
Abstract
The nucleus accumbens (NAc) is a key brain region for motivated behaviors, yet how distinct neuronal populations encode appetitive or aversive stimuli remains undetermined. Using microendoscopic calcium imaging in mice, we tracked NAc shell D1- or D2-medium spiny neurons’ (MSNs) activity during exposure to stimuli of opposing valence and associative learning. Despite drift in individual neurons’ coding, both D1- and D2-population activity was sufficient to discriminate opposing valence unconditioned stimuli, but not predictive cues. Notably, D1- and D2-MSNs were similarly co-recruited during appetitive and aversive conditioning, supporting a concurrent role in associative learning. Conversely, when contingencies changed, there was an asymmetric response in the NAc, with more pronounced changes in the activity of D2-MSNs. Optogenetic manipulation of D2-MSNs provided causal evidence of the necessity of this population in the extinction of aversive associations. Our results reveal how NAc shell neurons encode valence, Pavlovian associations and their extinction, and unveil mechanisms underlying motivated behaviors.
Similar content being viewed by others
Introduction
In a dynamic world, individuals are continuously flooded with sensory information of variable relevance. Our brains evolved to filter information and focus on relevant stimuli or cues predicting those stimuli. The ability to assign valence is essential to guide appropriate behavior and increase the chances of survival. Valence refers to the degree of attractiveness (positive valence) or aversiveness (negative valence) of a stimulus or outcome. From the behavioral point of view, a positive valence stimulus elicits approach whereas a negative valence stimulus triggers avoidance.
Several studies identified the NAc as essential in encoding rewarding and aversive information1,2,3. Seminal in vivo electrophysiological recordings report that NAc neurons innately respond to intraoral administration of appetitive unconditioned stimuli (US) such as sucrose, as well as to the aversive tastant quinine1,2. Interestingly, most neurons exclusively respond to positive or negative valence stimuli, though some respond to both2. NAc neurons also respond to reward/aversion-predictive cues (conditioned stimuli, CS)4, and accumbal activity is crucial for cue-outcome associations, i.e. Pavlovian conditioning4,5.
The NAc is mainly composed of GABAergic MSNs, segregated into those expressing dopamine receptor D1 (D1-MSNs) or D2 (D2-MSNs), though some neurons express both receptors6. The classical model in the field proposed a functional opposition of these two striatal subpopulations: D1-MSNs are thought to encode positive/rewarding stimuli whereas D2-MSNs encode negative/aversive stimuli7,8,9. However, this model fails to explain data from different studies10,11,12,13,14,15,16,17, which support a model where the two subpopulations work together to drive rewarding/aversive behaviors. Optical activation of either D1- or D2-MSNs supports self-stimulation, i.e. is reinforcing12. D1- or D2-MSN optical activation paired with a reward-predicting cue enhances motivation11,18,19. Moreover, distinct patterns of optical activation of D1- and D2-MSNs can trigger positive and negative reinforcement in the same animal10. Activation of NAc shell D1-MSNs inhibits palatable food consumption, and inhibition of these neurons promotes food consumption, even in satiated animals16,17.
Considering the available data and the proposed role for NAc as a crucial hub connecting limbic and motor systems, one can postulate that the NAc functions as a locus where bivalent valence information is encoded and used to guide directed approach/avoidance behavior20. Nevertheless, despite our understanding of the contribution of this region for behavior, it has remained technically challenging to differentiate the specific role of D1- and D2-MSNs in freely behaving animals due to their similar electrophysiological properties in extracellular recordings. In this context, the development of genetically encoded calcium indicators coupled with microendoscopic 1-photon imaging enables tracking the activity of the same neurons over multiple training and stimulus’ presentation sessions. Recent studies, including the one by Pedersen et al.21, show that NAc shell D1- and D2-MSNs present heterogeneous and divergent responses to rewards21,22,23. However, a study has proposed that NAc core neurons do not signal reward or valence24. Therefore, decades after the recognition of the NAc as a key brain region for cue-outcome association, a critical question remains unanswered: how do neurons in this brain region encode positive and negative valence stimuli and cue-outcome associations?
To address these questions, we monitored the activity of individual NAc medial shell neurons using the cre-dependent genetically encoded calcium indicator GCaMP6f through a miniaturized microscope in D1- or A2A-cre mice (A2A was used as a marker for D2-MSNs, since cholinergic interneurons also express D2 receptor) during exposure to appetitive and aversive stimuli and during Pavlovian conditioning. We decided to focus on NAc medial shell as most studies focused on the role of core subregion in reward/aversion and predictive cues encoding2. Core and shell subregions present distinct connectivity, and this translates into different characteristics (reviewed25). Our data shows that either NAc MSN population encode positive and negative valence unconditioned stimuli, but do not code CS valence. Surprisingly, individual neurons change their response to CS and US of either valence within trials (and across sessions), though population activity was stable and stereotypic and reliably encoded USs of opposing valence. Our data shows co-recruitment of both populations during appetitive and aversive Pavlovian conditioning, supporting a concurrent role in rewarding/aversive behaviors. We further show that D2-MSNs track US omission and extinction more prominently than D1-MSNs, and that optogenetic inhibition of D2-MSNs (but not D1-MSNs) delays extinction of aversive Pavlovian associations.
This work provides detailed functional and causal evidence regarding the role of NAc D1- and D2-MSNs in associative learning, which is of utmost importance to understand how this region contributes for motivated behaviors.
Results
Distinct representation of positive and negative valence stimuli in NAc medial shell D1- and D2-MSNs
To determine how NAc D1- and D2-MSNs represent stimuli of opposing valence, we recorded individual neurons in response to unsigned appetitive and aversive stimulus using a genetically encoded calcium indicator and a one-photon miniaturized microscope. For this, we injected an adeno-associated virus (AAV) expressing cre-dependent GCaMP6f in the NAc of D1-cre or A2A-cre mice, followed by gradient index (GRIN) lens implantation in the same location (Fig. 1A, B). Six weeks later, we imaged GCaMP6f signals during a sucrose session (US1; 15μl of 20% sucrose; 39 trials) and a shock session (US2; 0.5 mA, 1 s; 7 trials) (Fig. 1C). Accurate GRIN lens placement in the NAc and virus expression was assessed for all animals (Supplementary Fig. S1A).

A Schematic representation of calcium imaging recordings of GCaMP6f in NAc D1- or D2-MSNs using 1P-miniaturized microscopes; AP: anteroposterior. B Representative expression of GCaMP6f in the NAc, depicting GRIN lens location; scale bar: 250 μm. C Schematic representation of unexpected unconditioned stimulus (US) exposure sessions. US1 session consisted of 39 exposures to 15 ul of 20% of sucrose with 30 s ITI; US2 session consisted of 7 exposures to a 0.5 mA 1 s foot shock with variable ITI of 35–50 s. D Field of view of an example D1-cre mouse showing neurons responsive to US1 (green, left), to US2 (orange, middle) and to both (yellow, right); single traces of tracked neurons in both sessions in a sucrose session (left) and a shock session (right); vertical lines in traces represent events. E Heatmaps of responses of D1-MSNs to US1 and to US2 (nUS1 = 264; nUS2 = 227), and of F D2-MSNs (nUS1 = 164; nUS2 = 100). G Percentage of excited (red), inhibited (blue) and non-responsive (gray) D1-MSNs or H D2-MSNs in response to US1 and US2. I Percentage of each type of response of neurons tracked during US1 and US2 for D1-MSNs and J D2-MSNs. K Heatmap of individual D1-MSNs or L D2-MSNs showing variable responses throughout trials. M Correlation of activity of individual D1-MSNs or N D2-MSNs in response to US1 or US2. O Cartoon depicting population-based linear support vector machine (SVM) decoder (feature: population activity during stimulus; observations: average activity during a given trial; output: US1 or US2). P Classification accuracy for every mouse using population activity of D1- (two-tailed unpaired t-test) or Q D2-MSNs (two-tailed unpaired t-test) during US1 and US2 (0-3 s from US onset) (nD1-cre = 6, nA2A-cre = 6). The decoders were able to correctly identify sucrose and shock trials. Trajectories of trial-by-trial of R D1- or S D2-population responses to US1 and US2; Data from a representative mouse; Dots indicate stimulus onset; US1 and US2 trajectories move in orthogonal directions, supporting differential representation by D1- and D2-MSNs. Data are presented as mean values +/−SEM. **p < 0.01, ***p < 0.001. Source data are provided as a Source Data file. Created in BioRender. Pinto, L. (2024) BioRender.com/p41z755.
We aligned neuronal activity data to sucrose consumption, considering the first lick event after sucrose delivery, or to shock. We classified a neuron as being responsive if its activity during the stimulus was significantly different from baseline (p < 0.05, Permutation test). Neurons that responded to stimuli of either valence were distributed in the field of view with no obvious anatomical separation (Fig. 1D). Consistent with previous electrophysiological data2, sucrose consumption elicited mostly inhibitory responses in half of D1- or D2-MSNs (50% and 48% respectively); whereas shock triggered mostly excitations in both populations (85%; Fig. 1E–H).
To evaluate how the same neuron responded to stimuli of opposing valence, we analyzed the response of neurons that were tracked during sucrose and shock sessions (Fig. 1I, J) (tracked neurons activity was representative of the whole population - Supplementary Fig. S2A–D). The majority of D1- and D2-MSNs were responsive to both stimuli (67% and 78%, respectively; Fig. 1I, J). Either population presented mostly inhibitions to sucrose and excitations to shock (Fig. 1I, J). To further confirm these findings, we also characterized NAc D1- and D2-MSNs activity in response to other positive or negative valence stimuli - condensed milk and tail lift, respectively. Condensed milk led to the inhibition of 44% and 40% of D1- and D2-MSNs (Supplementary Fig. S2L–N), a smaller fraction than that observed for sucrose, which is largely consistent with the divergent response of NAc MSNs to different concentrations of sucrose21. Tail lift led to mostly excitatory responses in both subpopulations, in line with shock-evoked responses (Supplementary Fig. S2O–Q). These findings showing differential neuronal response to stimuli of opposing valence are in line to what is found in valence-encoding neurons from other brain regions26,27.
To assess the stability of neuronal responses to the same stimulus across trials, we categorized each unit into excited, inhibited or no change, based on unit average activity. Then, we calculated the fraction of persevered responses within sucrose or shock trials (analysis was performed for all recorded cells). Surprisingly, only a small percentage of D1- or D2-MSNs maintained their response to sucrose in more than 70% of the trials (example neurons in Fig. 1K, L; Supplementary Fig. S2E). Shock-responsive cells also changed responses throughout trials, albeit more consistent that for sucrose (Supplementary Fig. S2F). To further estimate response stability, we calculated the coefficient of correlation of activity of individual neurons across multiple trials of the same stimulus. Correlations were low for either stimulus in both MSN subpopulations (Fig. 1M, N). We also calculated Shannon’s entropy, a measure of the degree of variability in the neurons’ response. Entropy was high in sucrose trials for either D1- or D2-MSNs (Supplementary Fig. S2G), whereas it was lower in shock-related activity (Supplementary Fig. S2H). Importantly, the variability in individual response is not correlated with the time of the trial (Supplementary Fig. S2I). Altogether, these data indicate that individual D1- or D2-MSNs do not stably encode sucrose and shock throughout time.
Next, we trained a support vector machine (SVM) decoder to quantify how well individual neuron mean activity would predict trial type, i.e. distinguish sucrose from shock trials. There was a high variability in the accuracy of individual D1- or D2-MSNs (Supplementary Fig. S2J, K). Since individual NAc neurons appear to present time-dependent changes in coding properties, it is plausible that information is represented at the population level rather than at the individual level. Thus, we trained a new decoder using population responses (Fig. 1O), which accurately predicted sucrose and shock trials for D1- or D2-MSNs (86% and 77% accuracy, respectively; Fig. 1P, Q). We next sought to understand if the population represented the identity of individual stimuli or the valence of the stimuli. We observed that the decoder poorly distinguished between two positive valence stimuli - condensed milk and sucrose, though it accurately distinguished sucrose from tail lift (negative valence) (Supplementary Fig. S2R).
To further characterize how NAc activity to opposing valence stimuli unfolds over time, we calculated the neuronal trajectory, which describes the temporal evolution of neural population activity. For this, we examined trial-averaged trajectories of D1- or D2-population activity during each US. The trajectories of D1- or D2-MSNs during US1 moved in a distinct direction from US2 (Fig. 1R, S), supporting different representation of US1 and US2 by NAc neurons.
These results indicate that despite individual neuronal variability, NAc D1- and D2-population activity contains sufficient information to code positive and negative valence stimuli.
Representation of appetitive CS and US in NAc medial shell neurons during associative learning
After determining that NAc neurons distinguished opposing valence stimuli, we then sought to characterize how these neurons would respond during associative learning, in which an initially neutral cue acquires valence and motivational value with learning. Studies performed in other brain regions involved in Pavlovian conditioning have shown that neurons undergo dynamic modifications during learning, with the development of cue responses and/or transforming existing responses28,29. However, it is important to refer that cue-locked neuronal responses can reflect valence attribution, but may also reflect other features such as salience or prediction error24.
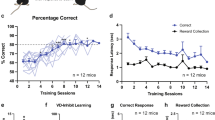
To understand how NAc neurons’ response to CS dynamically change with learning, we imaged animals during an appetitive Pavlovian associative learning task (Fig. 2A), in which animals learn to associate an auditory and visual cue (conditioned stimulus, CS1) with the delivery of sucrose (US1). As trial exposures progressed, mice increased the number of magazine pokes and presented reduced latency to obtain the reward after delivery, indicating successful learning (Fig. 2B, C; Supplementary Fig. S3A, B). We were able to record several hundreds of neurons for both populations throughout the days of conditioning (Fig. 2D–I). Throughout conditioning, 44% of D1-MSNs (Fig. 2D–F) and 48% of D2-MSNs presented excitations to CS1 (Fig. 2G–I). Around 1/3 of D1- or D2-MSNs presented cue inhibitions. Unexpectedly, the percentage of neurons that respond to CS1, US1 or both stimuli did not change significantly throughout learning (Fig. 2F, I; Supplementary Fig. S3C). The proportion of cue-excited or cue-inhibited neurons was remarkably stable throughout days, which contrasts with the amygdala, where an amplification in CS- (and US-) responsive neurons throughout learning was found28,29.

A Appetitive Pavlovian conditioning consisted in the association between a cue (conditioned stimulus - CS1; house light and 5-kHz tone) and the delivery of 15 μl of 20% sucrose solution (US1). B Average pokes during different days of Pavlovian conditioning (day 1, day 5 and day 10) of D1-cre mice or C A2A-cre mice, showing that animals increase pokes throughout learning (Friedman test; Dunn’s post hoc). D Heatmaps representing D1-MSN (nday1 = 708; nday5 = 761; nday10 = 699) or G D2-MSN (nday1 = 399; nday5 = 442; nday10 = 480) responses to CS1 (left) and US1 (right) on days 1, 5 and 10. In heatmaps, neurons are aligned by response to the CS. Percentage of responses of E D1-MSNs or H D2-MSNs to CS1 and US1 showing stability in the percentage of CS and US responses in both populations. F Pie chart with the percentage of responses throughout time for D1- or I D2-MSNs, demonstrating that the percentage of CS-US-responsive neurons does not change with learning. J Peri-stimulus time histogram (PSTHs) of cue-excited or L cue-inhibited D1-MSNs or N–P D2-MSNs tracked on different days of learning. K–M Area under the curve (AUCs) of mean cue response (0–3 s from cue onset) for D1- or O–Q D2-MSNs over the days; there is a substantial decrease in the magnitude of response to the cue of CS-excited neurons (and less for cue-inhibited) from day 1 to day 2, suggesting that part of the cue signal is due to novelty (Kruskal–Wallis test). The magnitude of response of CS-excited D2-MSNs during the cue period is higher than in D1-MSNs. Data are presented as mean values +/−SEM. *p < 0.05, **p < 0.01, ***p < 0.001. Source data are provided as a Source Data file. Created in BioRender. Pinto, L. (2024) BioRender.com/p41z755.
Learning can also link CS and US representations increasing correlation of responses throughout time28. Hence, we evaluated how CS responses were correlated with US responses on early, middle and late learning stages (day 1, 5 and 10, respectively). Contrary to what was expected, no increase in correlation between CS and US responses for either NAc D1- or D2-MSNs was found, even when we restrict to CS-US-responsive neurons (Supplementary Fig. S3D, E).
To better understand what happens to the activity of MSNs during learning, we plotted the average activity of cue-excited and cue-inhibited neurons during different learning stages. The magnitude of response of cue-excited (Fig. 2J, K, N, O), but not cue-inhibited (Fig. 2L, M, P, Q) neurons was higher in D2-MSNs in comparison to D1-MSNs, suggesting a differential contribution of these populations in cue encoding. However, this comparison should be interpreted with caution since magnitude changes may also arise from differences in sensor expression between groups.
Surprisingly, we observed a reduction in the magnitude of response of cue-excited and cue-inhibited D1- or D2-MSNs from day 1 to day 2 (Fig. 2J–Q), suggesting that part of the observed changes are due to novelty. In further support, when we compare the cue activity of the first trials with the latest trials of day 1, we observe a decrease in the magnitude of excitatory and inhibitory responses for D1-MSNs, though not significant for D2-MSNs (Supplementary Fig. S3F, G). Nevertheless, novelty per se does not explain the robust CS responses observed on later stages of conditioning. Recent work has proposed that NAc D2-MSNs encode valueless prediction error or signal errors24,30. In prediction error neurons, cue responses should develop with learning and the response to the US decrease as it becomes more predictable by the CS31, which we do not observe in either D1- or D2-MSNs.
Altogether, our data shows NAc cue- (and US-) evoked neuronal activity does not evolve with time, suggesting that cue responses may reflect other features of the stimulus, rather than classical prediction errors.
Drift in the representation of appetitive stimuli in NAc medial shell neurons throughout days
Next, we took advantage of the ability to monitor individual cells throughout time to allow a more comprehensive insight on how individual neurons change throughout associative learning. For this, we followed individual neurons on days 1, 5 and 10 (272 D1-MSNs; 172 D2-MSNs) corresponding to early, mid and late training stages (representative animal in Fig. 3A; all neurons in Fig. 2D–I, tracked neurons in Supplementary Fig. S4A–F).

A Field of view of a representative animal used for neuronal tracking during appetitive Pavlovian conditioning. B Number of neurons presenting each type of response of tracked D1-MSNs or C D2-MSNs on day 1, 5 and 10 of conditioning, showing variability in individual CS1-US1 responses over days (nD1 = 272, nD2 = 172). D Data from individual D1-MSNs’ activity was used to train a SVM decoder for the identification of CS1 and US1 (features: single neuron activity during stimulus; observations: average activity during given trial; output: CS1 or US1). Neurons are sorted and colored by decoding accuracy on day 1; the same was done for E D2-MSNs. F, G D1- or D2-MSNs with higher decoding accuracy on day 1 did not necessarily have relevant decoding accuracies throughout days. Colored dots represent accuracies on day 1 and gray dots represent the decoding accuracy of the same cells in days 5 or 10. H, I Decoding accuracies for all D1- or D2-MSNs and for top or bottom 20% cells in terms of accuracy. The decoding accuracy of the top 20% cells on day 1 largely overlapped with the entire population on days 5 and 10 for both populations. J Cartoon depicting Population-based SVM decoder (features: population activity during stimulus; observations: average activity during given trial; output: CS1 or US1). K SVM classification accuracy using population activity during CS1 and US1 (0–3 s from CS or US onset) for D1-MSNs (One-way ANOVA; Bonferroni post hoc) or L D2-MSNs (nD1-cre = 11, nA2A-cre = 9). The population decoders efficiently identified CS1 and US1 events for both populations. M Population vector correlation of CS1 responses of D1- or D2-MSNs within and between sessions (nD1-cre = 10, nA2A-cre = 9; Two-Way ANOVA, Bonferroni post hoc). N Population vector correlation of US1 responses of D1- or D2-MSNs, suggesting representational drift of US encoding between sessions (nD1-cre = 9, nA2A-cre = 9; Two-Way ANOVA, Bonferroni post hoc). Data are presented as mean values +/−SEM. *p < 0.05, **p < 0.01. Source data are provided as a Source Data file.
The majority of D1- and D2-MSNs (~70%) responded to both CS1 and US1 in all stages of learning (Supplementary Fig. S4C, F). In line with stochastic encoding of USs at a trial-by-trial level (Fig. 1), we also observed that D1- and D2-MSNs change their type of response to the CS1 and US1 trial-by-trial (Supplementary Fig. S4G, H), and between days (Fig. 3B, C; Supplementary Fig. S4I, J). Supporting previous data, a decoder trained with the activity of each individual neuron had low accuracy in distinguishing CS1 and US1 (Supplementary Fig. S4K). Even the top 20% D1- or D2-MSNs with high decoding accuracies on day 1 did not perform well on following days (Fig. 3D–I).
The previous data strongly suggested drift in the representation of stimuli by individual neurons, thus we hypothesized that CS and US representations were encoded at the population level. Thereafter, we trained a decoder using population responses to CS1 and US1 on days 1, 5 or 10 (Fig. 3J). For either population, the decoder efficiently distinguished CS1 and US1. Surprisingly, accuracy was higher on day 1 than on subsequent days (Fig. 3K, L).
Our data showed that despite changes in coding of individual cells, population activity patterns can be used to distinguish CS and US events on different days, so we next asked if these activity patterns would be stable throughout time. To study the stability of the population representation over timescales of minutes (within session) and days (between sessions), we computed the Pearson’s correlation between pairs of trial-averaged population vectors (PV) for each stimulus within and across sessions. We observed a reduction in PV correlation between sessions for CS and for US, in comparison to within session (Fig. 3M, N).
Altogether, these data support representation of CS1 and US1 at a population-level in both D1- and D2-MSNs and suggests that these populations exhibit representational drift over sessions.
Representation of aversive CS and US in NAc medial shell neurons during associative learning
Subsequently, we aimed to determine if NAc MSNs were involved in negative valence Pavlovian associations, thus we trained the same mice to associate another auditory and visual cue (CS2) with the delivery of a mild foot shock (US2) (Fig. 4A). Animals’ conditioned responses, measured as freezing behavior, increases throughout trials in D1-cre and A2A-cre mice (Fig. 4B–E), indicative of learning.

A Aversive Pavlovian conditioning involved learning to associate an auditory cue (CS2; 3-kHz tone) with a 0.5 mA 1 s foot shock (US2). B, C Freezing response progression over trials of the aversive conditioning session for D1-cre (4B: Friedman test; 4 C: two-tailed paired t-test) and D, E A2A-cre mice (4D: Friedman test; 4E: two-tailed paired t-test; nD1-cre = 14, nA2A-cre = 12). F Heatmaps representing D1-MSN responses to CS2 (left) and US2 (right). In heatmaps, neurons are aligned to response to CS2. G Percentage of each type of D1-MSNs response during the initial trials (1-2), last trials (6-7) and all trials. H Pie chart demonstrating that the percentage of CS-US-responsive D1-MSNs does not substantially change throughout trials. I Heatmaps representing D2-MSN responses to CS2 (left) and US2 (right). J Percentage of each type of D2-MSNs response. K Pie chart demonstrating that the percentage of CS-US-responsive D2-MSNs does not change throughout trials. L PSTHs of cue-excited or N cue-inhibited D1-MSNs or P–R D2-MSNs tracked on different trials. M–O AUCs of cue response (0–3 s from cue onset) for D1- or Q–S D2-MSNs over the days. No major differences were observed in the magnitude of responses throughout trials. T Activity of individual D1-MSNs from the first 2 trials and to the last 2 trials, showing variability in individual responses over trials; similar findings were observed for U D2-MSNs. V Cartoon depicting population based SVM decoder (features: population activity during stimulus; observations: average activity during given trial, output: CS2 or US2). W SVM classification accuracy for every mouse using population activity during CS2 and US2 (0-3 s from CS or US onset) for D1-MSNs or X D2-MSNs (two-tailed unpaired t-test). D2-MSNs population activity was more accurate in the categorization of CS2 and US2 events than D1-MSNs. Data are presented as mean values +/−SEM. ***p < 0.001. Source data are provided as a Source Data file. Created in BioRender. Pinto, L. (2024) BioRender.com/p41z755.
We observe mostly excitatory responses to CS2 or US2 in early trials (trials 1-2) and in late trials (trials 6-7) of aversive conditioning for both populations (Fig. 4F, G, I, J). The majority of D1- and D2-MSNs responded to both CS2 and US2 (Fig. 4H, K; Supplementary Fig. S5A). Regardless of the neuronal population, most neurons presented excitatory responses to CS2 and US2.
To evaluate the effect of learning cue-evoked activity, we plotted the average response of cue-excited and cue-inhibited neurons on early and late trials. The response of cue-excited or cue-inhibited neurons did not significantly change throughout time (Fig. 4L–S). However, the magnitude of excitatory response to the CS was higher in D2-MSNs than in D1-MSNs (Fig. 4L, P).
Next, we intended to observe if the responses to CS and US were more correlated in later trials. We observed a modest effect of learning in the correlation analysis between CS and US responses in NAc D1-MSNs, but not in D2-MSNs (Supplementary Fig. S5B, C).
As observed for appetitive conditioning, individual neurons changed their response throughout trials for both D1- and D2-neurons (Fig. 4T–U), as supported by the distribution of the correlation coefficients of single neuron activity (Figure S5D), but they could still decode CS2 and US2 at the single neuron level (Supplementary Fig. S5E). Because of the observed changes in individual neurons’ response, CS2 and US2 representations are also likely encoded at population level, akin to appetitive associations. Thus, we trained a decoder using the population responses to CS2 and US2 (Fig. 4V). The D1-population decoder distinguished CS2 or US2 events from shuffle data but presented lower accuracy (Fig. 4W); conversely, D2-population activity presented higher accuracy in classifying CS and US events (Fig. 4X).
In summary, we found that NAc D1- and D2-population contains sufficient information to distinguish the identities of CS2 and US2, despite individual neuronal variability.
Similar functional clusters between D1- and D2-MSNs during Pavlovian conditioning
After determining that D1- and D2-population activity could be used to distinguish opposing valence USs, and those events from cues, we sought to identify functional ensembles containing neurons with similar patterns of activity that could better represent the influence of learning in this region. To do this, we performed principal component analysis (PCA) on neuronal responses to each CS and US on day 1, followed by K-means clustering (Supplementary Fig. S6A–D). This unbiased approach identified three remarkably similar clusters on the appetitive conditioning for D1-MSNs and D2-MSNs (Supplementary Fig. S6E, F). Since we aimed to monitor the evolution of clusters’ activity throughout learning, we performed the same clustering analysis using only neurons tracked on days 1, 5 and 10. Analogous functional clusters were found for each neuronal population (Fig. 5A, B), that represented bona fide activity of the whole population (Supplementary Fig. S6E, F). Importantly, clustering analysis of either population based on the activity of day 10 originated similar functional clusters (Supplementary Fig. S6G, H), suggesting constancy of the pattern of activity at a population level.

Heatmaps and representation of neuronal clusters based on CS1 and US1 responses on day 1 of Pavlovian conditioning for A D1-MSNs and for B D2-MSNs, and the evolution of their response throughout days 5 and 10. Three similar functional clusters were found for each population. Neurons on heatmaps (left) are organized by CS response. C Representation of neuronal clusters based on CS2 and US2 responses during aversive Pavlovian conditioning for D1-MSNs and for D D2-MSNs, and the evolution of their response from early to late trials. Three similar functional clusters were found for each population. Neurons on heatmaps (left) are organized by CS response. PSTH data are presented as mean values +/−SEM.
Then, we trained a SVM decoder using the neuronal activity of each cluster. For either population, cluster 2, which present a robust CS1 excitation and US1 inhibition, presented the higher decoding accuracy on day 1 (Supplementary Fig. S6I–J; cluster 2 represented in Fig. 5A, B). To comprehend the temporal evolution of these clusters, we followed them throughout learning. Cluster 2 decrease the magnitude of CS response both for D1- and D2-MSNs (Supplementary Fig. S6K, L). This suggests that D1- or D2-MSNs’ cue responses do not robustly develop as a function of CS-US associative learning, as observed in VTA neurons for example32. Intriguingly, this cluster presents a subtle inversion in US1 response from day 1 to day 10 in both populations (Supplementary Fig. S6K, L).
We subsequently used the same unbiased clustering strategy to classify D1- or D2-MSNs into functional clusters for the aversive conditioning. For either D1- and D2-populations, three similar clusters were found (Fig. 5C, D). In the case of D1-MSNs, the cluster with higher decoding accuracy was cluster 2, that presents CS2 excitation followed by robust US2 excitation (Supplementary Fig. S6M). In the case of D2-MSNs, cluster 1 was the best performer (Supplementary Fig. S6N). Then, to observe if the activity of these clusters changes throughout conditioning, we aligned their activity from early to late trials. Throughout trials, D1-MSN cluster 2 and D2-MSN cluster 3 increase the magnitude of CS excitation, whereas US responses tend to be attenuated in later trials (Supplementary Fig. S6O, P).
Our findings indicate that D1- and D2-neuronal populations form remarkably similar functional clusters in either appetitive or aversive Pavlovian conditioning, indicating that both populations are similarly co-recruited during behavior, diverging from the classical model of striatal functional opposition.
Valence of USs, but not of CSs, is encoded by NAc medial shell MSNs
So far, our data showed that NAc D1- and D2-MSNs can encode positive and negative valence USs (Fig. 1) and distinguish those from CSs (Figs. 3 and 4). However, it was still unclear if these neuronal populations contain sufficient information to distinguish appetitive and aversive cues. To address this question, we tracked neuronal activity on day 10 of appetitive Pavlovian conditioning and on day 1 of aversive Pavlovian conditioning (Fig. 6A, C). More than 60% of D1- or D2-MSNs responded to both CS1 and CS2, and most of the CS-responsive neurons presented excitations, independently of CS valence (Fig. 6A–D). This observation challenges CS valence-encoding by these neurons, as one would expect differential responses to either valence cue28,29. Nevertheless, there was a smaller subset of neurons (24% of D1- and 27% of D2-MSNs) that responded in opposite manner for the two CSs, which could code valence. To observe if D1- or D2-neuronal activity could be used to classify event type, we used population activity patterns during CSs and USs to train a decoder. Population responses of D1-MSNs efficiently decoded sucrose and shock trials, but not the identity of CS1 and CS2 (Fig. 6E, F). Similarly, D2-neuronal population activity could be used to segregate sucrose and shock trials but poorly distinguished CS1 from CS2 (Fig. 6E, F).

Heatmaps representing single neuronal responses to CS1 and CS2 for D1-MSNs tracked during day 10 of appetitive conditioning and during day 1 of aversive conditioning for A D1-MSNs and C D2-MSNs. B Percentage of each type of CS1-CS2 response for both neuronal subtypes. D Percentage of each type of US1-US2 response for both neuronal subtypes. E, F The activity of D1- or D2-MSNs during CS1, CS2, US1 or US2 was used to create a population-based support vector machine decoder (features: population activity during each stimulus; observations: average activity during given trial; output: CS1 or CS2; US1 or US2). The decoder trained with D1-population data does not distinguish CSs of opposing valence, but clearly distinguishes USs of opposing valence (two-tailed paired t-test). The decoder trained with D2-population data poorly distinguishes CS1 and CS2 trials (two-tailed paired t-test), though it is accurate in identifying US1 and US2 trials (two-tailed paired t-test; nD1-cre = 14, nA2A-cre = 10. G Representative neuronal trajectories of one animal, depicting the trial responses to CSs or USs of opposing valence using trial-based activity data from day 10 of appetitive Pavlovian conditioning and from day 1 of aversive Pavlovian conditioning, and quantification of Euclidean distances for the trajectories of D1-MSNs (two-tailed unpaired t-test). H Representative neuronal trajectories and quantification of Euclidean distances for the trajectories of D2-MSNs (Mann–Whitney test). In trajectories, dashed lines represent trials, full lines represent median. Note that CS1 and CS2 trajectories are intermingled, whereas USs trajectories move in distinct directions in both populations. Data are presented as mean values +/− SEM. *p < 0.05, **p < 0.01, ***p < 0.001. Source data are provided as a Source Data file.
We also computed the neural trajectory to CSs and USs to visualize the population activity patterns. For both D1- and D2-MSNs, CS1 and CS2 trajectories presented high trial-to-trial variability, resulting in trajectories’ overlap and low mean Euclidean distances over time (Fig. 6G, H). Conversely, US1 and US2 trajectories remained distinct and exhibited higher mean Euclidean distances, like in the preconditioning phase (as depicted in Fig. 1R, S).
Our results show that the pattern of activity of D1- and D2-MSNs during CS1 and CS2 is similar, arguing against CS-valence encoding by these neurons. Since many D1- or D2-MSNs responded to both CS1 and CS2 in the same manner, this suggests that these neurons can encode CS salience, since it is expected that these types of neurons respond in the same direction regardless of valence33.
D1- and D2-MSNs respond to unexpected US omission
Our previous data suggest that CS responses encode other features of the stimuli rather than valence. The lack of CS-US changes throughout learning advocates against a classical prediction error encoding by NAc neurons. However, these findings do not preclude that NAc neurons are important for monitoring and updating outcomes and contribute for error signaling. To get further insight on this, we measured the activity of D1- and D2-MSNs in well-conditioned animals during unexpected US omission sessions.
After 10 days of appetitive conditioning, animals were subjected to a CS-US session in which reward was randomly omitted in 8 out of 30 trials. We evaluate neuronal activity during the first poke followed by lick event after CS1 (Supplementary Fig. S7A), to ensure that we align activity to consumption (or attempt to consume in the case of omission trials), and neurons were classified based on their average response to US consumption. No differences in CS1 responses were found between rewarded and omission trials, as expected, considering that animals do not anticipate US omission during the cue period (Supplementary Fig. S7B). D1- and D2-MSNs respond differently to reward delivery and in reward omission conditions. Sucrose-inhibited neurons no longer present inhibitions during omission in either population (Supplementary Fig. S7C, D), in further support of US-valence encoding by these neurons (Fig. 1). In fact, for both populations, neurons present an excitatory response to reward omission, with a more delayed response in D2-MSNs. Regarding sucrose-excited neurons, we observe that the two populations respond very differently, with D2-MSNs presenting a biphasic excitatory response during omission that was not observed in D1-MSNs (Supplementary Fig. S7C, D). This suggests that D2-MSNs can be important for error signaling, despite not behaving as a canonical prediction error neuron during conditioning (Figs. 2 and 4).
We also evaluated the activity of D1- and D2-MSNs during unexpected shock omission. In the day after the aversive conditioning, animals were recorded in a session in which shock was randomly omitted in 4 out of 11 trials (Supplementary Fig. S7E–I). No major differences in CS2 responses were found between trials, in agreement with the randomness of the shock omission (Supplementary Fig. S7F). Regarding response to shock omission, we observe a robust increase in the number of inhibited neurons in both populations (Supplementary Fig. S7G). This can reflect outcome updating, though another parsimonious explanation is that these changes signal a positive valence outcome, due the omission of an expected noxious stimulus. Importantly, there was higher percentage of D2-inhibited neurons during shock-omission in comparison to D1-MSNs (Supplementary Fig. S7G).
Altogether, our results suggest that, while D2-MSNs signal omissions of rewarding and aversive stimuli, they signal reward omission more prominently than D1-MSNs, supporting a model in where these neurons monitor and update outcomes.
Differential contribution of NAc medial shell D1- and D2-MSNs during extinction learning
Continuous omission of USs will eventually extinguish the learned association. Extinction is a form of learning that is thought to involve new brain plasticity that encodes a “CS-no US” association, though there can also occur the degradation of the previous association34. We recorded D1- and D2-MSNs activity in extinction conditions (CS, no US). After appetitive Pavlovian conditioning, animals were subjected to three extinction sessions, in which the CS1 was presented but no US1 (sucrose) was given (Supplementary Fig. S8A). D1- and A2A-cre animals present reduced conditioned responses since extinction session 1 (Supplementary Fig. S8B, C). Regardless of the neuronal population, most recorded cells presented excitatory responses to CS1 during extinction days (Supplementary Fig. S8D–G). These results were also confirmed with data of neurons tracked on day 10 of Pavlovian conditioning and on extinction days 1 and 3 (Supplementary Fig. S8H–K).
Next, we evaluated the responses of D1- and D2-MSNs during extinction of aversive Pavlovian associations. After aversive Pavlovian conditioning, animals were subjected to nine days of extinction, being exposed to CS2 but with no shock (US2) delivered (Fig. 7A). Freezing was used as a measure of conditioned responses. Throughout extinction sessions, both D1- and A2A-cre mice reduced freezing behavior, as expected (Fig. 7B, C). Of note, there was an uneven response of D1- and D2-MSNs to the CS in extinction conditions (Fig. 7D–H – tracked neurons; data from all recorded neurons in Supplementary Fig. S9A–E), since the percentage of excitatory and inhibitory responses throughout extinction is divergent. The percentage of CS2-excited D1-MSNs increases on extinction day 1 from 63% to 80% but substantially decreases on extinction days 5 and 9 (Fig. 7D). Conversely, the percentage of D1-inhibited cells increases throughout extinction. In contrast, D2-MSNs preserve the percentage of CS2-excited or CS-inhibited responses throughout extinction (Fig. 7D). The magnitude of CS-excitatory response throughout extinction appeared to be more prominent in D2-MSNs in comparison to D1-MSNs throughout days (Fig. 7E–H), though this should be interpreted with caution due to the caveats of comparing different experimental groups using fluorescent sensors.

A After aversive Pavlovian conditioning, animals were exposed to 9 days of extinction (CS2, no US2). B, C Freezing responses of D1-cre or A2A-cre animals, respectively, showing a decrease in aversive conditioned responses throughout extinction. D Percentage of excitatory and inhibitory responses to CS2 in the day of aversive conditioning and in extinction sessions 1, 5 and 9 for D1-MSNs and D2-MSNs. E Heatmaps of tracked D1-MSNs or G tracked D2-MSNs showing the CS2 responses throughout days. In heatmaps, neurons are aligned to CS2 response on the conditioning day. F AUCs of D1-MSNs or H D2-MSNs CS responses throughout the days. I Scheme of the optogenetic manipulation during aversive Pavlovian conditioning. D1- and A2A-cre mice were exposed to a fear conditioning session without optogenetic manipulation. After, mice were exposed to 9 days of extinction, with optical excitation or optical inhibition of D1- or D2-MSNs during the 10 seconds of CS exposure (excitation: 25 ms light pulses at 20 Hz; 473 nm; nD1-YFP = 19, nD1-ChR2 = 20; nA2A-YFP = 19, nA2A-ChR2 = 17; inhibition: constant light; 589 nm; nD1-YFP = 19 nD1-NpHR = 17; nA2A-YFP = 19, nA2A-NpHR = 18). J Optogenetic excitation or K inhibition of D1-MSNs during extinction sessions does not alter the extinction of conditioned responses. L Optogenetic excitation of D2-MSNs during extinction sessions does not alter conditioned responses. M Optogenetic inhibition of D2-MSNs during CS period in extinction sessions delays the extinction of conditioned responses, as shown by the increased freezing behavior in relation to YFP animals (Two-way ANOVA, Bonferroni post hoc). Data are presented as mean values +/−SEM. **p < 0.01, ***p < 0.001. Source data are provided as a Source Data file. Created in BioRender. Pinto, L. (2024) BioRender.com/p41z755.
Together, these results indicate that D2-MSNs have a sustained CS-excitatory response profile under extinction conditions, and that the change in D1-MSN activity throughout days may reflect a change in perceived salience of the CS.
Optogenetic manipulation of NAc medial shell D2-MSNs during aversive CS delays extinction of conditioned responses
The differential temporal dynamics in CS response between D1- and D2-MSNs, suggests that the two populations have distinct contributions in extinction conditions. Considering the sustained response of D2-MSNs and the magnitude of their response throughout extinction sessions, we hypothesized that by inhibiting these neurons, one could modulate the extinction association and change conditioned responses. To test this hypothesis, we injected D1- or A2A-cre animals with an AAV carrying cre-dependent expression of an inhibitory opsin in the NAc (eNpHR), or an excitatory opsin (ChR2) or with control YFP virus and implanted an optic fiber for optogenetic manipulation (Fig. 7I; Supplementary Fig. S1B, C).
Animals were trained in the aversive Pavlovian conditioning, in which CS2 was followed by the delivery of a foot shock (7 pairings). After conditioning, animals were subjected to 9 days of extinction (CS2 only, no shock), in which we optically inhibited or excited D1- or D2-MSNs during the full cue period (Fig. 7I). Optical excitation or inhibition of D1-MSNs during cue period in extinction conditions did not alter the slope of extinction of conditioned responses in comparison to YFP control animals (Fig. 7J, K). Optical excitation of D2-MSNs during extinction sessions also had no effect in freezing (Fig. 7L). Remarkably, and supporting our hypothesis, optical inhibition of D2-MSNs during CS2 in extinction sessions significantly delayed the extinction of the conditioned response, since A2A-NpHR animals presented higher freezing behavior in comparison to control YFP group (Fig. 7M). Importantly, inhibition of D2-MSNs (or D1-MSNs) during the CS period of the conditioning day had no significant impact in freezing behavior (Supplementary Fig. S10A, B). No major differences in locomotor behavior were observed in any of the groups (Supplementary Fig. S10C, D).
Overall, this experiment demonstrates the essential contribution of D2-MSNs for the extinction of aversive Pavlovian associations.
Discussion
In this study, we examined the specific features of NAc medial shell D1- and D2-neuronal responses to positive and negative valence stimuli within the same individual and decoded their role in associative learning. We show that despite stochastic encoding at individual level, NAc medial shell D1- or D2-population activity reliably encodes positive and negative valence unconditioned stimuli. The two populations form remarkably similar functional clusters during Pavlovian conditioning, supporting a model where both populations simultaneously work together to drive appropriate associative learning. However, contrary to other brain regions involved in associative learning28,29, cue-evoked accumbens medial shell activity does not encode valence or canonical prediction errors. We show that D2-MSNs present a constant and robust response to reward omission, supporting a key role in monitoring and updating outcome information. In line, optogenetic inhibition of medial shell D2-MSNs delays extinction of aversive Pavlovian associations.
Here, we demonstrate that NAc medial shell MSNs, regardless of being D1- or D2-MSNs, presented mostly inhibitions to positive valence stimuli and excitations to negative valence stimuli. Our findings are in line with previous seminal electrophysiological studies of non-identified accumbal neurons in response to sucrose and quinine2,4. Nevertheless, it is important to refer that the sensory modality of the positive and negative valence stimuli of our experimental design was different (physical vs tastant), while in the previously mentioned study both stimuli were of the same modality (different tastants). Considering the recent hypothesis that NAc core D1-MSNs encode perceived saliency24,35, in future studies would be interesting to evaluate if/how NAc medial shell neurons respond to different modalities and/or intensities of the same modality stimuli. In line, a recent study using 2-photon calcium imaging showed that NAc medial shell D1- and D2-MSNs respond to rewards of different concentrations21.
Unexpectedly, the vast majority of NAc medial shell neurons do not reliably respond to the same stimulus similarly within and between days, presenting representational drift. Still, a stable representation of USs (and CSs) emerges at a population level, despite inherent variability of individual responses. These findings are important to consider in the interpretation of studies showing that different stimuli are represented in different neurons36, as distinct recruited neurons may just reflect a novel reconfiguration of the population response. Population-level coding with single-neuron variability has also been shown for sensory representation in parietal cortex37 or odor coding in the piriform cortex38. While neuronal drift appears counterintuitive in terms of neuronal representation, it can provide the flexibility and robustness of encoding that a brain region like the NAc requires. In a constantly changing environment, the presence of multiple ways of encoding the information provides redundancy and guarantees that different dimensions of the stimulus are integrated to create a coherent representation. Drift can also allow the adjustment of the strength and structure of synaptic connections, facilitating the encoding of new information and/or refining existing representations39. While drift can provide these advantages, it is still necessary to reliably encode information, which we do observe at population level for D1- and D2-MSNs. One could hypothesize that in the case of the NAc medial shell, ensembles containing several neurons can be used to integrate multiple signals including valence signals arising from the amygdala40, sensory inputs to the NAc41, or context information from the hippocampus42,43, creating a unified and comprehensive representation of a positive or negative valence stimulus. It is tempting to speculate that the stability in population responses, despite individual drift, can still convey a stable representation of information to downstream areas such as the ventral pallidum, to where both D1- and D2-MSNs project to44, and that has been shown to be a crucial region in integrating and responding to appetitive and aversive stimuli and predicting cues45,46.
Lesion and pharmacological studies show that the NAc is crucial for CS-US associations and the expression of conditioned approach responses47,48,49,50. A key finding from our study was the remarkable similitude in D1- and D2-MSNs responses during Pavlovian conditioning, with appetitive and aversive functional neuronal clusters of each population mirroring the activity patterns of the other. These findings demonstrate that both populations act in synchrony to code rewarding/aversive information, akin to studies in the dorsal striatum showing concurrent activation of D1- and D2-MSNs in action initiation14, and previous studies in the NAc using fiber photometry recordings24,51. The neuronal activity data is in agreement with behavioral studies showing that optogenetic manipulation of either D1- or D2-MSNs can drive place preference or place aversion in the same animal, depending on the pattern of activation of MSNs10.
In line with a relevant role for NAc in associative learning, electrophysiological studies of unidentified NAc neurons showed robust responses to CSs that develop with time2,52,53, consistent with a classic reward prediction error and/or valence attribution. We also found that the majority of D1- and D2-MSNs responded to appetitive and aversive CSs throughout conditioning. Interestingly, the magnitude of cue-evoked activity decreases considerably in the second day, indicating that part of the observed cue signal is due to novelty. Yet, cue-evoked activity in later stages of conditioning (and in extinction) implies that these neurons encode other features besides novelty, such as prediction errors, valence or salience24,28,29,33,54. A decoder trained with either D1- or D2-MSNs activity could not distinguish between opposing valence CSs, even when the association if fully established, which implies that NAc neurons encode valueless information about the cue, and that CS-valence signals are likely encoded in other regions such as the amygdala28,29. One possibility is that these neurons encode salience. Salience reflects the importance of the stimulus and refers to the ability of the stimulus to capture attention and promotes associative learning55. It is very difficult to disambiguate salience encoding from other features, as for example unexpected US omission is an error and a salient event. Since our data suggests that the observed CS-neuronal responses are a composite of different features, we need more sophisticated behavioral tasks to isolate and track each dimension.
A recent study by Zachry and colleagues proposed that NAc core D2-MSNs (but not D1-MSNs) encode reward prediction errors (RPE)24. This was because throughout aversive conditioning learning, authors found an increase in the percentage of CS-recruited neurons. This is in contrast with our data, as we did not find changes in the type of CS responses in NAc medial shell neurons throughout either appetitive or aversive conditioning (either by amplification of recruited neurons or increased correlation of CS-US activities), which argues against a classical prediction error encoding. The discrepancy between the two studies may be explained by anatomical specificities, considering the differential contribution of core and medial shell regions for Pavlovian conditioning56,57,58. Another important consideration is that, while we were able to track individual responses to positive and negative valence stimuli in the same neurons, the other study was mostly based on photometry recordings measurements, which were in line with findings from other studies51. Still, we observe that NAc medial shell D2-MSNs are activated during unexpected omission and play an important role during extinction, suggesting that they are important in error signaling/updating outcome information.
The fact that we did not observe evidence for clear RPE in NAc medial shell neurons is not surprising. Recent studies show regional heterogeneity and temporal dissociation of dopaminergic signals throughout the striatum, including in NAc core and shell subregions59. Moreover, the evidence for RPE dopamine signals in the core is stronger that in the shell59. In fact, a very interesting study has shown that cue-evoked dopamine signals emerge in the NAc core but not in shell60, and dopaminergic terminals in NAc shell do not appear to be crucial for cue-reward learning61.
Sparse evidence suggests that the NAc is important for reward extinction34,62,63 and pharmacological blockade of dopamine receptors in the NAc impairs fear extinction learning64. Extinction is a fundamental form of inhibitory learning that is important for adapting to changing contingencies within the environment. Despite D1- and D2-MSNs presenting remarkable similarities in activity during Pavlovian conditioning, their response was asymmetric during extinction of appetitive and aversive conditioned responses. In the extinction of appetitive conditioning, the activity of each population during the CS period was similar and stable throughout days. Conversely, in the extinction of aversive conditioning, the response of D2-CS-excited neurons was more pronounced than in D1-MSNs, in line with an important role of D2-MSNs in updating information. Importantly, optogenetic inhibition of D2-MSNs (but not D1) during extinction delays the suppression of conditioned freezing response. This is reminiscent of another study in which optogenetic inhibition of hippocampal neurons that were active during extinction increased freezing conditioned response after extinction training65. These findings suggest that D2-MSNs play an important role in aversive extinction learning and are particularly interesting in light of recent evidence showing that VTA dopaminergic signals to distinct NAc subregions are essential for extinction responses62,66. Interestingly, NAc lateral shell D2-MSNs are not involved in appetitive extinction learning67, further supporting the notion that different NAc sub-regions distinctively encode learning. Importantly, though D1-MSN optogenetic manipulation did not produce observable changes, the change in D1-MSNs activity throughout extinction days may reflect a change in CS salience due to the new rules, implying that D1-MSNs are also relevant to the extinction process. Future techniques that specifically target activated or inhibited subpopulations of MSNs may help to elucidate their distinct roles.
In sum, we showed that population activity of either D1- or D2-MSNs can be used to represent and discriminate positive and negative valence stimuli. Our data strongly favors a model where the two subpopulations are co-recruited to encode CS-US associations and elicit appetitive/aversive motivated behaviors. Moreover, we show that when contingencies change, D2-MSNs are essential for the extinction of aversive associations. These findings have broad implications since extinction learning constitutes a crucial component of current anxiety and post-traumatic stress disorder therapeutic interventions. Remarkably, manipulation of D2-MSNs (but not D1-) projecting to the ventral pallidum generates anxiety-like behavior68. Moreover, NAc dysfunction has also been found in other neuropsychiatric disorders, namely depression and addiction69,70, which highlights the need for further investigations to unravel the distinct contribution of different NAc neurons in the development of maladaptive behaviors.
Methods
Lead contact
Further information and requests for resources and reagents should be directed to and will be fulfilled by the lead contacts, Ana João Rodrigues (ajrodrigues@med.uminho.pt) and Carina Soares-Cunha (carinacunha@med.uminho.pt).
Subjects
Male and female heterozygous D1-cre (line EY262, Gensat.org) and A2A-cre (line GK139, Gensat.org) transgenic mouse lines (2-3 months of age) with a C57BL/6J background were used. All animals were maintained under standard laboratory conditions: an artificial 12 h light/dark cycle with lights on from 8 am to 8 pm; with an ambient temperature of 21 ± 1 °C and a relative humidity of 50–60%. Mice were housed in type 2L home cages with a maximum of 6 mice per cage, with food (standard diet 4RF21, Mucedola, Italy) and water ad libitum, unless stated otherwise. After surgery, animals were maintained in pairs, without physical access to one another (using a cage divider) to avoid damaging of the implants.
Behavioral experiments were performed during the light period of the light/dark cycle. Handling was performed for 10 minutes a day, starting at least one week before behavioral experiments. Animals were habituated to behavioral apparatuses for 3 consecutive days for 15 min before the behavioral tasks. Sample size used in behavioral tests was chosen according to previous studies; the investigator was not blind to the group allocation during behavioral performance, but it was blind in data analysis.
All procedures involving mice were performed according to the guidelines for the welfare of laboratory mice as described in the European Union Directive 2010/63/EU. All protocols were approved by the Ethics Committee of the Life and Health Sciences Research Institute (ICVS) and by the national authority for animal experimentation, Direção-Geral de Alimentação e Veterinária (DGAV; approval reference #8332). Health monitoring was carried out according to FELASA guidelines and all experimenters and animal facilities are accredited by DGAV.
Surgeries
Surgeries were performed under sterile conditions and sevoflurane (2–3%, plus oxygen at 1–1.5 l/min) anesthesia on a stereotactic frame (David Kopf Instruments, Model 940). Throughout each surgery, mouse body temperature was maintained at 36 °C using an animal temperature controller (ATC2000, World Precision Instruments) and afterward, each mouse was allowed to recover from the anesthesia in its homecage under a heating lamp. The mouse head was shaved, cleaned with 70% alcohol and a small incision from anterior to posterior was made on the skin to allow for aligning the head and drilling the hole for the injection site.
Imaging
Each animal was unilaterally injected with 400 nl of AAV5-CAG-Flex-GCaMP6f-WPRE-SV40 (Addgene) into the right NAc (AP: 1.45 mm, ML: 0.6 mm, DV: 4.5 mm) using a Nanojet III Injector (Drummond Scientific, USA) at a rate of 1 nl per second. The injection pipette was left in place for 10 min post-injection before it was removed. After the injection, a 0.6-mm-diameter gradient index (GRIN) lens with a baseplate attached (Inscopix) was slowly lowered into the right mouse NAc (0.2 mm per minute) directly above the injection site after a slow pre-track was made with a 26-gauge blunt needle (until 0.4 mm above the target DV coordinate). Once in place, the craniotomy was closed with a low toxicity silicone adhesive (kwik-sil) and the lens was secured to the skull using dental cement (Superbond C&B kit).
Optogenetics
Each animal was unilaterally injected with 500 nl of cre-inducible AAV5-EF1a-DIO-hChR2(H134R)-eYFP, AAV5-EF1a-DIO-eNpHR-eYFP, or AAV5-EF1a-DIO-eYFP (UNC vector core) into the right NAc (AP: 1.45 mm, ML: 0.6 mm, DV: 4.5 mm) using a Nanojet III Injector (Drummond Scientific, USA) at a rate of 1 nl per second. The injection pipette was left in place for 10 min post-injection before it was removed. After the injection, a 0.2-mm-diameter optic fiber (Thorlabs) was slowly lowered into the right mouse NAc directly above the injection site (until 0.4 mm above the target DV coordinate). Once in place, the fiber was secured to the skull using dental cement (Superbond C&B kit).
At the end of the surgical procedure, mice were removed from the stereotaxic frame and postoperative care was carried out by administering analgesia (0.05mg kg-1 buprenorphine) 6 h post-procedure, as well as once every 24 h during three successive days. Animals were let to recover for 6 weeks before imaging recordings.
Behavioral experiments
Behavioral apparatus
Behavioral sessions were performed in a custom-made operant chamber using pyControl software and hardware (17.8 cm length × 19 cm width × 23 cm height) within a sound-attenuating box. For appetitive stimulus (sucrose), the chamber was composed by a central magazine, to provide access to 15 μl of sucrose solution (20% wt/vol in water) delivered by a solenoid (for liquid dispenser), a cue-sound (70 dB 5-kHz), a house-light (100 mA, 2.8 W) installed on the top and metallic floor. For the aversive stimulus (shock), the chamber contained a house-light (100 mA, 2.8 W) installed on the top of the chamber, a cue-sound (80 dB 2-kHz) and a cue-light installed in one side wall and a gridded floor with shocker. A computer was used to control the equipment and record the data and a webcam (CMOS OV2710, ELP, Shenzhen, China) was used to acquire video.
Exposure to distinct USs
Appetitive USs
Sucrose (US1)
After 3 days of habituation to the behavioral box and the miniscope, mice (nD1-cre = 15, nA2A-cre = 12) were exposed to 1 session of sucrose consumption, in which 15ul of a 20% sucrose solution were delivered every 30 seconds, for 20 minutes.
Condensed milk (US3)
After sucrose session, mice (nD1-cre = 7, nA2A-cre = 6) were exposed to 1 session of condensed milk consumption, in which 15ul of a condensed milk solution (10% sugar) were delivered every 30 s, for 30 min.
Aversive USs
Foot Shock (US2)
The same mice were familiarized to the aversive chamber apparatus for 3 days for 10 min with the patch cable connected. The foot shock session consisted of the unpredictable delivery of 7 mild foot shocks (0.5 mA, 1 s), separated by a random ITI (35–50 s).
Tail Lift (US4)
Mice were placed in an open arena covered with corn cob bedding and were allowed to explore the arena for 10 min. After that, a manual lift to the tail was applied with an interval of 30 s for 5 times.
Appetitive Pavlovian conditioning
All animals performed first the appetitive Pavlovian conditioning (protocol adapted from56) and posteriorly the aversive Pavlovian conditioning. After sucrose consumption session, mice started the appetitive Pavlovian conditioning in which a conditioned stimulus (CS1) consisting of a 70 dB 5-kHz tone and a house-light (100 mA, 2.8 W) was turned on for 10 seconds; 15 ul of 20% sucrose solution (unconditioned stimulus, US1) was made available at the 7th second after CS onset. CS-US pairings were repeated 30 times per session, with a variable inter-trial interval (ITI) of 15–35 s (randomly assigned). Mice underwent a total of 10 sessions of appetitive Pavlovian conditioning. The behavior apparatus and the sucrose receptacle were disinfected with 10% ethanol between animals to remove any odor. For all sessions, nose poke and licks (for half of the animals, as in one set the animals the lickometer was not properly working) data and imaging recordings were simultaneously obtained and synchronized through pyControl and IDAS (Inscopix) systems. To quantify CS-triggered behavior, number of nose pokes in the sucrose port were recorded during CS presentation; nose pokes and licks were also registered during the ITI period. Additionally, the area under the curve (AUC) was calculated for the ITI period and for the CS period using the Python package Scikit-learn (function sklearn.metrics.auc).
Unexpected reward omission and extinction sessions
In an additional session, animals were subjected to an unexpected reward omission session, in which reward was omitted in 25% of the trials (8/30 trials, randomly assigned). This session was followed by 3 extinction sessions, one session per day, in which CS1 was presented but no sucrose was given.
Aversive Pavlovian conditioning
After habituation to the chamber, mice started a 3-day aversive Pavlovian conditioning protocol. All sessions started with 60 s of habituation period, with the house light on. The CS2 consisted of an 80 dB, 2-kHz tone plus a cue light, paired with a mild foot shock (0.5 mA, over 1 s) (US2). During the first conditioning session, mice were exposed to 10 CS-US pairings. Each trial consisted of a random ITI (35–50 s) followed by a 10 s tone, which was immediately followed by electric foot shock delivered through the stainless-steel grid floor. All sessions were recorded with webcams. The freezing response was defined as the time (seconds) that mice spent immobile (lack of any movement including sniffing) except respiration during the CS period and calculated as percentage of total cue time ((freezing time ×100)/cue duration). To assess CS-triggered conditioned responses, two researchers evaluated freezing behavior during the CS period from all sessions in a blind manner. Since no differences between observers were detected, only data from one observer is presented in the manuscript. Researchers that performed freezing analysis were blind to group and condition.
Unexpected shock omission and extinction sessions
In an additional session, animals were subjected to an unexpected shock omission session, in which shock was omitted in 25% of the trials (4/12 trials, randomly assigned). This session was followed by 9 days of extinction, in which CS2 was presented but no shock was delivered.
Optogenetic manipulation
For all optogenetic experiments using ChR2 for optical excitation, 5 mW of blue light (at the tip of the fiberoptic) was generated by 473 nm DPSS laser (CNI Laser, Changchun, China) and unilaterally delivered to mice through fiberoptic patch cords (0.22NA, 200 μm diameter; Thorlabs, Newton, NJ, USA) that were attached to the implanted ferrule. For optogenetic experiments using eNpHR for optical inhibition, 5 mW of yellow light (at the tip of the fiberoptic) was generated by 589 nm DPSS laser (CNI Laser, Changchun, China) and unilaterally delivered as above. Laser output was controlled using a pulse generator (Master-8; AMPI, New Ulm, MN, USA) to deliver light.
Optogenetic manipulation of D1- or D2-MSNs was time-locked to cue onset on each trial and lasted the entire period of the cue (10 s). Stimulation was performed during the conditioning day (Supplementary Fig. 10) or during the nine days of extinction (Fig. 7).
Aversive Pavlovian conditioning with optogenetic manipulation
Modulation during the conditioning session
The same protocol as the one described above was performed, with optical inhibition (10 s of constant light 5 mW at the tip of the fiber) being paired with CS2.
Modulation during the extinction phase
Mice were exposed to a conditioning session like the one described above. On the extinction sessions optical manipulation (excitation: 25 ms light pulses of 20 Hz for 10 s; optical inhibition: 10 s of constant light 5 mW at the tip of the fiber) was paired with CS2 presentation, with no foot shock being delivered. Mice were exposed to 9 identical extinction sessions with optical manipulation.
Locomotor activity with optogenetic manipulation
Locomotor activity was evaluated in an open field arena (43.2 cm × 43.2 cm) with transparent acrylic walls and white floor (Med Associates Inc., St. Albans, VT, USA). Briefly, mice were attached to an optical fiber connected to a laser (473 nm or 589 nm) and immediately placed in the center of the arena. Locomotion was monitored online over a period of 10 minutes (stimulation was given similarly as in the aversive Pavlovian conditioning: 10 s of light stimulation followed by a 50 s no stimulation interval). Distance traveled during the 10-minute session was automatically detected using the Activity Monitor software (Med Associates Inc., St. Albans, VT, USA), through real-time tracking of the animal’s position by an infra-red tracking system mounted on the bottom of all walls of the arena. used as indicator of locomotor activity. Average of all stimulation (0–10 s) and post-stimulation (10–60 s) periods is presented.
Calcium imaging acquisition
GCaMP6f fluorescence signals were acquired using a miniaturized integrated fluorescence microscope system (nVoke, Inscopix, Palo Alto, CA) through GRIN lenses implanted in the NAc on freely behaving mice. Before each imaging session, the miniaturized microscope was attached to the baseplate, by gently restraining the mouse. The analog gain (3.2–6) and LED output power (0.8–1.5 mW) of the microscope were set to be constant for the same subject across imaging sessions. The microscope focus was adjusted such that the best dynamic fluorescence signals were at the focal plane, which was subsequently kept constant across imaging sessions. To synchronize behavioral events with imaging acquisition, the Data Acquisition Box of the Imaging system (Inscopix, Palo Alto, CA) was triggered by the pyControl behavioral software. Compressed gray scale images were then recorded at 20 frames per second and with spatial down sampling by a factor of 4. Timestamp of each video frame was synchronized with and recorded by the pyControl behavioral acquisition system. Calcium imaging videos were acquired during the sucrose and shock sessions, during every day of the appetitive Pavlovian conditioning for half of the animals, while the remaining animals were recorded on days 1, 2, 3, 4, 5, 6, 7 and 10. For the aversive Pavlovian conditioning, we registered all days. The number of neurons recorded in each animal is depicted in Supplementary Table 1.
Calcium imaging data processing
Data preprocessing
Using the Inscopix Data Processing Software (IDPS), we performed a field of view cropping to remove marginal areas and fixed this region for all recording session for each animal. Subsequently, using IDPS, we applied a low-pass filter for noise reduction and a first motion correction using standard IDPS settings, exporting the result as a single TIFF image stack. Next, we applied a second motion correction using the CaImAn toolbox in Python, which incorporates the NoRMCorre algorithm. This algorithm performs a fast non-rigid motion correction that collectively optimizes artifact removal caused by movements. Following these corrections, we used the extended constrained non-negative matrix factorization developed for one-photon analysis (CNMF-E) in CaImAn to perform source separation (automatic identification of regions of interest (ROIs)) and obtain denoised and deconvolved fluorescence temporal activity, termed F. This, combined with a custom Python script that estimates the noise level of temporal traces (F0) through the CaImAn internal function GetSn, yielded the normalized signals, termed ∆F/F0. Subsequently, we manually inspected all automatically detected ROIs to eliminate irregular shapes or noisy calcium activity and to confirm that the identified ROIs corresponded to cells. Examples of ROIs and normalized signals obtained from a field of view, after cropping to remove marginal areas, are shown in Fig. 1D.
Cell registration
After manual inspection, we extracted the spatial footprints of confirmed cells from CNMF-E for each session. Given a set of chronologically ordered sessions, we employed two methods for cell tracking: 1) CellReg61: a MATLAB script package that aligns spatial footprints across sessions using translation and rotation methods, with the first session as the reference map. For each cell pair, CellReg calculates a probability (P_same) of being the same cell, based on spatial correlations and centroid distance. Using default CellReg settings, a cell was considered tracked if P_same > 0.5; 2) Internal methods of the CaImAn toolbox: the register_multisession function uses spatial footprints of each cell and spatial correlation maps of each session to obtain an intersection over union metric for calculating distances between different cells in different sessions. It then solves a linear assignment problem using the Hungarian algorithm to determine the most likely cell pairings across sessions. Like CellReg, register_multisession uses the first session as the reference map. We adjusted the input parameters, setting the following variables “maximum distance considered” to 0.9 and the “max distance between centroids” to 100, providing greater flexibility compared to CellReg. We combined the results from both tracking methods and confirmed them through manual inspection.
Calcium data analysis
Alignment of activity to behavioral events
Sucrose and condensed milk
The occurrence of sucrose or condensed milk consumption events was defined as the detection of the first poke followed by a licking episode (1 licking episode was defined as having at least 2 lick events occurring less than 250 ms apart) in the behavior box within 10 s of sucrose delivery. This criterion ensured that events were accurately reflecting consumption. Since we observed a high correlation of the first poke with licking behavior, indicative of consumption, in the Pavlovian conditioning, activity data is aligned to the first poke after sucrose delivery. For each event, a 3-s window pre-event and a 3-s window post-event were analyzed.
Foot shock exposure
For foot shock event, a 3-s window pre-foot shock and a 3-s window post-foot shock were analyzed.
Tail lift
Tail lifts were similarly marked with an external TTL signal triggered manually. For each tail lift, a 3-s window pre-TTL and a 3-s window post-TTL were analyzed.
Permutation test
To classify the response of a cell to a stimulus, we analyzed changes in their average signal ∆F/F_0. We used a permutation test where the fluorescence signal was shuffled across the 6-second window (3 s before and after stimulus presentation) 1000 times. If the absolute difference between the average pre- and post-stimulus signal in a real trial was statistically less 5% than observed in the 1000 shuffled data, the neuron was considered excited when the post-stimulus signals increased and inhibited when it decreased. Neurons with non-significant differences were classified as non-responsive.
Signal normalization (z-score)
For subsequent analyses, we computed the z-score to represent the intensity of a cell’s response to a stimulus (CS or US). The z-score is calculated as z(t) = (F(t) − Favg)/FSD, where F(t) represents the normalized fluorescence signal ∆F/F0, and FAvg and FSD are the mean and standard deviation of ∆F/F0 measured across all 3-s pre-stimulus baselines in all trials, respectively. For trial-by-trial analyses, the z-score is computed using Favg and FSD measured only for the evaluated trial and in the 6 s immediately preceding the stimulus. In cases where the baseline contains only spurious activity, such as decay transients of the calcium signal or null activity, FSD is set to the standard deviation over all 6-second pre-stimulus baselines, which serves as the expected natural standard deviation.
Heatmaps
Heatmaps were constructed using the average z-score of individual cell activity using the seaborn package in Python (seaborn.heatmap). The interval was set from −3 to 3 s, where zero indicates the stimulus onset (CS or US). When heatmaps were shown for the activity of all cells recorded in each session for two stimuli, the responses were sorted in descending order based on the magnitude z-score of the first stimulus, and the same cell order was maintained for the heatmap of the second stimulus. When heatmaps were displayed for tracked cells, even for different sessions, the responses were aligned in descending order to the first stimulus of the first reference session. In all cases, the color bar was configured with ‘extend’ in seaborn.heatmap, indicating that the color scale extends beyond the upper and lower bounds of the data. The color scale was set to have an upper bound of 1.5 and a lower bound of −0.5 for the z-score.
PSTHs and AUCs
After classifying the cells responses to a stimulus, we constructed peristimulus time histograms (PSTHs) for each group of response types (excited, inhibited, and non-responsive). For each group, individual z-scores were averaged, and the mean with standard error of the mean (SEM) was plotted. Additionally, the AUC was calculated for each group during the 3 s following stimulus onset using the Python package Scikit-learn (function sklearn.metrics.auc). These methods were also applied to obtain PSTHs and AUCs for each cluster of cells based on their responses on day 1 of appetitive and aversive conditioning.
Percentage of persisted responses
After classifying a cell’s response to a stimulus as excited, inhibited, or non-responsive based on the mean across trials in the permutation test, we calculated the percentage of persisted responses. This involved determining the fraction of trials in which the same response was obtained when the permutation test was applied to each single trial. We defined that >=70% of persisted responses indicates that the cell’s response to the stimulus is consistent (Supplementary Fig. 2E, F).
Shannon’s entropy
To assess the variability of neuronal responses to each stimulus, we used Shannon’s entropy. For each presentation of the same stimulus, the resulting neuronal responses formed a series, which was subsequently utilized to calculate Shannon’s entropy using the expression -Σip(i)log2[p(i)], where p(i) denotes the probability of observing a specific type of response (e.g., excitatory, inhibitory, or non-responsive). In the most uncertain (random) scenario, where the neuron could produce any response for each stimulus presentation, p(i) = 1/3, resulting in an approximate maximum entropy of 1.58. Conversely, in the deterministic scenario, where the neuron consistently responds in the same manner for every stimulus presentation, the entropy is zero.
Single cell decoder analysis
We used the average z-score of the cell’s 3-second response to each stimulus presentation for single-cell decoding analyses. Given a cell with MA trials responding to stimulus A and MB trials responding to stimulus B, we employed balanced datasets for training a linear SVM model (using sklearn.svm.LinearSVC in Python with C = 0.8). If MA is greater than MB, we randomly subsampled MB trials from MA. Otherwise, we randomly subsampled MA trials from MB. From the actual data, we used 70% of the trials for each stimulus for training and reserved the remaining 30% for machine accuracy testing. To generate the corresponding shuffled data, we conducted 100 permutations of the ∆F/F0 signal for each window from −6 to 3 s relative to stimulus onset. We then proceeded with the z-score calculation for trial-by-trial as described previously. This procedure was mirrored to create a classifier using the actual data to decode the shuffled data. Finally, we generated 1000 independent machines for both the actual test data and the shuffled data. The average of the accuracy test results on these machines represents the single cell decoder accuracy. In Fig. 3F, G, the decoding accuracy of each neuron (neuron ID on X axis) on day 1 was plotted following the color code of the accuracy (red - high, blue – low), and gray dots represent the decoding accuracy of those same neurons on day 5 (left) or day 10 (right). For Fig. 3H, I, neurons were divided into 20% best decoders, or 20% worst decoders based on day 1 activity.
Population decoder analysis
For population-level decoding analyses, we use a population vector of size N, where N is the total number of cells responding to different stimuli. Each element in this vector represents the average z-score of a cell’s 3-second response to a stimulus presentation. Each trial generates a unique population vector that serves as a representation of the presented stimulus. A stimulus presentation trial generates a population vector that represents that stimulus. Like the single-cell analysis, we ensured balanced datasets for the SVM model by selecting an equal number of population vectors from each stimulus category. As before, we selected an equal number of population vectors from each stimulus to generate datasets for the SVM model. This way, we allocated 70% of the population vectors for model training and the remaining 30% for machine accuracy testing. We generated 1000 independent machines for both the actual test data and the shuffled data. The average of the accuracy test results on these machines represents the population decoder accuracy.
Correlation analysis
To assess individual variability in response dynamics to the same stimulus within a single session, we constructed an M x M matrix for each neuron, where M is the number of trials for the same stimulus. Each element Mij of the matrix was computed as the Pearson correlation coefficient between the 3-second z-scored responses to the stimulus for trial i and trial j. The single neuron activity correlation was then derived as the mean of the elements located above the main diagonal of matrix M, representing the average Pearson correlation coefficient across all combinations of trial pairs. This is exemplified in Fig. 1L, M, which shows the distribution of this correlation for neurons for US1 and US2.
In addition to single neuron activity correlation, we also assessed the tuning curve correlation, which quantifies the similarity in response profiles of individual neurons to the same stimulus across multiple presentations within a session71. For each trial i, we constructed an N x T matrix of z-scores, where N is the number of neurons recorded from an animal and T is the number of time frames in the analysis period (3 s post-stimulus, acquired at 20 Hz, resulting in T = 60). The tuning curve correlation between trials i and j was then calculated as the median of the Pearson correlation coefficients between corresponding neurons across trials. Supplementary Fig. 2I illustrates how the tuning curve correlation changes on average as we compare trials with increasing separation (“distance between trials”). For this figure, especially for sucrose consumption (US1), we only showed 13 consumptions per animal. This is because consumption varied between animals, and 13 represented the least common number of consumptions observed.
To evaluate the correlation in individual response dynamics to the same stimulus and across different sessions, for each tracked neuron, we calculated the Pearson correlation coefficient between the average signal of z-scored responses (3-second window) to the stimulus across distinct sessions. This assesses how consistently individual neurons respond to the same stimulus over time. For instance, in Supplementary Fig. 4I, J, the distributions of these correlations of all tracked neurons for different pairs of days during appetitive conditioning are shown, with the averages of the distributions of each pair being highlighted by vertical lines.
Furthermore, we examined correlation between the CS-US stimuli at the population level. For each neuron, we averaged its responses across all trials and within a 3-s window for each stimulus. For example, this data is visualized in Supplementary Fig. 3D, E, where each data point represents an average response pair CS-US. To quantify the association between these paired responses, we calculated the Pearson correlation coefficient and performed linear regression to identify possible trends in the data. We also measured the angle between the closest points of each response pair. This metric, in the form of a histogram, allows from another perspective to compare the evolution of these data when analyzing different sessions. These analyses were not only performed on all neurons but also on a subset of “CS-US responsive” neurons identified through a permutation test. This approach allowed us to concentrate on neurons that showed significant activity changes in response to both stimuli.
We also evaluated the population vector correlation (PV correlation), which is a measure of similarity between multiple responses to the same stimulus across a neuronal population. This measure has been applied in detecting drift in overall activity patterns over time, comparing “within-session” (single session) and “between-sessions” (different sessions) correlations (for more details, see ref. 71). Briefly, for each stimulus (CS or US) and tracked neurons, we split each session into two blocks (first and second half of trials). For each neuron, we then averaged the z-scores across all frames within each block. In these blocks, a population vector is a set of average z-score values at a given frame, thus, a block is formed by an N × T matrix, where N is the number of neurons and with T = 60 frames (3 s). We then calculate the PV correlation between pairs of blocks by averaging the Pearson correlation coefficients between their respective population vectors at each frame. When the two blocks are from the same session, the values of PV correlation are added to the “within-session” group, and when the blocks are from different sessions, the values of PV correlation are added to the “between-sessions” group. Finally, Fig. 3M, N displays the average values of these groups across animals for D1- and A2A-neurons.
Cluster analysis and representation
For cluster analysis, we concatenated the z-scores of the average activity over all trials from −3 to 3 s (where zero corresponds to stimulus onset) of both the CS and its corresponding US, totaling 12 s. Treating each timepoint (frame) as an independent dimension, we applied Principal Component Analysis (PCA, sklearn.decomposition.PCA function in Python) to a matrix of dimensions N x D, where N is the number of cells and D is the total number of dimensions. In our case, this resulted in 240 dimensions (12 s × 20 Hz, the sampling rate). Subsequently, we performed dimensionality reduction, selecting enough Principal Components (PCs) to account for at least 80% of the observed variance in our data (Supplementary Fig. 6A). This typically ranged from 12 to 15 PCs, which were then subjected to the K-means clustering method using the sklearn.cluster.KMeans function in Python. To determine the optimal number of clusters (nc) for the reduced data, we varied nc from 2 to 10 and performed the following assessments: 1) Identified the nc that marked the onset of stability in the average cosine similarity computed between cluster elements and the average cluster activity (Supplementary Fig. 6B); 2) Identified the nc that yielded a peak in silhouette score analysis using the Euclidean metric (Supplementary Fig. 6C). The concordance of these two measures defined the nc to be employed in subsequent analyses. As an illustrative example, we examined the average z-scores of D1 neurons recorded on day 1 of appetitive conditioning, and this analysis revealed that nc = 3 was the optimal number of clusters for this dataset. To visualize the identified clusters, in Supplementary Fig. 6D, we utilized two dimensionality reduction techniques: PCA (a linear method) and t-Distributed Stochastic Neighbor Embedding (t-SNE, a non-linear method), and for both, we use the first two components, and each color represents a cluster.
Neuronal trajectory
To investigate the evolution of neural activity patterns following positive and negative valence stimuli, we conducted neural trajectory analysis. This approach tracks changes in a tracked population response over time. For each tracked cell, we concatenated z-score of average activity across all trials within a 3-second window after stimulus onset. To analyze US1 and US2 in Fig. 1R, S, we generated a matrix of size N x 120 (2 stimuli, each comprising 60 frames). Similarly, to analyze CS1, CS2, US1, and US2 in Fig. 6G–I, we generated a matrix of size N x 240. Each matrix was generated per animal, where N represents the number of cells. To visualize the overall activity dynamics, we reduced the matrix dimensionality to two principal components (PC) using PCA treating each cell as an independent dimension. We then smoothed these components using one-dimensional Gaussian convolution, resulting in neural trajectories shown as solid lines in these figures.
To quantify the dissimilarity between trajectories over time for different stimuli, we calculated the Euclidean distance between their corresponding PC values at each time point. Specifically, we computed distances between (PC1_EST1, PC2_EST1) and (PC1_EST2, PC2_EST2) at each time point, where EST corresponds to CS or US.
Furthermore, we visualized single-trial variability by plotting individual trial trajectories as dashed lines (Fig. 1R, S and Fig. 6G–I). These trajectories were obtained by applying the same PCA transformation matrix used for average activity data. Notably, we only plotted 7 trials per stimulus (the maximum number of trials for footshock) to maintain a balanced representation, similar to the population decoder analysis.
Sacrifice and brain sectioning for histological analysis
At the end of all behavior procedures, mice were deeply anesthetized by a mixture of ketamine/medetomidine. Animals were then transcardially perfused with 0.9% saline, followed by 4% paraformaldehyde (PFA) solution. After, whole heads with the lenses or the optic fibers attached were immersed for 48 h in 4% PFA so that the lens track is clearly visible for histological analysis. Next, brains were extracted and then rinsed and stored in 30% sucrose at 4 °C until sectioning.
Sectioning was performed coronally, in 40 μm slices, on a vibrating microtome (VT1000S, Leica, Germany) and slices were stored at 4 °C on 12-well plates (or long-term storage in cryoprotectant solution at −20 °C) until use. Slices from the area of interest (NAc) were selected using the Mouse Brain Atlas62.
Immunohistochemistry
To assess GCaMPf6 expression in D1-cre and A2A-cre mice, brain slices containing NAc sections were washed with phosphate buffered saline (PBS), and then permeabilizated with PBS-Triton 0.3% (PBS-T 0.3%). Blocking was performed for 1 h using 5% Fetal Bovine Serum (FBS; Invitrogen, MA, USA) in PBS-T at RT. The primary antibody goat anti-GFP (1:750; ab6673, Lot 1033180-23, ABCAM, Cambridge, UK), was incubated overnight at 4 °C with agitation, followed by PBS-T washes and subsequent incubation with the secondary fluorescent antibody Alexa Fluor® 488 donkey anti-goat (1:500; A11055, Lot 2747580, Invitrogen, Carlsbad, CA, USA). All antibodies were diluted in PBS-T with 2% FBS. Slices were washed with PBS-T, incubated with 4’,6-Diamidino-2-Phenylindole Dihydrochloride (DAPI, 1:1000; 62248, Thermo ScientificTM, Waltham, MA, USA) for nucleus staining, washed with PBS and mounted using Permafluor (Invitrogen, MA, USA). Slides were stored at 4 °C and kept protected from light.
Image acquisition and analysis
Images from the NAc of D1-cre and A2A-cre mice were collected in an inverted confocal microscope (Olympus FV3000, Tokyo, Japan). About 6 slices per animals were used for each analysis. Optic fiber placement was assessed to confirm if the activity detected was from the NAc region. For that, slices where the optic fiber was detected were classified according to the Mouse brain atlas62 to estimate the stereotaxic coordinates.
Statistical analysis
Part of the statistical analysis was performed in GraphPad Prism 9.0 (GraphPad Software, Inc., La Jolla, CA, USA). Prior to any statistical comparison between groups, normality was assessed in all data analyzed by using the Kolmogorov–Smirnov (KS) test. Parametric tests were used whenever KS test >0.05. If normality assumptions were not met, non-parametric analysis (Mann–Whitney or Wilcoxon test) was performed.
For behavior, two-way analysis of variance (ANOVA) for repeated measures was used to assess learning in D1- and A2A-cre mice (factors used: nose pokes during CS versus nose pokes during ITI, across days of training); and to analyze percentage (%) of freezing throughout the trials on day 1. Bonferroni’s post hoc multiple comparison test was used for group differences determination. One Way ANOVA was done to compare the poke probability at the ITI or CS period on days 1, 5 and 10 of learning in the appetitive Pavlovian conditioning task. One Way ANOVA was also done to compare the latency to compare the time between delivery of the reward and the first poke on days 1, 5 and 10 of training. Statistical analysis between two time-points was made using two-tailed paired Student’s t test, to compare percentage of freezing in the first and last trials of the aversive conditioning. Two-way ANOVA for repeated measures was performed to compare the percentage of freezing across days of shock extinction.
Data are presented as mean ± standard error of mean (SEM). Statistical significance was considered for p ≤ 0.05. All statistical data of the main figures are depicted in Supplementary Table 2. All statistical data of the Supplementary figures are depicted in Supplementary Table 3.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Source Data are provided with this paper. All data generated in this study have been deposited in the Zenodo database under identifier https://doi.org/10.5281/zenodo.13890862. Source data are provided with this paper.
Code availability
This paper reports original code that is available on the GitHub platform github.com/rewave-lab and published the identifier https://doi.org/10.5281/zenodo.13890127.
References
Wheeler, R. A., Roitman, M., Grigson, P. & Carelli, R. M. Single Neurons in the Nucleus Accumbens Track Relative Reward. Int. J. Comp. Psychol. 18, 320–332 (2005).
Roitman, M. F., Wheeler, R. A. & Carelli, R. M. Nucleus Accumbens Neurons Are Innately Tuned for Rewarding and Aversive Taste Stimuli, Encode Their Predictors, and Are Linked to Motor Output. Neuron 45, 587–597 (2005).
Ottenheimer, D., Richard, J. M. & Janak, P. H. Ventral pallidum encodes relative reward value earlier and more robustly than nucleus accumbens. Nat. Commun. 9, 4350 (2018).
Day, J. J., Wheeler, R. A., Roitman, M. F. & Carelli, R. M. Nucleus accumbens neurons encode Pavlovian approach behaviors: evidence from an autoshaping paradigm. Eur. J. Neurosci. 23, 1341–1351 (2006).
Eyny, Y. S. & Horvitz, J. C. Opposing Roles of D 1 and D 2 Receptors in Appetitive Conditioning. J. Neurosci. 23, 1584–1587 (2003).
Perreault, M. L., Hasbi, A., O’Dowd, B. F. & George, S. R. The Dopamine D1–D2 Receptor Heteromer in Striatal Medium Spiny Neurons: Evidence for a Third Distinct Neuronal Pathway in Basal Ganglia. Front. Neuroanat. 5, 31 (2011).
Kravitz, A. V., Tye, L. D. & Kreitzer, A. C. Distinct roles for direct and indirect pathway striatal neurons in reinforcement. Nat. Neurosci. 15, 816–818 (2012).
Lobo, M. K. et al. Cell Type–Specific Loss of BDNF Signaling Mimics Optogenetic Control of Cocaine Reward. Science 330, 385–390 (2010).
Hikida, T., Kimura, K., Wada, N., Funabiki, K. & Nakanishi, S. Distinct roles of synaptic transmission in direct and indirect striatal pathways to reward and aversive behavior. Neuron 66, 896–907 (2010).
Soares-Cunha, C. et al. Nucleus accumbens medium spiny neurons subtypes signal both reward and aversion. Mol. Psychiatry 25, 3241–3255 (2020).
Soares-Cunha, C. et al. Activation of D2 dopamine receptor-expressing neurons in the nucleus accumbens increases motivation. Nat. Commun. 7, 11829 (2016).
Cole, S. L., Robinson, M. J. F. & Berridge, K. C. Optogenetic self-stimulation in the nucleus accumbens: D1 reward versus D2 ambivalence. PLOS ONE 13, e0207694 (2018).
Natsubori, A. et al. Ventrolateral Striatal Medium Spiny Neurons Positively Regulate Food-Incentive, Goal-Directed Behavior Independently of D1 and D2 Selectivity. J. Neurosci. J. Soc. Neurosci. 37, 2723–2733 (2017).
Cui, G. et al. Concurrent activation of striatal direct and indirect pathways during action initiation. Nature 494, 238–242 (2013).
Lafferty, C. K., Yang, A. K., Mendoza, J. A. & Britt, J. P. Nucleus Accumbens Cell Type- and Input-Specific Suppression of Unproductive Reward Seeking. Cell Rep. 30, 3729–3742.e3 (2020).
Thoeni, S., Loureiro, M., O’Connor, E. C. & Lüscher, C. Depression of Accumbal to Lateral Hypothalamic Synapses Gates Overeating. Neuron 107, 158–172.e4 (2020).
O’Connor, E. C. et al. Accumbal D1R Neurons Projecting to Lateral Hypothalamus Authorize Feeding. Neuron 88, 553–564 (2015).
Soares-Cunha, C. et al. Nucleus Accumbens Microcircuit Underlying D2-MSN-Driven Increase in Motivation. eNeuro 5, ENEURO.0386-18.2018 (2018).
Soares-Cunha, C. et al. Distinct role of nucleus accumbens D2-MSN projections to ventral pallidum in different phases of motivated behavior. Cell Rep. 38, 110380 (2022).
Humphries, M. D. & Prescott, T. J. The ventral basal ganglia, a selection mechanism at the crossroads of space, strategy, and reward. Prog. Neurobiol. 90, 385–417 (2010).
Pedersen, C. E. et al. Medial Accumbens Shell Spiny Projection Neurons Encode Relative Reward Preference. http://biorxiv.org/lookup/doi/10.1101/2022.09.18.508426 (2022).
Coss, A., Suaste, E. & Gutierrez, R. Lateral NAc Shell D1 and D2 Neuronal Ensembles Concurrently Predict Licking Behavior and Categorize Sucrose Concentrations in a Context-dependent Manner. Neuroscience 493, 81–98 (2022).
Chen, G. et al. Distinct reward processing by subregions of the nucleus accumbens. Cell Rep. 42, 112069 (2023).
Zachry, J. E. et al. D1 and D2 medium spiny neurons in the nucleus accumbens core have distinct and valence-independent roles in learning. Neuron S0896627323009261, https://doi.org/10.1016/j.neuron.2023.11.023 (2023).
Soares-Cunha, C., Coimbra, B., Sousa, N. & Rodrigues, A. J. Reappraising striatal D1- and D2-neurons in reward and aversion. Neurosci. Biobehav. Rev. 68, 370–386 (2016).
Tye, K. M. Neural Circuit Motifs in Valence Processing. Neuron 100, 436–452 (2018).
Lin, S.-C. & Nicolelis, M. A. L. Neuronal Ensemble Bursting in the Basal Forebrain Encodes Salience Irrespective of Valence. Neuron 59, 138–149 (2008).
Zhang, X. & Li, B. Population coding of valence in the basolateral amygdala. Nat. Commun. 9, 5195 (2018).
Yang, T. et al. Plastic and stimulus-specific coding of salient events in the central amygdala. Nature 616, 510–519 (2023).
Nishioka, T. et al. Error-related signaling in nucleus accumbens D2 receptor-expressing neurons guides inhibition-based choice behavior in mice. Nat. Commun. 14, 2284 (2023).
Rescorla, R. A. & Wagner, A. R. A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. In Classical Conditioning II: Current Research and Theory 64–99 (Appleton-Century Crofts, 1972).
Schultz, W. Dopamine reward prediction-error signalling: a two-component response. Nat. Rev. Neurosci. 17, 183–195 (2016).
Zhu, Y. et al. Dynamic salience processing in paraventricular thalamus gates associative learning. Science 362, 423–429 (2018).
Bouton, M. E., Westbrook, R. F., Corcoran, K. A. & Maren, S. Contextual and Temporal Modulation of Extinction: Behavioral and Biological Mechanisms. Biol. Psychiatry 60, 352–360 (2006).
Kutlu, M. G. et al. Dopamine release in the nucleus accumbens core signals perceived saliency. Curr. Biol. 31, 4748–4761.e8 (2021).
Bobadilla, A.-C. et al. Cocaine and sucrose rewards recruit different seeking ensembles in the nucleus accumbens core. Mol. Psychiatry 25, 3150–3163 (2020).
Driscoll, L. N., Pettit, N. L., Minderer, M., Chettih, S. N. & Harvey, C. D. Dynamic Reorganization of Neuronal Activity Patterns in Parietal Cortex. Cell 170, 986–999.e16 (2017).
Schoonover, C. E., Ohashi, S. N., Axel, R. & Fink, A. J. P. Representational drift in primary olfactory cortex. Nature 594, 541–546 (2021).
Rule, M. E., O’Leary, T. & Harvey, C. D. Causes and consequences of representational drift. Curr. Opin. Neurobiol. 58, 141–147 (2019).
Stuber, G. D. et al. Excitatory transmission from the amygdala to nucleus accumbens facilitates reward seeking. Nature 475, 377–380 (2011).
Ma, L., Chen, W., Yu, D. & Han, Y. Brain-Wide Mapping of Afferent Inputs to Accumbens Nucleus Core Subdomains and Accumbens Nucleus Subnuclei. Front. Syst. Neurosci. 14, 15 (2020).
Ito, R., Robbins, T. W., Pennartz, C. M. & Everitt, B. J. Functional Interaction between the Hippocampus and Nucleus Accumbens Shell Is Necessary for the Acquisition of Appetitive Spatial Context Conditioning. J. Neurosci. 28, 6950–6959 (2008).
Sjulson, L., Peyrache, A., Cumpelik, A., Cassataro, D. & Buzsáki, G. Cocaine Place Conditioning Strengthens Location-Specific Hippocampal Coupling to the Nucleus Accumbens. Neuron 98, 926–934.e5 (2018).
Kupchik, Y. M. et al. Coding the direct/indirect pathways by D1 and D2 receptors is not valid for accumbens projections. Nat. Neurosci. 18, 1230–1232 (2015).
Ottenheimer, D. J. et al. A quantitative reward prediction error signal in the ventral pallidum. Nat. Neurosci. 23, 1267–1276 (2020).
Stephenson-Jones, M. et al. Opposing Contributions of GABAergic and Glutamatergic Ventral Pallidal Neurons to Motivational Behaviors. Neuron 105, 921–933.e5 (2020).
Parkinson, J. A. et al. Nucleus accumbens dopamine depletion impairs both acquisition and performance of appetitive Pavlovian approach behaviour: implications for mesoaccumbens dopamine function. Behav. Brain Res. 137, 149–163 (2002).
Di Ciano, P., Cardinal, R. N., Cowell, R. A., Little, S. J. & Everitt, B. J. Differential Involvement of NMDA, AMPA/Kainate, and Dopamine Receptors in the Nucleus Accumbens Core in the Acquisition and Performance of Pavlovian Approach Behavior. J. Neurosci. 21, 9471–9477 (2001).
Dalley, J. W. et al. Time-limited modulation of appetitive Pavlovian memory by D1 and NMDA receptors in the nucleus accumbens. Proc. Natl Acad. Sci. 102, 6189–6194 (2005).
Parkinson, J. A., Olmstead, M. C., Burns, L. H., Robbins, T. W. & Everitt, B. J. Dissociation in Effects of Lesions of the Nucleus Accumbens Core and Shell on Appetitive Pavlovian Approach Behavior and the Potentiation of Conditioned Reinforcement and Locomotor Activity byd-Amphetamine. J. Neurosci. 19, 2401–2411 (1999).
Deseyve, C. et al. Nucleus accumbens neurons dynamically respond to appetitive and aversive associative learning. J. Neurochem. 168, 312–327 (2024).
Ray, M. H., Moaddab, M. & McDannald, M. A. Threat and Bidirectional Valence Signaling in the Nucleus Accumbens Core. J. Neurosci. 42, 817–833 (2022).
Morrison, S. E., McGinty, V. B., du Hoffmann, J. & Nicola, S. M. Limbic-motor integration by neural excitations and inhibitions in the nucleus accumbens. J. Neurophysiol. 118, 2549–2567 (2017).
Saddoris, M. P., Cacciapaglia, F., Wightman, R. M. & Carelli, R. M. Differential Dopamine Release Dynamics in the Nucleus Accumbens Core and Shell Reveal Complementary Signals for Error Prediction and Incentive Motivation. J. Neurosci. 35, 11572–11582 (2015).
Pearce, J. M. & Hall, G. A model for Pavlovian learning: Variations in the effectiveness of conditioned but not of unconditioned stimuli. Psychol. Rev. 87, 532–552 (1980).
Ambroggi, F., Ghazizadeh, A., Nicola, S. M. & Fields, H. L. Roles of Nucleus Accumbens Core and Shell in Incentive-Cue Responding and Behavioral Inhibition. J. Neurosci. 31, 6820–6830 (2011).
West, E. A. & Carelli, R. M. Nucleus Accumbens Core and Shell Differentially Encode Reward-Associated Cues after Reinforcer Devaluation. J. Neurosci. 36, 1128–1139 (2016).
Floresco, S. B., Montes, D. R., Tse, M. M. T. & Van Holstein, M. Differential Contributions of Nucleus Accumbens Subregions to Cue-Guided Risk/Reward Decision Making and Implementation of Conditional Rules. J. Neurosci. 38, 1901–1914 (2018).
Van Elzelingen, W. et al. A unidirectional but not uniform striatal landscape of dopamine signaling for motivational stimuli. Proc. Natl Acad. Sci. 119, e2117270119 (2022).
Engel, L. et al. Dopamine neurons drive spatiotemporally heterogeneous striatal dopamine signals during learning. Curr. Biol. 34, 3086–3101.e4 (2024).
Saunders, B. T., Richard, J. M., Margolis, E. B. & Janak, P. H. Dopamine neurons create Pavlovian conditioned stimuli with circuit-defined motivational properties. Nat. Neurosci. 21, 1072–1083 (2018).
Salinas-Hernández, X. I., Zafiri, D., Sigurdsson, T. & Duvarci, S. Functional architecture of dopamine neurons driving fear extinction learning. Neuron S0896627323006360, https://doi.org/10.1016/j.neuron.2023.08.025 (2023).
Belilos, A. et al. Nucleus accumbens local circuit for cue-dependent aversive learning. Cell Rep. 42, 113488 (2023).
Holtzman-Assif, O., Laurent, V. & Westbrook, R. F. Blockade of dopamine activity in the nucleus accumbens impairs learning extinction of conditioned fear. Learn. Mem. 17, 71–75 (2010).
Lacagnina, A. F. et al. Distinct hippocampal engrams control extinction and relapse of fear memory. Nat. Neurosci. 22, 753–761 (2019).
Luo, R. et al. A dopaminergic switch for fear to safety transitions. Nat. Commun. 9, 2483 (2018).
Iino, Y. et al. Dopamine D2 receptors in discrimination learning and spine enlargement. Nature 579, 555–560 (2020).
Correia, R. et al. Involvement of nucleus accumbens D2–medium spiny neurons projecting to the ventral pallidum in anxiety-like behaviour. J. Psychiatry Neurosci. 48, E267–E284 (2023).
Francis, T. C. & Lobo, M. K. Emerging Role for Nucleus Accumbens Medium Spiny Neuron Subtypes in Depression. Biol. Psychiatry 81, 645–653 (2017).
Lüscher, C. The Emergence of a Circuit Model for Addiction. Annu. Rev. Neurosci. 39, 257–276 (2016).
Deitch, D., Rubin, A. & Ziv, Y. Representational drift in the mouse visual cortex. Curr. Biol. 31, 4327–4339.e6 (2021).
Acknowledgements
This work was funded by the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement No 101003187) and by the “la Caixa” Foundation (ID 100010434), under the agreement LCF/PR/HR20/52400020. The work was also funded by a Bial Foundation grant (175/2020). Part of the work received funding from the FCT under the scope of the projects PTDC/MED-NEU/4804/2020 (https://doi.org/10.54499/PTDC/MED-NEU/4804/2020), PTDC/SAU-TOX/6802/2020 (https://doi.org/10.54499/PTDC/SAU-TOX/6802/2020), 2022.02201.PTDC (https://doi.org/10.54499/2022.02201.PTDC) and 2022.01467.PTDC (https://doi.org/10.54499/2022.01467.PTDC). CS-C, BC and LP have Scientific Employment Stimulus contracts from the Portuguese Foundation for Science and Technology (FCT) (CEECIND/03887/2017 (https://doi.org/10.54499/CEECIND/03887/2017/CP1458/CT0027) and 2023.08896.CEECIND; CEECIND/03898/2020 (https://doi.org/10.54499/2020.03898.CEECIND/CP1600/CT0015); CEECINST/00077/2018 (https://doi.org/10.54499/CEECINST/00077/2018/CP1640/CT0003)). AVD and RC have FCT PhD grants (SFRH/BD/147066/2019; 2022.12973.BD). This work was also supported by a FEBS (Federation of European Biochemical Societies) Excellence Award, IBRO Early Career Award, “Maria de Sousa” Award and Career Development Grant (CDG, International Society for Neurochemistry) attributed to Carina Soares-Cunha. Host laboratory is funded by National funds, through FCT - project UIDB/50026/2020 (https://doi.org/10.54499/UIDB/50026/2020), UIDP/50026/2020 (https://doi.org/10.54499/UIDP/50026/2020) and LA/P/0050/2020 (https://doi.org/10.54499/LA/P/0050/2020).
Author information
Authors and Affiliations
Contributions
C.S.-C. and A.J.R. conceived the project, designed the experiments and supervised the research; A.V.D. performed the majority of the experiments; T.T.A.C. analyzed most of the data; G.J.M. supported in the collection of 1-photon images; R.C. performed histological confirmation; B.C. supported in optogenetic experiments; R.B.-G. performed analysis of aversive Pavlovian conditioning data; R.G. was responsible for colony management and generation of transgenic mice; M.W. helped in the optogenetic experiment data analysis; L.P. provided support to imaging data collection and analysis; R.M.C. and N.S. supervised the research.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Fabien Ducrocq, Tom Macpherson, Kenji Tanaka, and Pierre Trifilieff for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Domingues, A.V., Carvalho, T.T.A., Martins, G.J. et al. Dynamic representation of appetitive and aversive stimuli in nucleus accumbens shell D1- and D2-medium spiny neurons. Nat Commun 16, 59 (2025). https://doi.org/10.1038/s41467-024-55269-9
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-024-55269-9
This article is cited by
-
Motivation meets sleep: tuning arousal via nucleus accumbens and Basal Ganglia circuits
npj Biological Timing and Sleep (2025)
