
Abstract
Investigating similarities and differences between alcohol use disorder (AUD) and obesity is important because both AUD and obesity are public health concerns and share neurobiological and periphery-brain mechanisms. Furthermore, AUD and obesity often present with similar medical consequences related to organ damage, including liver and cardiovascular diseases. There is also growing evidence of changes in alcohol drinking in people who undergo bariatric surgery for obesity. In this non-systematic critical review, we identified relevant articles through PubMed searches, previous knowledge, and recursive reference searching. A librarian also used PubMed and Google Scholar for additional relevant articles, using terms such as alcohol, metabolic disorders, obesity, glucagon-like peptide-1 (GLP-1), bariatric surgery, and gut-brain axis. We provide an overview of the neurobiological, pathophysiological, neuroimaging, and clinical features related to the overlap and crosstalk between AUD and obesity. We also provide a summary of the currently approved medications for obesity and those for AUD and note the potential for some of these medications to work for both disorders. Specific to the latter point, we place emphasis on GLP-1 therapies, given their recent approval for weight loss and the growing evidence suggesting their potential efficacy for AUD and other addictions. We further review studies of the relationship between bariatric surgery and AUD and discuss potential mechanisms and future directions. In summary, studying the overlap between obesity and AUD may shed light on the mechanisms underlying the development and maintenance of both diseases. This knowledge, in turn, may help identify new therapeutic targets for AUD, and possibly comorbid obesity and/or other metabolic disorders.
Similar content being viewed by others

Introduction
Approximately 42% of adults in the United States have obesity [1], and 10% meet criteria for alcohol use disorder (AUD) [2]. An estimated 8 million adults (approximately 3% of the United States adult population) have obesity and drink alcohol heavily [3]. This comorbidity is particularly alarming given that AUD and obesity independently contribute to a host of comorbidities, including cardiovascular disease, diabetes, liver disease, and cancer [4, 5]. Although on the surface, both diseases seem to arise from ‘over-consumption’ – either of food or alcohol – their etiologies are far more complex than simply eating or drinking too much. Obesity, defined as a body mass index (BMI) greater than 30 kg/m2, results from the storage of excess energy in the form of adipose tissue [6]. How this excess energy comes to be stored by an individual, however, is not a simple matter, reflecting a multitude of biological, psychological, and environmental factors that predispose to or protect from obesity. AUD, in contrast, results from “a problematic pattern of alcohol use leading to clinically significant impairment or distress”, as defined by the DSM-5 [7]. As with obesity, how a pattern of alcohol use becomes problematic also reflects a complex interaction of biological, psychological, and environmental factors, which predispose to or protect from AUD.
Critically, research has shown remarkable overlap in the biological, psychological, and environmental determinants of AUD and obesity. AUD and obesity not only share overlapping neurocircuitry within the brain’s reward and appetitive pathways, but are also influenced by similar neuroendocrine and neuroimmune signaling pathways. As a result, AUD and obesity often respond similarly to treatments that target their common pathophysiologies, and interventions for one disease can often have consequences for the other. The goal of this narrative critical review is thus to investigate the similarities and differences in AUD and obesity’s causes and sequelae, with an eye toward identifying novel interventions that reduce the consequences of these two diseases for personal and public health.
We identified relevant articles through PubMed searches, previous knowledge, and recursive reference searching. A librarian also used PubMed and Google Scholar for additional relevant articles, using terms such as alcohol, metabolic disorders, obesity, glucagon-like peptide-1 (GLP-1), bariatric surgery, and gut-brain axis. Our search did not include specific inclusion/exclusion criteria, nor did we apply timeframe constraints. While this approach might not be free from potential bias, we envisioned this narrative overview of the literature as a Critical Review, that identifies dominant, overlapping themes of AUD and obesity.
Environmental and genetic contributors to AUD and obesity
Owing largely to advancements in neuroscience and the development of effective biologically-based treatments (particularly pharmacotherapies), recent decades have seen remarkable changes in public opinion regarding the etiology of AUD and obesity [8,9,10]. Although significant stigma remains around both diseases [11,12,13], there has been a steady shift away from attributing AUD and obesity exclusively to individual failings of willpower or gluttony toward an understanding that both diseases arise from the multifactorial influence of genetics, environment, neurobiology, and culture.
The past two decades have witnessed an explosion of research on the genetics of obesity and AUD, with both shown to have substantial genetic heritability. The heritability of obesity is approximately 40–70%, with numerous genetic loci identified by genome-wide association studies [14]. Likewise, AUD is heritable (approximately 50%), although with a smaller number of loci as-yet identified [15, 16]. Although some epidemiological studies have provided support for a link between family history of alcohol problems and obesity, especially in women [17], the genetic correlation of AUD and obesity has been reported to be near zero [18, 19]. This is surprising given their overlapping pathophysiologies. More recently, evidence has emerged that the two diseases may have substantial genetic overlap, but with many of their shared gene variants having discordant (opposite) effects. For example, while the polygenic overlap of AUD and BMI is approximately 81%, only 48% of their shared loci have concordant effect directions [20, 21]. This finding may be critical to future efforts at identifying which obesity pharmacotherapies may be repurposed for AUD and vice versa.
As with genetics, significant progress has been made toward understanding how environmental and sociocultural factors influence AUD and obesity, although attention to specific environmental factors often differs between the two diseases. For example, the past decade has seen significant changes in the awareness that ‘built environment’ and access to nutritious foods (e.g., ‘food deserts’) greatly influence the development of obesity [22,23,24], but relatively less attention has been paid to evidence that access and availability of alcohol promotes problematic drinking [25, 26]. Furthermore, it is well-established that social factors and social networks play a role in both AUD and obesity; in fact, they influence alcohol consumption [27], and have effects that may be either obesogenic or protective [28,29,30]. More broadly, it is important to recognize that in addition to genetic and biological factors, social determinants of health, including socioeconomic status, employment, education, housing, and trauma history play a critical role in the development and maintenance of AUD and obesity. Therefore, prevention of chronic diseases like AUD and obesity should include the development of more and better community and national programs and policies aimed at addressing the social determinants of health that affect AUD and obesity and the associated medical consequences. This requires a comprehensive approach that includes not only biological but also social, environmental, and psychological factors [31, 32].
Notwithstanding their uneven application across obesity and AUD, we are encouraged by increased awareness that sociocultural factors considerably influence the development and course of both diseases. The present review, however, does not cover environmental, sociocultural, or genetic factors contributing to AUD and obesity, nor does it discuss related clinical phenotypes, including eating disorders, or the debate about whether obesity results from ‘food addiction’ [33, 34]. Rather, our focus is on how AUD and obesity’s shared neurobiological mechanisms, clinical features, and responses to extant treatments may inform our understanding of these two diseases, in the hope of highlighting new research opportunities and therapeutic targets that may ultimately lessen the burden of AUD and obesity.
Clinical consequences of AUD and obesity
Both AUD and obesity lead to several medical consequences. AUD is not solely a mental health but a “whole body” disorder, playing a causal role in more than 200 diseases, injuries, and other health conditions [35, 36]. Similarly, obesity plays a causal role in the development of several diseases [37]. One might postulate a simple model whereby the shared negative health outcomes of AUD and obesity result from an increase in caloric intake with heavy alcohol consumption which then promotes the development of obesity. This is unlikely, however, as experimental and epidemiological findings suggest that excess energy from alcohol has a relatively greater effect on non-daily alcohol drinkers than in daily heavy drinkers [38].
AUD and obesity are responsible for significant organ damage and consequent medical complications, including (but not limited to) on the liver, gut, pancreas, nervous system, cardiovascular system, endocrine system, and immune function, plus increased risk for other medical conditions like cancers [36, 39]. The mechanisms underlying these interactions are complex and not fully understood; some are distinct to AUD or obesity, while others overlap. Some alcohol-associated organ damage is related to the direct toxic effects of alcohol on cells, tissues, and organs, while some obesity-related medical consequences are attributed to increased fat mass and/or increased release of peptides from enlarged fat cells [37]. By contrast, inflammation, and especially low-grade inflammation, as well as dysregulated immunity (e.g., altered leucocyte counts as well as cell-mediated immune responses), is shared between AUD and obesity, contributing at multiple levels to their related medical complications, but also representing potential treatment targets [40, 41]. For comprehensive reviews on mechanisms and signaling pathways related to these complications, see e.g. [37, 39] for obesity and [36] for AUD. Some of the medical consequences linked to AUD or obesity arise from their comorbidities, either directly or indirectly. For example, both AUD and obesity are directly responsible for hypertension and cardiomyopathies, with hypertension potentially further exacerbating cardiomyopathies.
Clinically, there are some distinct features in the management of patients with AUD and those with obesity. Obesity-related medical consequences are typically chronic in nature and acute manifestations are secondary to the underlying organ damage. For example, obesity leads to cardiovascular disease (CVD), which in turn may lead to acute myocardial infarction (MI). Another example is that obesity leads to diabetes which in turn may lead to e.g., acute diabetic ketoacidosis. While these scenarios are also observed in people with AUD (AUD → CVD → MI; AUD → diabetes → acute diabetic ketoacidosis, etc.), AUD and alcohol drinking are also directly responsible for acute medical conditions, including after acute alcohol intoxication [42]. For example, while AUD chronically increases the risk of diabetes, acute alcohol intoxication (even in absence of AUD) may cause hypoglycemia. In addition to alcohol-related gastrointestinal chronic conditions like peptic ulcer disease and esophagitis, acute alcohol intoxication may lead to acute esophagitis and gastritis with severe nausea and vomiting and in some cases life-threating complications due to e.g., bleeding and Mallory-Weiss syndrome (tear or laceration of the gastroesophageal junction) [42]. These medical emergencies are often the result of an ‘acute on chronic’ worsening of alcohol-associated medical complications, e.g., acute on chronic gastritis, hepatitis, pancreatitis.
Given that AUD and obesity are common and frequently co-exist, the related medical consequences often arise from their additive or synergistic effects. For example, hypertension and cardiomyopathy are common in people with obesity and/or AUD; weight loss as well alcohol abstinence or even reduction are associated with improvement in cardiovascular comorbidities [37, 43, 44]. Importantly, patients with both AUD and obesity are even more likely to have comorbid cardiovascular complications due to the dual etiology.
Hepatic diseases offer another cogent example of the dual etiology of AUD and obesity in organ-related damage and consequences. People with alcohol-associated cirrhosis have a much lower life expectancy than the general population (on average 14 years shorter for men and 16 years shorter for women) [45]. Not only is alcohol a main cause of chronic liver disease and acute liver failure, but also with the development of new effective treatments for hepatitis C, alcohol has now become the leading cause of cirrhosis and the leading indication for liver transplantation in the United States [46]. Like alcohol use, obesity in the United States has also increased, leading to an increase in the previously named non-alcoholic fatty liver disease (NAFLD) [39, 47]. Several studies have shown accelerated liver disease in people with metabolic syndrome (central adiposity, diabetes, hypertension, and dyslipidemia) who also drink alcohol [48]. For example, a study from Northern Europe showed a ~ 7‐fold increase in incidence of severe liver disease in people with greater central adiposity who also consumed >1 and >2 alcohol-containing drinks/day for women and men, respectively [49]; such increase approached 20-fold in people with diabetes versus those without. In other words, not only are AUD and obesity two leading causes of liver diseases, but also their frequent comorbidity contributes to accelerated development and worsening of liver disease. The increased knowledge of overlapping biological processes that contribute to both NAFLD and alcohol-associated/related liver disease (ALD), coupled with the increased clinical and public relevance of this dual etiology, has led to a recent effort across multiple international liver societies to develop a new nomenclature. NAFLD has been replaced with metabolic dysfunction-associated steatotic liver disease (MASLD), and a new category, outside of pure MASLD, was created and named MetALD to describe those with MASLD who consume excessive amounts of alcohol (140 and 210 g/week for women and men, respectively) [50]. This new nomenclature acknowledges the common clinical feature of concomitant metabolic- and alcohol-related contributions to hepatic steatosis and subsequent liver inflammation, damage, and degeneration. This interdisciplinary approach may serve as a model for studying other medical conditions due to AUD and obesity comorbidity.
It is also key to consider the important role of sex differences in the development and clinical manifestations of both obesity and AUD, including differences in alcohol and food metabolism. In fact, not only sex differences exist in obesity and AUD per se, but also treatment response may vary as a function of sex. Furthermore, growing evidence shows sex differences in the medical consequences of these chronic diseases, including obesity-related conditions like obstructive sleep apnea and heart failure [51], and AUD-related conditions like ALD [46].
Neuroscientific knowledge and shared mechanisms of AUD and obesity
Both AUD and obesity are characterized by compulsive consummatory behaviors that persist despite adverse consequences, at least partially consistent with intersecting neurobiological mechanisms [33, 52]. Emerging research highlights common brain circuitries that contribute to these behaviors [33, 52]. Understanding the circuit-based mechanisms of AUD and obesity can provide valuable insights into their shared pathophysiology. This section will put special emphasis on GLP-1-related mechanisms in obesity and AUD, given the growing evidence of its important role in both disorders and the related treatment implications, as discussed later.
Mechanisms related to reward processing and dopamine-related pathways
Consumption of alcohol and energy-dense palatable foods increases the activity of dopamine neurons in ventral midbrain, leading to enhanced dopamine transmission in the nucleus accumbens (NAc) and other areas of the striatum. Dopamine-dependent and -independent signaling in the NAc and dorsal striatum regulate alcohol and palatable food consumption. Furthermore, prolonged alcohol drinking and weight gain resulting from overeating of energy-dense food are associated with deficits in mesostriatal dopamine transmission [53, 54], possibly involving mechanisms that originate in the gastrointestinal tract [55,56,57,58]. Positron emission tomography (PET) studies have established that striatal Dopamine D2 receptor (D2R) availability is decreased in individuals with AUD and obesity [59,60,61,62,63,64]. Disrupted D2R signaling in the striatum is thought to play an important role in AUD and diet-induced obesity. Indeed, the TaqIA allele of the DRD2/ANKK1 gene locus, which results in reduced striatal D2R expression, is associated with increased predisposition to AUD and obesity [65,66,67]. D2R gene knockout mice show increased aversion to alcohol, reduced alcohol consumption, and reduced sensitivity to the locomotor impairing effects of alcohol [68]. Furthermore, conditional deletion of D2Rs from striatal medium spiny neurons in mice reduces their sensitivity to the locomotor-impairing effects of alcohol and precipitates compulsive-like alcohol consumption, as reflected by drinking despite negative consequences [69]. In rats, RNA interference-mediated knockdown of D2Rs in the striatum similarly rendered consumption of palatable food similarly resistant to the appetite-suppressing effects of punished-associated conditioned stimuli [70]. These findings suggest that striatal D2R signaling contributes to the compulsive nature of both AUD and obesity. Notably, GLP-1 receptor agonists (GLP-1RAs) modulate dopamine signaling in the striatum and other brain regions [71,72,73,74,75,76,77,78,79,80,81,82,83], which is thought to contribute to their inhibitory effects on alcohol and food consumption.
Mechanisms related to stress, including hypothalamic and extrahypothalamic pathways
Stress plays an important role in the etiology and maintenance of both AUD and obesity [84,85,86]. The hypothalamic-pituitary-adrenal (HPA) axis mediates physiological and behavioral adaptations to prolonged stress, and chronic activation of this system can drive maladaptive behaviors, including over-consumption of alcohol and palatable food [87, 88]. In AUD, stress-induced activation of the HPA axis is thought to enhance alcohol craving and consumption in part to obtain the transient stress-alleviating actions of alcohol [89, 90]. Similarly, chronic stress is associated with overeating, particularly of palatable high-energy foods, a phenomenon sometimes referred to as “comfort eating” [91,92,93]. The central nucleus of the amygdala (CeA) is a core component of the extended amygdala and a critical node linking stress and reward pathways [94]. Dysregulated neural activity in the CeA is thought to contribute to stress-enhanced consumption of alcohol and palatable food through HPA-dependent and -independent mechanisms [95,96,97,98,99]. The bed nucleus of the stria terminalis (BNST) is another component of the extended amygdala heavily implicated in stress-related alcohol and palatable food consumption [100,101,102,103,104,105]. Neurons in the CeA and BNST express GLP-1Rs, and both structures are thought to regulate the effects of GLP-1RAs on the HPA axis and consummatory behaviors [106,107,108,109,110,111,112,113,114].
The hypothalamus is essential for maintaining energy homeostasis [115]. Major hypothalamic nuclei, including the arcuate nucleus, paraventricular nucleus, and lateral hypothalamus, integrate energy-related signals from peripheral hormones, including leptin, ghrelin, insulin, glucose-dependent insulinotropic polypeptide (GIP), and glucagon, to regulate feeding behavior and energy expenditure [115]. Weight gain resulting from the overconsumption of palatable food is associated with structural and functional adaptions in the hypothalamus, particularly in the lateral hypothalamus, which contribute to the development of compulsive-like food intake [116,117,118,119,120]. Alcohol drinking also affects hypothalamic function and can alter hunger and satiety signals to promote food consumption [121,122,123], while hunger-related hormones that act on the hypothalamus can stimulate alcohol craving and intake [122, 124]. The lateral hypothalamus also regulates alcohol seeking and consumption [125,126,127,128]. GLP-1R signaling in the lateral hypothalamus and other hypothalamic nuclei regulates alcohol and food consumption [129,130,131,132].
Mechanisms related to satiety and appetite
The nucleus of the solitary tract (NTS) and area postrema (AP) in the hindbrain contain transcriptionally heterogeneous populations of neurons that regulate satiety-related suppression of appetite, food avoidance, malaise, and nausea [133,134,135,136]. Diet-induced adaptations in NTS and AP neurons are thought to contribute to the development of overeating and obesity [133, 137,138,139]. Lesioning AP neurons increases alcohol consumption in rats through an unclear mechanism independent of alcohol-induced aversion or nausea [140]. Little is currently known about the role of NTS neurons in regulating alcohol consumption, although these cells regulate the consumption of other addictive drugs [141]. Notably, the NTS contains a population of preproglucagon-producing (PPG+) neurons that are the predominant or exclusive source of GLP-1 in the brain [142,143,144,145]. The NTS and AP also contain high concentrations of GLP-1Rs [146]. It was recently established that activation of GLP-1R-expressing (GLP1R+) neurons in the NTS triggered satiety and reduced food intake independent of aversion, while activation of GLP1R+ neurons in the AP elicited aversion accompanied by suppressed food intake. Little is currently known about the role of GLP1R+ neurons in the NTS and AP in regulating compulsive alcohol and food consumption.
In addition to PPG+ neurons in the NTS, GLP-1 is also produced by entero-endocrine cells in the gastrointestinal tract [147,148,149]. Peripheral GLP-1 is rapidly degraded by dipeptidylpeptidase-4 (DDP-4) and is unlikely to accumulate to levels sufficient to stimulate brain GLP-1R signaling [142,143,144,145]. However, peripheral GLP-1 can stimulate GLP-1R-expressing nodose (vagal) sensory neurons that innervate the gastrointestinal tract and hepatic portal vein and transmit feeding-related information to the NTS [150,151,152,153]. GLP1R+ nodose neurons play an important role in regulating meal termination and the appetite-suppressing effects of peripheral GLP-1 [154,155,156,157,158,159]. Impaired transmission of satiety-related information from the gastrointestinal tract to the NTS by nodose neurons may contribute to diet-induced obesity [160,161,162]. Nodose sensory neurons have also been implicated in the regulation of alcohol consumption [163], although few studies have directly investigated peripheral sensory mechanisms of alcohol reinforcement. Currently, it is unclear whether the suppression of alcohol and palatable food consumption by GLP-1RAs results from activation of GLP1R+ nodose neurons, stimulation of GLP1R+ neurons in the brain or a combination of both.
Prefrontal cortex mechanisms related to cognitive control/behavioral inhibition
In individuals with obesity and substance use disorders including AUD, increased prefrontal cortex (PFC) activity has been noted in responses to both food and drug cues respectively [164, 165]. Given this and the established role of the PFC in cognitive control and attention [166], it is notable that some studies have found evidence that both conditions are associated with decreased behavioral inhibition in the presence of either food- or alcohol-stimuli as well as altered PFC activity (particularly in the dorsolateral PFC and anterior cingulate cortex) while performing food- or alcohol-related cognitive control neuroimaging tasks (for review, see [167]). Regardless of whether this PFC dysfunction is a cause or consequence of either obesity or AUD, its implications for treatment adherence and persistence are clear: whether in weight loss for obesity or reducing alcohol consumption in the context of AUD, more cognitive control and behavioral inhibition is better than less. Cognitive and behavioral treatments that emphasize the development of cognitive control and behavioral inhibition have demonstrated efficacy in promoting weight loss and reducing alcohol consumption [168,169,170], and neuromodulation of the dorsolateral PFC via transcranial magnetic stimulation (TMS) has exhibited potential efficacy for both conditions as well [171, 172] (see neuromodulation section below).
Neuroimaging studies of AUD and obesity
There exist expansive human neuroimaging literature examining obesity and AUD. Although a thorough analysis of the similarities and differences among these literatures is beyond the scope of this review, a few general comments are worth making here. First, PET and functional magnetic resonance imaging (fMRI) studies using visual, olfactory, and gustatory stimuli overwhelmingly find that perception of food and alcohol cues activates overlapping distributed neural networks. These include striate and extrastriate visual areas to process visual features, gustatory and interoceptive regions in the insula to predict taste and the effects of food and alcohol on the body, and appetitive and reward-related regions in striatum and orbitofrontal cortex that represent the motivational salience and anticipated hedonic value of food and alcohol cues [164, 173,174,175,176,177].
Although neuroimaging studies have largely examined obesity and AUD phenotypes separately, their findings are generally overlapping and, within the limitations of neuroimaging’s spatial and temporal resolution, highly consistent with the preclinical studies reviewed above. For example, human PET and fMRI studies do not typically report engagement of sub-voxel-sized hypothalamic or brainstem sub-nuclei that powerfully impact food and alcohol consumption, but they do point to larger structures highlighted in preclinical studies. These include the ventral and dorsal striatum, anterior cingulate, orbitofrontal cortex, and amygdala, which typically exhibit greater activation to food or alcohol stimuli in people with obesity or AUD compared to controls [164, 173, 175, 178]. This presents an important question: if human neuroimaging is only able to measure a subset of the known circuitry underlying appetitive responses to food and alcohol, why bother? The answer is that it is challenging to reliably measure certain important phenomena in animals, such as the phenomenological experience of food or drug craving during voluntary abstinence [179,180,181]. Here, again, evidence suggests that the neural substrates of alcohol and food craving are largely overlapping, encompassing the ventromedial/orbitofrontal PFC, dorsal and subgenual anterior cingulate cortex, ventral striatum, parietal and temporal areas, cerebellum, and amygdala, with machine-learning classifier algorithms trained on one class of stimuli (e.g., food) reliably predicting craving for the other class (e.g., alcohol), and vice versa [182].
Treatments of AUD and obesity
Behavioral and pharmacological interventions, in addition to lifestyle modifications, constitute the mainstream treatment options to manage AUD as well as obesity.
For AUD, Alcoholics Anonymous and 12-Step facilitation treatments and other similar programs have historically played a key role in helping people maintain alcohol abstinence and reduce alcohol drinking [183]. Evidence-based behavioral treatments for AUD encompass brief interventions (including as part of the Screening, Brief Intervention, and Referral to Treatment (SBIRT) approach), motivational interviewing, cognitive behavioral therapies (CBT), contingency management, and mindfulness-based interventions [184,185,186]. Medications approved by the Food and Drug Administration (FDA) to treat AUD are naltrexone, acamprosate, and disulfiram. A few other medications have also shown efficacy and are sometimes used off-label for AUD, including topiramate, gabapentin, baclofen, and varenicline [186,187,188]. Optimal approach, albeit rarely implemented in clinical practice, would include pharmacotherapy plus an evidence-based behavioral therapy [189].
Many patients with AUD or other substance use disorders report an increase in food cravings and weight gain upon cessation of alcohol consumption and drug use [190,191,192]. The various accounts offered for this phenomenon generally fall into two camps [191]. One hypothesis asserts that individuals with substance use disorders have a strong neurobiological propensity for compulsive/addictive behaviors that include overeating, and that food motivation, which is often suppressed by the effects of alcohol and drugs, returns in force upon abstinence. An alternative ‘addiction transfer’ hypothesis asserts that following cessation of substance use, individuals may replace drug rewards with other substances (e.g., food) which stimulate similar neurobiological reward pathways. Although a full discussion of this topic is beyond the scope of this review, it is noteworthy that both accounts ground the phenomenon of increased food craving following cessation of alcohol or drug use in the shared neurobiological bases of obesity and alcohol and other substance use disorders.
The main treatment modalities for obesity considerably overlap with those used for AUD. Behavioral treatments for obesity are often delivered as part of multicomponent programs such as Weight Watchers and Diabetes Prevention Program and include goal setting, monitoring and adjustment of calorie intake and physical activity, stimulus control, stress management, and CBT [193,194,195]. FDA-approved medications for obesity include phentermine, phendimetrazine, and diethylpropion for short-term use and phentermine-topiramate, naltrexone-bupropion, orlistat, and more recently the GLP-1RAs liraglutide and semaglutide, and the GLP-1/GIP dual receptor agonist tirzepatide for long-term use. Other medications such as metformin, topiramate, and bupropion are also used off-label for weight loss [195, 196]. It is particularly intriguing that pharmacotherapies for AUD, e.g., naltrexone and topiramate, also reduce appetite and promote weight loss, a clinical observation consistent with the neurobiological overlap between AUD and obesity discussed before. Moreover, emerging research suggests that feeding-related and metabolic endocrine pathways such as ghrelin and GLP-1 could serve as promising pharmacotherapeutic targets for AUD [197].
The development, approval, and rapid clinical adoption of GLP-1RAs like semaglutide and tirzepatide has revolutionized the medical management of obesity and comorbid conditions such as diabetes, obstructive sleep apnea, liver and cardiovascular diseases [198, 199]. Following food intake, GLP-1 stimulates insulin secretion and inhibits glucagon release from the pancreas, hence regulating glucose homeostasis. Activation of GLP-1Rs also delays gastric emptying, reduces gastrointestinal motility, and overall suppresses appetite and food intake [200]. Compared to the first generation of GLP-1RAs (e.g., exenatide), newer GLP-1RAs (e.g., semaglutide) and poly agonists (e.g., tirzepatide, retatrutide) have higher affinity for the GLP-1R, longer half-lives, and are more potent, leading to high efficacy for glucose control and weight reduction [201,202,203].
Growing evidence suggests that GLP-1RAs also reduce alcohol use and may be repurposed to treat AUD. Preclinical experiments across different species, using various models and conducted by independent laboratories, show that central or peripheral administration of GLP-1RAs reduce alcohol intake and other related outcomes such as alcohol-induced conditioned place preference [197, 204, 205]. While initial studies were done with first-generation GLP-1RAs, more recent work has consistently found a robust effect with semaglutide reducing binge-like alcohol drinking in mice [110], operant self-administration, alcohol intake, and relapse-like drinking in rats [72, 110], and alcohol consumption in alcohol-preferring vervet monkeys [206]. In addition to their impact on appetite and consummatory behaviors [207, 208], several mechanisms have been proposed to mediate GLP-1RAs’ effects on alcohol intake, including interactions with reward-related pathways [209,210,211,212], stress [213,214,215], cognition and neuroprotection [216,217,218], pain and aversion [113, 219, 220], and inflammation [221,222,223].
Clinical work on the potential efficacy of GLP-1RAs for AUD is under way. In a 26-week clinical trial, exenatide 2 mg/week had no significant effect on the primary outcome of alcohol drinking in the full sample (dropout rate: 54.3%), while it reduced alcohol cue-reactivity (fMRI) and dopamine transporter availability (SPECT) in the brain. Exploratory analyses found a significant reduction in alcohol intake among exenatide-treated participants who had a BMI > 30 kg/m2, whereas an opposite effect was found among those with a BMI < 25 kg/m2 [224]. In a secondary analysis of a 12-week smoking cessation trial, dulaglutide 1.5 mg/week significantly reduced weekly alcohol consumption (most participants had a BMI > 29.9 kg/m2, and all received varenicline and behavioral counseling) [225]. Furthermore, recent case series and analyses of social media posts indicate that patients receiving semaglutide or tirzepatide report beneficial effects on alcohol-related outcomes [226,227,228,229]. With the exponential growth in clinical use of GLP-1RAs, pharmacoepidemiological studies have also analyzed the association between receipt of GLP-1RAs and alcohol-related outcome, using real-world electronic health records. A recent study using aggregate data from 61 healthcare organizations found that receipt of semaglutide, compared to other non-GLP-1RA anti-obesity and anti-diabetes medications, was associated with reduced risks of incident and recurrent AUD [230]. Other large-scale studies using different datasets have also found beneficial effects with GLP-1RAs on other outcomes, including the frequency and quantity of alcohol use as measured by alcohol use disorders identification test-consumption (AUDIT-C) [231], alcohol-related events [232], intoxication [233], and hospitalizations [234]. While these observational findings are promising, randomized controlled trials (RCTs) are needed to draw conclusions [235, 236]. A recent RCT showed that low-dose semaglutide was effective, compared to placebo, in reducing laboratory alcohol self-administration, as well as drinks per drinking days, heavy drinking, and alcohol craving during the 2-month trial duration [237]. Additional ongoing clinical trials are studying GLP-1RAs, mostly semaglutide, in individuals with AUD and comorbid conditions. Beyond the GLP-1 system, several other feeding/metabolic endocrine pathways have also shown promise as potential pharmacotherapeutic targets for AUD and/or obesity, including (but not limited to) ghrelin [238,239,240,241,242,243], orexins [244,245,246,247,248], amylin [249,250,251,252], and fibroblast growth factor 21 [253,254,255,256]. Further research is needed to investigate the safety and efficacy of novel medications targeting these and other systems for the treatment of AUD and/or obesity.
Beyond behavioral and pharmacological treatments, growing evidence supports the potential use of neuromodulation techniques such as repeated TMS (rTMS), transcranial direct current stimulation (tDCS), deep brain stimulation, and vagus nerve stimulation, as emerging treatments for addictions, including AUD. Among these promising neuromodulation approaches, rTMS is the most investigated technique, using several coils (including the most common 8-coil and the H-coil for deep-TMS) and targeting different brain regions, the most common being the dorsolateral PFC. Of note, the salience network represents a hub of several circuits and networks associated with AUD and recent evidence suggests that rTMS targeting the salience network may be particularly promising for AUD [257]. Similarly, growing evidence suggests a role of neuromodulation approaches for obesity. While previous systematic reviews did not observe clear evidence for these approaches with obesity [258], more recent meta-analytical evidence based on RCTs supports that rTMS as well as tDCS lead to weight loss in people with obesity, possibly via a reduction in food craving [172]. Nonetheless, larger RCTs are needed to assess the potential of rTMS and other neuromodulation approaches for treating AUD and obesity.
Bariatric surgery for obesity and increased risk of AUD
Bariatric surgery is the most effective long-term treatment for severe obesity [259], with over two million procedures in the past decade in the United States [260]. As prevalence of severe obesity grows and new guidelines endorse these surgeries for pediatric patients [261], procedure volumes are expected to climb. In the United States today, sleeve gastrectomy (SG) accounts for ~70% and Roux-en-Y gastric bypass (RYGB) for 27% of cases [262]. Laparoscopic gastric banding (LAGB), has dwindled to less than 1% of surgeries because of modest weight loss and greater long-term weight regain [262]. LAGB merely constricts gastric volume, whereas SG (resection of ~70–80% of the stomach, leaving a tubular remnant) and RYGB (creation of a small gastric pouch anastomosed to the jejunum) fundamentally change gastrointestinal anatomy and physiology. These alterations enhance signals such as GLP-1 [263, 264] reshaping appetite, reward processing, and glycemic control, earning SG and RYGB the label “metabolic surgeries.”
The concept of surgically treating obesity traces back to mid-20th-century observations that patients who underwent partial/total gastrectomy for ulcers or cancer often shed substantial weight [265]. Those same gastric resections, however, were accompanied by unexpectedly high rates of AUD [266, 267]. Only in the late 2000s was a similar association recognized for bariatric procedures [268].
Preclinical work supports a causal link. In several studies, rats with obesity that received RYGB consumed more alcohol or worked harder for oral [57, 269, 270] or intravenous [271, 272] alcohol than sham-operated controls suggesting a biological driver of postoperative alcohol misuse. In rodents, SG does not increase alcohol consumption [273,274,275] (for a detailed review, see also [276]). These divergent findings in rodent models hint that the magnitude and/or mechanisms related to AUD risk may vary by procedure.
One of the earliest clinical signals came from an alcohol treatment program that noted disproportionate enrollment of RYGB patients [277]. Two prospective cohorts in the United States subsequently documented a near-doubling in the incidence of AUD [278] and increased frequency of alcohol use [279] two years after RYGB versus LAGB. At five-year follow-up, ~20% of RYGB patients met AUD criteria determined by the Alcohol Use Disorders Identification Test [280]. A Swedish registry (n = 12,277, 1980–2006) showed that RYGB was associated with a two-fold greater risk of inpatient AUD treatment than LAGB [281] and the controlled Swedish Obese Subjects study confirmed an elevated risk after gastric bypass [282]. A Danish national cohort (n = 14309; 95% RYGB) extended these findings, revealing a 7% higher hazard of AUD five years post-surgery compared with both pre-surgery values and nonsurgical controls with obesity [283]. Because SG became widespread after 2014, the data set in humans is smaller than that for RYGB, but several [284,285,286] though not all [287,288,289] studies suggest SG approximates RYGB in raising AUD risk, particularly for de novo post-operative alcohol problems.
Multiple, potentially interacting mechanisms have been proposed for the increased risk of AUD after metabolic surgery. First, SG and RYGB accelerate alcohol absorption and reduce first-pass metabolism by quickening gastric emptying, which result in earlier and higher blood alcohol concentration (BAC) peaks [290,291,292,293,294,295]. Women can surpass the legal driving BAC limit (0.08%) within minutes of consuming less than two standard drinks [293, 294, 296]. Rapid central (brain) delivery is strongly linked to addictive potential [297]. Furthermore, a BAC of 0.08% meets the definition for binge alcohol drinking, which typically occurs after women consume 4 (and men 5) standard drinks within 2 h [298]. Therefore, RYGB/SG surgeries can turn moderate-seeming intake into pharmacological binge episodes that foster alcohol tolerance, which can further drive AUD [299, 300]. The magnitude of the alcohol-pharmacokinetic changes that follow metabolic surgeries is likely underestimated in studies that use breath analyzers, which cannot capture the very early post-ingestion BAC peaks after RYGB and SG [294, 301, 302].
Second, metabolic surgeries dramatically shift gut-peptide profiles that modulate reward circuits. GLP-1 rises sharply post-prandially [303] whereas fasting ghrelin falls (in SG and at least early in RYGB) [304, 305]. In rodent models, enhanced sensitivity of central ghrelin receptor (the growth hormone secretagogue receptor (GHSR)) mediates heightened motivation for alcohol [271]. Remarkably, 40-50% of patients with high-risk alcohol use before surgery reduced drinking within the first year after RYGB surgery [278, 306,307,308] yet AUD risk surges two or more years later [278, 279, 284]. This temporal pattern supports the idea of dynamic neuroadaptations —initial peptide changes may blunt reward [309], but chronic reductions in ghrelin, for example, could up-regulate GHSR and later amplify alcohol seeking [303].
Third, the marked post-operative caloric restriction inherent to SG and RYGB could sensitize mesolimbic dopamine pathways, paralleling findings that food deprivation enhances the rewarding effects of drugs in humans and animal models [310,311,312]. Finally, obesity itself is associated with reward-circuit alterations resembling those caused by AUD [64, 313], as reviewed above. Once highly palatable foods become less reinforcing after surgery, patients may “transfer” reward seeking to alcohol [314].
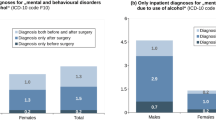
Key knowledge gaps remain. SG is linked to increased AUD in humans but not in rodent models; resolving whether species differences, surgical techniques, or alter alcohol pharmacokinetics accounts for this discrepancy is essential. Another critical research area involves surgery in the pediatric population, who as teenagers and young adults will have a higher baseline propensity for binge drinking. The Teen-LABS study found that 47% of adolescents who underwent metabolic surgery screened positive for excessive alcohol drinking or alcohol-related problems eight years later [315]. As metabolic surgery expands in youth, clinicians must recognize altered alcohol metabolism and implement age-appropriate screening and counseling. Finally, growing evidence links post-bariatric AUD to alcohol-related organ injury, including liver disease [316, 317]. Systematic screening algorithms as well as targeted preventive and therapeutic strategies are urgently needed to maximize the metabolic benefits of surgery while mitigating its unintended addiction-related harms.
Open questions, challenges, and final remarks
This narrative critical review provides an overview of the overlapping features of AUD and obesity (Fig. 1). AUD and obesity are two chronic endemic diseases which share common neurobiological mechanisms, identified in animal models and human neuroimaging studies, and both lead to many similar medical consequences. Another important evidence of the overlap between AUD and obesity is that some treatment modalities work for both weight loss and alcohol reduction. Unfortunately, both AUD and obesity are dramatically under-treated and stigmatized. Our healthcare structure is based on organ-specific specialized approaches and the focus is often treating medical consequences rather than root causes (e.g., treating hypertension but ignoring the underlying AUD and/or obesity). Second, many healthcare providers are not trained in the screening, diagnosis, and management of these diseases, although the addiction medicine / addiction psychiatry and obesity medicine fields are growing. Screening for alcohol problems is not routinely done in primary care. While weight is routinely measured, there is rarely a focused discussion about it. Third, patients often do not seek treatment for their obesity or AUD, due to a variety of reasons, including denial, lack of awareness that effective treatments exist, and the stigma surrounding obesity and AUD. People with obesity, as well as those with AUD, are subject to significant stigma with severe personal, societal, and healthcare consequences [318, 319]. Stigma causes physical and psychological harm to patients, and affected individuals are less likely to receive adequate evidence-based care and treatments [318, 319].

Development and course of alcohol use disorder (AUD) and obesity share several central and peripheral mechanisms, such as dysregulations in reward processing, stress pathways, consummatory behavior, inflammation, immunity, and endocrine systems. Some of the key brain regions with shared neurocircuitry between AUD and obesity include ventral and dorsal striatum, extended amygdala, prefrontal cortex, hypothalamus, nucleus of the solitary tract, and area postrema. Genetic, environmental, and psychosocial factors, some shared and some distinct, also contribute to the natural history of AUD and obesity. While some comorbidities are more prevalent or specific to one condition, AUD and obesity lead to several overlapping medical consequences, such as cancers, liver, gastrointestinal, and cardiovascular diseases, and disruptions in endocrine and immune systems. In addition to lifestyle modifications, treatment options for AUD and obesity include a range of interventions. Some behavioral psychological interventions such as motivational interviewing and cognitive behavioral therapies are used for both conditions, while others are more specific to one (e.g., contingency management for AUD). Food and Drug Administration (FDA)-approved and off-label medications used to treat AUD and obesity also have some commonalities (e.g., naltrexone, topiramate) and growing evidence suggests that, pending additional evidence, anti-obesity GLP-1 therapies (e.g., semaglutide, tirzepatide) have the potential to be repurposed for AUD. Furthermore, growing evidence supports the potential use of neuromodulation techniques such as repeated transcranial magnetic stimulation (rTMS) and others, as emerging novel treatments for obesity as well as addictions, including AUD. Finally, preclinical and human studies suggest a link between metabolic surgeries and increased risk of AUD.
The development of GLP-1RAs as new effective treatments for weight loss has revolutionized medicine. There is much that can be learned and applied to the addiction field, including for AUD. A few examples follow.
-
1)
The treatment of obesity is changing significantly because of the increased acceptance that it is medical condition, rather than simply a lifestyle choice. Likewise, it is critical to promote awareness of addiction, including AUD, as a chronic relapsing and treatable brain disease [320,321,322]. This notion does not negate the important role of environmental and social factors in AUD, but the medical disease model of addiction highlights the need to develop new treatments, including pharmacotherapies and neuromodulation approaches.
-
2)
It is crucial to reduce stigma not only at the individual patient level but also at the organizational and structural levels, including among healthcare providers and policy makers.
-
3)
In addition to increasing clinicians’ awareness of AUD as a medical disorder, an important lesson to learn from the “GLP-1” era is that these complex multifaceted medical conditions need to be studied and managed by multidisciplinary teams, not just a niche of highly specialized providers. The use of GLP-1RAs is impacting not only obesity medicine, but also primary care, cardiology, nephology, hepatology, and surgery. Similarly, AUD is a “whole body” disease, and the engagement of diverse clinicians/disciplines is critically important to improve the screening, diagnosis, and management of people with AUD. Initial efforts in this direction are under way between the addiction and hepatology fields in managing people with AUD and ALD [46,47,48, 323]; such approaches must be expanded across other relevant disciplines. Specific to the current efforts aimed at testing GLP-1RAs for addictions, including AUD, safety considerations are also of paramount importance. For example, patients with AUD may have malnutrition or metabolic conditions that may be worsened by these drugs e.g., loss of free fat mass. The latter is particularly important to keep in mind in patients with AUD and comorbid ALD who often present with sarcopenia. Furthermore, alcohol is one of the main causes of pancreatitis and GLP-1RAs are associated with increased risk of pancreatitis, therefore extra caution is needed in this patient population.
-
4)
The often-prohibitive costs associated with the newer GLP-1RAs for obesity serve as a reminder that similar equity issues may rise for people with AUD, should these or other medications be proven to be effective in AUD [235]. Therefore, it will be imperative to ensure that future new effective treatments have a wide implementation among people with AUD.
-
5)
Given some challenges with accessing GLP-1RAs, e.g., due to shortage in the market, prohibitive costs and other factors, the obesity medicine field has observed an uptake in the prescription of other approved medications for obesity. While there is a need for more pharmacotherapies for AUD, we already have FDA-approved medications for AUD (acamprosate, disulfiram, naltrexone). Despite meta-analytical evidence that naltrexone and acamprosate are effective in people with AUD [324], it’s estimated that <2% of people with AUD in the United States receive an FDA-approved medication for AUD [325]. Therefore, it’s critical to increase the use of FDA-approved medications for AUD for those seeking treatment, while the research field continues to work toward the discovery and development of additional effective treatments.
References
Fryar C, Carroll M, Afful J. Prevalence of overweight, obesity, and extreme obesity among adults aged 20 and over: United States, 1960–1962 through 2017–2018. 2021. 2021/02/05/T09:41:45Z. Accessed 2024/10/18/23:58:26. https://www.cdc.gov/nchs/data/hestat/obesity-adult-17-18/obesity-adult.htm.
Substance Abuse and Mental Health Services Administration. National Survey of Drug Use and Health (NSDUH) Releases. 2022.
Raza SA, Sokale IO, Thrift AP. Burden of high-risk phenotype of heavy alcohol consumption among obese U.S. population: results from National Health and Nutrition Examination Survey, 1999–2020. Lancet Reg Health Am. 2023;23:100525. https://doi.org/10.1016/j.lana.2023.100525.
Hruby A, Manson JE, Qi L, Malik VS, Rimm EB, Sun Q, et al. Determinants and consequences of obesity. Am J Public Health. 2016;106:1656–62. https://doi.org/10.2105/AJPH.2016.303326.
Kranzler HR. Overview of alcohol use disorder. Am J Psychiatry. 2023;180:565–72. https://doi.org/10.1176/appi.ajp.20230488.
Schwartz MW, Seeley RJ, Zeltser LM, Drewnowski A, Ravussin E, Redman LM, et al. Obesity pathogenesis: an endocrine society scientific statement. Endocr Rev. 2017;38:267–96. https://doi.org/10.1210/er.2017-00111.
American Psychiatric Association. Diagnostic and statistical manual of mental disorders (DSM-5®). 5th ed. Washington, DC: American Psychiatric Publishing; 2013.
Mann K. One hundred years of alcoholism: the twentieth century. Alcohol Alcohol. 2000;35:10–15. https://doi.org/10.1093/alcalc/35.1.10.
Lusk JL, Ellison B. Who is to blame for the rise in obesity? Appetite. 2013;68:14–20. https://doi.org/10.1016/j.appet.2013.04.001.
Stanford FC, Tauqeer Z, Kyle TK. Media and its influence on obesity. Curr Obes Rep. 2018;7:186–92. https://doi.org/10.1007/s13679-018-0304-0.
Hammarlund RA, Crapanzano KA, Luce L, Mulligan LA, Ward KM. Review of the effects of self-stigma and perceived social stigma on the treatment-seeking decisions of individuals with drug- and alcohol-use disorders. SAR. 2018;9:115–36. https://doi.org/10.2147/SAR.S183256.
Keyes KM, Hatzenbuehler ML, McLaughlin KA, Link B, Olfson M, Grant BF, et al. Stigma and treatment for alcohol disorders in the United States. Am J Epidemiol. 2010;172:1364–72. https://doi.org/10.1093/aje/kwq304.
Westbury S, Oyebode O, Van Rens T, Barber TM. Obesity stigma: causes, consequences, and potential solutions. Curr Obes Rep. 2023;12:10–23. https://doi.org/10.1007/s13679-023-00495-3.
Loos RJF, Yeo GSH. The genetics of obesity: from discovery to biology. Nat Rev Genet. 2022;23:120–33. https://doi.org/10.1038/s41576-021-00414-z.
Verhulst B, Neale MC, Kendler KS. The heritability of alcohol use disorders: a meta-analysis of twin and adoption studies. Psychol Med. 2015;45:1061–72. https://doi.org/10.1017/S0033291714002165.
Zhou H, Gelernter J. Human genetics and epigenetics of alcohol use disorder. J Clin Invest. 2024;134:e172885. https://doi.org/10.1172/JCI172885.
Grucza RA, Krueger RF, Racette SB, Norberg KE, Hipp PR, Bierut LJ. The emerging link between alcoholism risk and obesity in the United States. Arch Gen Psychiatry. 2010;2010:67.
Deak JD, Miller AP, Gizer IR. Genetics of alcohol use disorder: a review. Curr Opin Psychol. 2019;27:56–61. https://doi.org/10.1016/j.copsyc.2018.07.012.
Saunders GRB, Wang X, Chen F, Jang SK, Liu M, Wang C, et al. Genetic diversity fuels gene discovery for tobacco and alcohol use. Nature. 2022;612:720–4. https://doi.org/10.1038/s41586-022-05477-4.
Zhou H, Kember RL, Deak JD, Xu H, Toikumo S, Yuan K, et al. Multi-ancestry study of the genetics of problematic alcohol use in over 1 million individuals. Nat Med. 2023;29:3184–92. https://doi.org/10.1038/s41591-023-02653-5.
Malone SG, Davis CN, Piserchia Z, Setzer MR, Toikumo S, Zhou H, et al. Alcohol use disorder and body mass index show genetic pleiotropy and shared neural associations. Nat Hum Behav. 2025;9:1056–66. https://doi.org/10.1038/s41562-025-02148-y.
Ghosh-Dastidar B, Cohen D, Hunter G, Zenk SN, Huang C, Beckman R, et al. Distance to store, food prices, and obesity in urban food deserts. Am J Prev Med. 2014;47:587–95. https://doi.org/10.1016/j.amepre.2014.07.005.
Papas MA, Alberg AJ, Ewing R, Helzlsouer KJ, Gary TL, Klassen AC. The built environment and obesity. Epidemiol Rev. 2007;29:129–43. https://doi.org/10.1093/epirev/mxm009.
Anza-Ramirez C, Lazo M, Zafra-Tanaka JH, Avila-Palencia I, Bilal U, Hernández-Vásquez A, et al. The urban built environment and adult BMI, obesity, and diabetes in Latin American cities. Nat Commun. 2022;13:7977. https://doi.org/10.1038/s41467-022-35648-w.
Dimova ED, Lekkas P, Maxwell K, Clemens TL, Pearce JR, Mitchell R, et al. Exploring the influence of local alcohol availability on drinking norms and practices: a qualitative scoping review. Drug Alcohol Rev. 2023;42:691–703. https://doi.org/10.1111/dar.13596.
Freisthler B, Wernekinck U. Examining how the geographic availability of alcohol within residential neighborhoods, activity spaces, and destination nodes is related to alcohol use by parents of young children. Drug Alcohol Depend. 2022;233:109352. https://doi.org/10.1016/j.drugalcdep.2022.109352.
Van Den Ende MWJ, Van Der Maas HLJ, Epskamp S, Lees MH. Alcohol consumption as a socially contagious phenomenon in the Framingham Heart Study social network. Sci Rep. 2024;14:4499. https://doi.org/10.1038/s41598-024-54155-0.
Lim J, Cornwell B. Social network-structural position and obesity: evidence from a national study. Soc Sci Med. 2023;329:116006. https://doi.org/10.1016/j.socscimed.2023.116006.
Christakis NA, Fowler JH. The spread of obesity in a large social network over 32 years. N Engl J Med. 2007;357:370–9. https://doi.org/10.1056/NEJMsa066082.
Wirtz Baker JM, Pou SA, Niclis C, Haluszka E, Aballay LR. Non-traditional data sources in obesity research: a systematic review of their use in the study of obesogenic environments. Int J Obes (Lond). 2023;47:686–96. https://doi.org/10.1038/s41366-023-01331-3.
Karriker-Jaffe KJ, Goode SA, Blakey SM, Humphrey JL, Williams PA, Borja IR, et al. Area-level social determinants of alcohol-related mortality: knowledge gaps and implications for community health. Alcohol Res. 2024;44:06. https://doi.org/10.35946/arcr.v44.1.06.
Capoccia D, Milani I, Colangeli L, Parrotta ME, Leonetti F, Guglielmi V. Social, cultural and ethnic determinants of obesity: from pathogenesis to treatment. Nutr Metab Cardiovasc Dis. 2025;35:103901. https://doi.org/10.1016/j.numecd.2025.103901.
Fletcher PC, Kenny PJ. Food addiction: a valid concept? Neuropsychopharmacology. 2018;43:2506–13. https://doi.org/10.1038/s41386-018-0203-9.
Florio L, Lassi DLS, De Azevedo-Marques Perico C, Vignoli NG, Torales J, Ventriglio A, et al. Food addiction: a comprehensive review. J Nerv Ment Dis. 2022;210:874–9. https://doi.org/10.1097/NMD.0000000000001555.
World Health Organization. Alcohol. https://www.who.int/news-room/fact-sheets/detail/alcohol.
Addolorato G, Mirijello A, Vassallo GA, D’Angelo C, Ferrulli A, Antonelli M, et al. Physical considerations for treatment complications of alcohol and drug use and misuse. In: Johnson B, ed. Addiction medicine. Philadelphia, PA: Elsevier; 2020;pp. 811-32: Chapter 59 (Part IX).
Bray GA. Medical consequences of obesity. J Clin Endocrinol Metab. 2004;89:2583–9. https://doi.org/10.1210/jc.2004-0535.
Suter PM. Is alcohol consumption a risk factor for weight gain and obesity? Crit Rev Clin Lab Sci. 2005;42:197–227. https://doi.org/10.1080/10408360590913542.
Chandrasekaran P, Weiskirchen R. The signaling pathways in obesity-related complications. J Cell Commun Signal. 2024;18:e12039. https://doi.org/10.1002/ccs3.12039.
O’Brien CJO, Domingos AI. An anti-obesity immunotherapy? Science. 2021;373:24–25. https://doi.org/10.1126/science.abj5072.
Meredith LR, Burnette EM, Grodin EN, Irwin MR, Ray LA. Immune treatments for alcohol use disorder: a translational framework. Brain Behav Immun. 2021;97:349–64. https://doi.org/10.1016/j.bbi.2021.07.023.
Vonghia L, Leggio L, Ferrulli A, Bertini M, Gasbarrini G, Addolorato G. Acute alcohol intoxication. Eur J Intern Med. 2008;19:561–7. https://doi.org/10.1016/j.ejim.2007.06.033.
Farinelli LA, Piacentino D, Browning BD, Brewer BB, Leggio L. Cardiovascular consequences of excessive alcohol drinking via electrocardiogram: a systematic review. J Addict Nurs. 2021;32:39–45. https://doi.org/10.1097/jan.0000000000000384.
Knox J, Scodes J, Witkiewitz K, Kranzler HR, Mann K, O’Malley SS, et al. Reduction in world health organization risk drinking levels and cardiovascular disease. Alcohol Clin Exp Res. 2020;44:1625–35. https://doi.org/10.1111/acer.14386.
Wester A, Shang Y, Stål P, Hagström H Time trends in mortality and life expectancy in 22,658 patients hospitalized with alcohol-associated cirrhosis: a nationwide cohort study. Hepatol Commun. 2023;7. https://doi.org/10.1097/hc9.0000000000000279.
Haque LY, Leggio L. Integrated and collaborative care across the spectrum of alcohol-associated liver disease and alcohol use disorder. Hepatology. 2024;80:1408–23. https://doi.org/10.1097/hep.0000000000000996.
Arab JP, Izzy M, Leggio L, Bataller R, Shah VH. Management of alcohol use disorder in patients with cirrhosis in the setting of liver transplantation. Nat Rev Gastroenterol Hepatol. 2022;19:45–59. https://doi.org/10.1038/s41575-021-00527-0.
Leggio L, Mellinger JL. Alcohol use disorder in community management of chronic liver diseases. Hepatology. 2023;77:1006–21. https://doi.org/10.1002/hep.32531.
Åberg F, Helenius-Hietala J, Puukka P, Färkkilä M, Jula A. Interaction between alcohol consumption and metabolic syndrome in predicting severe liver disease in the general population. Hepatology. 2018;67:2141–9. https://doi.org/10.1002/hep.29631.
Rinella ME, Lazarus JV, Ratziu V, Francque SM, Sanyal AJ, Kanwal F, et al. A multisociety Delphi consensus statement on new fatty liver disease nomenclature. Hepatology. 2023;78:1966–86. https://doi.org/10.1097/hep.0000000000000520.
Ghanta A, Wilson E, Chao AM. Sex differences in obesity and its treatment. Curr Psychiatry Rep. 2025;27:278–85. https://doi.org/10.1007/s11920-025-01601-z.
Kenny PJ. Reward mechanisms in obesity: new insights and future directions. Neuron. 2011;69:664–79. https://doi.org/10.1016/j.neuron.2011.02.016.
Martel P, Fantino M. Influence of the amount of food ingested on mesolimbic dopaminergic system activity: a microdialysis study. Pharmacol Biochem Behav. 1996;55:297–302. https://doi.org/10.1016/s0091-3057(96)00087-1.
Geiger BM, Haburcak M, Avena NM, Moyer MC, Hoebel BG, Pothos EN. Deficits of mesolimbic dopamine neurotransmission in rat dietary obesity. Neuroscience. 2009;159:1193–9. https://doi.org/10.1016/j.neuroscience.2009.02.007.
Tellez LA, Medina S, Han W, Ferreira JG, Licona-Limón P, Ren X, et al. A gut lipid messenger links excess dietary fat to dopamine deficiency. Science. 2013;341:800–2. https://doi.org/10.1126/science.1239275.
Hankir MK, Seyfried F, Hintschich CA, Diep TA, Kleberg K, Kranz M, et al. Gastric bypass surgery recruits a Gut PPAR-alpha-striatal D1R pathway to reduce fat appetite in obese rats. Cell Metab. 2017;25:335–44. https://doi.org/10.1016/j.cmet.2016.12.006.
Sirohi S, Richardson BD, Lugo JM, Rossi DJ, Davis JF. Impact of Roux-en-Y gastric bypass surgery on appetite, alcohol intake behaviors, and midbrain ghrelin signaling in the rat. Obesity (Silver Spring). 2017;25:1228–36. https://doi.org/10.1002/oby.21839.
Jadhav KS, Peterson VL, Halfon O, Ahern G, Fouhy F, Stanton C, et al. Gut microbiome correlates with altered striatal dopamine receptor expression in a model of compulsive alcohol seeking. Neuropharmacology. 2018;141:249–59. https://doi.org/10.1016/j.neuropharm.2018.08.026.
Tupala E, Hall H, Bergström K, Särkioja T, Räsänen P, Mantere T, et al. Dopamine D(2)/D(3)-receptor and transporter densities in nucleus accumbens and amygdala of type 1 and 2 alcoholics. Mol Psychiatry. 2001;6:261–7. https://doi.org/10.1038/sj.mp.4000859.
Tupala E, Hall H, Mantere T, Räsänen P, Särkioja T, Tiihonen J. Dopamine receptors and transporters in the brain reward circuits of type 1 and 2 alcoholics measured with human whole hemisphere autoradiography. Neuroimage. 2003;19:145–55. https://doi.org/10.1016/s1053-8119(03)00060-0.
Hietala J, West C, Syvälahti E, Någren K, Lehikoinen P, Sonninen P, et al. Striatal D2 dopamine receptor binding characteristics in vivo in patients with alcohol dependence. Psychopharmacology (Berl). 1994;116:285–90. https://doi.org/10.1007/bf02245330.
Volkow ND, Wang GJ, Fowler JS, Logan J, Hitzemann R, Ding YS, et al. Decreases in dopamine receptors but not in dopamine transporters in alcoholics. Alcohol Clin Exp Res. 1996;20:1594–8. https://doi.org/10.1111/j.1530-0277.1996.tb05936.x.
Ribeiro G, Maia A, Cotovio G, Oliveira FPM, Costa DC, Oliveira-Maia AJ. Striatal dopamine D2-like receptors availability in obesity and its modulation by bariatric surgery: a systematic review and meta-analysis. Sci Rep. 2023;13:4959. https://doi.org/10.1038/s41598-023-31250-2.
Wang GJ, Volkow ND, Logan J, Pappas NR, Wong CT, Zhu W, et al. Brain dopamine and obesity. Lancet. 2001;357:354–7. https://doi.org/10.1016/s0140-6736(00)03643-6.
Stice E, Spoor S, Bohon C, Small DM. Relation between obesity and blunted striatal response to food is moderated by TaqIA A1 allele. Science. 2008;322:449–52. https://doi.org/10.1126/science.1161550.
Noble EP. Addiction and its reward process through polymorphisms of the D2 dopamine receptor gene: a review. Eur Psychiatry. 2000;15:79–89. https://doi.org/10.1016/s0924-9338(00)00208-x.
Smith L, Watson M, Gates S, Ball D, Foxcroft D. Meta-analysis of the association of the Taq1A polymorphism with the risk of alcohol dependency: a HuGE gene-disease association review. Am J Epidemiol. 2008;167:125–38. https://doi.org/10.1093/aje/kwm281.
Phillips TJ, Brown KJ, Burkhart-Kasch S, Wenger CD, Kelly MA, Rubinstein M, et al. Alcohol preference and sensitivity are markedly reduced in mice lacking dopamine D2 receptors. Nat Neurosci. 1998;1:610–5. https://doi.org/10.1038/2843.
Bocarsly ME, da Silva E Silva D, Kolb V, Luderman KD, Shashikiran S, Rubinstein M, et al. A mechanism linking two known vulnerability factors for alcohol abuse: heightened alcohol stimulation and low striatal dopamine D2 receptors. Cell Rep. 2019;29:1147–1163.e5. https://doi.org/10.1016/j.celrep.2019.09.059.
Johnson PM, Kenny PJ. Dopamine D2 receptors in addiction-like reward dysfunction and compulsive eating in obese rats. Nat Neurosci. 2010;13:635–41. https://doi.org/10.1038/nn.2519.
Anderberg RH, Anefors C, Bergquist F, Nissbrandt H, Skibicka KP. Dopamine signaling in the amygdala, increased by food ingestion and GLP-1, regulates feeding behavior. Physiol Behav. 2014;136:135–44. https://doi.org/10.1016/j.physbeh.2014.02.026.
Aranäs C, Edvardsson CE, Shevchouk OT, Zhang Q, Witley S, Blid Sköldheden S, et al. Semaglutide reduces alcohol intake and relapse-like drinking in male and female rats. EBioMedicine. 2023;93:104642. https://doi.org/10.1016/j.ebiom.2023.104642.
Egecioglu E, Engel JA, Jerlhag E. The glucagon-like peptide 1 analogue, exendin-4, attenuates the rewarding properties of psychostimulant drugs in mice. PLoS ONE. 2013;8:e69010. https://doi.org/10.1371/journal.pone.0069010.
Egecioglu E, Engel JA, Jerlhag E. The glucagon-like peptide 1 analogue Exendin-4 attenuates the nicotine-induced locomotor stimulation, accumbal dopamine release, conditioned place preference as well as the expression of locomotor sensitization in mice. PLoS ONE. 2013;8:e77284. https://doi.org/10.1371/journal.pone.0077284.
Falk S, Petersen J, Svendsen C, Romero-Leguizamón CR, Jørgensen SH, Krauth N, et al. GLP-1 and nicotine combination therapy engages hypothalamic and mesolimbic pathways to reverse obesity. Cell Rep. 2023;42:112466. https://doi.org/10.1016/j.celrep.2023.112466.
Fortin SM, Chartoff EH, Roitman MF. The aversive agent lithium chloride suppresses phasic dopamine release through central GLP-1 receptors. Neuropsychopharmacol. 2016;41:906–15. https://doi.org/10.1038/npp.2015.220.
Fortin SM, Roitman MF. Central GLP-1 receptor activation modulates cocaine-evoked phasic dopamine signaling in the nucleus accumbens core. Physiol Behav. 2017;176:17–25. https://doi.org/10.1016/j.physbeh.2017.03.019.
Sandoval-Rodriguez R, Parra-Reyes JA, Han W, Rueda-Orozco PE, Perez IO, de Araujo IE, et al. D1 and D2 neurons in the nucleus accumbens enable positive and negative control over sugar intake in mice. Cell Rep. 2023;42:112190. https://doi.org/10.1016/j.celrep.2023.112190.
Sorensen G, Reddy IA, Weikop P, Graham DL, Stanwood GD, Wortwein G, et al. The glucagon-like peptide 1 (GLP-1) receptor agonist exendin-4 reduces cocaine self-administration in mice. Physiol Behav. 2015;149:262–8. https://doi.org/10.1016/j.physbeh.2015.06.013.
Vallof D, Kalafateli AL, Jerlhag E. Brain region specific glucagon-like peptide-1 receptors regulate alcohol-induced behaviors in rodents. Psychoneuroendocrinology. 2019;103:284–95. https://doi.org/10.1016/j.psyneuen.2019.02.006.
Vallof D, Maccioni P, Colombo G, Mandrapa M, Jörnulf JW, Egecioglu E, et al. The glucagon-like peptide 1 receptor agonist liraglutide attenuates the reinforcing properties of alcohol in rodents. Addict Biol. 2016;21:422–37. https://doi.org/10.1111/adb.12295.
Wang XF, Liu JJ, Xia J, Liu J, Mirabella V, Pang ZP. Endogenous Glucagon-like peptide-1 suppresses high-fat food intake by reducing synaptic drive onto mesolimbic dopamine neurons. Cell Rep. 2015;12:726–33. https://doi.org/10.1016/j.celrep.2015.06.062.
Zhang Y, Kahng MW, Elkind JA, Weir VR, Hernandez NS, Stein LM, et al. Activation of GLP-1 receptors attenuates oxycodone taking and seeking without compromising the antinociceptive effects of oxycodone in rats. Neuropsychopharmacol. 2020;45:451–61. https://doi.org/10.1038/s41386-019-0531-4.
Tomiyama AJ. Stress and obesity. Annu Rev Psychol. 2019;70:703–18. https://doi.org/10.1146/annurev-psych-010418-102936.
Edwards S, Little HJ, Richardson HN, Vendruscolo LF. Divergent regulation of distinct glucocorticoid systems in alcohol dependence. Alcohol. 2015;49:811–6. https://doi.org/10.1016/j.alcohol.2015.04.004.
McGinn MA, Pantazis CB, Tunstall BJ, Marchette RCN, Carlson ER, Said N, et al. Drug addiction co-morbidity with alcohol: neurobiological insights. Int Rev Neurobiol. 2021;157:409–72. https://doi.org/10.1016/bs.irn.2020.11.002.
Sominsky L, Spencer SJ. Eating behavior and stress: a pathway to obesity. Front Psychol. 2014;5:434 https://doi.org/10.3389/fpsyg.2014.00434.
Stephens MA, Wand G. Stress and the HPA axis: role of glucocorticoids in alcohol dependence. Alcohol Res. 2012;34:468–83.
Wemm SE, Sinha R. Drug-induced stress responses and addiction risk and relapse. Neurobiol Stress. 2019;10:100148. https://doi.org/10.1016/j.ynstr.2019.100148.
Blaine SK, Nautiyal N, Hart R, Guarnaccia JB, Sinha R. Craving, cortisol and behavioral alcohol motivation responses to stress and alcohol cue contexts and discrete cues in binge and non-binge drinkers. Addict Biol. 2019;24:1096–108. https://doi.org/10.1111/adb.12665.
Sinha R. Role of addiction and stress neurobiology on food intake and obesity. Biol Psychol. 2018;131:5–13. https://doi.org/10.1016/j.biopsycho.2017.05.001.
Dallman MF, Pecoraro NC, la Fleur SE. Chronic stress and comfort foods: self-medication and abdominal obesity. Brain Behav Immun. 2005;19:275–80. https://doi.org/10.1016/j.bbi.2004.11.004.
Ip CK, Rezitis J, Qi Y, Bajaj N, Koller J, Farzi A, et al. Critical role of lateral habenula circuits in the control of stress-induced palatable food consumption. Neuron. 2023;111:2583–2600.e6. https://doi.org/10.1016/j.neuron.2023.05.010.
Gilpin NW, Herman MA, Roberto M. The central amygdala as an integrative hub for anxiety and alcohol use disorders. Biol Psychiatry. 2015;77:859–69. https://doi.org/10.1016/j.biopsych.2014.09.008.
Roberto M, Madamba SG, Moore SD, Tallent MK, Siggins GR. Ethanol increases GABAergic transmission at both pre- and postsynaptic sites in rat central amygdala neurons. Proc Natl Acad Sci USA. 2003;100:2053–8. https://doi.org/10.1073/pnas.0437926100.
Nie Z, Schweitzer P, Roberts AJ, Madamba SG, Moore SD, Siggins GR. Ethanol augments GABAergic transmission in the central amygdala via CRF1 receptors. Science. 2004;303:1512–4. https://doi.org/10.1126/science.1092550.
Hardaway JA, Halladay LR, Mazzone CM, Pati D, Bloodgood DW, Kim M, et al. Central amygdala prepronociceptin-expressing neurons mediate palatable food consumption and reward. Neuron. 2019;102:1037–1052.e7. https://doi.org/10.1016/j.neuron.2019.03.037.
Torruella-Suarez ML, Vandenberg JR, Cogan ES, Ipton GJ, Teklezghi A, Dange K, et al. Manipulations of central amygdala neurotensin neurons alter the consumption of ethanol and sweet fluids in mice. J Neurosci. 2020;40:632–47. https://doi.org/10.1523/JNEUROSCI.1466-19.2019.
Funk CK, O’Dell LE, Crawford EF, Koob GF. Corticotropin-releasing factor within the central nucleus of the amygdala mediates enhanced ethanol self-administration in withdrawn, ethanol-dependent rats. J Neurosci. 2006;26:11324–32. https://doi.org/10.1523/jneurosci.3096-06.2006.
Micioni Di Bonaventura MV, Ciccocioppo R, Romano A, Bossert JM, Rice KC, et al. Role of bed nucleus of the stria terminalis corticotrophin-releasing factor receptors in frustration stress-induced binge-like palatable food consumption in female rats with a history of food restriction. J Neurosci. 2014;34:11316–24. https://doi.org/10.1523/JNEUROSCI.1854-14.2014.
Dumont EC, Mark GP, Mader S, Williams JT. Self-administration enhances excitatory synaptic transmission in the bed nucleus of the stria terminalis. Nat Neurosci. 2005;8:413–4. https://doi.org/10.1038/nn1414.
Sotelo-Hitschfeld T, Minere M, Klemm P, et al. GABAergic disinhibition from the BNST to PNOC(ARC) neurons promotes HFD-induced hyperphagia. Cell Rep. 2024;43:114343. https://doi.org/10.1016/j.celrep.2024.114343.
de Araujo Salgado I, Li C, Burnett CJ, Rodriguez Gonzalez S, Becker JJ, et al. Toggling between food-seeking and self-preservation behaviors via hypothalamic response networks. Neuron. 2023;111:2899–2917.e6. https://doi.org/10.1016/j.neuron.2023.06.006.
Hao S, Yang H, Wang X, He Y, Xu H, Wu X, et al. The lateral hypothalamic and BNST GABAergic projections to the anterior ventrolateral periaqueductal gray regulate feeding. Cell Rep. 2019;28:616–624.e5. https://doi.org/10.1016/j.celrep.2019.06.051.
Wang Y, Kim J, Schmit MB, Cho TS, Fang C, Cai H. A bed nucleus of stria terminalis microcircuit regulating inflammation-associated modulation of feeding. Nat Commun. 2019;10:2769. https://doi.org/10.1038/s41467-019-10715-x.
Cork SC, Richards JE, Holt MK, Gribble FM, Reimann F, Trapp S. Distribution and characterisation of Glucagon-like peptide-1 receptor expressing cells in the mouse brain. Mol Metab. 2015;4:718–31. https://doi.org/10.1016/j.molmet.2015.07.008.
Qiao H, Ren WN, Li HZ, Hou YX. Inhibitory effects of peripheral administration of exendin-4 on food intake are attenuated by lesions of the central nucleus of amygdala. Brain Res Bull. 2019;148:131–5. https://doi.org/10.1016/j.brainresbull.2019.03.002.
Zeng N, Cutts EJ, Lopez CB, Kaur S, Duran M, Virkus SA, et al. Anatomical and functional characterization of central amygdala Glucagon-Like peptide 1 receptor expressing neurons. Front Behav Neurosci. 2021;15:724030. https://doi.org/10.3389/fnbeh.2021.724030.
Farokhnia M, Browning BD, Crozier ME, Sun H, Akhlaghi F, Leggio L. The glucagon-like peptide-1 system is modulated by acute and chronic alcohol exposure: findings from human laboratory experiments and a post-mortem brain study. Addict Biol. 2022;27:e13211. https://doi.org/10.1111/adb.13211.
Chuong V, Farokhnia M, Khom S, Pince CL, Elvig SK, Vlkolinsky R, et al. The glucagon-like peptide-1 (GLP-1) analogue semaglutide reduces alcohol drinking and modulates central GABA neurotransmission. JCI Insight. 2023;8. https://doi.org/10.1172/jci.insight.170671.
Baumgartner I, Pacheco-Lopez G, Ruttimann EB, Arnold M, Asarian L, Langhans W, et al. Hepatic-portal vein infusions of glucagon-like peptide-1 reduce meal size and increase c-Fos expression in the nucleus tractus solitarii, area postrema and central nucleus of the amygdala in rats. J Neuroendocrinol. 2010;22:557–63. https://doi.org/10.1111/j.1365-2826.2010.01995.x.
Turton MD, O’Shea D, Gunn I, Beak SA, Edwards CM, Meeran K, et al. A role for glucagon-like peptide-1 in the central regulation of feeding. Nature. 1996;379:69–72. https://doi.org/10.1038/379069a0.
Kinzig KP, D’Alessio DA, Seeley RJ. The diverse roles of specific GLP-1 receptors in the control of food intake and the response to visceral illness. J Neurosci. 2002;22:10470–6. https://doi.org/10.1523/JNEUROSCI.22-23-10470.2002.
Williams DL, Lilly NA, Edwards IJ, Yao P, Richards JE, Trapp S. GLP-1 action in the mouse bed nucleus of the stria terminalis. Neuropharmacology. 2018;131:83–95. https://doi.org/10.1016/j.neuropharm.2017.12.007.
Grill HJ. Distributed neural control of energy balance: contributions from hindbrain and hypothalamus. Obesity (Silver Spring). 2006;14(Suppl 5):216S–221S. https://doi.org/10.1038/oby.2006.312.
Nieh EH, Matthews GA, Allsop SA, Presbrey KN, Leppla CA, Wichmann R, et al. Decoding neural circuits that control compulsive sucrose seeking. Cell. 2015;160:528–41. https://doi.org/10.1016/j.cell.2015.01.003.
Stamatakis AM, Van Swieten M, Basiri ML, Blair GA, Kantak P, Stuber GD. Lateral hypothalamic area glutamatergic neurons and their projections to the lateral habenula regulate feeding and reward. J Neurosci. 2016;36:302–11. https://doi.org/10.1523/JNEUROSCI.1202-15.2016.
Rossi MA, Stuber GD. Overlapping brain circuits for homeostatic and hedonic feeding. Cell Metab. 2018;27:42–56. https://doi.org/10.1016/j.cmet.2017.09.021.
Rossi MA, Basiri ML, McHenry JA, Kosyk O, Otis JM, van den Munkhof HE, et al. Obesity remodels activity and transcriptional state of a lateral hypothalamic brake on feeding. Science. 2019;364:1271–4. https://doi.org/10.1126/science.aax1184.
Jennings JH, Rizzi G, Stamatakis AM, Ung RL, Stuber GD. The inhibitory circuit architecture of the lateral hypothalamus orchestrates feeding. Science. 2013;341:1517–21. https://doi.org/10.1126/science.1241812.
Eiler WJ 2nd, Dzemidzic M, Case KR, Soeurt CM, Armstrong CL, et al. The aperitif effect: alcohol’s effects on the brain’s response to food aromas in women. Obesity (Silver Spring). 2015;23:1386–93. https://doi.org/10.1002/oby.21109.
Barson JR, Leibowitz SF. Hypothalamic neuropeptide signaling in alcohol addiction. Prog Neuropsychopharmacol Biol Psychiatry. 2016;65:321–9. https://doi.org/10.1016/j.pnpbp.2015.02.006.
Marty VN, Mulpuri Y, Munier JJ, Spigelman I. Chronic alcohol disrupts hypothalamic responses to stress by modifying CRF and NMDA receptor function. Neuropharmacology. 2020;167:107991. https://doi.org/10.1016/j.neuropharm.2020.107991.
Farokhnia M, Grodin EN, Lee MR, Oot EN, Blackburn AN, Stangl BL, et al. Exogenous ghrelin administration increases alcohol self-administration and modulates brain functional activity in heavy-drinking alcohol-dependent individuals. Mol Psychiatry. 2018;23:2029–38. https://doi.org/10.1038/mp.2017.226.
Wayner MJ. Craving for alcohol in the rat: adjunctive behavior and the lateral hypothalamus. Pharmacol Biochem Behav. 2002;73:27–43. https://doi.org/10.1016/s0091-3057(02)00780-3.
Marchant NJ, Rabei R, Kaganovsky K, Caprioli D, Bossert JM, Bonci A, et al. A critical role of lateral hypothalamus in context-induced relapse to alcohol seeking after punishment-imposed abstinence. J Neurosci. 2014;34:7447–57. https://doi.org/10.1523/JNEUROSCI.0256-14.2014.
Greene LW, Hollander CS. Alcohol and the hypothalamus. Prog Biochem Pharmacol. 1981;18:15–23.
Campbell EJ, Hill MK, Maddern XJ, Jin S, Pang TY, Lawrence AJ. Orexin-1 receptor signaling within the lateral hypothalamus, but not bed nucleus of the stria terminalis, mediates context-induced relapse to alcohol seeking. J Psychopharmacol. 2020;34:1261–70. https://doi.org/10.1177/0269881120959638.
Colvin KJ, Killen HS, Kanter MJ, Halperin MC, Engel L, Currie PJ Brain site-specific inhibitory effects of the GLP-1 analogue Exendin-4 on alcohol intake and operant responding for palatable food. Int J Mol Sci. 2020;21. https://doi.org/10.3390/ijms21249710.
Richard JE, Farkas I, Anesten F, Anderberg RH, Dickson SL, Gribble FM, et al. GLP-1 receptor stimulation of the lateral parabrachial nucleus reduces food intake: neuroanatomical, electrophysiological, and behavioral evidence. Endocrinology. 2014;155:4356–67. https://doi.org/10.1210/en.2014-1248.
Lopez-Ferreras L, Richard JE, Noble EE, Eerola K, Anderberg RH, Olandersson K, et al. Lateral hypothalamic GLP-1 receptors are critical for the control of food reinforcement, ingestive behavior and body weight. Mol Psychiatry. 2018;23:1157–68. https://doi.org/10.1038/mp.2017.187.
Burmeister MA, Ayala JE, Smouse H, Landivar-Rocha A, Brown JD, Drucker DJ, et al. The hypothalamic Glucagon-Like peptide 1 receptor is sufficient but not necessary for the regulation of energy balance and glucose homeostasis in mice. Diabetes. 2017;66:372–84. https://doi.org/10.2337/db16-1102.
Cheng W, Gordian D, Ludwig MQ, Pers TH, Seeley RJ, Myers MG Jr. Hindbrain circuits in the control of eating behaviour and energy balance. Nat Metab. 2022;4:826–35. https://doi.org/10.1038/s42255-022-00606-9.
Dowsett GKC, Lam BYH, Tadross JA, Cimino I, Rimmington D, Coll AP, et al. A survey of the mouse hindbrain in the fed and fasted states using single-nucleus RNA sequencing. Mol Metab. 2021;53:101240. https://doi.org/10.1016/j.molmet.2021.101240.
Ludwig MQ, Cheng W, Gordian D, Lee J, Paulsen SJ, Hansen SN, et al. A genetic map of the mouse dorsal vagal complex and its role in obesity. Nat Metab. 2021;3:530–45. https://doi.org/10.1038/s42255-021-00363-1.
Zhang C, Kaye JA, Cai Z, Wang Y, Prescott SL, Liberles SD. Area postrema cell types that mediate nausea-associated behaviors. Neuron. 2021;109:461–472.e5. https://doi.org/10.1016/j.neuron.2020.11.010.
Cheng W, Ndoka E, Maung JN, Pan W, Rupp AC, Rhodes CJ, et al. NTS Prlh overcomes orexigenic stimuli and ameliorates dietary and genetic forms of obesity. Nat Commun. 2021;12:5175. https://doi.org/10.1038/s41467-021-25525-3.
Hsu JY, Crawley S, Chen M, Ayupova DA, Lindhout DA, Higbee J, et al. Non-homeostatic body weight regulation through a brainstem-restricted receptor for GDF15. Nature. 2017;550:255–9. https://doi.org/10.1038/nature24042.
Edwards GL, Ritter RC. Area postrema lesions: cause of overingestion is not altered visceral nerve function. Am J Physiol. 1986;251:R575–81. https://doi.org/10.1152/ajpregu.1986.251.3.R575.
Stewart RB, Perlanski E, Grupp LA. Area postrema and alcohol: effects of area postrema lesions on ethanol self-administration, pharmacokinetics, and ethanol-induced conditioned taste aversion. Alcohol Clin Exp Res. 1988;12:698–704. https://doi.org/10.1111/j.1530-0277.1988.tb00268.x.
Tuesta LM, Chen Z, Duncan A, Fowler CD, Ishikawa M, Lee BR, et al. GLP-1 acts on habenular avoidance circuits to control nicotine intake. Nat Neurosci. 2017;20:708–16. https://doi.org/10.1038/nn.4540.
Kastin AJ, Akerstrom V, Pan W. Interactions of glucagon-like peptide-1 (GLP-1) with the blood-brain barrier. J Mol Neurosci. 2002;18:7–14. https://doi.org/10.1385/JMN:18:1-2:07.
Merchenthaler I, Lane M, Shughrue P. Distribution of pre-pro-glucagon and glucagon-like peptide-1 receptor messenger RNAs in the rat central nervous system. J Comp Neurol. 1999;403:261–80.
Larsen PJ, Tang-Christensen M, Holst JJ, Orskov C. Distribution of glucagon-like peptide-1 and other preproglucagon-derived peptides in the rat hypothalamus and brainstem. Neuroscience. 1997;77:257–70. https://doi.org/10.1016/s0306-4522(96)00434-4.
Holt MK, Richards JE, Cook DR, Brierley DI, Williams DL, Reimann F, et al. Preproglucagon neurons in the nucleus of the solitary tract are the main source of brain GLP-1, mediate stress-induced hypophagia, and limit unusually large intakes of food. Diabetes. 2019;68:21–33. https://doi.org/10.2337/db18-0729.
Huang KP, Acosta AA, Ghidewon MY, McKnight AD, Almeida MS, Nyema NT, et al. Dissociable hindbrain GLP1R circuits for satiety and aversion. Nature. 2024;632:585–93. https://doi.org/10.1038/s41586-024-07685-6.
Hayashi M, Kaye JA, Douglas ER, Joshi NR, Gribble FM, Reimann F, et al. Enteroendocrine cell lineages that differentially control feeding and gut motility. Elife. 2023;12 https://doi.org/10.7554/eLife.78512.
Beumer J, Geurts MH, Geurts V, Andersson-Rolf A, Akkerman N, Völlmy F, et al. Description and functional validation of human enteroendocrine cell sensors. Science. 2024;386:341–8. https://doi.org/10.1126/science.adl1460.
Wettergren A, Petersen H, Orskov C, Christiansen J, Sheikh SP, Holst JJ. Glucagon-like peptide-1 7-36 amide and peptide YY from the L-cell of the ileal mucosa are potent inhibitors of vagally induced gastric acid secretion in man. Scand J Gastroenterol. 1994;29:501–5. https://doi.org/10.3109/00365529409092462.
Bai L, Mesgarzadeh S, Ramesh KS, Huey EL, Liu Y, Gray LA, et al. Genetic identification of vagal sensory neurons that control feeding. Cell. 2019;179:1129–1143.e23. https://doi.org/10.1016/j.cell.2019.10.031.
Brierley DI, Holt MK, Singh A, de Araujo A, McDougle M, Vergara M, et al. Central and peripheral GLP-1 systems independently suppress eating. Nat Metab. 2021;3:258–73. https://doi.org/10.1038/s42255-021-00344-4.
Kim DH, D’Alessio DA, Woods SC, Seeley RJ. The effects of GLP-1 infusion in the hepatic portal region on food intake. Regul Pept. 2009;155:110–4. https://doi.org/10.1016/j.regpep.2009.03.002.
Ruttimann EB, Arnold M, Hillebrand JJ, Geary N, Langhans W. Intrameal hepatic portal and intraperitoneal infusions of glucagon-like peptide-1 reduce spontaneous meal size in the rat via different mechanisms. Endocrinology. 2009;150:1174–81. https://doi.org/10.1210/en.2008-1221.
Diepenbroek C, Quinn D, Stephens R, Zollinger B, Anderson S, Pan A, et al. Validation and characterization of a novel method for selective vagal deafferentation of the gut. Am J Physiol Gastrointest Liver Physiol. 2017;313:G342–G352. https://doi.org/10.1152/ajpgi.00095.2017.
Plamboeck A, Veedfald S, Deacon CF, Hartmann B, Wettergren A, Svendsen LB, et al. The effect of exogenous GLP-1 on food intake is lost in male truncally vagotomized subjects with pyloroplasty. Am J Physiol Gastrointest Liver Physiol. 2013;304:G1117–27. https://doi.org/10.1152/ajpgi.00035.2013.
Kanoski SE, Fortin SM, Arnold M, Grill HJ, Hayes MR. Peripheral and central GLP-1 receptor populations mediate the anorectic effects of peripherally administered GLP-1 receptor agonists, liraglutide and exendin-4. Endocrinology. 2011;152:3103–12. https://doi.org/10.1210/en.2011-0174.
Abbott CR, Monteiro M, Small CJ, Sajedi A, Smith KL, Parkinson JR, et al. The inhibitory effects of peripheral administration of peptide YY(3-36) and glucagon-like peptide-1 on food intake are attenuated by ablation of the vagal-brainstem-hypothalamic pathway. Brain Res. 2005;1044:127–31. https://doi.org/10.1016/j.brainres.2005.03.011.
Gonzalez MF, Deutsch JA. Vagotomy abolishes cues of satiety produced by gastric distension. Science. 1981;212:1283–4. https://doi.org/10.1126/science.7233218.
Borgmann D, Ciglieri E, Biglari N, Brandt C, Cremer AL, Backes H, et al. Gut-brain communication by distinct sensory neurons differently controls feeding and glucose metabolism. Cell Metab. 2021;33:1466–1482.e7. https://doi.org/10.1016/j.cmet.2021.05.002.
Daly DM, Park SJ, Valinsky WC, Beyak MJ. Impaired intestinal afferent nerve satiety signalling and vagal afferent excitability in diet induced obesity in the mouse. J Physiol. 2011;589:2857–70. https://doi.org/10.1113/jphysiol.2010.204594.
Covasa M, Grahn J, Ritter RC. Reduced hindbrain and enteric neuronal response to intestinal oleate in rats maintained on high-fat diet. Auton Neurosci. 2000;84:8–18. https://doi.org/10.1016/S1566-0702(00)00176-4.
Covasa M, Ritter RC. Reduced sensitivity to the satiation effect of intestinal oleate in rats adapted to high-fat diet. Am J Physiol. 1999;277:R279–85. https://doi.org/10.1152/ajpregu.1999.277.1.R279.
Keller BN, Snyder AE, Coker CR, Aguilar EA, O’Brien MK, Lookfong NA, et al. Vagus nerve damage increases alcohol intake and preference in a nonpreferring rat line: relationship to vagal regulation of the hypothalamic-pituitary-adrenal axis. Alcohol Clin Exp Res (Hoboken). 2024;48:488–98. https://doi.org/10.1111/acer.15264.
Zeng J, Yu S, Cao H, Su Y, Dong Z, Yang X. Neurobiological correlates of cue-reactivity in alcohol-use disorders: a voxel-wise meta-analysis of fMRI studies. Neurosci Biobehav Rev. 2021;128:294–310. https://doi.org/10.1016/j.neubiorev.2021.06.031.
Gluck ME, Viswanath P, Stinson EJ. Obesity, appetite, and the prefrontal cortex. Curr Obes Rep. 2017;6:380–8. https://doi.org/10.1007/s13679-017-0289-0.
Friedman NP, Robbins TW. The role of prefrontal cortex in cognitive control and executive function. Neuropsychopharmacol. 2022;47:72–89. https://doi.org/10.1038/s41386-021-01132-0.
Michaud A, Vainik U, Garcia-Garcia I, Dagher A. Overlapping neural endophenotypes in addiction and obesity. Front Endocrinol (Lausanne). 2017;8:127. https://doi.org/10.3389/fendo.2017.00127.
Eichen DM, Pasquale EK, Twamley EW, Boutelle KN. Targeting executive function for weight loss in adults with overweight or obesity. Physiol Behav. 2021;240:113540. https://doi.org/10.1016/j.physbeh.2021.113540.
Veling H, van Koningsbruggen GM, Aarts H, Stroebe W. Targeting impulsive processes of eating behavior via the internet. Effects on body weight. Appetite. 2014;78:102–9. https://doi.org/10.1016/j.appet.2014.03.014.
Wilcox CE, Dekonenko CJ, Mayer AR, Bogenschutz MP, Turner JA. Cognitive control in alcohol use disorder: deficits and clinical relevance. Rev Neurosci. 2014;25:1–24. https://doi.org/10.1515/revneuro-2013-0054.
Maatoug R, Bihan K, Duriez P, Podevin P, Silveira-Reis-Brito L, Benyamina A, et al. Non-invasive and invasive brain stimulation in alcohol use disorders: a critical review of selected human evidence and methodological considerations to guide future research. Compr Psychiatry. 2021;109:152257. https://doi.org/10.1016/j.comppsych.2021.152257.
Alhindi YA, Khalifa N, Al-Khyatt W, Idris I. The use of non-invasive brain stimulation techniques to reduce body weight and food cravings: a systematic review and meta-analysis. Clin Obes. 2023;13:e12611. https://doi.org/10.1111/cob.12611.
Schacht JP, Anton RF, Myrick H. Functional neuroimaging studies of alcohol cue reactivity: a quantitative meta‐analysis and systematic review. Addict Biol. 2013;18:121–33. https://doi.org/10.1111/j.1369-1600.2012.00464.x.
Noori HR, Cosa Linan A, Spanagel R. Largely overlapping neuronal substrates of reactivity to drug, gambling, food and sexual cues: a comprehensive meta-analysis. Eur Neuropsychopharmacol. 2016;26:1419–30. https://doi.org/10.1016/j.euroneuro.2016.06.013.
Yang Y, Wu Q, Morys F. Brain responses to high-calorie visual food cues in individuals with normal-weight or obesity: an activation likelihood estimation meta-analysis. Brain Sci. 2021;11:1587. https://doi.org/10.3390/brainsci11121587.
Van Der Laan LN, De Ridder DTD, Viergever MA, Smeets PAM. The first taste is always with the eyes: a meta-analysis on the neural correlates of processing visual food cues. Neuroimage. 2011;55:296–303. https://doi.org/10.1016/j.neuroimage.2010.11.055.
Tang DW, Fellows LK, Small DM, Dagher A. Food and drug cues activate similar brain regions: a meta-analysis of functional MRI studies. Physiol Behav. 2012;106:317–24. https://doi.org/10.1016/j.physbeh.2012.03.009.
Li G, Hu Y, Zhang W, et al. Brain functional and structural magnetic resonance imaging of obesity and weight loss interventions. Mol Psychiatry. 2023;28:1466–79. https://doi.org/10.1038/s41380-023-02025-y.
Fredriksson I, Venniro M, Reiner DJ, Chow JJ, Bossert JM, Shaham Y. Animal models of drug relapse and craving after voluntary abstinence: a review. Pharmacol Rev. 2021;73:1050–83. https://doi.org/10.1124/pharmrev.120.000191.
Venniro M, Banks ML, Heilig M, Epstein DH, Shaham Y. Improving translation of animal models of addiction and relapse by reverse translation. Nat Rev Neurosci. 2020;21:625–43. https://doi.org/10.1038/s41583-020-0378-z.
Avery JA, Burrows K, Kerr KL, Bodurka J, Khalsa SS, Paulus MP, et al. How the brain wants what the body needs: the neural basis of positive alliesthesia. Neuropsychopharmacol. 2017;42:822–30. https://doi.org/10.1038/npp.2016.128.
Koban L, Wager TD, Kober H. A neuromarker for drug and food craving distinguishes drug users from non-users. Nat Neurosci. 2023;26:316–25. https://doi.org/10.1038/s41593-022-01228-w.
Kelly JF, Abry A, Ferri M, Humphreys K. Alcoholics anonymous and 12-step facilitation treatments for alcohol use disorder: a distillation of a 2020 cochrane review for clinicians and policy makers. Alcohol Alcohol. 2020;55:641–51. https://doi.org/10.1093/alcalc/agaa050.
McCrady BS, Owens MD, Borders AZ, Brovko JM Psychosocial approaches to alcohol use disorders since 1940: a review. J Stud Alcohol Drugs Suppl. 2014:68-78. https://doi.org/10.15288/jsads.2014.s17.68.
Witkiewitz K, Marlatt A. Behavioral therapy across the spectrum. Alcohol Res Health. 2011;33:313–9.
Witkiewitz K, Litten RZ, Leggio L. Advances in the science and treatment of alcohol use disorder. Sci Adv. 2019;5:eaax4043. https://doi.org/10.1126/sciadv.aax4043.
Lékó AH, Farokhnia M, Leggio L. Approved, promising, and experimental medications for treatment of alcohol use disorder. In: Mueller S, Heilig M, eds. Alcohol and alcohol-related diseases. Cham, Switzerland: Springer International Publishing; 2023. pp. 261-88.
Heilig M, Witkiewitz K, Ray LA, Leggio L Novel medications for problematic alcohol use. J Clin Invest. 2024;134. https://doi.org/10.1172/jci172889.
Ray LA, Meredith LR, Kiluk BD, Walthers J, Carroll KM, Magill M. Combined pharmacotherapy and cognitive behavioral therapy for adults with alcohol or substance use disorders: a systematic review and meta-analysis. JAMA Netw Open. 2020;3:e208279. https://doi.org/10.1001/jamanetworkopen.2020.8279.
Cowan J, Devine C. Food, eating, and weight concerns of men in recovery from substance addiction. Appetite. 2008;50:33–42. https://doi.org/10.1016/j.appet.2007.05.006.
Gottfredson NC, Sokol RL. Explaining excessive weight gain during early recovery from addiction. Subst Use Misuse. 2019;54:769–78. https://doi.org/10.1080/10826084.2018.1536722.
Nolan LJ, Scagnelli LM. Preference for sweet foods and higher body mass index in patients being treated in long-term methadone maintenance. Subst Use Misuse. 2007;42:1555–66. https://doi.org/10.1080/10826080701517727.
Gudzune KA, Doshi RS, Mehta AK, et al. Efficacy of commercial weight-loss programs: an updated systematic review. Ann Intern Med. 2015;162:501–12. https://doi.org/10.7326/m14-2238.
Diabetes Prevention Program (DPP) Research Group. The Diabetes Prevention Program (DPP): description of lifestyle intervention. Diabetes Care. 2002;25:2165–71. https://doi.org/10.2337/diacare.25.12.2165.
Elmaleh-Sachs A, Schwartz JL, Bramante CT, Nicklas JM, Gudzune KA, Jay M. Obesity management in adults: a review. JAMA. 2023;330:2000–15. https://doi.org/10.1001/jama.2023.19897.
Yanovski SZ, Yanovski JA. Approach to obesity treatment in primary care: a review. JAMA Intern Med. 2024;184:818–29. https://doi.org/10.1001/jamainternmed.2023.8526.
Bruns Vi N, Tressler EH, Vendruscolo LF, Leggio L, Farokhnia M. IUPHAR review - Glucagon-like peptide-1 (GLP-1) and substance use disorders: an emerging pharmacotherapeutic target. Pharmacol Res. 2024;207:107312. https://doi.org/10.1016/j.phrs.2024.107312.
Drucker DJ. GLP-1 physiology informs the pharmacotherapy of obesity. Mol Metab. 2022;57:101351. https://doi.org/10.1016/j.molmet.2021.101351.
Samuels JM, Patel MB, Niswender KD. Time to rethink the approach to treating obesity. JAMA Surg. 2024;159:841–2. https://doi.org/10.1001/jamasurg.2024.1502.
Holst JJ. The physiology of Glucagon-like peptide 1. Physiol Rev. 2007;87:1409–39. https://doi.org/10.1152/physrev.00034.2006.
Knudsen LB, Lau J. The discovery and development of liraglutide and semaglutide. Front Endocrinol (Lausanne). 2019;10:155. https://doi.org/10.3389/fendo.2019.00155.
Drucker DJ. Discovery of GLP-1-based drugs for the treatment of obesity. N Engl J Med. 2025;392:612–5. https://doi.org/10.1056/NEJMcibr2409089.
Jastreboff AM, Kaplan LM, Frías JP, Wu Q, Du Y, Gurbuz S, et al. Triple–hormone-receptor agonist retatrutide for obesity — a phase 2 trial. N Engl J Med. 2023;389:514–26. https://doi.org/10.1056/NEJMoa2301972.
Klausen MK, Thomsen M, Wortwein G, Fink-Jensen A. The role of glucagon-like peptide 1 (GLP-1) in addictive disorders. Br J Pharmacol. 2022;179:625–41. https://doi.org/10.1111/bph.15677.
Jerlhag E. The therapeutic potential of glucagon-like peptide-1 for persons with addictions based on findings from preclinical and clinical studies. Front Pharmacol. 2023;14:1063033. https://doi.org/10.3389/fphar.2023.1063033.
Fink-Jensen A, Wörtwein G, Klausen MK, Holst JJ, Hartmann B, Thomsen M, et al. Effect of the glucagon-like peptide-1 (GLP-1) receptor agonist semaglutide on alcohol consumption in alcohol-preferring male vervet monkeys. Psychopharmacology (Berl). 2025;242:63–70. https://doi.org/10.1007/s00213-024-06637-2.
Douton JE, Norgren R, Grigson PS. Effects of a glucagon-like peptide-1 analog on appetitive and consummatory behavior for rewarding and aversive gustatory stimuli in rats. Physiol Behav. 2021;229:113279. https://doi.org/10.1016/j.physbeh.2020.113279.
Bettadapura S, Dowling K, Jablon K, Al-Humadi AW, le Roux CW. Changes in food preferences and ingestive behaviors after glucagon-like peptide-1 analog treatment: techniques and opportunities. Int J Obes. 2025;49:418–26. https://doi.org/10.1038/s41366-024-01500-y.
Reddy IA, Stanwood GD, Galli A. Moving beyond energy homeostasis: new roles for glucagon-like peptide-1 in food and drug reward. Neurochem Int. 2014;73:49–55. https://doi.org/10.1016/j.neuint.2013.10.003.
Skibicka KP. The central GLP-1: implications for food and drug reward. Front Neurosci. 2013;7:181. https://doi.org/10.3389/fnins.2013.00181.
Eren-Yazicioglu CY, Yigit A, Dogruoz RE, Yapici-Eser H. Can GLP-1 Be a target for reward system related disorders? A qualitative synthesis and systematic review analysis of studies on palatable food, drugs of abuse, and alcohol. Front Behav Neurosci. 2020;14:614884. https://doi.org/10.3389/fnbeh.2020.614884.
Badulescu S, Tabassum A, Le GH, Wong S, Phan L, Gill H, et al. Glucagon-like peptide 1 agonist and effects on reward behaviour: a systematic review. Physiol Behav. 2024;283:114622. https://doi.org/10.1016/j.physbeh.2024.114622.
Guerrero-Hreins E, Goldstone AP, Brown RM, Sumithran P. The therapeutic potential of GLP-1 analogues for stress-related eating and role of GLP-1 in stress, emotion and mood: a review. Prog Neuropsychopharmacol Biol Psychiatry. 2021;110:110303. https://doi.org/10.1016/j.pnpbp.2021.110303.
Holt MK, Trapp S. The physiological role of the brain GLP-1 system in stress. Cogent Biol. 2016;2:1229086. https://doi.org/10.1080/23312025.2016.1229086.
Ghosal S, Myers B, Herman JP. Role of central glucagon-like peptide-1 in stress regulation. Physiol Behav. 2013;122:201–7. https://doi.org/10.1016/j.physbeh.2013.04.003.
Yaribeygi H, Rashidy-Pour A, Atkin SL, Jamialahmadi T, Sahebkar A. GLP-1 mimetics and cognition. Life Sci. 2021;264:118645. https://doi.org/10.1016/j.lfs.2020.118645.
Reich N, Hölscher C. The neuroprotective effects of glucagon-like peptide 1 in Alzheimer’s and Parkinson’s disease: an in-depth review. Front Neurosci. 2022;16:970925. https://doi.org/10.3389/fnins.2022.970925.
Kopp KO, Glotfelty EJ, Li Y, Lahiri DK, Greig NH. Type 2 diabetes mellitus/obesity drugs: a neurodegenerative disorders savior or a bridge too far? Ageing Res Rev. 2024;98:102343. https://doi.org/10.1016/j.arr.2024.102343.
Halloum W, Dughem YA, Beier D, Pellesi L. Glucagon-like peptide-1 (GLP-1) receptor agonists for headache and pain disorders: a systematic review. J Headache Pain. 2024;25:112. https://doi.org/10.1186/s10194-024-01821-3.
Thiele TE, Van Dijk G, Campfield LA, Smith FJ, Burn P, Woods SC, et al. Central infusion of GLP-1, but not leptin, produces conditioned taste aversions in rats. Am J Physiol Regul Integr Comp Physiol. 1997;272:R726–R730.
Diz-Chaves Y, Maastor Z, Spuch C, Lamas JA, González-Matías LC, Mallo F. Glucagon-like peptide 1 receptor activation: anti-inflammatory effects in the brain. Neural Regen Res. 2024;19:1671–7. https://doi.org/10.4103/1673-5374.389626.
Kopp KO, Glotfelty EJ, Li Y, Greig NH. Glucagon-like peptide-1 (GLP-1) receptor agonists and neuroinflammation: implications for neurodegenerative disease treatment. Pharmacol Res. 2022;186:106550. https://doi.org/10.1016/j.phrs.2022.106550.
Yoon G, Kim Y-K, Song J. Glucagon-like peptide-1 suppresses neuroinflammation and improves neural structure. Pharmacol Res. 2020;152:104615. https://doi.org/10.1016/j.phrs.2019.104615.
Klausen MK, Jensen ME, Møller M, Le Dous N, Jensen AØ, Zeeman VA, et al. Exenatide once weekly for alcohol use disorder investigated in a randomized, placebo-controlled clinical trial. JCI Insight. 2022;7. https://doi.org/10.1172/jci.insight.159863.
Probst L, Monnerat S, Vogt DR, Lengsfeld S, Burkard T, Meienberg A, et al. Effects of dulaglutide on alcohol consumption during smoking cessation. JCI Insight. 2023;8. https://doi.org/10.1172/jci.insight.170419.
Richards JR, Dorand MF, Royal K, Mnajjed L, Paszkowiak M, Simmons WK Significant decrease in alcohol use disorder symptoms secondary to semaglutide therapy for weight loss: a case series. J Clin Psychiatry. 2023;85. https://doi.org/10.4088/JCP.23m15068.
Quddos F, Hubshman Z, Tegge A, Sane D, Marti E, Kablinger AS, et al. Semaglutide and Tirzepatide reduce alcohol consumption in individuals with obesity. Sci Rep. 2023;13:20998. https://doi.org/10.1038/s41598-023-48267-2.
Bremmer MP, Hendershot CS. Social media as pharmacovigilance: the potential for patient reports to inform clinical research on glucagon-like peptide 1 (GLP-1) receptor agonists for substance use disorders. J Stud Alcohol Drugs. 2024;85:5–11. https://doi.org/10.15288/jsad.23-00318.
Arillotta D, Floresta G, Papanti Pelletier GD, Guirguis A, Corkery JM, Martinotti G, et al. Exploring the potential impact of GLP-1 receptor agonists on substance use, compulsive behavior, and libido: insights from social media using a mixed-methods approach. Brain Sci. 2024;14:617.
Wang W, Volkow ND, Berger NA, Davis PB, Kaelber DC, Xu R. Associations of semaglutide with incidence and recurrence of alcohol use disorder in real-world population. Nat Commun. 2024;15:4548. https://doi.org/10.1038/s41467-024-48780-6.
Farokhnia M, Tazare J, Pince CL, Bruns N 6th, Gray JC, Lo Re V 3rd, et al. Glucagon-like peptide-1 receptor agonists, but not dipeptidyl peptidase-4 inhibitors, reduce alcohol intake. J Clin Invest. 2025;135. https://doi.org/10.1172/jci188314.
Wium-Andersen IK, Wium-Andersen MK, Fink-Jensen A, Rungby J, Jørgensen MB, Osler M. Use of GLP-1 receptor agonists and subsequent risk of alcohol-related events. A nationwide register-based cohort and self-controlled case series study. Basic Clin Pharmacol Toxicol. 2022;131:372–9. https://doi.org/10.1111/bcpt.13776.
Qeadan F, McCunn A, Tingey B. The association between glucose-dependent insulinotropic polypeptide and/or glucagon-like peptide-1 receptor agonist prescriptions and substance-related outcomes in patients with opioid and alcohol use disorders: a real-world data analysis. Addiction. 2025;120:236–50. https://doi.org/10.1111/add.16679.
Lähteenvuo M, Tiihonen J, Solismaa A, Tanskanen A, Mittendorfer-Rutz E, Taipale H Repurposing semaglutide and liraglutide for alcohol use disorder. JAMA Psychiatry. 2024. https://doi.org/10.1001/jamapsychiatry.2024.3599.
Leggio L, Hendershot CS, Farokhnia M, Fink-Jensen A, Klausen MK, Schacht JP, et al. GLP-1 receptor agonists are promising but unproven treatments for alcohol and substance use disorders. Nat Med. 2023;29:2993–5. https://doi.org/10.1038/s41591-023-02634-8.
Volkow ND, Xu R GLP-1R agonist medications for addiction treatment. Addiction. 2024. https://doi.org/10.1111/add.16626.
Hendershot CS, Bremmer MP, Paladino MB, Kostantinis G, Gilmore TA, Sullivan NR, et al. Once-weekly semaglutide in adults with alcohol use disorder: a randomized clinical trial. JAMA Psychiatry. 2025;82:395–405. https://doi.org/10.1001/jamapsychiatry.2024.4789.
Farokhnia M, Faulkner ML, Piacentino D, Lee MR, Leggio L. Ghrelin: from a gut hormone to a potential therapeutic target for alcohol use disorder. Physiol Behav. 2019;204:49–57. https://doi.org/10.1016/j.physbeh.2019.02.008.
Richardson RS, Sulima A, Rice KC, Kucharczk JA, Janda KD, Nisbett KE, et al. Pharmacological GHSR (ghrelin receptor) blockade reduces alcohol binge-like drinking in male and female mice. Neuropharmacology. 2023;238:109643. https://doi.org/10.1016/j.neuropharm.2023.109643.
Faulkner ML, Farokhnia M, Lee MR, Farinelli L, Browning BD, Abshire K, et al. A randomized, double-blind, placebo-controlled study of a GHSR blocker in people with alcohol use disorder. JCI Insight. 2024;9. https://doi.org/10.1172/jci.insight.182331.
Horvath TL, Castañeda T, Tang-Christensen M, Pagotto U, Tschöp MH. Ghrelin as a potential anti-obesity target. Curr Pharm Des. 2003;9:1383–95. https://doi.org/10.2174/1381612033454748.
Chollet C, Meyer K, Beck-Sickinger AG. Ghrelin-a novel generation of anti-obesity drug: design, pharmacomodulation and biological activity of ghrelin analogues. J Pept Sci. 2009;15:711–30. https://doi.org/10.1002/psc.1177.
Gupta D, Ogden SB, Shankar K, Varshney S, Zigman JM. A LEAP 2 conclusions? Targeting the ghrelin system to treat obesity and diabetes. Mol Metab. 2021;46:101128. https://doi.org/10.1016/j.molmet.2020.101128.
Walker LC, Lawrence AJ. The role of orexins/hypocretins in alcohol use and abuse. Curr Top Behav Neurosci. 2017;33:221–46. https://doi.org/10.1007/7854_2016_55.
Moorman DE. The hypocretin/orexin system as a target for excessive motivation in alcohol use disorders. Psychopharmacology (Berl). 2018;235:1663–80. https://doi.org/10.1007/s00213-018-4871-2.
Cason AM, Smith RJ, Tahsili-Fahadan P, Moorman DE, Sartor GC, Aston-Jones G. Role of orexin/hypocretin in reward-seeking and addiction: implications for obesity. Physiol Behav. 2010;100:419–28. https://doi.org/10.1016/j.physbeh.2010.03.009.
Kotz C, Nixon J, Butterick T, Perez-Leighton C, Teske J, Billington C. Brain orexin promotes obesity resistance. Ann N Y Acad Sci. 2012;1264:72–86. https://doi.org/10.1111/j.1749-6632.2012.06585.x.
Perez-Leighton CE, Butterick-Peterson TA, Billington CJ, Kotz CM. Role of orexin receptors in obesity: from cellular to behavioral evidence. Int J Obes (Lond). 2013;37:167–74. https://doi.org/10.1038/ijo.2012.30.
Aranäs C, Caffrey A, Edvardsson CE, Vestlund J, Schmidt HD, Jerlhag E. Synergistic-like decreases in alcohol intake following combined pharmacotherapy with GLP-1 and amylin in male rats. Br J Pharmacol. 2025;182:1292–305. https://doi.org/10.1111/bph.17406.
Boukabara S, Farokhnia M, Leggio L. Amylin in alcohol addiction: a potential new treatment target or an adjuvant to other treatments? ACS Chem Neurosci. 2024;15:1609–10. https://doi.org/10.1021/acschemneuro.4c00133.
Walker CS, Aitken JF, Vazhoor Amarsingh G, Zhang S, Cooper GJS Amylin: emergent therapeutic opportunities in overweight, obesity and diabetes mellitus. Nat Rev Endocrinol. 2025. https://doi.org/10.1038/s41574-025-01125-9.
Boyle CN, Lutz TA, Le Foll C. Amylin - its role in the homeostatic and hedonic control of eating and recent developments of amylin analogs to treat obesity. Mol Metab. 2018;8:203–10. https://doi.org/10.1016/j.molmet.2017.11.009.
Flippo KH, Trammell SAJ, Gillum MP, Aklan I, Perez MB, Yavuz Y, et al. FGF21 suppresses alcohol consumption through an amygdalo-striatal circuit. Cell Metab. 2022;34:317–328.e6. https://doi.org/10.1016/j.cmet.2021.12.024.
Wang T, Tyler RE, Ilaka O, Cooper D, Farokhnia M, Leggio L. The crosstalk between fibroblast growth factor 21 (FGF21) system and substance use. iScience. 2024;27:110389. https://doi.org/10.1016/j.isci.2024.110389.
Sonoda J, Chen MZ, Baruch A FGF21-receptor agonists: an emerging therapeutic class for obesity-related diseases. Horm Mol Biol Clin Investig. 2017;30. https://doi.org/10.1515/hmbci-2017-0002.
Babaknejad N, Nayeri H, Hemmati R, Bahrami S, Esmaillzadeh A. An overview of FGF19 and FGF21: the therapeutic role in the treatment of the metabolic disorders and obesity. Horm Metab Res. 2018;50:441–52. https://doi.org/10.1055/a-0623-2909.
Padula CB, Tenekedjieva LT, McCalley DM, Al-Dasouqi H, Hanlon CA, Williams LM, et al. Targeting the salience network: a mini-review on a novel neuromodulation approach for treating alcohol use disorder. Front Psychiatry. 2022;13:893833. https://doi.org/10.3389/fpsyt.2022.893833.
Gouveia FV, Silk E, Davidson B, Pople CB, Abrahao A, Hamilton J, et al. A systematic review on neuromodulation therapies for reducing body weight in patients with obesity. Obes Rev. 2021;22:e13309. https://doi.org/10.1111/obr.13309.
Eisenberg D, Shikora SA, Aarts E, Aminian A, Angrisani L, Cohen RV, et al. 2022 American Society of Metabolic and Bariatric Surgery (ASMBS) and International Federation for the Surgery of Obesity and Metabolic Disorders (IFSO) indications for metabolic and bariatric surgery. Obes Surg. 2023;33:3–14. https://doi.org/10.1007/s11695-022-06332-1
American Society for Metabolic and Bariatric Surgery. Estimate of Bariatric Surgery Numbers, 2011-2022. Accessed Septemer 3, 2024. https://asmbs.org/resources/estimate-of-bariatric-surgery-numbers/.
Hampl SE, Hassink SG, Skinner AC, Armstrong SC, Barlow SE, Bolling CF, et al. Clinical practice guideline for the evaluation and treatment of children and adolescents with obesity. Pediatrics. 2023;151:e2022060640. https://doi.org/10.1542/peds.2022-060640.
Angrisani L, Santonicola A, Iovino P, Palma R, Kow L, Prager G, et al. IFSO Worldwide survey 2020–2021: current trends for bariatric and metabolic procedures. Obes Surg. 2024;34:1075–85. https://doi.org/10.1007/s11695-024-07118-3.
Madsbad S, Dirksen C, Holst JJ. Mechanisms of changes in glucose metabolism and bodyweight after bariatric surgery. Lancet Diabetes Endocrinol. 2014;2:152–64. https://doi.org/10.1016/s2213-8587(13)70218-3.
Hutch CR, Sandoval D. The role of GLP-1 in the metabolic success of bariatric surgery. Endocrinology. 2017;158:4139–51. https://doi.org/10.1210/en.2017-00564.
Mason EE, Ito C. Gastric bypass. Ann Surg. 1969;170:329–39. https://doi.org/10.1097/00000658-196909010-00003.
Navratil L. Alcoholism and gastric resection. Munch Med Wochenschr. 1959;101:1088–90. Alkoholismus und Magenresektion
Navratil L, Wenger R. [Gastrectomy and alcoholism]. Munch Med Wochenschr. 1957;99:546–50. Magenresektion und Trunksucht
Sogg S. Alcohol misuse after bariatric surgery: epiphenomenon or “Oprah” phenomenon? Surg Obes Relat Dis. 2007;3:366–8. https://doi.org/10.1016/j.soard.2007.03.004.
Thanos PK, Subrize M, Delis F, Cooney RN, Culnan D, Sun M, et al. Gastric bypass increases ethanol and water consumption in diet-induced obese rats. Obes Surg. 2012;22:1884–92. https://doi.org/10.1007/s11695-012-0749-2.
Davis JF, Tracy AL, Schurdak JD, Magrisso IJ, Grayson BE, Seeley RJ, et al. Roux en Y gastric bypass increases ethanol intake in the rat. Obes Surg. 2013;23:920–30. https://doi.org/10.1007/s11695-013-0884-4.
Hajnal A, Zharikov A, Polston JE, Fields MR, Tomasko J, Rogers AM, et al. Alcohol reward is increased after Roux-en-Y gastric bypass in dietary obese rats with differential effects following ghrelin antagonism. PLoS ONE. 2012;7:e49121. https://doi.org/10.1371/journal.pone.0049121.
Polston JE, Pritchett CE, Tomasko JM, Rogers AM, Leggio L, Thanos PK, et al. Roux-en-Y gastric bypass increases intravenous ethanol self-administration in dietary obese rats. PLoS ONE. 2013;8:e83741. https://doi.org/10.1371/journal.pone.0083741.
Orellana ER, Jamis C, Horvath N, Hajnal A. Effect of vertical sleeve gastrectomy on alcohol consumption and preferences in dietary obese rats and mice: a plausible role for altered ghrelin signaling. Brain Res Bull. 2018;138:26–36. https://doi.org/10.1016/j.brainresbull.2017.08.004.
Sirohi S, Skripnikova E, Davis JF. Vertical sleeve gastrectomy attenuates hedonic feeding without impacting alcohol drinking in rats. Obesity (Silver Spring). 2019;27:603–11. https://doi.org/10.1002/oby.22415.
Orellana E, Horvath N, Farokhnia M, Leggio L, Hajnal A. Changes in plasma ghrelin levels following surgical and non-surgical weight-loss in female rats predict alcohol use. Brain Res Bull. 2022;188:179–86. https://doi.org/10.1016/j.brainresbull.2022.07.019.
Blackburn AN, Hajnal A, Leggio L. The gut in the brain: the effects of bariatric surgery on alcohol consumption. Addict Biol. 2017;22:1540–53. https://doi.org/10.1111/adb.12436.
Saules KK, Wiedemann A, Ivezaj V, Hopper JA, Foster-Hartsfield J, Schwarz D. Bariatric surgery history among substance abuse treatment patients: prevalence and associated features. Surg Obes Relat Dis. 2010;6:615–21. https://doi.org/10.1016/j.soard.2009.12.008.
King WC, Chen JY, Mitchell JE, Kalarchian MA, Steffen KJ, Engel SG, et al. Prevalence of alcohol use disorders before and after bariatric surgery. JAMA. 2012;307:2516–25. https://doi.org/10.1001/jama.2012.6147.
Conason A, Teixeira J, Hsu CH, Puma L, Knafo D, Geliebter A. Substance use following bariatric weight loss surgery. JAMA Surg. 2013;148:145–50. https://doi.org/10.1001/2013.jamasurg.265.
King WC, Chen JY, Courcoulas AP, Dakin GF, Engel SG, Flum DR, et al. Alcohol and other substance use after bariatric surgery: prospective evidence from a U.S. multicenter cohort study. Surg Obes Relat Dis. 2017;13:1392–402. https://doi.org/10.1016/j.soard.2017.03.021.
Ostlund MP, Backman O, Marsk R, Stockeld D, Lagergren J, Rasmussen F, et al. Increased admission for alcohol dependence after gastric bypass surgery compared with restrictive bariatric surgery. JAMA Surg. 2013;148:374–7. https://doi.org/10.1001/jamasurg.2013.700.
Svensson PA, Anveden Å, Romeo S, Peltonen M, Ahlin S, Burza MA, et al. Alcohol consumption and alcohol problems after bariatric surgery in the Swedish obese subjects study. Obesity (Silver Spring). 2013;21:2444–51. https://doi.org/10.1002/oby.20397.
Bramming M, Becker U, Jørgensen MB, Neermark S, Bisgaard T, Tolstrup JS. Bariatric surgery and risk of alcohol use disorder: a Register-Based Cohort study. Int J Epidemiol. 2021;49:1826–35. https://doi.org/10.1093/ije/dyaa147.
Ibrahim N, Alameddine M, Brennan J, Sessine M, Holliday C, Ghaferi AA. New onset alcohol use disorder following bariatric surgery. Surg Endosc. 2019;33:2521–30. https://doi.org/10.1007/s00464-018-6545-x.
Wong E, Fleishman A, Brem A, Jones DB, Wee CC. High-risk alcohol use and disordered eating behavior before and 1 year after sleeve gastrectomy. Obes Surg. 2022;32:593–8. https://doi.org/10.1007/s11695-021-05847-3.
Maciejewski ML, Smith VA, Berkowitz TSZ, Arterburn DE, Mitchell JE, Olsen MK, et al. Association of bariatric surgical procedures with changes in unhealthy alcohol use among US veterans. JAMA Netw Open. 2020;3:e2028117. https://doi.org/10.1001/jamanetworkopen.2020.28117.
Coluzzi I, Iossa A, Spinetti E, Silecchia G. Alcohol consumption after laparoscopic sleeve gastrectomy: 1-year results. Eat Weight Disord. 2019;24:1131–6. https://doi.org/10.1007/s40519-018-0486-1.
Mahmud N, Panchal S, Abu-Gazala S, Serper M, Lewis JD, Kaplan DE. Association between bariatric surgery and alcohol use-related hospitalization and all-cause mortality in a veterans affairs cohort. JAMA Surg. 2023;158:162–71. https://doi.org/10.1001/jamasurg.2022.6410.
Speed SN, Sherman GT, Wang T, Pince CL, Sanfilippo JE, Montemitro C, et al. Bariatric surgery and alcohol and substance use: a case-control survey study. Drug Alcohol Depend. 2024;267:112529. https://doi.org/10.1016/j.drugalcdep.2024.112529.
Klockhoff H, Näslund I, Jones AW. Faster absorption of ethanol and higher peak concentration in women after gastric bypass surgery. Br J Clin Pharmacol. 2002;54:587–91. https://doi.org/10.1046/j.1365-2125.2002.01698.x.
Woodard GA, Downey J, Hernandez-Boussard T, Morton JM. Impaired alcohol metabolism after gastric bypass surgery: a case-crossover trial. J Am Coll Surg. 2011;212:209–14. https://doi.org/10.1016/j.jamcollsurg.2010.09.020.
Steffen KJ, Engel SG, Pollert GA, Li C, Mitchell JE. Blood alcohol concentrations rise rapidly and dramatically after Roux-en-Y gastric bypass. Surg Obes Relat Dis. 2013;9:470–3. https://doi.org/10.1016/j.soard.2013.02.002.
Pepino MY, Okunade AL, Eagon JC, Bartholow BD, Bucholz K, Klein S. Effect of Roux-en-Y gastric bypass surgery: converting 2 alcoholic drinks to 4. JAMA Surg. 2015;150:1096–8. https://doi.org/10.1001/jamasurg.2015.1884.
Acevedo MB, Eagon JC, Bartholow BD, Klein S, Bucholz KK, Pepino MY. Sleeve gastrectomy surgery: when 2 alcoholic drinks are converted to 4. Surg Obes Relat Dis. 2018;14:277–83. https://doi.org/10.1016/j.soard.2017.11.010.
Seyedsadjadi N, Acevedo MB, Alfaro R, Ramchandani VA, Plawecki MH, Rowitz B, et al. Site of alcohol first-pass metabolism among women. JAMA Netw Open. 2022;5:e223711 https://doi.org/10.1001/jamanetworkopen.2022.3711.
Acevedo MB, Teran-Garcia M, Bucholz KK, Eagon JC, Bartholow BD, Burd NA, et al. Alcohol sensitivity in women after undergoing bariatric surgery: a Cross-Sectional study. Surg Obes Relat Dis. 2020;16:536–44. https://doi.org/10.1016/j.soard.2020.01.014.
de Wit H, Bodker B, Ambre J. Rate of increase of plasma drug level influences subjective response in humans. Psychopharmacology (Berl). 1992;107:352–8. https://doi.org/10.1007/bf02245161.
National Institute on Alcohol Abuse and Alcoholism. What colleges need to know now. An update on college drinking research. Bethesda, MD: NIAAA; 2024. Available from: https://www.collegedrinkingprevention.gov/sites/cdp/files/documents/1College_Q7%20Bulletin-508_361C4E.pdf.
Linsenbardt DN, Moore EM, Griffin KD, Gigante ED, Boehm SL 2nd. Tolerance to ethanol’s ataxic effects and alterations in ethanol-induced locomotion following repeated binge-like ethanol intake using the DID model. Alcohol Clin Exp Res. 2011;35:1246–55. https://doi.org/10.1111/j.1530-0277.2011.01459.x.
Elvig SK, McGinn MA, Smith C, Arends MA, Koob GF, Vendruscolo LF. Tolerance to alcohol: a critical yet understudied factor in alcohol addiction. Pharmacol Biochem Behav. 2021;204:173155. https://doi.org/10.1016/j.pbb.2021.173155.
Changchien EM, Woodard GA, Hernandez-Boussard T, Morton JM. Normal alcohol metabolism after gastric banding and sleeve gastrectomy: a case-cross-over trial. J Am Coll Surg. 2012;215:475–9. https://doi.org/10.1016/j.jamcollsurg.2012.06.008.
Gallo AS, Berducci MA, Nijhawan S, Nino DF, Broderick RC, Harnsberger CR, et al. Alcohol metabolism is not affected by sleeve gastrectomy. Surg Endosc. 2015;29:1088–93. https://doi.org/10.1007/s00464-014-3790-5.
Jerlhag E. GLP-1 signaling and alcohol-mediated behaviors; preclinical and clinical evidence. Neuropharmacology. 2018;136:343–9. https://doi.org/10.1016/j.neuropharm.2018.01.013.
Jerlhag E, Egecioglu E, Landgren S, Salomé N, Heilig M, Moechars D, et al. Requirement of central ghrelin signaling for alcohol reward. Proc Natl Acad Sci USA. 2009;106:11318–23. https://doi.org/10.1073/pnas.0812809106.
Leggio L, Zywiak WH, Fricchione SR, Edwards SM, de la Monte SM, Swift RM, et al. Intravenous ghrelin administration increases alcohol craving in alcohol-dependent heavy drinkers: a preliminary investigation. Biol Psychiatry. 2014;76:734–41. https://doi.org/10.1016/j.biopsych.2014.03.019.
Davis JF, Schurdak JD, Magrisso IJ, Mul JD, Grayson BE, Pfluger PT, et al. Gastric bypass surgery attenuates ethanol consumption in ethanol-preferring rats. Biol Psychiatry. 2012;72:354–60. https://doi.org/10.1016/j.biopsych.2012.01.035.
Ivezaj V, Saules KK, Schuh LM. New-onset substance use disorder after gastric bypass surgery: rates and associated characteristics. Obes Surg. 2014;24:1975–80. https://doi.org/10.1007/s11695-014-1317-8.
Wee CC, Mukamal KJ, Huskey KW, Davis RB, Colten ME, Bolcic-Jankovic D, et al. High-risk alcohol use after weight loss surgery. Surg Obes Relat Dis. 2014;10:508–13. https://doi.org/10.1016/j.soard.2013.12.014.
Bradley D, Magkos F, Eagon JC, Varela JE, Gastaldelli A, Okunade AL, et al. Matched weight loss induced by sleeve gastrectomy or gastric bypass similarly improves metabolic function in obese subjects. Obesity (Silver Spring). 2014;22:2026–31. https://doi.org/10.1002/oby.20803.
Carroll ME, France CP, Meisch RA. Food deprivation increases oral and intravenous drug intake in rats. Science. 1979;205:319–21. https://doi.org/10.1126/science.36665.
Carr KD, Kim GY, Cabeza de Vaca S. Rewarding and locomotor-activating effects of direct dopamine receptor agonists are augmented by chronic food restriction in rats. Psychopharmacology (Berl). 2001;154:420–8. https://doi.org/10.1007/s002130000674.
Cheskin LJ, Hess JM, Henningfield J, Gorelick DA. Calorie restriction increases cigarette use in adult smokers. Psychopharmacology (Berl). 2005;179:430–6. https://doi.org/10.1007/s00213-004-2037-x.
Volkow ND, Wang GJ, Fowler JS, Telang F. Overlapping neuronal circuits in addiction and obesity: evidence of systems pathology. Philos Trans R Soc Lond B Biol Sci. 2008;363:3191–200. https://doi.org/10.1098/rstb.2008.0107.
Nance K, Acevedo MB, Pepino MY. Changes in taste function and ingestive behavior following bariatric surgery. Appetite. 2020;146:104423 https://doi.org/10.1016/j.appet.2019.104423.
White GE, Boles RE, Courcoulas AP, Yanovski SZ, Zeller MH, Jenkins TM, et al. A prospective cohort of alcohol use and alcohol-related problems before and after metabolic and bariatric surgery in adolescents. Ann Surg. 2023;278:e519–e525. https://doi.org/10.1097/sla.0000000000005759.
Mellinger JL, Shedden K, Winder GS, Fernandez AC, Lee BP, Waljee J, et al. Bariatric surgery and the risk of alcohol-related cirrhosis and alcohol misuse. Liver Int. 2021;41:1012–9. https://doi.org/10.1111/liv.14805.
Alvarado-Tapias E, Marti-Aguado D, Kennedy K, Fernández-Carrillo C, Ventura-Cots M, Morales-Arraez D, et al. Bariatric surgery is associated with alcohol-related liver disease and psychiatric disorders associated with AUD. Obes Surg. 2023;33:1494–505. https://doi.org/10.1007/s11695-023-06490-w.
Rubino F, Puhl RM, Cummings DE, Eckel RH, Ryan DH, Mechanick JI, et al. Joint international consensus statement for ending stigma of obesity. Nat Med. 2020;26:485–97. https://doi.org/10.1038/s41591-020-0803-x.
Volkow ND. Stigma and the toll of addiction. N Engl J Med. 2020;382:1289–90. https://doi.org/10.1056/NEJMp1917360.
Leshner AI. Addiction is a brain disease, and it matters. Science. 1997;278:45–7. https://doi.org/10.1126/science.278.5335.45.
Volkow ND, Koob GF, McLellan AT. Neurobiologic advances from the brain disease model of addiction. N Engl J Med. 2016;374:363–71. https://doi.org/10.1056/NEJMra1511480.
Heilig M, MacKillop J, Martinez D, Rehm J, Leggio L, Vanderschuren L. Addiction as a brain disease revised: why it still matters, and the need for consilience. Neuropsychopharmacol. 2021;46:1715–23. https://doi.org/10.1038/s41386-020-00950-y.
DiMartini AF, Leggio L, Singal AK. Barriers to the management of alcohol use disorder and alcohol-associated liver disease: strategies to implement integrated care models. Lancet Gastroenterol Hepatol. 2022;7:186–95. https://doi.org/10.1016/S2468-1253(21)00191-6.
McPheeters M, O’Connor EA, Riley S, Kennedy SM, Voisin C, Kuznacic K, et al. Pharmacotherapy for alcohol use disorder: a systematic review and meta-analysis. JAMA. 2023;330:1653–65. https://doi.org/10.1001/jama.2023.19761.
Han B, Jones CM, Einstein EB, Powell PA, Compton WM. Use of medications for alcohol use disorder in the US: results from the 2019 national survey on drug use and health. JAMA Psychiatry. 2021;78:922–4. https://doi.org/10.1001/jamapsychiatry.2021.1271.
Acknowledgements
The authors would like to thank Ms. Lauren Brick (Visual Media, NIDA IRP) for graphical design and Ms. Diane Cooper for editing and bibliographic assistance.
Funding
Drs. Leggio and Farokhnia are supported by the National Institutes of Health (NIH) Intramural Research Program funding ZIA-DA000635 (Section on Clinical Psychoneuroendocrinology and Neuropsychopharmacology, PI: Dr. Lorenzo Leggio) jointly supported by the National Institute on Drug Abuse (NIDA) Intramural Research Program (IRP) and the National Institute on Alcohol Abuse and Alcoholism (NIAAA) Division of Intramural Clinical and Biological Research (DICBR). Dr. Kenny is supported by NIDA/NIH, under Award Number R37DA020686. Dr. Pepino is supported by NIAAA/NIH, under Award Number R01AA024103. Dr. Simmons is supported by the Hardesty Family Foundation. This research was supported in part by the Intramural Research Program of the National Institutes of Health (NIH). The contributions of the NIH authors were made as part of their official duties as NIH federal employees, are in compliance with agency policy requirements, and are considered works of the United States Government. However, the findings and conclusions presented in this paper are those of the authors and do not necessarily reflect the views of the NIH or the U.S. Department of Health and Human Services. Open access funding provided by the National Institutes of Health.
Author information
Authors and Affiliations
Contributions
LL, MF, PJK, MYP and WKS jointly drafted and contributed to the writing and final approval of the paper.
Corresponding author
Ethics declarations
Competing interests
Dr. Leggio reports, outside his federal employment at the NIH, honoraria from the UK Medical Council on Alcohol (Editor-in-Chief for Alcohol and Alcoholism) and book royalties from Routledge (as editor of a textbook). Dr. Simmons serves on a clinical trial steering committee for Apollo Therapeutics, and is a site PI for a clinical trial sponsored by Altimmune Inc. The other authors report no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Leggio, L., Farokhnia, M., Kenny, P.J. et al. Crosstalk between alcohol use disorder and obesity: two sides of the same coin?. Mol Psychiatry 30, 5938–5952 (2025). https://doi.org/10.1038/s41380-025-03259-8
Received:
Revised:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41380-025-03259-8
