
Abstract
DEAD-box RNA helicase 3 (DDX3) and its homologs play a vital role in translation initiation by unwinding secondary structures of selected mRNAs. The human DDX3 gene is located on the sex chromosomes, so there are DDX3X and DDX3Y. DDX3X is ubiquitously expressed in almost all tissues and critical for embryonic development, whereas DDX3Y is only expressed in the testis and essential for male fertility. Drosophila belle (bel) is the single ortholog of DDX3, and mutations in bel cause male and female infertility. Using Drosophila bel mutants and Ddx3x conditional knockout (cKO) mice, we confirmed the pivotal role of DDX3 in female fertility and ovarian development. Drosophila bel mutants exhibited female infertility and immature egg chambers. Consistently, oocyte-specific Ddx3x knockout in mice resulted in female infertility and impaired oogenesis. We further found that immature egg chambers in Drosophila bel mutants and impaired follicular development in oocyte-specific Ddx3x cKO mice were caused by excessive apoptosis. We also identified a set of DDX3 target genes involved in oocyte meiosis and maturation and demonstrated that DDX3 is involved in their translation in human cells. Our results suggest that DDX3 is critical for female fertility via translational control in oogenesis.
Similar content being viewed by others

Introduction
The human DEAD-box protein DDX3 plays a crucial role in cellular RNA metabolism. DDX3 and its homologs have been implicated in multiple cellular processes, including translation initiation [1,2,3,4,5,6]. The yeast DDX3 homolog, Ded1, is required for translation of bulk mRNAs [1]. However, human DDX3 may affect the translation of only a subset of mRNAs. We previously reported that DDX3 is required for the translation of selected mRNAs that contain a long or structured 5′ UTR in human cells [3, 6]. Consistently, depletion of DDX3 in HCT116 cells represses translation of mRNAs with complex 5′ UTRs, such as those with high GC content or containing the cytosine-enriched regulator of translation (CERT) motif [5]. Given that the helicase activity of DDX3 is required for its function in translation [3], DDX3 may facilitate ribosome scanning by resolving secondary structures in the 5′ UTR of selected mRNAs.
DDX3 is a multifunctional protein implicated in a variety of biological processes, including cell cycle control [6,7,8], tumorigenesis and cancer progression [9,10,11,12], cell apoptosis [13,14,15], viral replication [16,17,18], innate immunity [19,20,21], and neurodevelopment [22,23,24]. The human DDX3 genes are located on both the X and Y chromosomes, so there are DDX3X and DDX3Y. Although DDX3X and DDX3Y share ~92% of the protein sequence identity and can functionally complement each other in translation [25], they show distinct expression patterns and biological functions [26]. DDX3X is ubiquitously expressed in almost all tissues and critical for embryonic development [7, 27], whereas DDX3Y is only expressed in the testis, especially in spermatogonia and early spermatocytes, and essential for male fertility [28]. The most common male infertility cases are caused by deletions in the Y chromosome’s azoospermia factor (AZF) regions [29]. Deletion analysis of the Y chromosome revealed three common deletions: AZFa, AZFb, and AZFc [30]. AZFa deletion causes the most severe azoospermia phenotype, a complete absence of germ cells in the seminiferous tubules of the testis, and Sertoli cell-only (SCO) syndrome in humans [31]. The human AZFa region contains three genes: USP9Y, DDX3Y, and UTY [32]. Uty knockout in mice did not affect fertility [33]. Loss of the human USP9Y gene did not cause male infertility [34], suggesting that USP9Y is not the causative gene. It has been reported that DDX3Y has a higher mutation rate in SCO syndrome than the other two genes in the AZFa region [35]. Overexpression of DDX3Y in AZFa-deleted iPSCs restored germ cell formation [36]. Therefore, DDX3Y is regarded as the major gene in AZFa deletion-induced spermatogenic failure [35].
Mouse Ddx3x mRNA is predominantly expressed in the ovary and embryo [37], implying that Ddx3x may play a role in the ovary and embryo. Conventional knockout of Ddx3x in mice causes early embryonic lethality [27]. Mouse Ddx3x protein is expressed in germinal vesicle (GV) oocytes, and its expression is remarkably increased in metaphase II oocytes [7]. This finding suggests a role for mouse Ddx3x in oocyte development. Drosophila belle (bel), the single ortholog of human DDX3, is required for larval growth and can functionally substitute for the yeast Ded1 in vivo [38], suggesting the functional conservation of DDX3 homologs. It has been reported that Drosophila bel mutants are male- and female-sterile [38]. Ectopic expression of bel ATPase mutants results in decreases in female fertility and ovary mass [39]. A recent study showed that DDX3 is required for ovarian development and oocyte maturation in Locusta migratoria [40]. These results reveal an evolutionarily conserved role of DDX3 in the development of germ cells. However, the molecular mechanisms of DDX3 in gametogenesis (spermatogenesis and oogenesis) remain largely unclear.
Here, we used Drosophila bel mutants and Ddx3x conditional knockout (cKO) mouse models to confirm the importance of DDX3 in female fertility and ovarian development. We previously identified many target mRNAs whose translation is regulated by DDX3 in HeLa cells [6]. Pathway enrichment analysis showed that ten candidate DDX3 target genes are involved in oocyte meiosis, oocyte maturation, and the gonadotropin-releasing hormone (GnRH) signaling pathway. Therefore, we assume that DDX3 is critical for female fertility via translational control in oogenesis.
Results
Drosophila bel mutants are female-sterile
Drosophila bel is the single ortholog of human DDX3. It has been reported that Drosophila bel mutations cause male and female infertility [38, 39]. We first need to verify the experimental results. The belneo30 is a hypomorphic mutation with a P-element insertion at the first intron of the bel gene. The bel6 is a lethal mutation with a point mutation that causes a premature stop codon. To verify whether Drosophila bel is required for female fertility, the belneo30/TM6B heterozygotes and the bel6/TM6B heterozygotes were crossed to produce the trans-heterozygous allelic combination belneo30/bel6. Western blot analysis showed that the expression of bel/DDX3 protein was markedly decreased in Drosophila belneo30/bel6 female mutants (Fig. 1A). Compared to the w1118 strain and heterozygous mutations (belneo30/TM6B and bel6/TM6B), Drosophila belneo30/bel6 females laid a lot fewer eggs, and did not produce any viable adults (Fig. 1B). The results indicate that Drosophila belneo30/bel6 mutants are female-sterile.

A Adult females were lysed in 1× RIPA Lysis Buffer and subjected to western blot analysis using antibodies against DDX3 and α-tubulin proteins. Detection of α-tubulin served as a loading control. B Three virgin females were mated with two w1118 males for 5 days. Fecundity was assessed by the number of eggs laid within 24 h. Fertility was evaluated by the number of offspring after 19 days of ovulation. The strain w1118 served as a control. Data are shown as mean and standard deviation (n = 10/group). C Adult females were fed diets containing yeast for 3 days and then dissected to collect their ovaries. Body length and ovary size were measured and recorded under a dissecting microscope. Data are shown as mean and standard deviation (n = 10/group). Statistical significance was determined using the Student’s t-test (ns not significant; **p < 0.01; ****p < 0.0001).
Ovarian development is impaired in Drosophila bel mutants
In Drosophila, body size is one of the most determining traits in sexual selection [41]. The size of the Drosophila body was measured and recorded under a dissecting microscope. There was no significant change in the mean length of the Drosophila body in belneo30/bel6 female mutants compared to the w1118 strain or single-allele mutations (belneo30/TM6B and bel6/TM6B) (Fig. 1C). To understand why Drosophila belneo30/bel6 mutants are female-sterile, we dissected adult females to observe their ovaries. An adult female has two ovaries. Notably, we observed that the size and volume of the ovaries were much smaller in belneo30/bel6 female mutants compared to the w1118 strain or single-allele mutations (Fig. 1C). The results suggest that Drosophila bel/DDX3 plays a critical role in ovarian development.
Apoptosis results in immature egg chambers in Drosophila bel mutants
To explore why Drosophila belneo30/bel6 mutants lose female fertility, dissected ovaries were stained with DAPI and observed using a fluorescence microscope. We did not find mature oocytes in belneo30/bel6 (Fig. 2A). The disruption of nuclei in nurse cells (NCs) of egg chambers was noticeable (Fig. 2A). We therefore speculate that apoptosis of egg chambers may occur in the development of the belneo30/bel6 ovaries, thus resulting in the arrest of oocyte maturation. To test this hypothesis, we performed immunofluorescence (IF) and terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) staining to detect apoptosis in Drosophila ovaries. Cleaved Dcp-1(cDcp-1) in Drosophila, a homolog of mammalian caspase 3, is a specific marker of apoptosis [42]. Apoptotic cells can be labeled with cDcp-1 and TUNEL in Drosophila tissues [43]. The results of IF and TUNEL staining showed that massive apoptosis of egg chambers occurs in the belneo30/bel6 ovaries compared to the w1118 ovaries (Fig. 2B, C). Therefore, mutations in Drosophila bel resulted in apoptosis of egg chambers.

Adult females (w1118 and belneo30/bel6) were fed diets containing yeast for 3 days and then dissected to collect their ovaries. Ovaries were fixed with 4% formaldehyde and washed with 1× PBS containing 0.3% Triton X-100. A DAPI staining shows the presence of nuclei in Drosophila ovaries. NCs: nurse cells. B Ovaries were permeabilized with 1% Triton X-100. Immunofluorescence staining was performed using antibodies against cleaved Dcp-1 (cDcp-1), commonly used as a marker of apoptosis in Drosophila. DAPI staining shows the presence of nuclei in ovaries. C Ovaries were permeabilized with 1% Triton X-100. TUNEL assay was performed to detect apoptotic DNA fragmentation in Drosophila ovaries. DAPI staining shows the presence of nuclei in ovaries. Scale bars, 100 μm.
Generation of oocyte-specific Ddx3x conditional knockout (cKO) mice
As mentioned above, conventional knockout of Ddx3x in mice results in early embryonic lethality [27]. We therefore used oocyte-specific Ddx3x cKO mice to study the importance of Ddx3x in oogenesis. A Ddx3x cKO mouse model, in which the Ddx3x gene is flanked with loxP sites using a CRISPR/Cas9-based gene editing, has been established (Fig. 3A). According to the Cre-loxP system [44], gene deletion can be precisely controlled by spatiotemporally defined Cre recombinase expression [45]. A transgenic mouse line Zp3-Cre in which Cre recombinase expression is controlled by the promoter of the zona pellucida 3 (Zp3) gene, which is exclusively expressed in the growing oocytes [46]. The Zp3-Cre line is used to delete the specific gene in oocytes [47]. To generate oocyte-specific Ddx3x cKO female mice (Ddx3xloxP/loxP; Zp3-Cre), heterozygous Ddx3xloxP mice were inbred to produce homozygous Ddx3xloxP/loxP female mice and then crossed with Zp3-Cre male mice (Fig. 3B). Oocyte-specific Ddx3x cKO female mice were confirmed by PCR-based genotyping (Fig. 3C).

A The loxP site (ATAACTTCGTATAATGTATGCTATACGAAGTTAT), followed by an ApaI restriction site (GGGCCC), was inserted into intron 1 and intron 10 of the Ddx3x gene using a CRISPR/Cas9-based gene editing strategy. The length of a PCR product with a loxP site will increase by 40 bp, and it can be cut into two fragments by the ApaI restriction enzyme. B Diagram of breeding strategy for generation of Cre-loxP-mediated oocyte-specific Ddx3x cKO mice. C Homozygous Ddx3xloxP/loxP female mice were detected by PCR product (636 bp) compared to wild-type mice (596 bp). Zp3-Cre mice were detected by PCR product (~300 bp). Among the 13 female mice, 7 were oocyte-specific Ddx3x cKO mice (Ddx3xloxP/loxP; Zp3-Cre) (red circle).
Female infertility in oocyte-specific Ddx3x cKO mice
To examine whether oocyte-specific deletion of Ddx3x affects female fertility, the fertility test was carried out twice to reduce experimental errors. The number of offspring produced by mating 10-week-old Ddx3xloxP/loxP or Ddx3xloxP/loxP; Zp3-Cre female mice with normal C57BL/6 male mice were counted. Male mice were randomly assigned to female mice. A female mouse is considered to be fertile if she gives birth to pups. The number of offspring from each pregnancy was recorded after birth. The average litter size was evaluated in Ddx3xloxP/loxP; Zp3-Cre female mice compared to Ddx3xloxP/loxP female mice. As expected, Ddx3xloxP/loxP; Zp3-Cre female mice did not give birth to any pups, whereas Ddx3xloxP/loxP female mice had an average of 7–8 pups per litter (Fig. 4A). The results indicated that oocyte-specific Ddx3x cKO mice are female-sterile.

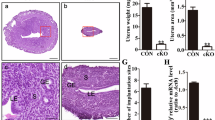
A Ten-week-old Ddx3xloxP/loxP female mice (n = 9) and Ddx3xloxP/loxP; Zp3-Cre female mice (n = 10) were mated with wild-type C57BL/6 male mice for 10 days (1 female:1 male). Females were then separated from the males and allowed to rest for 21 days, the average gestation period in mice. The number of pups was counted after birth. Fertility tests were carried out twice. Data are shown as mean ± SD. B Ten-week-old Ddx3xloxP/loxP female mice (n = 6) and Ddx3xloxP/loxP; Zp3-Cre female mice (n = 6) were dissected to collect their uteruses (with ovaries). The weight of the body and uterus (with ovaries) was measured and recorded. The ratio of uterus weight to body weight of mice was calculated. Data are shown as mean ± SD. C Experimental mice are as described in (B). Mouse ovaries were observed under a dissecting microscope. Representative images of mouse ovaries were shown. D Experimental mice are as described in (B). Mouse ovaries were fixed in formalin, embedded in paraffin, and then sectioned at a thickness of 5 μm. Ovarian sections were stained with hematoxylin and eosin (H&E). Representative images of H&E-stained ovarian sections were shown. E Experimental mice were prepared and treated as described in (D). The number of follicles in the ovarian sections at different stages (primordial, primary, secondary, and antral) was counted. The bar graph shows the number of follicles at different stages in the ovaries of oocyte-specific Ddx3x cKO mice (Ddx3xloxP/loxP; Zp3-Cre) compared to control mice (Ddx3xloxP/loxP). Data are shown as mean and standard deviation (n = 6/group). Statistical significance was determined using the Student’s t-test (ns not significant; *p < 0.05; **p < 0.01; ***p < 0.001).
Follicular development is impaired in oocyte-specific Ddx3x cKO mice
There was no significant difference in appearance and body weight between Ddx3xloxP/loxP and Ddx3xloxP/loxP; Zp3-Cre female mice (Fig. 4B). To understand why oocyte-specific Ddx3x cKO mice lose female fertility, we dissected Ddx3xloxP/loxP and Ddx3xloxP/loxP; Zp3-Cre female mice to observe their ovaries. The weight of the uterus (with ovaries) was measured and recorded. The uterus index, defined as the ratio of uterus weight to body weight of mice, was calculated and compared to the control group. There was only a slight decrease but no significant difference in the uterus index of Ddx3xloxP/loxP; Zp3-Cre female mice compared to Ddx3xloxP/loxP female mice (Fig. 4B). In contrast, the size of the ovaries of Ddx3xloxP/loxP; Zp3-Cre female mice were much smaller than that of Ddx3xloxP/loxP female mice (Fig. 4C). The ovaries of Ddx3xloxP/loxP and Ddx3xloxP/loxP; Zp3-Cre female mice were collected, fixed in formalin, and embedded in paraffin for long-term storage. Follicular development was observed in hematoxylin and eosin (H&E)-stained ovarian sections. Notably, it is difficult to find secondary and antral follicles in the ovaries of Ddx3xloxP/loxP; Zp3-Cre female mice (Fig. 4D). The ovarian tissue of Ddx3xloxP/loxP; Zp3-Cre female mice showed the absence of corpus luteum (Fig. 4D). The accumulation of primary follicles in oocyte-specific Ddx3x cKO female mice suggested that deletion of Ddx3x inhibits the primary to secondary follicle transition (Fig. 4E). The results supported that Ddx3x is required for the development of ovaries and mature follicles in female mice. However, the molecular mechanisms of DDX3 in ovarian development and oogenesis remain to be further studied.
Extensive apoptosis occurs in the ovarian follicles of oocyte-specific Ddx3x cKO female mice
We also performed immunohistochemistry (IHC) staining with DDX3 antibody to detect oocyte-specific knockout of Ddx3x in the ovaries of Ddx3xloxP/loxP; Zp3-Cre female mice. However, the IHC staining of Ddx3x in ovarian oocytes is too light to distinguish between Ddx3xloxP/loxP and Ddx3xloxP/loxP; Zp3-Cre female mice (Fig. 5A). Among the observed ovarian sections of oocyte-specific Ddx3x cKO female mice, only one antral follicle was seen (Fig. 5A). Notably, oocyte collapse/degeneration was observed in the ovaries of oocyte-specific Ddx3x cKO female mice (Fig. 5A). The phenotype of oocyte collapse/degeneration suggested that cells may undergo apoptosis. To examine whether oocyte-specific deletion of Ddx3x results in apoptosis in the ovarian follicles, we performed IHC and TUNEL staining to detect apoptosis in mouse ovarian sections. Cleaved caspase 3 is a specific marker of apoptosis in mouse oocytes [48]. IHC staining with cleaved caspase 3 antibody and TUNEL assay were performed to detect apoptotic cells in mouse ovarian sections. The results of cleaved caspase 3 and TUNEL staining showed that extensive apoptosis occurs in ovarian sections of Ddx3xloxP/loxP; Zp3-Cre female mice compared to Ddx3xloxP/loxP female mice (Fig. 5B, C). Therefore, loss of Ddx3x in mouse oocytes results in a female infertility phenotype due to apoptosis of ovarian follicles.

10-week-old Ddx3xloxP/loxP female mice (n = 6) and Ddx3xloxP/loxP; Zp3-Cre female mice (n = 6) were dissected to collect their ovaries. Mouse ovaries were fixed in formalin, embedded in paraffin, and then sectioned at a thickness of 5 μm. A Ovarian sections were processed for immunohistochemistry (IHC) to stain endogenous mouse Ddx3x. Representative images of IHC-stained ovarian sections were shown. An antral follicle and nearby degenerated oocytes were enlarged in Ddx3xloxP/loxP; Zp3-Cre ovarian sections. B Ovarian sections were processed for immunohistochemistry (IHC) to stain cleaved caspase 3. Representative images of IHC-stained ovarian sections were shown. Brown signals were frequently observed in Ddx3xloxP/loxP; Zp3-Cre ovarian sections. C TUNEL assay was performed to detect apoptotic DNA fragmentation in mouse ovarian sections. DAPI staining shows the presence of nuclei in mouse ovaries. Scale bars, 100 μm.
DDX3-regulated candidate genes are involved in oogenesis
Because it is difficult to collect mouse oocytes from oocyte-specific Ddx3x cKO female mice, we re-analyzed a set of translational targets of DDX3 that were previously identified in HeLa cells [6]. To gain insight into the biological functions of DDX3, pathway enrichment analysis was performed using the DAVID Bioinformatics Resources software (https://david.ncifcrf.gov/) to map genes to biological pathways defined by the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. The result showed that a set of candidate genes are involved in oocyte meiosis, progesterone-mediated oocyte maturation, and the GnRH signaling pathway (Table 1). These candidate DDX3 target genes include APC1, CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, and GNAS (Table 1). Since these candidate genes have been implicated in oocyte meiosis and maturation (Table 2), we speculate that DDX3 has a role in oogenesis.
DDX3 regulates the protein expression of oogenesis-associated genes in HeLa cells
We identified a set of DDX3 target genes involved in oogenesis from HeLa cells (Table 1). Next, we evaluated whether DDX3 regulates their protein expression. Using short hairpin RNAs (shDDX3-1 and shDDX3-2), we observed that the level of DDX3 protein was considerably reduced in HeLa cells (Fig. 6A). The protein levels of APC1, cyclin E1, PP2Aβ, PKACα, FBXW11, Rac1, and GNAS were prominently decreased in DDX3 knockdown HeLa cells compared to mock-treated cells (pLKO.1) (Fig. 6A). However, the level of p38 MAPK and B56β proteins were moderately decreased. The results suggest that DDX3 positively regulates the expression of APC1, cyclin E1, p38 MAPK, B56β, PP2Aβ, PKACα, FBXW11, Rac1, and GNAS proteins in HeLa cells. To examine whether the knockdown of DDX3 affects the expression of APC1, CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, and GNAS mRNAs, we used quantitative real-time RT-PCR to detect the mRNA levels. The results showed that the expression of DDX3 mRNA was dramatically depleted by shDDX3-1 and shDDX3-2 in HeLa cells (Fig. 6B). There was no significant change in the mRNA levels of APC1, PP2Aβ, PKACα, and GNAS in DDX3 knockdown HeLa cells compared to mock-treated cells (Fig. 6B). However, CCNE1 and RAC1 mRNAs were significantly decreased in DDX3 knockdown HeLa cells, suggesting that DDX3 knockdown may affect the transcription or stability of CCNE1 and RAC1 mRNAs. Since depletion of DDX3 does not decrease the mRNA expression of most candidate DDX3 target genes, DDX3 may promote their expression at the translation level.

HeLa cells were transduced with the empty lentiviral vector (pLKO.1) or the pLKO.1 vector expressing DDX3 shRNAs (shDDX3-1 and shDDX3-2). Cells were harvested for analysis at day 3 post-transduction. A Western blot analysis was performed using antibodies against DDX3, APC1, cyclin E1, p38 MAPK, B56β, PP2Aβ, PKACα, FBXW11, Rac1, GNAS, and α-tubulin proteins. Detection of α-tubulin served as a loading control. B The expression of APC1, CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, GNAS, DDX3, and GAPDH mRNAs were detected by quantitative real-time RT-PCR. The bar graph shows the relative mRNA levels normalized to GAPDH as mean and standard deviation from three independent experiments. Statistical significance was tested by one-way ANOVA (ns not significant; **p < 0.01; ***p < 0.001; ****p < 0.0001).
Translational efficiency of CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, and GNAS mRNAs is decreased in DDX3 knockdown 293T cells
To verify whether DDX3 regulates the translation of its target mRNAs, we performed sucrose gradient sedimentation and polysome profile analysis to evaluate the translational status of DDX3-knockdown and mock-treated 293T cells. As in HeLa cells (Fig. 6), shDDX3-1 significantly downregulated the protein level of DDX3 in 293T cells (Fig. 7A). Translational efficiency of a transcript was defined as the ratio of the mRNA abundance in the polysome fractions to the mRNA abundance in the total fractions [49]. We assessed translational efficiency of APC1, CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, GNAS, and GAPDH mRNAs by polysome profiling and quantitative real-time RT-PCR. Among all transcripts examined, only APC1 and GAPDH mRNAs were not significantly decreased in DDX3 knockdown 293T cells compared to mock-treated cells (Fig. 7B). Therefore, DDX3 may regulate translation of CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, and GNAS mRNAs in 293T cells. Since depletion of DDX3 does not significantly affect translational efficiency of APC1 mRNA, the downregulation of APC1 protein in DDX3 knockdown 293T cells may result from protein destabilization. However, it remains to be studied further.

293T cells were transduced with the empty lentiviral vector (pLKO.1) or the pLKO.1 vector expressing shDDX3-1. Cells were harvested for analysis at day 3 post-transduction. A Western blot analysis was performed using antibodies against DDX3 and α-tubulin proteins. Detection of α-tubulin served as a loading control. B Cytoplasmic extracts were loaded on a linear 15-40% sucrose gradient ultracentrifugation. Total RNA was extracted from each fraction for analysis. Polysome profile analysis and quantitative real-time RT-PCR were performed to assess translational efficiency of APC1, CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, GNAS, and GAPDH mRNAs in DDX3 knockdown 293T cells (shDDX3-1) compared to control cells (pLKO.1). Detection of GAPDH mRNA served as a negative control. The bar graph shows the changes in translational efficiency as mean and standard deviation from three independent experiments. Statistical significance was determined using the Student’s t-test (ns not significant; *p < 0.05; **p < 0.01; ***p < 0.001).
DDX3 may facilitate the translation of oogenesis-related target mRNAs by resolving their 5′ UTRs
We have reported that DDX3 is required for efficient translation of selected mRNAs that contain a long or structured 5′ UTR [3, 6]. Computational analysis showed that oogenesis-related DDX3 target mRNAs contain long and GC-rich 5′ UTRs that are likely to form stable secondary structures; their average length is 300 nucleotides (nt), with a GC content of 78.5% (Fig. 8A). In contrast, the average length of 5′ UTRs in randomly selected mRNAs is only about 246 nt, with a GC content of 63% [6]. Secondary structures were predicted using the RNAfold web server in ViennaRNA Web Services (http://rna.tbi.univie.ac.at/cgi-bin/ RNAWebSuite/RNAfold.cgi). The RNAfold program was used to calculate the minimum free energy (MFE) of RNA secondary structures (Fig. 8A). The secondary structures within the 5′ UTRs of candidate DDX3 target mRNAs are shown in Fig. 8B. To determine whether DDX3 participates in ribosome scanning by resolving secondary structures in the 5′ UTR of candidate target mRNAs, we performed the dual-luciferase reporter assay in 293T cells. Firefly luciferase (Fluc) reporter genes containing the 5′ UTRs of the selected candidates were constructed. A Renilla luciferase (Rluc) reporter with an unstructured 5′ UTR was co-transfected with the Fluc reporter as an internal control. The translation activity of the reporter mRNAs was assessed using the firefly to Renilla luciferase activity ratio (Fluc/Rluc). The results showed that the relative Fluc/Rluc activity of reporters was significantly decreased in DDX3 knockdown 293T cells compared to mock-treated cells (Fig. 8C). Therefore, DDX3 may facilitate translation of oogenesis-related target mRNAs by resolving secondary structures within the 5′ UTRs.

A The 5′ UTR sequences of APC1, CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, and GNAS mRNAs obtained from the NCBI Reference Sequence (RefSeq) database were analyzed. The length, GC content, and minimum free energy (MFE) of the 5′ UTR sequences were indicated. B Secondary structures within the 5′ UTRs were predicted using the RNAfold web server. The nucleotides are colored according to their probabilities in the structure. C 293T cells were transduced with the empty lentiviral vector (pLKO.1) or the pLKO.1 vector expressing shDDX3-1. After 48 h, 293T cells were co-transfected with firefly luciferase (Fluc) reporters containing the 5′ UTRs of APC1, CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, or GNAS mRNAs in combination with the control pRL-SV40 vector encoding the Renilla luciferase (Rluc). 293T cells were lysed for analysis at 24 h post transfection. The Fluc activity was normalized for each transfectant to that of the Rluc control. The bar graph shows the relative Fluc/Rluc activities in DDX3 knockdown 293T cells (shDDX3-1) compared to control cells (pLKO.1). Data are shown as mean and standard deviation from three independent experiments. Statistical significance was determined using the Student’s t-test (**p < 0.01; ***p < 0.001; ****p < 0.0001).
Discussion
Although the human DDX3Y gene is widely transcribed in various tissues, the DDX3Y protein is only detected in male germ cells (spermatogonia) [26]. Loss of the DDX3Y gene is the most likely cause of the SCO syndrome, observed in AZFa deletion-induced spermatogenic failure [26, 35]. The SCO syndrome is one of the most common causes of male infertility, and the patients have very low or absent spermatogenesis (oligozoospermia and azoospermia). More recently, DDX3Y was identified as the critical spermatogenic factor in AZFa deletion-induced azoospermia [50]. Infertility patients with loss-of-function mutations in the DDX3Y gene presented the SCO phenotype, suggesting that DDX3Y is essential for spermatogenesis and male fertility [50]. However, the molecular mechanisms by which DDX3Y functions in spermatogenesis remain largely unclear.
Here, we identified several DDX3-regulated target genes involved in oocyte meiosis and maturation (Tables 1 and 2), suggesting a role for DDX3 in oogenesis. These DDX3-regulated target genes may act directly on oocyte meiosis or play a regulatory role during the processes of oogenesis. Meiosis occurs during the process of spermatogenesis and oogenesis. It is conceivable that loss of DDX3Y may affect sperm meiosis and maturation (spermatogenesis). This study used Drosophila bel mutants and oocyte-specific Ddx3x cKO mice to determine that DDX3X is required for female fertility and ovarian development. Drosophila bel is the single ortholog of human DDX3, and bel mutations cause male and female infertility [38]. Similarly, the ortholog of DDX3 in Locusta migratoria is ubiquitously expressed in various tissues with a high abundance in the testis and ovary [40]. Depletion of LmDDX3 also blocked ovarian development and oocyte maturation [40]. In mice, there are three homologs of the DDX3 subfamily: Ddx3x, Ddx3y, and Pl10. Mouse Ddx3x exhibits 99% similarity to human DDX3X [7]. Mouse Ddx3x is predominantly expressed in the ovary and embryo [37], suggesting that Ddx3x is required for ovarian and embryonic development [27]. Interestingly, the expression of mouse Ddx3x protein is remarkably increased in metaphase II oocytes [7]. This finding is consistent with the requirement of Ddx3x for oocyte meiosis and oogenesis.
Recently, Ddx3y KO male mice were generated using CRISPR/Cas9 but showed normal spermatogenesis [51]. It was proposed that mouse Ddx3y may be dispensable in germ cells for male fertility. Nevertheless, the mouse contains an autosomal homolog of DDX3, Pl10, which is thought to be a pseudogene in humans [52]. Mouse Pl10 is predominantly expressed in meiotic spermatocytes [37]. Pl10-deficient male mice were shown to be infertile [53], suggesting a role for Pl10 in spermatogenesis. Pl10 may replace the role of Ddx3y during spermatogenesis in mice.
Although mutations in the bel gene do not affect the size of the Drosophila body, the size and volume of the ovaries are obviously reduced in Drosophila bel mutants compared to the control group (Fig. 1C). The results indicate that Drosophila bel/DDX3 plays a critical role in ovarian development. Consistently, oocyte-specific deletion of Ddx3x in mice does not significantly affect the body weight and uterus index (Fig. 4B). The size of the ovaries is obviously reduced in oocyte-specific Ddx3x cKO mice compared to the control group (Fig. 4C). It is surprised that only deletion of Ddx3x from oocytes results in impaired ovarian development, indicating the importance of maintaining oocyte integrity for the development of follicles and ovaries. Oocyte-specific deletion of Ddx3x causes extensive apoptosis in oocytes and adjacent granulosa cells (Fig. 5). This result demonstrates that precise communication between oocytes and granulosa cells is vital for follicle and ovarian development.
Although DDX3 can bind to most mRNAs, only a subset of mRNAs is predominantly affected by DDX3 knockdown at the translation level [5, 6]. DDX3 prefers to bind to 5′ UTRs of mRNAs and 18S rRNA [54], supporting its role in ribosome scanning during translation initiation. DDX3 regulates translation of mRNAs with complex 5′ UTRs, such as high GC content [3, 5, 6], cytosine-enriched regulator of translation (CERT) motif [5], and upstream open reading frame (uORF) [55]. Although less prevalent than in the 5′ UTR, DDX3 also binds to the coding sequence (CDS) and the 3′ UTR of mRNAs [56, 57]. It has been recently reported that DDX3 target mRNAs have high GC content in the CDS [5]. This may explain why translational efficiency of PKACα mRNA decreased significantly in DDX3 knockdown cells (Fig. 7B). However, the Fluc reporter with PKACα 5′ UTR is less sensitive to DDX3 knockdown (Fig. 8C). There is a high GC region at the beginning of the CDS of PKACα mRNA, and it may also involve DDX3-mediated translational control. DDX3 was recently shown to control translation of specific mRNAs through the 3′ UTR [56, 57]. Therefore, DDX3-mediated translational control may have different mechanisms.
DDX3 was identified as a component of an anti-apoptotic protein complex associated with death receptors [58]. Loss of Ddx3x results in cell cycle arrest and apoptosis in mouse embryos [27], suggesting a role for DDX3 in anti-apoptosis. DDX3 protects cells from sanguinarine-induced intrinsic apoptosis in HeLa cells [14]. After sanguinarine treatment, DDX3 upregulates the expression of anti-apoptotic gene (Bcl-xL) and downregulates the expression of pro-apoptotic genes (CASP3 and BAX) [14]. Consistent with previous studies, we also found that DDX3 interacts with USP9X and exerts anti-apoptotic effects by deubiquitinating and stabilizing MCL1 [15]. Depletion of DDX3 results in increased cell apoptosis [15], supporting a role for DDX3 in anti-apoptosis. DDX3 may activate the anti-apoptotic Bcl-xL gene and repress the pro-apoptotic CASP3 and BAX genes through transcriptional or translational regulation. DDX3 also stabilizes anti-apoptotic MCL1 protein through USP9X-mediated MCL1 deubiquitination [15]. However, the molecular mechanism of DDX3 in anti-apoptosis remains to be further studied.
Materials and methods
Drosophila stocks
Drosophila bel mutants belneo30 and bel6 were obtained from the Bloomington Drosophila Stock Center (BDSC) at Indiana University. Dr. Haiwei Pi provided the control strain w1118. All fly stocks were maintained and raised at 25 °C.
Western blot analysis
Proteins were transferred onto a polyvinylidene difluoride (PVDF) transfer membrane (Millipore). Protein blots were blocked with 3% skim milk in TBST (100 mM Tris–HCl [pH 7.6], 150 mM NaCl, and 0.1% Tween-20) at room temperature for 1 h. The primary antibodies included affinity-purified rabbit anti-DDX3 (0.1 μg/ml) [3], mouse anti-α-tubulin (0.2 μg/ml; sc-32293, Santa Cruz Biotechnology), rabbit anti-APC1 (1:1000 dilution; #13329, Cell Signaling), mouse anti-cyclin E1 (0.2 μg/ml; sc-247, Santa Cruz Biotechnology), rabbit anti-p38 MAPK (1:1000 dilution; #8690, Cell Signaling), mouse anti-PP2A-B56-β (0.4 μg/ml; sc-515676, Santa Cruz Biotechnology), rabbit anti-PP2Aβ (1:1000 dilution; GTX101690, GeneTex), rabbit anti-PKACα (1:1000 dilution; GTX104934, GeneTex), rabbit anti-FBXW11 (1:1000 dilution; GTX33193, GeneTex), mouse anti-Rac1 (1:1000 dilution; 05-389, Millipore), and rabbit anti-GNAS (1:1000 dilution; GTX113200, GeneTex). Blots were incubated with primary antibodies in the blocking buffer at room temperature for 2 h, followed by horseradish peroxidase (HRP)-conjugated secondary antibodies at room temperature for 2 h. Signals were detected using Immobilon Western chemiluminescent HRP substrate (Millipore), and images from autoradiograms were captured with the ChemiDoc touch imaging system (Bio-Rad).
Drosophila fecundity and fertility tests
Three young virgin females (0–1 day old) were mated with two w1118 males for five days. After mating, females were transferred to an agar plate (9-cm Petri dish) for the fecundity test. After 24 h of ovulation, the number of eggs was observed and calculated with a dissecting microscope. For the fertility test, females were transferred to a fresh culture tube after mating. Adult females were removed 24 h after spawning, and eggs were incubated for 19 days. The number of adult offspring was counted.
DAPI and immunofluorescence staining of Drosophila ovaries
Adult females were anesthetized by CO2 and dissected to get their ovaries in cold 1× PBS. Ovaries were fixed with 4% formaldehyde in 1× PBS at room temperature for 20 min. For DAPI staining, ovaries were washed with 1× PBS containing 0.3% Triton X-100 and mounted using ProLong Gold Antifade Mountant with DNA Stain DAPI (P36935, Invitrogen). For immunofluorescence staining, ovaries were washed with 1× PBS containing 0.3% Triton X-100 and permeabilized with 1% Triton X-100 in PBS at room temperature for 1 h. After washing with 1× PBS containing 0.3% Triton X-100, ovaries were blocked by 1% BSA in 1× PBS for 1 h and then incubated with cleaved Drosophila Dcp-1 (1:200 dilution; #9578, Cell Signaling) in 1× PBS at 4 °C overnight. Ovaries were washed with 1× PBS containing 0.3% Triton X-100, followed by incubation with fluorescent secondary antibody in 1× PBS at room temperature for 1 h. After extensive washing with 1× PBS containing 0.3% Triton X-100, ovaries were washed with 1× PBS and then mounted using ProLong Gold Antifade Mountant with DNA Stain DAPI (P36935, Invitrogen). Samples were observed using an inverted fluorescence microscope (Zeiss Axio Observer A1) equipped with a camera (Zeiss AxioCam MRm).
TUNEL assay of Drosophila ovaries
Drosophila ovaries were dissected in cold 1× PBS, fixed with 4% formaldehyde in 1× PBS for 20 min, and then washed with 1× PBS containing 0.3% Triton X-100. Ovaries were permeabilized with 1% Triton X-100 in PBS for 1 h at room temperature and washed with 1× PBS containing 0.3% Triton X-100. TUNEL assay was performed using One-step TUNEL In Situ Apoptosis Kit (Elabscience) according to the supplier’s recommendations. Briefly, ovaries were incubated in 100 μl of TdT Equilibration Buffer at 37 °C for 30 min. After adding 50 μl Labeling Working Solution, ovaries were incubated at 37 °C for 60 min with shading light in a humidified chamber. Ovaries were washed with 1× PBS containing 0.3% Triton X-100 three times, followed by 1× PBS three times, and then mounted using ProLong Gold Antifade Mountant with DNA Stain DAPI (P36935, Invitrogen). Samples were observed using an inverted fluorescence microscope (Zeiss Axio Observer A1) equipped with a camera (Zeiss AxioCam MRm).
Generation of oocyte-specific Ddx3x cKO mice
All C57BL/6 (C57BL/6JNarl) mice were purchased from the National Laboratory Animal Center in Taiwan. To generate Ddx3xloxP/loxP mice, two loxP sites were placed in both intron 1 and intron 10 of the Ddx3x gene using a CRISPR/Cas9 nuclease-based technology (Gene Knockout Mouse Core Lab, National Taiwan University). Zp3-Cre (C57BL/6-Tg(Zp3-cre)93Knw/J, Stock No. 003651) transgenic mice were purchased from the Jackson Laboratory and used to excise the floxed Ddx3x sequence in oocytes specifically. Ddx3xloxP/loxP females were mated with the Zp3-Cre transgenic males to generate Ddx3x cKO female mice with the genotype Ddx3xloxP/loxP; Zp3-Cre. Mice were housed in specific pathogen-free (SPF) conditions with free access to the standard rodent diet and water. The experimental procedures using mice were approved by the Institutional Animal Care and Use Committee at Chang Gung University (IACUC No. CGU110-208).
Mouse genotyping
Mouse ear samples were lysed in lysis buffer (50 mM KCl, 1.5 mM MgCl2, 50 mM Tris–HCl [pH 8.5], 0.45% IGEPAL CA-630, 0.45% Tween-20, 1% proteinase K) at 56 °C for 2 h, and then heated at 96 °C for 10 min to remove proteinase K activity. Genomic DNA was extracted from mouse ear lysates and genotyped by PCR. Ddx3xloxP/loxP mice were genotyped by PCR analysis using primers: 5VF1 (5′GAGCAGTGTGTAGCA TTTTTGT3′), 5VR1 (5′AGAGGTTTGAAGTCAGGGCAG3′), 3VF1 (5′AGAGAGGGAAGATCGGGTT3′), and 3VR1 (5′TAAACATCATAGTGTGGCGGA3′). Zp3-Cre transgenic mice were genotyped by PCR analysis using primers: Cre-F (5′GCATAACCAGTGAAACAGCATTGCTG3′) and Cre-R (5′GGACATGTT CAGGGATCGCCAGGCG3′). PCR reactions were carried out for 32 cycles (94 °C, 30 s; 55 °C, 30 s; 72 °C, 1 min).
Mouse fertility test
Ddx3xloxP/loxP and Ddx3xloxP/loxP; Zp3-Cre female mice were mated with C57BL/6 male mice (1:1), respectively. Male mice were removed 10 days after mating. The number of pups was counted. Male and female mice were randomly assigned, and the fertility test was repeated twice.
Immunohistochemistry staining
Mouse ovaries were fixed in 10% formalin overnight at room temperature, embedded in paraffin, and then cut into 5 μm serial sections. Ovarian sections were stained with hematoxylin and eosin (H&E). For immunohistochemistry staining, ovarian sections were de-paraffinized, rehydrated, and washed in 1× PBS three times. Antigen retrieval was performed by boiling the sections for 20 min in citrate buffer (BOND Epitope Retrieval Solution 1; Leica). For quenching endogenous peroxidase activity, ovarian sections were treated with 3% H2O2 in 1× PBS at room temperature for 10 min and rinsed briefly in 1× PBS. Ovarian sections were blocked by Mouse on Mouse (M.O.M.) Blocking Reagent (Vector Laboratories) at room temperature for 1 h. Ovarian sections were rinsed in 1× PBS and incubated with mouse anti-DDX3 antibody (0.4 μg/ml; sc-365768, Santa Cruz Biotechnology) or cleaved caspase 3 antibody (1:200 dilution; #9661, Cell Signaling) at room temperature for 1 h. After washing with 1× PBS, ovarian sections were incubated with a secondary antibody at room temperature for 1 h. Amplification of antibody responses to antigen was achieved by polymeric amplification system at room temperature for 1 h. The color reaction was precipitated using diaminobenzidine (DAB) at room temperature for 10 min. Ovarian sections were stained with H&E. After washing with 1× PBS, ovarian sections were dried and mounted using Dako Mounting Medium (Agilent Technologies). Samples were observed using the Olympus Microscope Digital Camera Model DP71.
TUNEL assay of mouse ovarian sections
Paraffin-embedded ovarian sections were de-paraffinized by xylene for 10 min twice, then hydrated with a sequential of hydrated ethanol of different percentages (100%, 95%, 90%, 80%, 75%) for 3 min each time. Ovarian sections were washed with 1× PBS for 5 min twice, then treated with 100 μg/ml proteinase K in 1× PBS at 37 °C for 20 min. Ovarian sections were washed with 1× PBS for 5 min three times. TUNEL assay was performed using the One-step TUNEL In Situ Apoptosis Kit (Elabscience) according to the supplier’s recommendations. Briefly, ovarian sections were incubated in 100 μl of TdT Equilibration Buffer at 37 °C for 30 min. After adding 50 μl Labeling Working Solution, ovarian sections were incubated at 37 °C for 60 min with shading light in a humidified chamber. Ovarian sections were washed with 1× PBS three times and then mounted using ProLong Gold Antifade Mountant with DNA Stain DAPI (P36935; Invitrogen). Samples were observed using an inverted fluorescence microscope (Zeiss Axio Observer A1) equipped with a camera (Zeiss AxioCam MRm).
Cell culture and transfection
HeLa and 293T cells were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). The cell lines used in this study were validated using short tandem repeat (STR) profiling and were free from mycoplasma contamination. Cells were grown in Dulbecco’s modified Eagles’ medium (DMEM) supplemented with 10% fetal bovine serum, 100 U/ml penicillin, 100 μg/ml streptomycin, and 2 mM L-glutamine at 37 °C in 5% CO2 incubator. Cell transfection was performed using the Lipofectamine® 2000 (Thermo Fisher Scientific) according to the manufacturer’s instructions.
Lentivirus-mediated RNAi knockdown
The RNAi Core Facility (Academia Sinica, Taiwan) provided all of the plasmids required for lentivirus production. The two pLKO.1-shRNA vectors used to knockdown DDX3 were as follows: TRCN0000000002 (shDDX3-1) and TRCN0000000004 (shDDX3-2). The transfection reagent Lipofectamine® 2000 (Thermo Fisher Scientific) was used for lentiviral production in 293T cells with a packaging plasmid (psPAX2), an envelope plasmid (pMD2.G), and a shRNA-expressing plasmid (pLKO.1). To knock down endogenous DDX3, cells were transduced with shRNA-expressing lentivirus at a multiplicity of infection (MOI) of 5 virus particles/cell in growth medium containing 8 µg/ml polybrene at 37 °C, 5% CO2. After 24 h incubation, puromycin (2 μg/ml) was added to the medium for selecting infected cells. Cells were harvested 3 days after transduction for analysis.
Quantitative real-time RT-PCR
Extracted total RNA was reverse transcribed into cDNA using the High-Capacity cDNA Reverse Transcription Kits (Thermo Fisher Scientific) according to the manufacturer’s instructions. According to the supplier’s recommendations, the resulting cDNA was subjected to quantitative real-time PCR analysis using StepOnePlus™ Real-Time PCR Systems (Thermo Fisher Scientific). Quantitative real-time PCR was performed using the Fast SYBR Green Master Mix (Thermo Fisher Scientific) and the forward and reverse primers (Table 3). The levels of mRNAs were detected by the measurement of threshold cycle (Ct) values during the exponential phase of amplification. Relative quantitation values were calculated using the 2−ΔΔCt method.
Sucrose gradient sedimentation and polysome profile analysis
293T cells were collected in cold PBS containing 100 µg/ml cycloheximide. All subsequent steps were performed at 4 °C. Cell pellets were resuspended in RSB-150 (10 mM Tris-HCl [pH 7.4], 3 mM MgCl2, and 150 mM NaCl) containing 100 µg/ml cycloheximide, 40 µg/ml digitonin (Calbiochem), 20 U/ml RNasin (Promega), and 1× protease inhibitor cocktail (Thermo Fisher Scientific). After incubation on ice for 5 min, cells were disrupted by passage through a 26-gauge needle five times. Cytoplasmic extracts were collected by centrifugation at 3000 × g for 2 min and clarified by further centrifugation at 11,000 × g for 15 min. The samples were loaded on a linear 15-40% sucrose gradient and centrifuged at 38,000 rpm for 3 h in a Beckman SW41 rotor. After centrifugation, total RNA was extracted from each fraction using phenol/chloroform extraction in the presence of 1% SDS and 0.25 M NaCl, followed by ethanol precipitation. The translational efficiency of a transcript was calculated as previously described [21].
Plasmid construction
For in vivo translation assay, the 5′ UTR fragments of APC1, CCNE1, P38γ, B56β, PP2Aβ, PKACα, FBXW11, RAC1, P38β, and GNAS mRNAs were obtained by RT-PCR using total RNA from 293T cells as the template and inserted into a unique HindIII site upstream of the firefly luciferase coding region in the pFL-SV40 vector [3]. All plasmid constructs were confirmed by DNA sequencing.
In vivo translation assay
293T cells (6 × 104 cells/well) were transduced with the empty lentiviral vector pLKO.1 or the pLKO.1 vector expressing shDDX3-1. At 48 h post transduction, cells were co-transfected with firefly luciferase reporter containing the 5′ UTR of DDX3 target mRNAs and the control pRL-SV40 vector encoding the Renilla luciferase. After 24 h post-transfection, cells were lysed in 1× Passive Lysis Buffer (Promega). The activities of firefly luciferase and Renilla luciferase were measured using the Dual-Luciferase Reporter Assay System (Promega).
Statistical analysis
For each experiment, at least three independent experiments were performed. Data from independent experiments were calculated and expressed as mean ± SD. Statistical analysis was performed using one-way ANOVA or unpaired Student’s t-test. A p value < 0.05 was considered statistically significant.
Data availability
Original data are available upon request.
References
Chuang RY, Weaver PL, Liu Z, Chang TH. Requirement of the DEAD-Box protein ded1p for messenger RNA translation. Science. 1997;275:1468–71.
Soto-Rifo R, Rubilar PS, Limousin T, de Breyne S, Decimo D, Ohlmann T. DEAD-box protein DDX3 associates with eIF4F to promote translation of selected mRNAs. EMBO J. 2012;31:3745–56.
Lai MC, Lee YH, Tarn WY. The DEAD-box RNA helicase DDX3 associates with export messenger ribonucleoproteins as well as tip-associated protein and participates in translational control. Mol Biol Cell. 2008;19:3847–58.
Lee CS, Dias AP, Jedrychowski M, Patel AH, Hsu JL, Reed R. Human DDX3 functions in translation and interacts with the translation initiation factor eIF3. Nucleic Acids Res. 2008;36:4708–18.
Calviello L, Venkataramanan S, Rogowski KJ, Wyler E, Wilkins K, Tejura M, et al. DDX3 depletion represses translation of mRNAs with complex 5′ UTRs. Nucleic Acids Res. 2021;49:5336–50.
Lai MC, Chang WC, Shieh SY, Tarn WY. DDX3 regulates cell growth through translational control of cyclin E1. Mol Cell Biol. 2010;30:5444–53.
Li Q, Zhang P, Zhang C, Wang Y, Wan R, Yang Y, et al. DDX3X regulates cell survival and cell cycle during mouse early embryonic development. J Biomed Res. 2014;28:282–91.
Heerma van Voss MR, Kammers K, Vesuna F, Brilliant J, Bergman Y, Tantravedi S, et al. Global effects of DDX3 inhibition on cell cycle regulation identified by a combined phosphoproteomics and single cell tracking approach. Transl Oncol. 2018;11:755–63.
Chen HH, Yu HI, Cho WC, Tarn WY. DDX3 modulates cell adhesion and motility and cancer cell metastasis via Rac1-mediated signaling pathway. Oncogene. 2015;34:2790–800.
He TY, Wu DW, Lin PL, Wang L, Huang CC, Chou MC, et al. DDX3 promotes tumor invasion in colorectal cancer via the CK1epsilon/Dvl2 axis. Sci Rep. 2016;6:21483.
Botlagunta M, Vesuna F, Mironchik Y, Raman A, Lisok A, Winnard P Jr, et al. Oncogenic role of DDX3 in breast cancer biogenesis. Oncogene. 2008;27:3912–22.
Yang F, Fang E, Mei H, Chen Y, Li H, Li D, et al. Cis-acting circ-CTNNB1 promotes beta-catenin signaling and cancer progression via DDX3-mediated transactivation of YY1. Cancer Res. 2019;79:557–71.
Sun M, Zhou T, Jonasch E, Jope RS. DDX3 regulates DNA damage-induced apoptosis and p53 stabilization. Biochim Biophys Acta. 2013;1833:1489–97.
Nguyen CN, Nguyen KVA, Eom S, Choi YJ, Kang LJ, Lee J, et al. Regulation of p21 expression for anti-apoptotic activity of DDX3 against sanguinarine-induced cell death on intrinsic pathway. Phytomedicine. 2019;65:153096.
Lai MC, Chen YP, Li DA, Yu JS, Hung HY, Tarn WY. DDX3 interacts with USP9X and participates in deubiquitination of the anti-apoptotic protein MCL1. FEBS J. 2022;289:1043–61.
Ariumi Y, Kuroki M, Abe K, Dansako H, Ikeda M, Wakita T, et al. DDX3 DEAD-box RNA helicase is required for hepatitis C virus RNA replication. J Virol. 2007;81:13922–6.
Yedavalli VS, Neuveut C, Chi YH, Kleiman L, Jeang KT. Requirement of DDX3 DEAD box RNA helicase for HIV-1 Rev-RRE export function. Cell. 2004;119:381–92.
Li C, Ge LL, Li PP, Wang Y, Dai JJ, Sun MX, et al. Cellular DDX3 regulates Japanese encephalitis virus replication by interacting with viral un-translated regions. Virology. 2014;449:70–81.
Schroder M, Baran M, Bowie AG. Viral targeting of DEAD box protein 3 reveals its role in TBK1/IKKepsilon-mediated IRF activation. EMBO J. 2008;27:2147–57.
Soulat D, Burckstummer T, Westermayer S, Goncalves A, Bauch A, Stefanovic A, et al. The DEAD-box helicase DDX3X is a critical component of the TANK-binding kinase 1-dependent innate immune response. EMBO J. 2008;27:2135–46.
Ku Y-C, Lai M-H, Lo C-C, Cheng Y-C, Qiu J-T, Tarn W-Y, et al. DDX3 participates in translational control of inflammation induced by infections and injuries. Mol Cell Biol. 2019;39:e00285–18.
Lennox AL, Hoye ML, Jiang R, Johnson-Kerner BL, Suit LA, Venkataramanan S, et al. Pathogenic DDX3X mutations impair RNA metabolism and neurogenesis during fetal cortical development. Neuron. 2020;106:404–20.e8.
Chen HH, Yu HI, Tarn WY. DDX3 modulates neurite development via translationally activating an RNA regulon involved in Rac1 activation. J Neurosci. 2016;36:9792–804.
Hoye ML, Calviello L, Poff AJ, Ejimogu NE, Newman CR, Montgomery MD, et al. Aberrant cortical development is driven by impaired cell cycle and translational control in a DDX3X syndrome model. Elife. 2022;11:e78203.
Venkataramanan S, Gadek M, Calviello L, Wilkins K, Floor SN. DDX3X and DDX3Y are redundant in protein synthesis. RNA. 2021;27:1577–88.
Ditton HJ, Zimmer J, Kamp C, Rajpert-De Meyts E, Vogt PH. The AZFa gene DBY (DDX3Y) is widely transcribed but the protein is limited to the male germ cells by translation control. Hum Mol Genet. 2004;13:2333–41.
Chen CY, Chan CH, Chen CM, Tsai YS, Tsai TY, Wu Lee YH, et al. Targeted inactivation of murine Ddx3x: essential roles of Ddx3x in placentation and embryogenesis. Hum Mol Genet. 2016;25:2905–22.
Gueler B, Sonne SB, Zimmer J, Hilscher B, Hilscher W, Graem N, et al. AZFa protein DDX3Y is differentially expressed in human male germ cells during development and in testicular tumours: new evidence for phenotypic plasticity of germ cells. Hum Reprod. 2012;27:1547–55.
Reijo R, Alagappan RK, Patrizio P, Page DC. Severe oligozoospermia resulting from deletions of azoospermia factor gene on Y chromosome. Lancet. 1996;347:1290–3.
Vogt PH, Edelmann A, Kirsch S, Henegariu O, Hirschmann P, Kiesewetter F, et al. Human Y chromosome azoospermia factors (AZF) mapped to different subregions in Yq11. Hum Mol Genet. 1996;5:933–43.
Kamp C, Huellen K, Fernandes S, Sousa M, Schlegel PN, Mielnik A, et al. High deletion frequency of the complete AZFa sequence in men with Sertoli-cell-only syndrome. Mol Hum Reprod. 2001;7:987–94.
Foresta C, Moro E, Rossi A, Rossato M, Garolla A, Ferlin A. Role of the AZFa candidate genes in male infertility. J Endocrinol Investig. 2000;23:646–51.
Shpargel KB, Sengoku T, Yokoyama S, Magnuson T. UTX and UTY demonstrate histone demethylase-independent function in mouse embryonic development. PLoS Genet. 2012;8:e1002964.
Luddi A, Margollicci M, Gambera L, Serafini F, Cioni M, De Leo V, et al. Spermatogenesis in a man with complete deletion of USP9Y. N Engl J Med. 2009;360:881–5.
Foresta C, Ferlin A, Moro E. Deletion and expression analysis of AZFa genes on the human Y chromosome revealed a major role for DBY in male infertility. Hum Mol Genet. 2000;9:1161–9.
Ramathal C, Angulo B, Sukhwani M, Cui J, Durruthy-Durruthy J, Fang F, et al. DDX3Y gene rescue of a Y chromosome AZFa deletion restores germ cell formation and transcriptional programs. Sci Rep. 2015;5:15041.
Vong QP, Li Y, Lau YF, Dym M, Rennert OM, Chan WY. Structural characterization and expression studies of Dby and its homologs in the mouse. J Androl. 2006;27:653–61.
Johnstone O, Deuring R, Bock R, Linder P, Fuller MT, Lasko P. Belle is a Drosophila DEAD-box protein required for viability and in the germ line. Dev Biol. 2005;277:92–101.
Liao SE, Kandasamy SK, Zhu L, Fukunaga R. DEAD-box RNA helicase Belle posttranscriptionally promotes gene expression in an ATPase activity-dependent manner. RNA. 2019;25:825–39.
Wang J, Li T, Deng S, Ma E, Zhang J, Xing S. The RNA helicase DDX3 is required for ovarian development and oocyte maturation in Locusta migratoria. Arch Insect Biochem Physiol. 2021;106:e21775.
Promislow DE, Smith EA, Pearse L. Adult fitness consequences of sexual selection in Drosophila melanogaster. Proc Natl Acad Sci USA. 1998;95:10687–92.
Song Z, McCall K, Steller H. DCP-1, a Drosophila cell death protease essential for development. Science. 1997;275:536–40.
Sarkissian T, Timmons A, Arya R, Abdelwahid E, White K. Detecting apoptosis in Drosophila tissues and cells. Methods. 2014;68:89–96.
Pluck A. Conditional mutagenesis in mice: the Cre/loxP recombination system. Int J Exp Pathol. 1996;77:269–78.
Tsien JZ, Chen DF, Gerber D, Tom C, Mercer EH, Anderson DJ, et al. Subregion- and cell type-restricted gene knockout in mouse brain. Cell. 1996;87:1317–26.
Lan ZJ, Xu X, Cooney AJ. Differential oocyte-specific expression of Cre recombinase activity in GDF-9-iCre, Zp3cre, and Msx2Cre transgenic mice. Biol Reprod. 2004;71:1469–74.
Lewandoski M, Wassarman KM, Martin GR. Zp3-cre, a transgenic mouse line for the activation or inactivation of loxP-flanked target genes specifically in the female germ line. Curr Biol. 1997;7:148–51.
Ghafari F, Gutierrez CG, Hartshorne GM. Apoptosis in mouse fetal and neonatal oocytes during meiotic prophase one. BMC Dev Biol. 2007;7:87.
Thomas JD, Johannes GJ. Identification of mRNAs that continue to associate with polysomes during hypoxia. RNA. 2007;13:1116–31.
Dicke AK, Pilatz A, Wyrwoll MJ, Punab M, Ruckert C, Nagirnaja L, et al. DDX3Y is likely the key spermatogenic factor in the AZFa region that contributes to human non-obstructive azoospermia. Commun Biol. 2023;6:350.
Matsumura T, Endo T, Isotani A, Ogawa M, Ikawa M. An azoospermic factor gene, Ddx3y and its paralog, Ddx3x are dispensable in germ cells for male fertility. J Reprod Dev. 2019;65:121–8.
Kim YS, Lee SG, Park SH, Song K. Gene structure of the human DDX3 and chromosome mapping of its related sequences. Mol Cells. 2001;12:209–14.
Inoue H, Ogonuki N, Hirose M, Hatanaka Y, Matoba S, Chuma S, et al. Mouse D1Pas1, a DEAD-box RNA helicase, is required for the completion of first meiotic prophase in male germ cells. Biochem Biophys Res Commun. 2016;478:592–8.
Oh S, Flynn RA, Floor SN, Purzner J, Martin L, Do BT, et al. Medulloblastoma-associated DDX3 variant selectively alters the translational response to stress. Oncotarget. 2016;7:28169–82.
Chen HH, Yu HI, Yang MH, Tarn WY. DDX3 activates CBC-eIF3-mediated translation of uORF-containing oncogenic mRNAs to promote metastasis in HNSCC. Cancer Res. 2018;78:4512–23.
Chen HH, Yu HI, Chang JJ, Li CW, Yang MH, Hung MC, et al. DDX3 regulates cancer immune surveillance via 3′ UTR-mediated cell-surface expression of PD-L1. Cell Rep. 2024;43:113937.
Chen HH, Yu HI, Rudy R, Lim SL, Chen YF, Wu SH, et al. DDX3 modulates the tumor microenvironment via its role in endoplasmic reticulum-associated translation. iScience. 2021;24:103086.
Sun M, Song L, Li Y, Zhou T, Jope RS. Identification of an antiapoptotic protein complex at death receptors. Cell Death Differ. 2008;15:1887–900.
Pesin JA, Orr-Weaver TL. Regulation of APC/C activators in mitosis and meiosis. Annu Rev Cell Dev Biol. 2008;24:475–99.
Martinerie L, Manterola M, Chung SS, Panigrahi SK, Weisbach M, Vasileva A, et al. Mammalian E-type cyclins control chromosome pairing, telomere stability and CDK2 localization in male meiosis. PLoS Genet. 2014;10:e1004165.
Perdiguero E, Pillaire MJ, Bodart JF, Hennersdorf F, Frodin M, Duesbery NS, et al. Xp38gamma/SAPK3 promotes meiotic G(2)/M transition in Xenopus oocytes and activates Cdc25C. EMBO J. 2003;22:5746–56.
Bel Borja L, Soubigou F, Taylor SJP, Fraguas Bringas C, Budrewicz J, Lara-Gonzalez P, et al. BUB-1 targets PP2A:B56 to regulate chromosome congression during meiosis I in C. elegans oocytes. Elife. 2020;9:e65307.
Wlodarchak N, Xing Y. PP2A as a master regulator of the cell cycle. Crit Rev Biochem Mol Biol. 2016;51:162–84.
Pirino G, Wescott MP, Donovan PJ. Protein kinase A regulates resumption of meiosis by phosphorylation of Cdc25B in mammalian oocytes. Cell Cycle. 2009;8:665–70.
Xie J, Jin Y, Wang G. The role of SCF ubiquitin-ligase complex at the beginning of life. Reprod Biol Endocrinol. 2019;17:101.
Halet G, Carroll J. Rac activity is polarized and regulates meiotic spindle stability and anchoring in mammalian oocytes. Dev Cell. 2007;12:309–17.
Suzanne M, Irie K, Glise B, Agnes F, Mori E, Matsumoto K, et al. The Drosophila p38 MAPK pathway is required during oogenesis for egg asymmetric development. Genes Dev. 1999;13:1464–74.
Mehlmann LM, Jones TL, Jaffe LA. Meiotic arrest in the mouse follicle maintained by a Gs protein in the oocyte. Science. 2002;297:1343–5.
Acknowledgements
We thank Dr. Li-Mei Pai (Graduate Institute of Biomedical Sciences, Chang Gung University, Taoyuan, Taiwan) for technical assistance in DAPI staining of Drosophila ovaries, and Dr. Jiuan-Jiuan Hwang (Department and Institute of Physiology, National Yang Ming Chiao Tung University, Taipei, Taiwan) for observing mouse ovarian sections. We thank the National RNAi Core Facility (Academia Sinica, Taipei, Taiwan) for providing shRNA reagents and related services. We also thank the technical services of the Transgenic Mouse Models Core Facility of the National Core Facility for Biopharmaceuticals (National Science and Technology Council, Taiwan) and the Gene Knockout Mouse Core Laboratory of National Taiwan University Center of Genomic and Precision Medicine. Immunohistochemical stains were performed by the Pathology Core Laboratory (Molecular Medicine Research Center, Chang Gung University, Taoyuan, Taiwan).
Funding
This work was supported by grants from the National Science and Technology Council, Taiwan (112-2320-B-182-014), and Chang Gung Memorial Hospital, Taiwan (CMRPD1M0731).
Author information
Authors and Affiliations
Contributions
M-CL designed research; S-YT, C-HL, and Y-TJ performed research; M-CL, S-YT, C-HL, Y-TJ, G-JH, and HP analyzed data; M-CL wrote the paper; G-JH, HP, H-YH, and W-YT revised the paper.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval
All animal studies were carried out in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the NIH. The experimental procedures using mice were reviewed and approved by the Institutional Animal Care and Use Committee at Chang Gung University, Taiwan (IACUC:CGU110-208).
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tsai, SY., Lin, CH., Jiang, YT. et al. DDX3 is critical for female fertility via translational control in oogenesis. Cell Death Discov. 10, 472 (2024). https://doi.org/10.1038/s41420-024-02242-6
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41420-024-02242-6
