
Abstract
Pyroglutamylated RF-amide peptide (QRFP) is a peptide hormone with a C-terminal RF-amide motif. QRFP selectively activates a class A G-protein-coupled receptor (GPCR) GPR103 to exert various physiological functions such as energy metabolism and appetite regulation. Here, we report the cryo-electron microscopy structure of the QRFP26-GPR103-Gq complex at 3.19 Å resolution. QRFP26 adopts an extended structure bearing no secondary structure, with its N-terminal and C-terminal sides recognized by extracellular and transmembrane domains of GPR103 respectively. This movement, reminiscent of class B1 GPCRs except for orientation and structure of the ligand, is critical for the high-affinity binding and receptor specificity of QRFP26. Mutagenesis experiments validate the functional importance of the binding mode of QRFP26 by GPR103. Structural comparisons with closely related receptors, including RY-amide peptide-recognizing GPCRs, revealed conserved and diversified peptide recognition mechanisms, providing profound insights into the biological significance of RF-amide peptides. Collectively, this study not only advances our understanding of GPCR-ligand interactions, but also paves the way for the development of novel therapeutics targeting metabolic and appetite disorders and emergency medical care.
Similar content being viewed by others

Introduction
Neuropeptides, a diverse array of signaling molecules, orchestrate a multitude of physiological processes in organisms. Among them, neuropeptides possessing the Arg-Phe-NH2 (RF-amide) motif at their C-termini are designated as RF-amide peptides and have attracted considerable attention because of their pivotal roles in various biological functions1,2,3. The RF-amide family encompasses five distinct peptides: neuropeptide FF (NPFF), prolactin-releasing peptide (PrRP), kisspeptin (Kiss1), gonadotropin-inhibitory hormone (GnIH), and pyroglutamylated RF-amide peptide (QRFP). Each peptide interacts with specific class A G-protein-coupled receptors (GPCRs), initiating a cascade of intracellular events to modulate physiological responses. Similarly, the RY-amide peptides, characterized by the signature RY motif at their C-termini, present a similar mode of action, interacting with their respective receptors to regulate physiological processes4. This categorization based on the C-terminal residues highlights the specificity and diversity within these peptide families. The intricate interplay between these peptides and their receptors represents a complex network critical for maintaining homeostasis and responding to environmental changes.
QRFP is a 43-amino acid RF-amide peptide with a pyroglutamylated N-terminus (namely QRFP43)5 and demonstrates specific activity for GPR103 with orexigenic activity5,6,7. GPR103 is a Gi/o- and Gq-coupled GPCR expressed in the adrenal gland and various regions of the brain such as the hypothalamus, which plays a crucial role in the regulation of energy metabolism and appetite control. QRFP and GPR103 are implicated in a variety of physiological functions ranging from the modulation of feeding behavior to the regulation of energy homeostasis8, cardiovascular function, and bone formation9. This QRFP-GPR103 pair is remarkably conserved across various vertebrate species10, highlighting its fundamental significance in biological systems. GPR103 is not only pivotal in maintaining physiological balance but also presents therapeutic targets for disorders related to metabolism and appetite dysregulation. For example, QRFP administration reportedly increased food intake and fat mass while reducing glucose-induced insulin release, and may also cause osteopenia and facilitate nociception11. Thus, GPR103 antagonists are expected to be useful in the prevention and treatment of various metabolic disorders such as bulimia, vasospasm, obesity, diabetes, endocrine disorders, hypercholesterolemia, hyperlipidemia, gout, and fatty liver. They may also be useful as therapeutic agents for cardiovascular and renal diseases, including atherosclerosis and heart failure. Several nonpeptidic GPR103 antagonists (e.g. pyrrolo[2,3-c]pyridine) have been discovered from peptidomimetics and high-throughput screening, and have demonstrated anorexic activity in mice.
Recent advancements in cryo-electron microscopy (cryo-EM) have unveiled numerous GPCR–G-protein complex structures, including those associated with C-terminally amidated peptides such as cholecystokinin, orexin, and RY-amide neuropeptide Y12,13,14. These analyses have shed light on the intricacies of ligand-receptor interactions and their activation mechanisms. Nevertheless, a notable gap remains in our understanding of the mechanisms distinguishing RF- and RY-amides. This also includes an understanding of the selective activation of GPR103 by QRFP and the fundamental principles dictating ligand specificity among related receptors. These gaps in structural data significantly hinder the strategic development of therapeutic agents targeting GPR103. Here, we show a cryo-EM structure of the QRFP-bound GPR103•Gq complex, offering insights into its ligand recognition and selectivity, and dynamics.
Results
Overall structure
For the structural study, we used an N-terminally truncated form of QRFP, known as QRFP26 (Fig. 1a). QRFP26 is also found in vivo and possesses agonist activity comparable to that of QRFP437,8,15. In this study, we focused on the structural analysis complexed with Gq. To facilitate expression and purification, we truncated the C-terminal residues after G366 of human GPR103. The truncated mutant activated Gq to the same extent as the wild-type in the TGFα shedding assay16 (Supplementary Fig. 1a–d and Supplementary Table 1). We used a mini-Gαq (mini-Gsqi), an engineered mini-Gαs protein whose N-terminal and C-terminal residues are replaced by Gαi1 and Gαq, respectively17. To efficiently purify the stable GPCR–G-protein complex, the receptor, and mini-Gsqi were incorporated in a ‘Fusion-G system’ by combining two complex stabilization techniques18,19 (Supplementary Fig. 2a, b). The modified receptor and G-protein were co-expressed in HEK293 cells and purified by FLAG affinity chromatography. After an incubation with Nb35 and scFv16 that bind to mini-Gsqi, the complex was purified by size-exclusion chromatography (Supplementary Fig. 2c, d). The structure of the purified complex was determined by single-particle cryo-EM analysis with an overall resolution of 3.19 Å (Supplementary Fig. 3, Supplementary Table 2, and “Methods”). As the extracellular portion of the receptor was poorly resolved, we performed receptor-focused refinement, yielding a density map with a nominal resolution of 3.29 Å, which was combined with the overall refined map. The resulting composite map allowed us to precisely build the atomic model of all the components, including the receptor (residues 3–243 and 263–346), QRFP26 (residues 3–26), G-proteins, and antibodies (Fig. 1b, c).

a Amino acid sequences of QRFP43 and QRFP26. b Cryo-EM density map of the GPR103–mini-Gsqiβ1γ2–scFv16–Nb35 individually colored. c Refined structures of the complex are shown as a ribbon representation. d Diagram of GPR103. N-terminal forms a helix-loop-helix motif. ECL2 forms a long β-sheet. e Ribbon representation of the QRFP26 and GPR103. Density focused on QRFP26 (pink). Two disulfide bonds are represented by stick models. The one is the highly conserved disulfide bond between C1183.25 and C201ECL2, and the other is atypical disulfide bond between C2856.47 and C3277.48. The C1183.25A and C201ECL2A mutations abolished the QRFP26 potency (Supplementary Fig. 1a–d). By contrast, the C2856.47A and C3277.48A mutations did not alter the potency, indicating their lesser importance for receptor function.
The receptor consists of the canonical 7 transmembrane helices (TM) connected by three intracellular (ICL1–3) and three extracellular (ECL1–3) loops, the amphipathic helix 8 at the C-terminus (H8), and the N-terminal residues exposed on the extracellular side (Fig. 1d, e). ICL3 was disordered in the cryo-EM map. At the secondary-structure level, ICL2 and ECL3 contain short α-helices, and ECL2 forms a long β-sheet. The N-terminal residues extend above ECL2, constituting the extracellular domain (ECD) together with ECL2. An unambiguous density was observed from the interior of the transmembrane domain (TMD) to the ECD, allowing us to model residues 3–26 of QRFP26 (Fig. 1e). The residues 5–13 adopt an α-helix, consistent with the NMR analysis of QRFP alone20. The G-protein docks in the intracellular cavity of the receptor, forming similar interactions as in other G-protein complexes13,17,19,21,22,23,24,25 involving conserved D3.49R3.50Y3.51 (superscripts indicate Ballesteros–Weinstein numbers26) (modified to E3.49R3.50H3.51 in GPR103) and N7.49P7.50xxY7.53 motifs (Supplementary Fig. 4a–j).
QRFP26 binding site in the transmembrane domain
QRFP26 binds to both the ECD and TMD with its C-terminal amide directed toward the TMD core (Fig. 1e). The C-terminal heptapeptide of QRFP26 (GGFSFRF) fits vertically within the TMD and creates an extensive interaction network with TMs 2–7 and ECL2 (Fig. 2a, b and Supplementary Fig. 5). This interaction is broadly divided into the residues 20–24 (GGFSF) and the RF-amide moiety (R25 and F26). In the former part, the peptide backbone of QRFP26 forms hydrogen bonds with Q1052.64, Q1844.64, and Q3187.39. The Q1844.64A and Q3187.39A mutations reduced the potency of QRFP26 in the TGFα shedding assay, while the Q1052.64A mutation retained the potency (Fig. 2c, Supplementary Fig. 1a–c and Supplementary Table 1). Furthermore, F22 and F24 form stacking interactions with W111ECL1 (Fig. 2b), and the W111ECL1A mutation abolished the response for QRFP26 (Fig. 2c, Supplementary Fig. 1a–c). W111ECL1 stabilizes the upright structure of QRFP26 and plays an essential role in QRFP26 binding.

a, b Binding pocket for QRFP26 in the TMD. Residues involved in the QRFP-GPR103 interaction within 4.5 Å are shown as pink and blue sticks, respectively. Black dashed lines indicate hydrogen bonds. c Effects of mutations in the ligand-binding pocket of GPR103. QRFP26-induced activation of GPR103 was analyzed by the TGFα shedding assay. From the concentration–response curves (Supplementary Fig. 1b), ΔpEC50 values relative to the wild-type were calculated. Colors in the mutant bars indicate an expression level matching that of titrated wild-type. NA, parameter not available because of lack of the ligand response. WT, wild-type. Statistical analyses were performed using the ordinary one-way ANOVA followed by Dunnett tests with the expression-matched (colored) wild-type response. ns, p > 0.05; *p < 0.05; **p < 0.01; ***p < 0.001. Data are presented as mean values ± SEM from at least three independent experiments performed in triplicate (n = 5 for the wild-type and n = 3 for the mutants). Source data are provided as a Source Data file. d, e Close-up views of F26 (d) and R25 (e) of QRFP26. f Surface representation of GPR103 viewed from the extracellular side. Positive and negative charges of the receptor are colored in blue and red, respectively.
In comparison, RF-amide binds deep in the pocket (Fig. 2a), consistent with its significance in receptor binding reported in previous studies with mutant peptides27,28. The F26 side chain is present at the deepest position in the TMD, surrounded by bulky hydrophobic residues in TM2, TM3, and TM6 (Fig. 2d). The C-terminal amide forms hydrogen-bonding interactions with T1022.61, Q1253.32, and Q3187.39, and these mutations reduced the QRFP26 potency by over 3-fold (Fig. 2c). R25 forms diverse interactions with receptors, including hydrogen bonds with T2155.39, electrostatic interactions with E203ECL2 and E2976.59, and stacking interactions with W205ECL2 and Y2145.38 (Fig. 2e). While the T2155.39A mutation minimally affected the potency, the E203ECL2A and E2976.59A mutations reduced the potency by 3- and 10-fold, respectively, and the E203ECL2A/E2976.59A double mutation abolished the potency (Fig. 2c, Supplementary Fig. 1a–c, Supplementary Table 1). These results indicate that electrostatic interaction with E203ECL2A and E2976.59A plays an important role in QRFP26 binding. E203ECL2 and E2976.59 make the ligand-binding pocket of the TMD negatively charged (Fig. 2f). Thus, the C-terminal amidation functions not only in interactions with the receptor but also in enhancing the charge complementarity with the pocket, by neutralizing the negative charge of the C-terminal carboxylate. Moreover, the W205ECL2A and Y2145.38A mutations dramatically reduced the potency (Fig. 2c, Supplementary Fig. 1b, c, Supplementary Table 1). These aromatic residues would also be involved in pocket formation through the stacking interactions between them (Fig. 2e), suggesting their importance. Overall, our structural and mutational studies demonstrate the structure-activity relationship of QRFP26 binding in the TMD.
Previous studies with mutant peptides have shown that the C-terminal heptapeptide is sufficient to activate GPR10327,28, despite its reduced affinity. The heptapeptide is conserved from fish to mammals, except for S23 (Supplementary Fig. 6a), consistent with the fact that the S23 side chain poorly interacts with the receptor. The receptor residues interacting with the heptapeptide are highly conserved among the homologs (Supplementary Fig. 6b, c). Accordingly, this observed heptapeptide-TMD interaction plays a key role in evolutionarily conserved GPR103 activation.
Architecture of the N-terminal region
We observed an unambiguous density above ECL2, despite the suboptimal local resolution ranging from 4 to 6 Å (Supplementary Fig. 3). The density aligned with the N-terminal structure predicted by AlphaFold29,30,31 and allowed us to discuss the potential interactions between the ECD and QRFP26. The N-terminal residues 2–40 of GPR103 extend from TM1 to above ECL2, partially covering the ligand-binding pocket in the TMD. The residues 2–18 form a helix-loop-helix (HLH) above ECL2 (Fig. 3a). QRFP26 extends vertically from G20, interacting predominantly with the N-terminal region. The peptide backbone of R17 and K18 form chain-to-chain hydrogen-bonding interactions with L34N-term and V35N-term (Fig. 3b), while the α-helix is recognized by HLH (Fig. 3c). Specifically, the HLH structure is stabilized by stacking between centrally oriented F11N-term, F25N-term, and Y29N-term. These aromatic residues, together with surrounding hydrophobic residues, sandwich the α-helix of QRFP and form an extensive hydrophobic interaction. Moreover, the position of HLH is stabilized by hydrophobic interactions with ECL2 among F11N-term, I26N-term, L193ECL2, and Y194ECL2 (Fig. 3c). The alanine mutations of these aromatic residues reduced the QRFP26 potency by about 10-fold (Fig. 3d, Supplementary Fig. 1a–c, Supplementary Table 1). Overall, our structure and mutational study indicate that N-terminal HLH plays a critical role in the binding of QRPF26.

a Ribbon representation of GPR103 focused on the extracellular side. b Hydrogen-bonding interactions of R17-P33N-term and K18-Y36N-term are indicated by black dashed lines. c Hydrophobic interaction of the N-terminal HLH with ECL2 and QRFP26. d–f Effects of mutations in the ECD of GPR103. QRFP26-induced activation of GPR103 was analyzed by the TGFα shedding assay. From the concentration–response curves (Supplementary Fig. 1a), ΔpEC50 values relative to the wild-type were calculated (d). The concentration–response curves of the N-terminus-truncated mutants and the ECL2 deletion mutant were shown in e and f, respectively. Data for the wild-type response was obtained from the same experiment as Fig. 2c. Colors in the mutant bars indicate an expression level matching that of titrated wild type. NA, parameter not available because of lack of the ligand response. WT, wild-type. Statistical analyses were performed using the ordinary one-way ANOVA followed by Dunnett tests with the expression-matched (colored) wild-type response. ns, p > 0.05; *p < 0.05; **p < 0.01; ***p < 0.001. Data are presented as mean values ± SEM from at least three independent experiments performed in triplicate (n = 5 for the wild-type and n = 3 for the mutants). Source data are provided as a Source Data file.
To further examine the functional role of the ECD, we made truncated mutants of the N-terminal region and ECL2 (Fig. 3a). Δ1–18 or Δ1–30 is the mutant in which the N-terminal HLH is partially and completely truncated, respectively. Δ1–43 or ΔECL2 is the mutant in which the N-terminal region or ECL2 is completely truncated, respectively. Although the expression levels of these mutants were greatly reduced (Supplementary Fig. 1b), we could analyze the effects of the mutations by comparing them with the wild-type, whose expression levels were reduced to the same extent. Δ1–43 and ΔECL2 completely lost the QRFP26 potency in the TGFα shedding assay (Fig. 3e, f). Such truncation would completely break the ECD architecture and further affect the shape of the ligand-binding pocket in the TMD, suggesting that the ECD plays an indispensable role in QRFP binding. By contrast, we measured the QRFP26-dependent activity of Δ1–18 and Δ1–30 at concentrations of 10−6 to 10−5 M (Fig. 3e), although their potencies were greatly reduced to the point that their EC50 values could not be calculated. These data suggest that the HLH fractions as an affinity trap, achieving high compatibility through interactions with the N-terminal residues of QRFP26. Notably, Δ1–18 was more active than Δ1–30 (Fig. 3e), suggesting the remaining function as the affinity trap in Δ1–18 and further validating our cryo-EM structure. This N-terminal HLH motif is evolutionarily conserved among GPR103 homologs from zebrafish to humans (Supplementary Fig. 6c), and our homology searches have failed to identify any similar sequences in other proteins, indicating an unusual feature of GPR103.
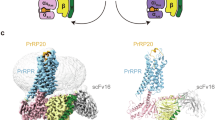
Next, we performed a 3D flexible refinement implemented in cryoSPARCv4.4.032 (Supplementary Fig. 7a–f) and uncovered a significant conformational change of the ECD (Supplementary Fig. 7g, h and Supplementary Movie 1). We then built the models of this alteration on the two most significantly changed maps among the output (Fig. 4a. b and Supplementary Table 2). The results revealed the upright and tilted states of the ECD. A structural comparison of the two states elucidated the dynamic movement of the N-terminal HLH by 4.6 Å (Fig. 4b). Moreover, QRFP26, the ECD, and the extracellular half of the TMD oscillate like a pendulum with the C-terminus of QRFP26 as the base point (Supplementary Movie 1). In the original map, the ECD of the refined structure is positioned in between the upright and tilted states, whereas the transmembrane helices are more closely aligned with the upright, implying the fundamental stability of the upright state.

Similar fluctuating action is observed in the parathyroid hormone receptor 1 (PTH1R), a class B1 GPCRs that feature an ECD with a ~150 amino acid ‘hotdog-like’ domain. The ECD acts as an affinity trap by encasing the agonist peptide, and cryo-EM structural analysis elucidated the structural polymorphism in its ECD33,34,35 (Fig. 4c, d). Despite significant differences in sequence homology and length, a functional analogy exists between the ECDs of GPR103 and PTH1R. However, the extracellular half of the GPR103-TMD undergoes a structural change following the ECD (Fig. 4b), while the PTH1R-TMD of PTH1R does not (Fig. 4d). Consequently, the structure and dynamics of the ECD and QRFP26 observed in this study are characteristic to GPR103, thus playing a critical role in the GPR103 selectivity of QRFP26.
Conservation among the RF-amide receptors
The RF-amide moiety is the only conserved part of the RF-amide peptide, while the length and sequence of their N-terminal portions vary widely (Supplementary Fig. 8a). Thus, ligand interactions in the extracellular region are likely to be different for individual RF-amide receptors. Indeed, amino acid sequence alignment indicates that only GPR103 has an N-terminal HLH (Supplementary Fig. 8b). To examine the conservation of the RF-amide recognition, we compared the residues interacting with the RF-amide with the corresponding RF-amide receptors; PrRPR (GPR10), KISS1R (GPR54), NPFFR1 (GPR147), and NPFFR2 (GPR74) (Fig. 5a). The recognition of the F26 side chain is less stringent but is shared by a bulky hydrophobic amino acid. By contrast, T1022.61 and Q1253.32, which recognize the C-terminal amide of QRFP26, are completely conserved. Although Q3187.39 is replaced by histidine in all the other RF-amide receptors, it would form hydrogen bonds with the oxygen atom of the C-terminal amide of QRFP26. These considerations suggest that amide recognition by hydrogen-bonding interactions via these three residues is a common mechanism in RF-amide receptors. The two negative charges near R25 are also conserved in RF-amide receptors, although E2976.59 is replaced by alanine in KISS1R. Instead, Q2115.35 is replaced by E2015.35 in KISS1R, suggesting the conserved recognition of R25 by the two negative charges. The triple mutants of E203ECL2A, E2976.59A, and Q2115.35E increased the QRFP26 potency compared with the E203ECL2A/E2976.59A double mutant (Fig. 2c, Supplementary Fig. 1a–d, and Supplementary Table 1). Thus, the RF-amide recognition mechanism observed in QRFP26–GPR103 is highly conserved in RF-amide receptors.

a Comparison of residues interacting with the RX-amide moiety in the RF- and RY-amide receptors. b Superimposition of GPR103 (blue), Y1R (PDB 7X9A (NPY–Y1R–Gi structure), green), CCK1R (PDB 7MBY (CCK-8–CCK1R–mini-Gsqi structure), magenta), and OX2R (PDB 7L1U (OxB–OX2R–mini-Gsqi structure), purple) structures in complex with QRFP26, NPY, CCK-8, and OxB, peptides, respectively. NPY, neuropeptide Y; CCK-8, cholecystokinin octapeptide; OxB, orexin-B. c, d Structural comparison of the binding mode of the RF-amide moiety in GPR103 (c) and the RY-amide moiety in Y1R (d). The residues involved in ligand-receptor interactions are represented by stick models. Black dashed lines indicate hydrogen bonds. e Schematic representations of RX-amide recognition conserved in RF- or RY-amide receptors. The most conserved residues are indicated based on the structures of GPR103 and Y1R.
Structural comparison with related peptide receptors
GPR103 exhibits a sequence homology with cholecystokinin receptors (CCKRs), orexin receptors (OXRs), and RY-amide neuropeptide Y receptors (YRs), which commonly recognize peptides with amidated C-termini (Supplementary Fig. 8a). To elucidate the characteristics of the structure and peptide recognition mechanisms of GPR103, we compared the endogenous ligand-bound structures of Y1R, CCK1R, and OX2R12,13,14 (Fig. 5b). These receptors have a long ECL2 in common, with diversity in length and angle depending on the type of ligand. While the short peptide ligands cholecystokinin octapeptide (CCK-8) and orexin-B (OxB) are buried in the TMD, QRPF26, and neuropeptide Y (NPY) have an α-helix at the N-terminus. The α-helix of NPY is recognized by ECL2 that of QRFP26 is by N-terminal HLH, further highlighting the distinctiveness of the N-terminal structure for peptide recognition. Despite the differences on the extracellular side, the C-termini of the agonist peptides as well as the intracellular sides of the receptors, superimposed well, suggesting an evolutionary linkage among these receptors.
We compared the interactions of the C-terminal residues of the ligand in GPR103 and Y1R (Fig. 5c, d), representing the RF- and RY- amide receptors, respectively. In both instances, the C-terminal arginine of the ligand forms electrostatic interactions with two negatively charged residues. The carbonyl oxygen of the C-terminal amide hydrogen bonds with Q3.32 and Q/H7.39. These four residues are highly conserved in the RF-amide and RY-amide receptors, indicating a shared recognition mechanism for the C-termini of the RX-amide peptide ligands (Fig. 5e). It should be noted that C2.57 is completely conserved and forms a hydrogen bond with the C-terminal nitrogen of NPY in Y1R, while the hydrogen bond is doubtful in GPR103 due to bond angle issue. However, the C982.57A mutation reduced the QRFP potency by 3-fold, suggesting the potential involvement in QRFP26 binding (Fig. 5e).
Next, we focused on the difference between the recognition of the RY-amide and RF-amide. In Y1R, Q2195.46 forms a hydrogen bond with the hydroxyl group of the C-terminal Y36 of NPY14,36 (Fig. 5c), which is highly conserved in the NPY receptors (Fig. 5a). Exceptionally, Q2195.46 is replaced by L2275.46 in Y2R (Fig. 5a), but its cryo-EM structure revealed that S2235.42, situated one turn above, alternatively forms a hydrogen bond with Y3614,37. Contrarily, in the QRFP26–GPR103 complex, the hydrogen bond between Y36 and Q2195.46 is replaced by a hydrophobic interaction between F26 and L2225.46. L5.46 is highly conserved in the RF-amide receptors (Fig. 5a). Although L5.46 is replaced by the polar residue T5.46 in PrRPR, it is not expected to interact with the C-terminal phenylalanine, due to its shorter side chain. The presence or absence of a residue capable of hydrogen bonding with the hydroxyl group of tyrosine at the C-terminus may distinguish the RF- and RY-amide receptors (Fig. 5e).
Discussion
Our study has revealed the binding mode of the C-terminal QRFP heptapeptide to the TMD, which is sufficient for the activation of GPR103. The conserved recognition mechanism of the C-terminal RF-amide in various RF-amide receptors suggests broader biological significance. Comparisons with other evolutionarily close peptide receptors demonstrated the commonality and diversity in the recognition mechanisms of the C-terminal two residues of peptide ligands. In particular, the presence or absence of residues that can form hydrogen bonds with the C-terminal side chain determines whether they function as RF- or RY-amide receptors. We identified the N-terminal HLH structure of GPR103, which captures the N-terminal side of QRFP and is quite unusual compared to class A GPCRs. We observed a pendulum-like motion of the ECD, including the N-terminus and the entire pocket, reminiscent of class B1 GPCRs. It should be noted that the orientation and secondary structure of the peptide ligands totally differ since the C-terminus of the α-helical ligand binds the extracellular domain in class B1 GPCRs. The structure and dynamics of these extracellular regions are important for promoting the specific, high-affinity binding of QRFP to GPR103. The distinctive structure and function of GPR103 determined in this study will be useful in the design of potential therapeutics targeting GPR103 for energy metabolism and appetite regulation.
Although the current active conformation was modeled based on the AlphaFold2 predicted structure31, there are considerable differences between them. Notably, the intracellular side of TM6 is in a closed configuration (Supplementary Fig. 9a, b), and R1433.50 of the D3.49R3.50Y3.51 motif (modified to E3.49R3.50H3.51 in GPR103) forms a salt bridge with E1423.49 and is not oriented toward the intracellular face (Supplementary Fig. 9c), indicating that the predicted conformation represents the inactive state. This observation allowed us to discuss the mechanism of receptor activation upon QRFP26 binding, by comparing the cryo-EM structure with the predicted structure. Within the ligand-binding pocket in the TMD, conformational changes were observed in TM5 and TM6 (Supplementary Fig. 9d, e). The hydrogen-bonding interaction between R25 and T2155.39 displaces TM5 inward by 1.4 Å. Conversely, F26 in QRFP induces the steric outward movement of F2896.51, resulting in the outward displacement of TM6 by 1.6 Å. This movement of F26 drives W2866.48 downward, thereby exerting a downward force on F2826.44 in the underlying P5.50I3.40F6.44 motif. This action is an important component of the P5.50I3.40F6.44 motif reorganization frequently observed in class A GPCRs21 and leads to the opening of the intracellular machinery of TM6. The mutations of F2826.44A, W2866.48A, and F2896.51A almost abolished the potency of QRFP26 (Supplementary Fig. 1b, c, 9f, and Supplementary Table 1), supporting our proposed importance for receptor activation. Such receptor activation, characterized by conformational changes in W2866.48, is a widespread phenomenon in class A GPCRs (e.g., human endothelin ETB receptor)17,18,19,38,39,40,41,42.
In summary, these insights offer a comprehensive understanding of the mechanism of GPR103 activation upon QRFP binding. In the apo state, the N-terminal structure is hypothesized to be more labile than observed (Fig. 6a), due to its instability with merely two helices and a lower AlphaFold predictive score. QRFP may initially interact with either the ECD or TMD (Fig. 6b). Eventually, QRFP establishes stable binding to both domains, presumably maintaining the interaction of the C-terminus with the receptor while allowing for some fluctuation in the ECD and ligand-binding pocket in the TMD (Fig. 6c). This binding would reorganize the aromatic residue cluster in TM6, leading to the intracellular opening for receptor activation.

a–c Schematic representations of GPR103 activation model upon QRFP binding. From the apo state (a), the peptide binds (b) and changes to the active state (c).
GPCRs except for class A commonly possess N-terminal domains with various lengths and functional roles (e.g. PTH1R33) (Fig. 7a), depending on the class. Although most class A GPCRs simply consist of 7TM, recent cryo-EM structures have visualized functional N-terminal regions, A typical example, in thyroid-stimulating hormone receptor (TSHR43), a large extracellular domain with multiple leucine-rich repeats captures the ligand protein (Fig. 7b). In chemokine receptor CXCR244 and C5a receptor (C5aR)45, the N-terminus is elongated to the extracellular side and participates in the interaction with ligand protein (Fig. 7c, d). Although not as large as the LRR or the hotdog-line domain, the N-terminal HLH of GPR103, interacts with QRFP26 together with its elongated N-terminal region (Fig. 7e). In this respect, GPR103 is intermediate between GPCRs with large extracellular domains and others (CXCR2 and C5aR).

Furthermore, structural comparisons revealed unexpected homology in peptide ligand recognition between C5aR and GPR103 (Fig. 7d, e). As QRFP26, C5a has its C-terminus buried in the TMD, from which the α-helix extends vertically. C5a has several helices in its additional region, some of which closely resemble the N-terminal HLH of GPR103 and interact similarly with ECL2. This similarity is probably coincidental since C5aR and GPR103 are not strikingly sequence homologous compared to other GPCRs. For stable receptor-ligand interactions at the extracellular side, the C5a–C5aR pair has a larger ligand. By contrast, the QRFP26–GPR103 pair has the HLH at the N-terminus of the receptor to capture the ligand.
In the current study, we have not only elucidated the architecture of the receptor but also shed light on the structure of QRFP itself, potentially paving the way for the modulation of QRFP-neurons (Q-neurons)46. QRFP is a biomarker for neurons and is capable of inducing an extended state of hypothermia and reduced metabolism, akin to hibernation. The QRFP-induced hibernation-like state (QIH) provides an unusual model to study metabolic suppression for therapeutic purposes, as well as artificial hibernation. QIH through pharmacogenetics47 or optogenetics48 is achievable in murine models, but the prospect of non-genetically triggering these neurons might represent a groundbreaking stride in the realm of synthetic hibernation research. Nevertheless, Q-neurons constitute a remarkably limited subset, and currently, there are no established methodologies for their selective pharmacological activation. Among the myriad strategies proposed, one promising avenue is the engineering of synthetic low-molecular-weight binders targeting QRFP, with their integration into novel drug delivery systems. The field of protein design is undergoing a rapid metamorphosis, fueled by recent innovations in structural prediction via AlphaFold31 and the utilization of generative AI46. This is exemplified by the development of soluble GPCR variants and binders based on GPCR structures. Harnessing our structural insights to conceive binders that specifically target QRFP could facilitate studies of Q-neurons. Although this idea is still in its nascent stage, it harbors substantial promise for future applications, e.g. in emergency medical care.
Methods
Expression and purification of scFv16 and Nb35
The His8-tagged scFv16 was expressed and secreted by Sf9 insect cells, as previously reported49. The Sf9 cells were removed by centrifugation at 5000 × g for 10 min, and the secreta-containing supernatant was combined with 5 mM CaCl2, 1 mM NiCl2, 20 mM HEPES (pH 8.0), and 150 mM NaCl. The supernatant was mixed with Ni Superflow resin (GE Healthcare Life Sciences) and stirred for 1 h at 4 °C. The collected resin was washed with buffer containing 20 mM Tris (pH 8.0), 500 mM NaCl, and 20 mM imidazole, and further washed with 10 column volumes of buffer containing 20 mM HEPES (pH 8.0), 500 mM NaCl, and 20 mM imidazole. Next, the protein was eluted with 20 mM Tris (pH 8.0), 500 mM NaCl, and 400 mM imidazole. The eluted fraction was concentrated and loaded onto a Superdex200 10/300 Increase size-exclusion column, equilibrated in a buffer containing 20 mM Tris (pH 8.0) and 150 mM NaCl. Peak fractions were pooled, concentrated to 5 mg/mL using a centrifugal filter device (Millipore 10 kDa MW cutoff), and frozen in liquid nitrogen.
Nb35 was prepared as previously reported50,51. In brief, Nb35 was expressed in the periplasm of E. coli. The harvested cells were disrupted by sonication. Nb35 was purified by nickel affinity chromatography, followed by gel-filtration chromatography, and frozen in liquid nitrogen.
Constructs for expression of GPR103 and Gq
The human GPR103 gene (UniProtKB, Q96P65) was subcloned into a modified pEG bacmamvector, with an N-terminal haemagglutinin signal peptide followed by the FLAG epitope (DYKDDDDK) and the LgBiT fused to its C-terminus followed by a 3C protease site and EGFP-His8 tag. A 15 amino sequence of GGSGGGGSGGSSSGG was inserted into both the N-terminal and C-terminal sides of LgBiT. Rat Gβ1 and bovine Gγ2 were subcloned into pEG vectors respectively. In detail, rat Gβ1 was cloned with a C-terminal HiBiT connected with a 15 amino sequence of GGSGGGGSGGSSSGG. Moreover, mini-Gsqi was subcloned into the C-terminus of the bovine Gγ2 with a nine amino sequence GSAGSAGSA linker. The two resulting pEG vectors can express the mini-Gsqi trimer.
For the TGFα shedding assay and the flow cytometry analysis, we used human full-length QRFPR-encoding plasmids. Specifically, GPR103 was N-terminally modified with an HA signal sequence followed by a FLAG epitope and then a flexible linker (MKTIIALSYIFCLVFADYKDDDDKGGSGGGGSGGSSSGGG; the FLAG epitope is underlined) and inserted into the pCAGGS vector. Plasmid encoding GPR103 mutants were generated using Quickchange Site-Directed Mutagensis kit (Agilent).
TGFα shedding assay
The TGFα shedding assay was performed as described previously with minor modifications16. Plasmid transfection was performed in a 6-well plate with a mixture of 500 ng (per well in a 6-well plate) AP-TGFα-encoding plasmid and 200 ng GPR103-encoding plasmid. After 1-day culture, the transfected cells were harvested by trypsinization, pelleted by centrifugation at 190 × g for 5 min, and washed once with 5 mM HEPES (pH 7.4)-containing Hank’s Balanced Salt Solution (HBSS). After centrifugation, the cells were resuspended in 8 mL of the HEPES-containing HBSS. The cell suspension was seeded in a 96-well culture plate (cell plate) at a volume of 90 μL (per well hereafter) and incubated in a 37 °C incubator with 5% CO2 for 30 min. The cells were mixed with 10 µL of titrated concentrations of 10× QRFP26 (Peptide institute) (diluted in HBSS containing 5 mM HEPES (pH 7.4) and 0.01% (w/v) fatty acid-free BSA (Serva)), and incubated for 60 min. After spinning the cell plates at 190 × g for 2 min, 80 μL of conditioned media was transferred to an empty 96-well plate (conditioned media (CM) plate). Then, 80 μL of AP reaction solution (10 mM p-nitrophenylphosphate (p-NPP), 120 mM Tris-HCl (pH 9.5), 40 mM NaCl, and 10 mM MgCl2) was dispensed into the cell plates and the CM plates. Absorbance at 405 nm of the plates was measured using a microplate reader (SpectraMax 340 PC384, Molecular Devices) before and after 1-h incubation at room temperature. Ligand-induced AP-TGFα release was calculated by subtracting spontaneous AP-TGFα release signal from ligand-induced AP-TGFα release signal. The AP-TGFα release signals were fitted to a four-parameter sigmoidal concentration–response curve, using the Prism 10 software (GraphPad Prism). For each replicate experiment, the pEC50 parameter (negative logarithmic values of EC50 values) of the individual GPR103 mutants were normalized to that of the wild-type GPR103 performed in parallel, and resulting ∆pEC50 values were used to denote ligand response activity of the mutants.
Flow cytometry analysis
Cell surface expression of GPR103 was measured as described previously50. HEK293A cells were seeded in a 6-well culture plate at a concentration of 2 × 105 cells per mL (2 mL per dish) and cultured for 1 day before transfection. The cells were transfected with N-terminally FLAG-tagged GPCR construct along with the AP-TGFα reporter by following the same procedure as described in the TGFα shedding assay section. After 1-day culture, the cells were collected by adding 200 µL of 0.53 mM EDTA-containing Dulbecco’s PBS (D-PBS), followed by 200 µL of 5 mM HEPES (pH 7.4)-containing HBSS. The cell suspension was transferred to a 96-well V-bottom plate in duplicate and fluorescently labeled with an anti-Flag epitope (DYKDDDDK) tag monoclonal antibody (Clone 1E6, FujiFilm Wako Pure Chemicals) diluted in 2% goat serum and 2 mM EDTA-containing D-PBS (blocking buffer) and a goat anti-mouse IgG (H+L) secondary antibody conjugated with Alexa Fluor 488 (Thermo Fisher Scientific) diluted in the blocking buffer. After washing with D-PBS, the cells were resuspended in 200 μL of 2 mM EDTA-containing D-PBS and filtered through a 40-μm filter. The fluorescent intensity of single cells was quantified by a CytoFLEX flow cytometer (Beckman Coulter) equipped with a 488-nm laser. The flow cytometry data were analyzed with the FlowJo software (FlowJo). For each experiment, the mean fluorescent intensity (MFI) value of mutants was normalized to that of the wild-type GPR103 performed in parallel. Values of mean fluorescence intensity from approximately 20,000 cells per sample were used for analysis.
Expression and purification of the human GPR103 – Gq complex
The recombinant baculovirus was prepared using the Bac-to-Bac baculovirus expression system (Thermo Fisher Scientific). For expression, 0.8 L of HEK293S GnTI− (N-acetylglucosaminyl-transferase I–negative) cells (American Type Culture Collection, Cat. No. CRL-3022) at a density of 3 × 106 cells/mL were co-infected with baculovirus encoding GPR103, mini-Gsqi trimer and Gβ1 at the ratio of 2:1:1. Twenty hours after infection, 10 mM of Sodium Butyrate was added, and the cells were incubated at 30 °C. After 48 h, the collected cells were resuspended and dounce-homogenized in 20 mM Tris-HCl, pH 8.0, 150 mM NaCl, 10% Glycerol, 4 μM QRFP26, 5.2 μg/mL aprotinin, 2.0 μg/mL leupeptin, and 100 μM PMSF. After homogenization, Apyrase was added to the lysis at a final concentration of 25 mU/mL, and the lysate was incubated at room temperature for 1 h. The crude membrane fraction was collected by ultracentrifugation at 180,000 × g for 1 h and solubilized in buffer, containing 50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1.5% Lauryl Maltose Neopentyl Glycol (LMNG) (Anatrace), 0.15% cholesteryl hemisuccinate (CHS) (Merck), 10% glycerol, 5.2 μg/mL aprotinin, 2.0 μg/mL leupeptin, 100 μM PMSF, 25 mU/mL Apyrase, and 4 μM QRFP26 for 2 h at 4 °C. The supernatant was separated from the insoluble material by ultracentrifugation at 180,000 × g for 30 min and incubated with 4 mL of Anti-DYKDDDDK G1 resin (Genscript) for 1 h at 4 °C. The resin was washed with 20 column volumes of buffer containing 20 mM Tris-HCl, pH 8.0, 500 mM NaCl, 10% Glycerol, 0.01% LMNG, 0.001% CHS, and 0.1 μM QRFP26. The complex was eluted in buffer containing 20 mM Tris-HCl, pH 8.0, 150 mM NaCl, 0.01% LMNG, 0.001% CHS, 10 μM QRFP26, and 0.2 mg/mL FLAG peptide. The eluate was incubated with the Nb35 and scFv16 at 4 °C. The complex was concentrated and purified by size-exclusion chromatography on a Superose 6 increase (GE) column in 20 mM Tris-HCl, pH 8.0, 150 mM NaCl, 0.01% LMNG, 0.001% CHS, and 0.1 μM QRFP26. QRFP26 was added to the peak fraction to the final concentration of 4 µM and concentrated up to 13.8 mg/ml.
Sample vitrification and cryo-EM data acquisition
The purified complex was applied onto a freshly glow-discharged Quantifoil holey carbon grid (R1.2/1.3, 300 mesh), and plunge-frozen in liquid ethane by using a Vitrobot Mark IV. Data collections were performed on a 300 kV Titan Krios G3i microscope (Thermo Fisher Scientific) equipped with a BioQuantum K3 imaging filter and a K3 direct electron detector (Gatan).
First, 9555 movies were acquired with a calibrated pixel size of 0.83 Å pix−1 and with a defocus range of −0.8 to −1.6 μm, using EPU. Each movie was acquired for 2.6 s and split into 48 frames, resulting in an accumulated exposure of about 49.6 electrons per Å2 at the grid.
Image processing
All acquired movies were dose-fractionated and subjected to beam-induced motion correction implemented in RELION 3.152. The contrast transfer function (CTF) parameters were estimated using CTFFIND 4.053. A total of 11,085,317 particles were extracted. The particles were subjected to 2D classifications, Ab-initio reconstruction, and several rounds of hetero refinement in cryoSPARC32. Next, the particles were re-extracted and exported to RELION 3.154, then subjected to 3D classification with a mask on the receptor. Then, the particles were subjected to Bayesian polishing in RELION55. The particle sets were exported to cryoSPARCv4.4.0, and subjected to CTF refinement, and Non-uniform refinement, yielding a map with a global nominal resolution of 3.19 Å, with the gold standard Fourier Shell Correlation (FSC = 0.143) criteria56. Moreover, the 3D model was refined with a mask on the receptor. As a result, the receptor has a higher resolution with a nominal resolution of 3.29 Å. The overall and receptor-focused maps were combined by phenix57. The processing strategy is described in Supplementary Fig. 3.
To investigate the flexibility of QRFP26 and ECD of GPR101, the batch of particles was further processed by 3D flexible refinement32. After an overall non-uniform refinement, the mesh was prepared using a micelle-removed density map. Then, following the 3D flex train job and the reconstruction job, the flexibility of the QRFP26 and the ECD of GPR103 were visualized by the flex generate job. The processing strategy is described in Supplementary Fig. 6.
Model building and refinement
The density map was sharpened by phenix.auto_sharpen58 and the quality of the density map was sufficient to build a model manually in COOT59,60. The model building was facilitated by the Alphafold-predicted structure and cryo-EM structure of OX2R (PDB 7L1U (OxB-OX2R-mini-Gsqi structure))12. We manually fitted GPR103, the Gq heterotrimer, and scFv16 into the map. We then manually readjusted the model using COOT and refined it using phenix.real_space_refine57,61 (v.1.19) with the secondary-structure restraints using phenix secondary_structure_restraints. For the densities derived from 3D Flex, the rigid body refinement and all-atom refinement were performed on two representative frames from the default 40 frames using the model without 3D flex.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The cryo-EM density map and atomic coordinates for the QRFP26-GPR103 complex have been deposited in the Electron Microscopy Data Bank and the PDB, under accession codes: EMD-60096 and PDB 8ZH8. The tilted and upright state generated by the 3D are available at the Zenodo data repository (https://doi.org/10.5281/zenodo.11408292). All other data are available from the corresponding authors upon request. Source data are provided with this paper.
References
Bechtold, D. A. & Luckman, S. M. The role of RFamide peptides in feeding. J. Endocrinol. 192, 3–15 (2007).
Findeisen, M., Rathmann, D. & Beck-Sickinger, A. G. RFamide peptides: structure, function, mechanisms and pharmaceutical potential. Pharmaceuticals 4, 1248–1280 (2011).
Fukusumi, S., Fujii, R. & Hinuma, S. Recent advances in mammalian RFamide peptides: the discovery and functional analyses of PrRP, RFRPs and QRFP. Peptides 27, 1073–1086 (2006).
Pedragosa-Badia, X., Stichel, J. & Beck-Sickinger, A. G. Neuropeptide Y receptors: how to get subtype selectivity. Front. Endocrinol. 4, 5 (2013).
Fukusumi, S. et al. A new peptidic ligand and its receptor regulating adrenal function in rats. J. Biol. Chem. 278, 46387–46395 (2003).
Chartrel, N. et al. Identification of 26RFa, a hypothalamic neuropeptide of the RFamide peptide family with orexigenic activity. Proc. Natl Acad. Sci. USA 100, 15247–15252 (2003).
Takayasu, S. et al. A neuropeptide ligand of the G protein-coupled receptor GPR103 regulates feeding, behavioral arousal, and blood pressure in mice. Proc. Natl Acad. Sci. USA 103, 7438–7443 (2006).
Chartrel, N. et al. The neuropeptide 26RFa (QRFP) and its role in the regulation of energy homeostasis: a mini-review. Front. Neurosci. 10, 549 (2016).
Baribault, H. et al. The G-protein-coupled receptor GPR103 regulates bone formation. Mol. Cell. Biol. 26, 709–717 (2006).
Ukena, K., Osugi, T., Leprince, J., Vaudry, H. & Tsutsui, K. Molecular evolution of GPCRs: 26Rfa/GPR103. J. Mol. Endocrinol. 52, T119–T131 (2014).
Georgsson, J. et al. GPR103 antagonists demonstrating anorexigenic activity in vivo: design and development of pyrrolo[2,3-c]pyridines that mimic the C-terminal Arg-Phe motif of QRFP26. J. Med. Chem. 57, 5935–5948 (2014).
Hong, C. et al. Structures of active-state orexin receptor 2 rationalize peptide and small-molecule agonist recognition and receptor activation. Nat. Commun. 12, 815 (2021).
Mobbs, J. I. et al. Structures of the human cholecystokinin 1 (CCK1) receptor bound to Gs and Gq mimetic proteins provide insight into mechanisms of G protein selectivity. PLoS Biol. 19, e3001295 (2021).
Tang, T. et al. Receptor-specific recognition of NPY peptides revealed by structures of NPY receptors. Sci. Adv. 8, eabm1232 (2022).
Bruzzone, F. et al. Anatomical distribution and biochemical characterization of the novel RFamide peptide 26RFa in the human hypothalamus and spinal cord. J. Neurochem 99, 616–627 (2006).
Inoue, A. et al. TGFα shedding assay: an accurate and versatile method for detecting GPCR activation. Nat. Methods 9, 1021–1029 (2012).
Kim, K. et al. Structure of a hallucinogen-activated Gq-coupled 5-HT2A serotonin receptor. Cell 182, 1574–1588.e19 (2020).
Shihoya, W., Iwama, A., Sano, F. K. & Nureki, O. Cryo-EM advances in GPCR structure determination. J. Biochem. https://doi.org/10.1093/jb/mvae029 (2024).
Sano, F. K., Akasaka, H., Shihoya, W. & Nureki, O. Cryo-EM structure of the endothelin-1-ETB-Gi complex. eLife 12, e85821 (2023).
Thuau, R. et al. Structural studies on 26RFa, a novel human RFamide-related peptide with orexigenic activity. Peptides 26, 779–789 (2005).
Venkatakrishnan, A. J. et al. Molecular signatures of G-protein-coupled receptors. Nature 494, 185–194 (2013).
Xia, R. et al. Cryo-EM structure of the human histamine H1 receptor/Gq complex. Nat. Commun. 12, 2086 (2021).
Cao, C. et al. Structure, function and pharmacology of human itch GPCRs. Nature 600, 170–175 (2021).
Rasmussen, S. G. F. et al. Crystal structure of the β2 adrenergic receptor-Gs protein complex. Nature 477, 549–555 (2011).
Ji, Y. et al. Structural basis of peptide recognition and activation of endothelin receptors. Nat. Commun. 14, 1258 (2023).
Ballesteros, J. A. & Weinstein, H. Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. Methods Neurosci. 25, 63 (1995).
Le Marec, O. et al. Structure-activity relationships of a series of analogues of the RFamide-related peptide 26RFa. J. Med Chem. 54, 4806–4814 (2011).
Neveu, C. et al. Rational design of a low molecular weight, stable, potent, and long-lasting GPR103 aza-β3-pseudopeptide agonist. J. Med Chem. 55, 7516–7524 (2012).
Jumper, J. et al. Highly accurate protein structure prediction with AlphaFold. Nature 596, 583–589 (2021).
Tunyasuvunakool, K. et al. Highly accurate protein structure prediction for the human proteome. Nature 596, 590–596 (2021).
Varadi, M. et al. AlphaFold Protein Structure Database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 50, D439–D444 (2022).
Punjani, A. & Fleet, D. J. 3DFlex: determining structure and motion of flexible proteins from cryo-EM. Nat. Methods 20, 860–870 (2023).
Kobayashi, K. et al. Endogenous ligand recognition and structural transition of a human PTH receptor. Mol. Cell 82, 3468–3483.e5 (2022).
Kobayashi, K. et al. Cryo-EM structure of the human PAC1 receptor coupled to an engineered heterotrimeric G protein. Nat. Struct. Mol. Biol. 27, 274–280 (2020).
Fukuhara, S. et al. Structure of the human secretin receptor coupled to an engineered heterotrimeric G protein. Biochem Biophys. Res. Commun. 533, 861–866 (2020).
Park, C. et al. Structural basis of neuropeptide Y signaling through Y1 receptor. Nat. Commun. 13, 853 (2022).
Kang, H. et al. Structural basis for Y2 receptor-mediated neuropeptide Y and peptide YY signaling. Structure 31, 44–57.e6 (2023).
Akasaka, H. et al. Structure of the active Gi-coupled human lysophosphatidic acid receptor 1 complexed with a potent agonist. Nat. Commun. 13, 5417 (2022).
Chen, H. et al. Structure of S1PR2-heterotrimeric G13 signaling complex. Sci. Adv. 8, eabn0067 (2022).
García-Nafría, J., Lee, Y., Bai, X., Carpenter, B. & Tate, C. G. Cryo-EM structure of the adenosine A2A receptor coupled to an engineered heterotrimeric G protein. eLife 7, e35946 (2018).
Koehl, A. et al. Structure of the µ-opioid receptor-Gi protein complex. Nature 558, 547–552 (2018).
Shihoya, W., Sano, F. K. & Nureki, O. Structural insights into endothelin receptor signalling. J. Biochem 174, 317–325 (2023).
Duan, J. et al. Hormone- and antibody-mediated activation of the thyrotropin receptor. Nature 609, 854–859 (2022).
Saha, S. et al. Molecular basis of ligand promiscuity, structural mimicry, and atypical dimerization in the chemokine receptors. Preprint at bioRxiv https://doi.org/10.1101/2024.02.01.578380 (2024).
Wang, Y. et al. Revealing the signaling of complement receptors C3aR and C5aR1 by anaphylatoxins. Nat. Chem. Biol. 19, 1351–1360 (2023).
Watson, J. L. et al. De novo design of protein structure and function with RFdiffusion. Nature 620, 1089–1100 (2023).
Takahashi, T. M. et al. A discrete neuronal circuit induces a hibernation-like state in rodents. Nature 583, 109–114 (2020).
Takahashi, T. M. et al. Optogenetic induction of hibernation-like state with modified human Opsin4 in mice. Cell Rep. Methods 2, 100336 (2022).
Okamoto, H. H. et al. Cryo-EM structure of the human MT1-Gi signaling complex. Nat. Struct. Mol. Biol. 28, 694–701 (2021).
Nagiri, C. et al. Cryo-EM structure of the β3-adrenergic receptor reveals the molecular basis of subtype selectivity. Mol. Cell 81, 3205–3215.e5 (2021).
Nureki, I. et al. Cryo-EM structures of the β3 adrenergic receptor bound to solabegron and isoproterenol. Biochem. Biophys. Res. Commun. 611, 158–164 (2022).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Rohou, A. & Grigorieff, N. CTFFIND4: fast and accurate defocus estimation from electron micrographs. J. Struct. Biol. 192, 216–221 (2015).
Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. Elife 7, e42166 (2018).
Zivanov, J., Nakane, T. & Scheres, S. H. W. A Bayesian approach to beam-induced motion correction in cryo-EM single-particle analysis. IUCrJ 6, 5–17 (2019).
Rosenthal, P. B. & Henderson, R. Optimal determination of particle orientation, absolute hand, and contrast loss in single-particle electron cryomicroscopy. J. Mol. Biol. 333, 721–745 (2003).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 66, 213–221 (2010).
Terwilliger, T. C., Sobolev, O. V., Afonine, P. V. & Adams, P. D. Automated map sharpening by maximization of detail and connectivity. Acta Crystallogr. D Struct. Biol. 74, 545–559 (2018).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 66, 486–501 (2010).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132 (2004).
Afonine, P. V. et al. Real-space refinement in PHENIX for cryo-EM and crystallography. Acta Crystallogr. D Struct. Biol. 74, 531–544 (2018).
Acknowledgements
We thank K. Ogomori and C. Harada for technical assistance; Kayo Sato, Shigeko Nakano, and Ayumi Inoue (Tohoku University) for their assistance in the cell-based assays; Masataka Yanagawa (Tohoku University) for the setup of the flow cytometer. This work was supported by grants from the JSPS KAKENHI, grant numbers JP21H05037 (O.N.), JP22K19371 and JP22H02751 (W.S.), and JP21H04791, JP21H05113, and JP21H05037 (Asuka I.); the ONO Medical Research Foundation (W.S.); the Kao Foundation for Arts and Sciences (W.S.); the Takeda Science Foundation (W.S.); the Uehara Memorial Foundation (W.S.); the Lotte Foundation (W.S.); Japan Science and Technology Agency (JST), grant numbers JPMJFR215T and JPMJMS2023 (Asuka I.); the Japan Agency for Medical Research and Development (AMED), grant numbers JP22ama121038 and JP22zf0127007 (Asuka I.), JP233fa627001 (O.N.); 22ck0106533h0003 (O.N.); and the Platform Project for Supporting Drug Discovery and Life Science Research (Basis for Supporting Innovative Drug Discovery and Life Science Research (BINDS)) from AMED, grant numbers JP23ama121002 (support number 3272, O.N.) and JP23ama121012 (supporting number 6204, Asuka.I.).
Author information
Authors and Affiliations
Contributions
Aika I. performed the cryo-EM structural study. R.K. and Asuka I. performed and oversaw the mutagenesis study. H.A., F.S., and H.S.O. assisted with the cryo-EM data collection and the single-particle analysis. W.S. designed the experiments. The manuscript was mainly prepared by Aika I., and W.S., with assistance from O.N. Asuka I., W.S., and O.N. supervised the research.
Corresponding authors
Ethics declarations
Competing interests
O.N. is a co-founder and scientific advisor for Curreio. All other authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Iwama, A., Kise, R., Akasaka, H. et al. Structure and dynamics of the pyroglutamylated RF-amide peptide QRFP receptor GPR103. Nat Commun 15, 4769 (2024). https://doi.org/10.1038/s41467-024-49030-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-49030-5
This article is cited by
-
Structural insights into the selective recognition of RF-amide peptides by neuropeptide FF receptor 2
EMBO Reports (2025)
-
Insights into G-protein coupling preference from cryo-EM structures of Gq-bound PTH1R
Nature Chemical Biology (2025)
-
Structural insights into the agonist selectivity of the adenosine A3 receptor
Nature Communications (2024)
