Toth SL, Rogosch FA. Early-occurring maternal depression and maternal negativity in predicting young Children’s emotion regulation and socioemotional difficulties. J Abnorm Child Psychol. 2007;35:685–703.
Feldman R, Granat A, Pariente C, Kanety H, Kuint J, Gilboa-Schechtman E. Maternal depression and anxiety across the postpartum year and infant social engagement, fear regulation, and stress reactivity. J Am Acad Child Adolesc Psychiatry. 2009;48:919–27.
Slomian J, Honvo G, Emonts P, Reginster J-Y, Bruyère O. Consequences of maternal postpartum depression: A systematic review of maternal and infant outcomes. Women’s Health. 2019;15:1745506519844044.
Goodman SH, Rouse MH, Connell AM, Broth MR, Hall CM, Heyward D. Maternal depression and child psychopathology: a meta-analytic review. Clin Child Family Psychol Rev. 2011;14:1–27.
Duan C, Hare MM, Staring M, Deligiannidis KM. Examining the relationship between perinatal depression and neurodevelopment in infants and children through structural and functional neuroimaging research. Int Rev Psychiatry. 2019;31:264–79.
Rifkin-Graboi A, Meaney MJ, Chen H, Bai J, Hameed WB, Tint MT, et al. Antenatal maternal anxiety predicts variations in neural structures implicated in anxiety disorders in newborns. J Am Acad Child Adolesc Psychiatry. 2015;54:313–321.e312.
Graham RM, Jiang L, McCorkle G, Bellando BJ, Sorensen ST, Glasier CM, et al. Maternal anxiety and depression during late pregnancy and newborn brain white matter development. Am J Neuroradiology. 2020;41:1908–15.
Rifkin-Graboi A, Bai J, Chen H, Hameed WB, Sim LW, et al. Prenatal maternal depression associates with microstructure of right amygdala in neonates at birth. Biol Psychiatry. 2013;74:837–44.
Lehtola SJ, Tuulari JJ, Scheinin NM, Karlsson L, Parkkola R, Merisaari H, et al. Newborn amygdalar volumes are associated with maternal prenatal psychological distress in a sex-dependent way. NeuroImage: Clin. 2020;28:102380.
Lee A, Poh JS, Wen DJ, Guillaume B, Chong Y-S, Shek LP, et al. Long-term influences of prenatal maternal depressive symptoms on the amygdala–prefrontal circuitry of the offspring from birth to early childhood. Biol Psychiatry: Cognit Neurosci Neuroimaging. 2019;4:940–7.
Qiu A, Anh TT, Li Y, Chen H, Rifkin-Graboi A, Broekman BFP, et al. Prenatal maternal depression alters amygdala functional connectivity in 6-month-old infants. Transl Psychiatry. 2015;5:e508.
Burghy CA, Stodola DE, Ruttle PL, Molloy EK, Armstrong JM, Oler JA, et al. Developmental pathways to amygdala-prefrontal function and internalizing symptoms in adolescence. Nat Neurosci. 2012;15:1736–41.
Erk S, Mikschl A, Stier S, Ciaramidaro A, Gapp V, Weber B, et al. Acute and sustained effects of cognitive emotion regulation in major depression. J Neurosci. 2010;30:15726.
Uddin LQ, Yeo BTT, Spreng RN. Towards a universal taxonomy of macro-scale functional human brain networks. Brain Topogr. 2019;32:926–42.
Nolvi S, Tuulari JJ, Lavonius T, Scheinin NM, Lehtola SJ, Lavonius M, et al. Newborn white matter microstructure moderates the association between maternal postpartum depressive symptoms and infant negative reactivity. Soc Cognit Affect Neurosci. 2020;15:nsaa081.
Dufford AJ, Salzwedel AP, Gilmore JH, Gao W, Kim P. Maternal trait anxiety symptoms, frontolimbic resting‐state functional connectivity, and cognitive development in infancy. Developmental Psychobiology. 2021;63:e22166.
Hendrix CL, Dilks DD, McKenna BG, Dunlop AL, Corwin EJ, Brennan PA. Maternal childhood adversity associates with frontoamygdala connectivity in neonates. Biol Psychiatry: Cognit Neurosci Neuroimaging. 2021;6:470–8.
Hanson JL, Hair N, Shen DG, Shi F, Gilmore JH, Wolfe BL, et al. Family poverty affects the rate of human infant brain growth. PLoS ONE. 2013;8:e80954.
Brady RG, Rogers CE, Prochaska T, Kaplan S, Lean RE, Smyser TA, et al. The effects of prenatal exposure to neighborhood crime on neonatal functional connectivity. Biol Psychiatry. 2022;92:139–48.
Jansen AG, Mous SE, White T, Posthuma D, Polderman TJC. What twin studies tell us about the heritability of brain development, morphology, and function: a review. Neuropsychol Rev. 2015;25:27–46.
Kremen WS, Fennema‐Notestine C, Eyler LT, Panizzon MS, Chen CH, Franz CE, et al. Genetics of brain structure: contributions from the vietnam era twin study of aging. Am J Méd Genet Part B: Neuropsychiatr Genet. 2013;162:751–61.
Sudre G, Choudhuri S, Szekely E, Bonner T, Goduni E, Sharp W, et al. Estimating the heritability of structural and functional brain connectivity in families affected by Attention-Deficit/Hyperactivity disorder. JAMA Psychiatry. 2016;74:76.
Teeuw J, Brouwer RM, Guimarães JPOFT, Brandner P, Koenis MMG, et al. Genetic and environmental influences on functional connectivity within and between canonical cortical resting-state networks throughout adolescent development in boys and girls. NeuroImage. 2019;202:116073.
Posner J, Cha J, Wang Z, Talati A, Warner V, Gerber A, et al. Increased default mode network connectivity in individuals at high familial risk for depression. Neuropsychopharmacology. 2016;41:1759–67.
Swain JE, Lorberbaum JP, Kose S, Strathearn L. Brain basis of early parent–infant interactions: psychology, physiology, and in vivo functional neuroimaging studies. J Child Psychol Psychiatry. 2007;48:262–87.
Swain JE. The human parental brain: in vivo neuroimaging. Prog Neuro-psychopharmacol Biol Psychiatry. 2011;35:1242–54.
Swain JE. Baby stimuli and the parent brain: functional neuroimaging of the neural substrates of parent-infant attachment. Psychiatry. 2008;5:28.
Nishimura Y, Kanakogi Y, Myowa-Yamakoshi M. Infants’ emotional states influence maternal behaviors during holding. Infant Behav Dev. 2016;43:66–74.
Wu Q, Feng X. Infant emotion regulation and cortisol response during the first 2 years of life: association with maternal parenting profiles. Developmental Psychobiology. 2020;62:1076–91.
Hu S, Pruessner JC, Coupé P, Collins DL. Volumetric analysis of medial temporal lobe structures in brain development from childhood to adolescence. Neuroimage. 2013;74:276–87.
Hay RE, Reynolds JE, Grohs MN, Paniukov D, Giesbrecht GF, Letourneau N, et al. Amygdala-prefrontal structural connectivity mediates the relationship between prenatal depression and behavior in preschool boys. J Neurosci. 2020;40:6969–77.
Albaugh MD, Nguyen T-V, Ducharme S, Collins DL, Botteron KN, D’Alberto N, et al. Age-related volumetric change of limbic structures and subclinical anxious/depressed symptomatology in typically developing children and adolescents. Biol Psychol. 2017;124:133–40.
Lichtin RD, Merz EC, He X, Desai PM, Simon KR, Melvin SA, et al. Material hardship, prefrontal cortex–amygdala structure, and internalizing symptoms in children. Developmental Psychobiology. 2021;63:364–77.
Fox NA, Henderson HA, Rubin KH, Calkins SD, Schmidt LA. Continuity and discontinuity of behavioral inhibition and exuberance: psychophysiological and behavioral influences across the first four years of life. Child Dev. 2001;72:1–21.
Kagan J, Snidman N, Arcus D. Childhood derivatives of high and low reactivity in infancy. Child Dev. 1998;69:1483–93.
Snidman N, Kagan J, Riordan L, Shannon DC. Cardiac function and behavioral reactivity during infancy. Psychophysiology. 1995;32:199–207.
Huot RL, Brennan PA, Stowe ZN, Plotsky PM, Walker EF. Negative affect in offspring of depressed mothers is predicted by infant cortisol levels at 6 months and maternal depression during pregnancy, but not postpartum. Ann N Y Acad Sci. 2004;1032:234–6.
Rinaman L, Banihashemi L, Koehnle TJ. Early life experience shapes the functional organization of stress-responsive visceral circuits. Physiol Behav. 2011;104:632–40.
Banihashemi L, Peng CW, Verstynen T, Wallace ML, Lamont DN, Alkhars HM, et al. Opposing relationships of childhood threat and deprivation with stria terminalis white matter. Hum Brain Mapp. 2021;42:2445–60.
Acknowledgements
This project was supported by the National Institutes of Health (R01MH115466, PIs: Drs. Mary L. Phillips and Alison E. Hipwell and The Pittsburgh Foundation, PI: Dr. Mary L. Phillips. This research was also supported in part by the University of Pittsburgh Center for Research Computing, RRID:SCR_022735, through the resources provided. Specifically, this work used the HTC cluster, which is supported by NIH award number S10OD028483.
Author information
Authors and Affiliations
Contributions
MLP (Conceptualization, writing—original draft, review and editing); AEH (Conceptualization, writing—original draft, review and editing); LB (Figures, writing—additional sections, review and editing); YZ (Table, writing—additional sections, review and editing). All authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Banihashemi, L., Zhang, Y., Hipwell, A.E. et al. The promise of infant MRI in psychiatry: toward a framework for neural network measures in early emotional and behavioral risk identification and new intervention targets. Mol Psychiatry 31, 444–455 (2026). https://doi.org/10.1038/s41380-025-03197-5
Received:
Revised:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41380-025-03197-5
