
Abstract
High-mountain lakes were historically fishless due to natural barriers, but human introductions have led to widespread fish presence. Although particularly intensive during the last decades, historical documents indicate introductions in European high mountains already during the 14th and 15th centuries, but they could have occurred before, provided the intensive land use of the high mountain had started earlier. We used ancient environmental DNA from lake sediments (sedDNA) to investigate this hypothesis. Fish ectoparasites from various clades were identified using the 18S rRNA gene in the sediment record of a deep, high-mountain Pyrenean lake, with Ichthyobodo (Kinetoplastea) being of particular interest due to its consistent occurrence. The study shows a continued presence of fish parasites in the lake since the 7th century, which coincides with the Late-Roman and Visigothic extensive mountain use for sheep pasturing as supported by nearby archeological remains and increased lake primary production evidenced by photosynthetic pigments.
Similar content being viewed by others
Introduction
Most high-elevation mountain lakes are naturally fishless worldwide because of the glacial steep relief that isolated the cirque lakes from colonization. Unfortunately, many have been stocked with downstream or non-native fish species1. Sport fishing and helicopter availability have widely spread lake fish during the last decades, becoming a conservation concern2,3. Historical documents indicate that fish stocking started much earlier, at least in the European ranges, during the 14th and 15th centuries4. Most existing documents were related to the rights to use specific lakes for fishing and trade, showing an advanced socio-economical use of those high-mountain lands. There were also philanthropic initiatives to recover otherwise “lifeless waters,” as the stated motivation of Emperor Maximillian I in the Tyrolian Alps during the 15th century5. It has been suspected, however, that fish stocking could have started much earlier4 than documents indicate, likely during the intensive occupation of the mountains for pasturing and foresting in the first millennium of the current era.
Without documentary records, lake sediments could be used to assess the historical introduction of fish. Sparse fish remains are unlikely to be sampled in a sediment core, and obtaining a continuous, reliable record of them is nearly impossible. The traditional approach has been to investigate the lake community shift that could have caused the occurrence of a previously non-existing top predator1,6. Amphibians and some large macroinvertebrates decline or even become extinct in mountain lakes with fish7. Unfortunately, the record of these most impacted organisms is also poor in the sediments, in contrast to cladoceran remains, which are abundant. As fish are visual predators, they prey on cladocerans selectively according to their size, which differs markedly among species; hence, changes in the community composition have been used to assess fish introductions in lakes8, with particular success in shallow, productive lakes studies9,10,11. However, the possibility of applying the method in large, oligotrophic mountain lakes has raised doubts12 because large cladocerans and trout coexist in many of these lakes. Furthermore, cladoceran population dynamics at centuries scales are influenced by land use and climate shifts13,14, whose effects could be confounded with fish introductions. Indirect fish impacts through top-down cascading to primary producers using other paleolimnological indicators (e.g., diatoms, pigments)11 could be even more challenging to distinguish from the effects of other drivers. Indeed, determining historical fish introductions in lakes through their food web impact is somewhat of a circular rationale because the cause is inferred by some presumed effects that should be, in fact, the next object of study. Ideally, a method providing direct evidence of fish occurrence could facilitate a more coherent evaluation of the causes and consequences15.
Environmental sediment DNA (sedDNA) increases the possibility of finding a signature of the fish introduction independent of the impact16. Recovering fish DNA from the sediments would be the most direct way, but it is technically challenging, and recovery is not always successful, even in lakes where fish is relatively abundant17. A complementary and overlooked possibility of detecting fish presence is the molecular signature of their parasites, which have been previously found in sedDNA studies18. Natural fish populations may host a wide variety of parasites, some achieving high densities without compromising the viability of the fish population19. Fish offer a large surface area for parasite-host encounter and colonization, facilitating infection, and are highly mobile, helping parasite dispersal. Microscopic ectoparasites may have large populations and distribute more homogenously across the water column during some life phases than their host. Because of direct sinking or scavenging by larger particles, there can be a small but sustained flux of parasites toward the sediment. In contrast, fish carcasses are occasional and locally distributed. Therefore, looking for direct fish evidence and of their parasites becomes a complementary strategy for assessing fish’s early introduction in mountain lakes using sedimentary DNA.
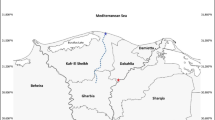
We used sedDNA to determine when fish were introduced in a large, 73 m deep lake of the Pyrenees (Fig. 1). This lake is isolated from the fluvial network by a steep waterfall over 100 m high, and thus, fish could not have naturally colonized the lake in the past. However, currently, the lake supports a large brown trout population ( ~ 60,000 individuals)20. The only documented evidence of fish occurrence in the past is a royal bureaucracy questionnaire from the 18th century, stating that the lake was hard for fishing but had excellent trout21. It is unlikely that once a large fish population was established in this lake, natural or human causes could extinguish it. Therefore, we can assume that trout have existed in the lake for over two hundred years and ask how much before it was first introduced. Because of its size and location, Lake Redon is an excellent site for testing whether fish introduction could have occurred in parallel with the early socio-economic development of the mountain area.

A Location of Lake Redon in the Central Pyrenees. B Lake Redon view facing northwestward. C Orthoimage of the catchment and lake bathymetry indicating the coring point. Source orthoimages in (A and B): Institut Cartogràfic i Geològic de Catalunya 2021. Ortofoto 25 cm (OF-25C) Mapa 1:2500 (CC BY 4.0).
Archeological evidence22 and an entire Holocene sediment record previously studied in the lake indicate that low-intensity pasturing in the catchment and mining in nearby areas occurred since the late Roman period23,24. Furthermore, the Central Southern Pyrenees experienced economic and cultural splendor during the 12th and 13th centuries, as evidenced by the flourishment of Romanesque art in the valleys’ towns25. Across these centuries, land use in the area achieved an exceptional intensity above any period after the Middle Ages. Therefore, it could be expected that fish introduction in the lake occurred at any time during these centuries, and since then, the lake community could have been conditioned by the fish’s presence.
We tested a collection of DNA primers to detect fish using top sediment samples, including some targeting vertebrates in general and others specific for trout (see methods). Despite the presence of the current fish population, all tests were unsuccessful. Therefore, we used the V9 region of the 18S rRNA gene across the sediment record to target a broad spectrum of eukaryotes, including fish parasites. The nucleotide sequence of this hypervariable region ( ~ 168 bp) is shorter than alternative regions (e.g., V4, V7), and thus, the results are likely less conditioned by the DNA fragmentation that may occur in the sediments when aging26. In addition to dating the sediment core, we used subfossil photosynthetic pigments27 to evaluate the changes in the lake’s trophic state as an indicator of the intensity of the pasturing in the catchment. The erosion caused by sheep herds during the visits to the area affects the lake’s primary productivity more than climatic fluctuations, always within a range of highly oligotrophic conditions that the lake has maintained throughout the Holocene24. Cladoceran remains were also analyzed to evaluate the potential fish occurrence with a complementary approach8 and how they compare with the sedDNA method.
In this work, we show a continued presence of fish parasites in Lake Redon since the 7th century, which coincides with the Late-Roman and Visigothic extensive mountain use for sheep pasturing, supported by nearby archeological remains and increased lake production evidenced by photosynthetic pigments. Cladoceran fluctuations do not appear primarily related to fish presence and, thus, could have hardly been used as evidence of fish introduction. SedDNA of fish parasites appears to be a reliable approach for investigating the history of fish lake introduction across worldwide high mountains.
Results
SedDNA fish evidence
The 30-cm long core RC18 from Lake Redon was stratigraphically homogeneous, consisting of a gyttja (dark organic mud) with a few layers containing small dropstones (Supplementary Fig. 1). Water content was high but similar throughout the core (83.4 ± 2.3%). Dating indicated a maximum age of 3233 cal years BP (1283 BCE), and the sediment accumulation rate shifted at ~4 cm, with an average after that initial part of 0.08 mm year-1 (Supplementary Fig. 2, Supplementary Table 1). Except for markedly high concentrations in samples corresponding to the last few decades, DNA distribution along the core showed a progressive decline from present to past (Supplementary Fig. 3A, B). The number of exact sequence variants (ESVs) recovered from equivalent amounts of amplified DNA from each sample showed a similar decline but more asymptotic, flattening from ca. 300 CE to older samples (Supplementary Fig. 3C). Only a fraction, about one-fourth per sample, of those ESVs could be taxonomically assigned (Supplementary Fig. 3D). Therefore, although the sequencing depth was equivalent for each sample and thus the number of reads obtained, the reads with taxonomic assignment declined progressively up to samples from 300 CE (Supplementary Fig. 3E). Eventually, the yield of reads with taxonomic assignment per DNA sediment concentration showed an abrupt shift at ~1300 CE (Supplementary Fig. 3F), suggesting a more fragmented and altered DNA for older ages, with consequent more limited taxonomic assignment. Occasional spikes of high reads across the core corresponded to oligochaete ESVs.
Although fish was currently present in Lake Redon, no direct DNA fish evidence was found with a battery of primers (Supplementary Table 2) for directly assessing fish occurrence using top sediment samples. In contrast, some trials did provide positive fish detection in a shallow lake (Llebreta) with high fish density in a nearby valley (Supplementary Table 2). Therefore, we concluded that it was a matter of fish density rather than inadequate primers, and they were not used to analyze downcore samples. In contrast, using primers for the region V9 of the18S rRNA gene for a general eukaryote screening along the RC18 core, we detected several reads ( < 8 per sample, usually 1 or 2) in nine samples between 1900 CE and the present and one sample around 1200 CE, which were assigned to Teleostei (support, 0.86—we will use the term “support” to refer the bootstrap confidence of the taxonomic assignment, 1 is the highest value, see methods for details). In contrast with this poor direct fish evidence, several potential ectoparasites of fish (i.e., 46 ESVs) belonging to four major clades were identified: Kinetoplastea (19 ESVs), Oomycota (12), Ichthyosporea (11), and Ciliophora (4) (Fig. 2). Overall, after denoising and taxonomic assignment, 12,718 ESV reads were assigned to fish parasites of a total of 1,151,243. They were detected in 72 core samples, with an average of 1.8% with respect to the total sample reads, a median of 0.7%, and a maximum of 28%.

Fish parasites were assessed by amplifying the region V9 of the 18S rRNA gene in sedDNA. Each plot corresponds to the four main clades identified: Kinetoplastea (A), Ciliophora (B), Icthyosporea (C), and Oomycota (D) The taxonomic clusters (TCs) distinguished are indicated with different colors. They were named with the lowest taxonomic level that the sequence supported with bootstrap confidence. In all cases, the organism most likely to support the sequence was a fish or suspected fish parasite (Supplementary Table 3). In the background (grey), the total number of reads for the entire high-taxonomic clade indicates the group’s DNA preservation, which was better for Kinetoplastea. Source data can be found in Supplementary Data 1.
Kinetoplastea parasites showed the earliest occurrence of fish parasites and the highest continuity over time, not shown by other groups (Fig. 2). Fifteen ESVs were highly related to the genus Ichthyobodo sequences in the databases (Supplementary Table 3). Most of them showed high support to the genus taxonomic assignment (0.9–1) (Fig. 3A). Four ESVs with lower support (0.65–0.76) were similar among them (Fig. 3B) and showed a higher proportion of thymine in the sequence (Fig. 3B), which could indicate degradation from other sequences28. All the ESVs were merged into a single taxonomic cluster (TC) (Fig. 2) as a paleorecord of similar ecological and archeological information. A few other ESVs were assigned to Prokinetoplastina with high support to the subclass but low to any genus (Supplementary Table 3). Alignment of the sequences showed that all were characterized by insertions of different lengths within the Ichthyododo sequences (Supplementary Fig. 4A) and a higher proportion of thymine bases in the sections in common. They were clustered as Prokinetoplastina-1 (Fig. 2), as they could be alterations of extracellular Ichthyobodo’s DNA. In any case, considering them or not does not make any difference in terms of ecological or archeological main results and conclusions that could be raised from the well-preserved sequences. Ichthyobodo is an intensely studied parasite of fish, including salmonid species such as Salmo trutta. It was first identified in the RC18 core record in samples of the early 7th century (i.e., the first in 609 CE ± 170 years, 95% probability) and consistently since 800 CE and up to the present, with a maximum of reads at 1300–1400 CE and a secondary maximum at 1800–1900 CE. It is worth highlighting that the class Kinetoplastea presented a non-declining record with sample age in the core (Fig. 2A), which suggests exceptional DNA preservation compared to other eukaryotic groups.

A Sequence alignment indicates the consensus sequence and the variants ordered by their supporting bootstrap assignment in the references databases, which is shown following the ESV label and taxonomic assignment. B Neighbor-joining tree74, depicting the similarity between the sequences. Values within the circles indicate the bootstrapping support of the branch; >70% can be considered good, and 50–70% moderate77. C Distribution of reads for each ESV along the sediment core. The changing color indicates the bootstrap confidence of the taxonomic assignment to Ichthyobodo; the lighter, the higher. ESV894 was the most common in four-weekly sediment trap samples during 2017–2018, and six other ESVs were also found (ESV957, ESV2118, ESV487, ESV7556, ESV516, ESV2505). Source data can be found in NCBI GenBank ref. OR539619-OR539672.
Provided the Ichthyobodo’s continuous sediment record until the top sediment, the parasite should also be in the lake’s current water column. Indeed, in a four-weekly survey in Lake Redon during 2017-2018, seven sediment ESVs assigned to Ichthyobodo were confirmed in the samples of the sediment traps, including sequences with high and low support. They showed a seasonal pattern, with a marked maximum in August during the two years sampled and fewer reads during the ice-covered period (Supplementary Fig. 5). The number of Ichthyobodo reads was always low in the trap samples (10-80 reads per 50.000–80.000 total ESV reads), which explains why it was not detected in the integrated water samples collected during the visits.
The suspected fish parasites of the other three main clades showed a more fragmented occurrence (Fig. 2). Within Oomycota, ten ESVs were well supported for assignment to the Aphanomyces genus, and two with poor taxonomic resolution were labeled as Oomycota-5 (Supplementary Table 3). The support for the taxonomic assignment of ESVs to Aphanomyces also declined with the increase of thymine bases in the sequences (Supplementary Fig. 4B). The alignment of Oomycota-5 sequences indicated a high similarity to Aphanomyces but enriched in adenine or thymine (Supplementary Fig. 4B). Aphanomyces can be a parasite of different organisms (e.g., crop plants, crayfish), but in the context of this lake, it most likely corresponds to fish in which it produces epizoic ulcers29. They occurred frequently and showed a relatively large number of reads compared to other groups (Fig. 2).
In contrast, reads were consistently low for the third large clade, Ichthyosporea (Fig. 2), yet they were detected frequently since 900 CE. The two orders found (Ichthyophonida and Dermocystida) include primarily fish and amphibian parasites, but they can have other hosts30. Some ESV sequences highly supported those of fish parasite genera (e.g., Dermocystidium) (Supplementary Table 3). However, the ESV alignment and clustering showed a higher variation within this clade than in the other three classes, and most of it does not seem related to DNA degradation (Supplementary Fig. 4C). Therefore, the ESVs were classified into five TCs according to the taxonomic resolution of the assignments and sequence similarities. The taxonomic assignment confidence was very high for most ESVs but at relatively high-rank taxonomic levels, perhaps due to the lack of adequate sequences in the databases; only in one case was there high support at the genus level (Dermoscystidium, 0.94).
In the case of the Ciliophora, the fish parasite evidence was weaker than in the other tree clades. Nevertheless, it is worth commenting as there was a rich record of ESVs assigned to genera identified in previous studies of ciliates in water samples from the lake using microscopy31,32, and a distinctive group of four ESVs differentiated from them clustering together in an Oligohymenophorea branch. Some of them were assigned to Trichodina (Supplementary Table 3), a fish parasite, although with meager bootstrap support. Alignment of the sequences showed that they were closely related (Supplementary Fig. 4D) and that a slight variation in the sequence can cause a marked decline in the assignment support to Trichodina sequences in the reference databases. For instance, the change of only one nucleotide between ESV4044 and ESV1452 caused a decline from 0.72 to 0.58, which made us suspect that perhaps all four ESVs could be degradation forms of the same organism. We divided them into two TCs for visualization (Fig. 2). They occurred in a few samples, but the number of reads was much higher than in the case of the other clades. These features suggest poor preservation in the sediments of Trichodina DNA if this is the origin of the recovered sequences.
Oomycota, Ichthyosporea, and Ciliophora entire ESV records declined the older the samples in contrast to Kinetoplastea (Fig. 2). Therefore, we may expect that the probability of finding the fish parasite forms of these groups as early as Ichthyobodo was low. All in all, fish parasite forms of Ichthyosporea and Oomycota were detected regularly after 900 CE, well before expected from historical documents.
Lake historical ecological context
The ecological context framing the fish parasite presence was evaluated using the sediment record of photosynthetic pigments as indicators of the primary producer community in the lake and cladocerans as potential indicators of fish impacts in the food web. Many pigments corresponding to photosynthetic organisms and some of their degradation products were found (Supplementary Table 4). A Principal Components Analysis (PCA) of their concentration in the sediment showed a first axis, accounting for 72% of the total variance, which indicated a general coherence of the fluctuations among the main algal groups with changes in lake productivity (Supplementary Fig. 6). The second axis accounted only for 15% and was related to the diagenetic process and the differential degradation of some particularly unstable pigments in the sediments (e.g., fucoxanthin)33.
All eukaryotic primary producers have chlorophyll-a as the main photosynthetic pigment. Therefore, a-phorbins concentration (chlorophyll-a and its related degradation forms: pheophytin, pheophorbide, and chlorophyllide) was used as an indicator of lake primary productivity over time (Fig. 4). The first prominent change in a-phorbins occurred around 200 BCE (Fig. 4A), indicating a shift in lake productivity. The pigment composition also showed a discontinuity (Supplementary Fig. 6). The more productive conditions continued until today, although with marked fluctuations. In particular, the period 1000–1300 CE showed a distinctive higher productivity across the entire record. Afterward, the values dropped abruptly to about half in less than one hundred years. The decline continued smoothly for about six centuries and only recently ( ~ 1975 CE) increased again. Despite the long progressive decline during the last centuries, values never reached low levels similar to those before 200 BCE.

A a-phorbins as indicators of the changes in total lake productivity. The individual phorbins are indicated in different colors corresponding to chlorophyll-a (Chla), pheophytin-a (Pheoa), pheophorbide-a (Phora), and chlorophyllide-a (Chlpa). B The alloxanthin/a-phorbin ratio indicates the contribution of cryptophytes, an exclusively planktonic group, to lake productivity. C Diatoxanthin/a-phorbin ratio indicates the diatoms’ contribution to lake productivity. D Lutein/a-phorbin ratio indicates the chlorophytes’ contribution to lake productivity. Source data can be found in Supplementary Data 2.
Because of the general coherence among pigments, as indicated by the high variance explanation by PC1, ratios of other indicative pigments with a-phorbins can reveal details about community changes. Alloxanthin is a pigment exclusive of cryptophytes with excellent sediment preservation34. Lake Redon phytoplankton is currently dominated by chrysophytes, characteristic of ultraoligotrophic waters35, and cryptophytes, the first to respond to episodic nutrient enrichments36. Cryptophytes are exclusively planktonic and generally follow the productivity in this habitat over an extensive range of nutrient conditions37. The alloxanthin/a-phorbin ratio peaked at about 900 CE (Fig. 4B), suggesting an increased contribution of benthic habitats after that date. Diatoms and chlorophytes occur across all habitats in the lake. The ratio of their primary indicative pigments (diatoxanthin and lutein, respectively) to a-phorbins showed a shift around 450 CE (Fig. 4C, D): diatoms declined, and chlorophytes increased their relative contribution. During the highest lake productivity period (900–1300 CE), the relative contribution of chlorophytes (i.e., lutein/a-phorbins) was markedly higher, consistent with a situation of increased external phosphorus loading, which is the limiting nutrient in the lake, as shown experimentally36.
Six cladoceran species were identified in the sediment remains: Daphnia pulicaria Forbes, 1893 is planktonic; the rest are benthic dwellers in the littoral zone: Chydorus sphaericus (O. F. Müller, 1776); Alona affinis (Leydig, 1860); Alona sp.; Acroperus harpae (Baird, 1834); and Eurycercus lamellatus (O. F. Müller, 1776). E. lamellatus is the largest cladoceran in the lake (up to 6 mm), followed by D. pulicaria (<3 mm), and the rest are small species ( < 1.5 mm). The abundance of cladoceran remains roughly followed a-phorbin general patterns (Supplementary Fig. 7). Most species populations increased when the lake became more productive up to the maximum at 900-1400 CE. However, the synchrony decayed after that period. Some cladoceran species declined following a-phorbins (i.e., E. lamellatus, C. sphaericus), but others maintained or even increased their populations with respect to a-phorbins concentrations (A. affinis, A. harpae, and D. pulicaria) (Supplementary Fig. 8). The abundance shift was abrupt at ~1400 CE in the species that declined, with Eurycercus, the largest species, showing a marked relative change compared to the more abundant species (Fig. 5).

Eurycercus lamellatus remains against the other two most abundant littoral cladocerans: Alona affinis (A, C), and Chydorus sphaericus (B, D). E. lamellatus is the largest species (up to 6 mm), and the other two species are smaller ( < 1.5 mm). A, B indicate the ratio distribution during three main periods corresponding to (i) an early phase of low or medium lake productivity (before 800 CE), (ii) the period of the highest lake productivity (800−1400 CE), and (iii) a period of intermediate productivity during the Little Ice Age and the posterior decades ( > 1400 CE). Box plot elements include: center line, median; box limits, first and third quartiles; whiskers, 1.5x interquartile range; and upper characters indicate distributions with a mean significantly different from those with another character (Pairwise two-sided Tukey test after significant ANOVA, p-values < 0.05 were considered significant. The number of cases is indicated in red at the bottom of each distribution). C, D show the temporal series of the ratio. Red bars indicate the periods. Source data can be found in Supplementary Data 3.
Discussion
Observations and documents suggested continued fish populations in Lake Redon for the last two centuries. We found evidence of much earlier fish introduction using sediment DNA fingerprints of fish parasites. Despite trying several eDNA approaches, including those specific for trout and some more taxonomically comprehensive, we could not find any direct fish signal in the lake top sediments, likely because of the low fish density as some methods were successful in another nearby lake, shallower and richer in fish. However, using a general eukaryotic indicator, the 18S rRNA gene, across a sediment core encompassing the last millennia, we found a rich record of organisms with life forms suspected of fish parasitism belonging to four major clades (Kinetoplastea, Oomycota, Ichthyospora, and Ciliophora). Except for those of Ciliophora, all suspected fish parasites indicated earlier fish presence in the lake than expected from historical documents, specifically concerning Lake Redon or other Pyrenean and European sites.
The ESVs identified as indicators of potential fish parasites showed contrasting precision and accuracy in their taxonomic assignments. Resolution and confidence of taxonomic assignments depend on the length and variation of the targeted region, DNA preservation in the sediments, and adequate reference sequences in the databases. All these aspects probably influenced our results to some degree. We used region V9 of the 18S rRNA gene, which is short compared to other regions and thus, in some cases, could limit the achievable taxonomic resolution. However, the bootstrapping confidence at the genus level was the highest for several relevant sequences. On the other hand, short regions could be less sensible to the fragmentation of ancient DNA. Increased fragmentation with sample age was present. The fraction of total ESVs that could be taxonomically assigned declined asymptotically with age, stabilizing at samples >1700 years old. The probability of finding a specific taxa declined in old samples. However, the asymptotic behavior suggests differential decaying rates among taxa. Indeed, there was a high contrast between total reads assigned to Kinetoplastea and Ciliophora ESVs: the former showed a flat profile of detection from top to bottom of the core, whereas the latter declined rapidly, becoming rare in the older samples, despite there is no doubt that a large variety of ciliate species exist in a lake at any time38. Ichthyosporea and Oomycota did not show sufficient non-parasite records to evaluate the general behavior of the clades. In addition, some sequenced DNA fragments can correspond to alteration forms of the original sequences39. Hydrolysis and oxidation damage in the nucleotide bases after cell depth or in extracellular DNA can enhance the probability that polymerases increase the fraction of typically thymine and occasionally adenine during amplification28,40. Interestingly, the support to the taxonomic assignment decreased with the relative presence of thymine bases in the sequences of the same TC (i.e., Ichthyobodo, Aphanomyces). These potentially altered sequences were much less abundant in the samples than those with high support, and their proportion did not show a relationship with the sample’s age. This kind of damage is more likely in oxic conditions; therefore, the pattern found suggests that it could have happened during the long transit from surface waters to sediments in this transparent and relatively deep lake. If this interpretation is correct, the variation in these otherwise similar sequences will indicate a false biological diversity, an artifact that will not be solved using a fixed identity distance for grouping sequences. Interestingly, the sequences assigned to Ichthyobodo in the sediment traps included many found in the sediments, emphasizing the value of the paleorecord. Understanding the origin of the observed variation will require more specific research beyond the scope of this study.
On the other hand, if sequence alteration is high, taxonomic assignment is only reliable at high taxonomic ranks and with relatively low confidence (e.g., Oomycota-5). In this context, the case of poorly supported assignments to Trichodina motivates further research because it is a common fish parasite, and ciliates show a singular genomic system41. In this study, when detected, the number of reads was much higher than for other parasite ESVs; thus, we can speculate about a very episodic presence or a difficult DNA preservation in the sediments of these taxa. Finally, when the taxonomic assignment support is high at intermediate levels and drops markedly at the genus level, it may indicate poor reference sequences in the databases; this was the case for several of the Ichthyosporea ESVs. Improvement of the availability of reference sequences in databases and a better understanding of the natural variability of barcoding sequences, including taphonomic aspects, should increase the precision and confidence of the taxonomic assignments of organisms hard to collect or observe by other means. All things considered, our results provide robust evidence of an early introduction of fish in Lake Redon and suggest that the population has experienced a potentially high diversity of parasitic interactions across the centuries.
The Ichthyobodo (Kinetoplastea) record was particularly relevant because of its early appearance, persistence over time, unequivocal DNA signal, and being a common salmonid parasite42. It was first detected in a few samples during the 7th century and recurrently since ~800 CE to the present, suggesting fish presence in the lake for thirteen centuries. We confirmed the current occurrence of this parasite in the lake, which was detected permanently over the year, with peaks in August. The excellent preservation of other Kinetoplastea ESVs, unrelated to fish parasites, across the ~3400 years of the core record supports the reliability of Ichthyobodo’s chronology.
Documented evidence of first fish occurrences in high-mountain lakes is scattered across space and time. In the Pyrenees, the first written evidence dates from the 14th century4, and it is one of the worldwide ranges with earlier documentation2. Even so, these documented records are more recent than the socio-economic splendor in many valleys of this range during the Middle Ages and centuries later than the beginning of extensive mountain land use for pasturing livestock43. Therefore, the results are coherent with the initial hypothesis that early fish introduction could have occurred during the early centuries of extensive and intensive use of the mountain’s resources (Fig. 6). Our finding aligns with the emerging evidence based on natural registers of relevant human activities in high mountains (e.g., mining) before the culture of writing extended in the Middle Ages44.

Photosynthetic pigments (green siluette) indicate fluctuations in the lake’s primary productivity, driven mainly by sheep livestock impact since 200 CE, with a high incidence during feudalism, which overlapped with the Medieval Climate Anomaly. Fish parasites (orange silluete), represented by Ichthyobodo, indicate a sustained fish occurrence since at least the 6th century. The variation in the Eurycercus lamellatus (purple silluette), the cladoceran most susceptible to fish predation, suggests that other drivers than fish impact were more relevant as drivers of the cladoceran community composition. The upper blue scale indicates the main socio-economic periods in the region. Source data can be found in Supplementary Data 1, 2, and 3.
Previous studies of a sediment record of Lake Redon (RC94), covering the entire Holocene, showed that the low sediment accumulation rate maintained during millennia began to increase at about 450 CE due to enhanced erosion in the catchment, achieving a new sediment accumulation regime that lasted until present45. The pigment record followed the same pattern24, indicating that erosion enhanced phosphorus external loading and, thus, lake productivity. The RC18 core studied here has higher temporal resolution than core RC9445, allowing us to infer that the increase in lake productivity could have started about two centuries earlier, during the 3rd century. As a matter of fact, archeological prospections have described and dated (135-335 CE) a historical site at the Tuc deth Lac Redon (2400 m a.s.l), a mountain pass between Lake Rius and Lake Redon43 in the easternmost part of the watershed. Fourteen livestock enclosures were identified that were likely used to hold 50–100 animals, given their size. According to the sedimentary record, the first prominent productivity peak and increased chlorophyte relevance occurred in the 5th century. The fish parasite presence started not much later and was consolidated during the period of higher lake productivity (900–1300 CE), which corresponds with the period of greater socio-economic wealth in the valleys of this Southern Central Pyrenees area, with flourishment of Romanesque art25 and expansion of transhumance46. It also corresponds to a historical peak of lead concentration in the lake sediments corresponding to nearby mining activity in the down valley that started in the Ibero-Roman period23.
During this period of higher lake productivity, livestock evolved to practices with fewer shepherds carrying larger flocks43. However, it also corresponded with the Medieval Climate Anomaly (MCA), which included warmer and drier conditions in the region47. Pastures in the Lake Redon watershed are poor because of the basin’s rocky and sloppy nature. Currently, sheep flocks only graze in the lake’s watershed for a short period at the end of summer (i.e., a few days to weeks) when better pastures at lower elevations are exhausted. In drier years, the upward movement of the flocks occurs earlier. Therefore, it might happen that the more productive period responded to new livestock practices and earlier and longer sheep presence in the watershed during summer. In any case, fish introduction in the lake happened at an early stage of intensive use of the mountain, which makes sense in a society used to fish in streams for fresh food.
Around 1300–1400 CE, due to the feudal crisis and the Little Ice Age (LIA), the human population significantly reduced in the area with the abandonment of numerous farms. Our results showed an abrupt decrease in the lake’s primary production at these dates and a steady low decline throughout the following centuries until the 1950s. The interaction between humans and the environment and the resilience of societies to climate change is controversial. The feudalism system could not have been resilient because low political participation appears to be more vulnerable to catastrophic climate-related disasters48. However, the climatic severity of the Little Ice Age and its potential for driving socio-economic crises is still uncertain. Average climatic conditions were not radically different, but droughts, floods, and cold/heat waves increased with high spatiotemporal variation across the Iberian mountains49. The Little Ice Age was not climatically homogenous, and 1300–1400 CE initial cooling was not harsh enough to cause a sharp decline in livestock impacts. It could be speculated that climate accelerated an already developing socio-economic break, particularly facilitating epidemics50. The sharp decline in lake productivity did not substantially modify the fish parasite profile, indicating a well-established fish community (Fig. 6), including the first part of the Little Ice Age ( ~ 1400 CE), which was climatically harder in the area according to reconstructions of winter conditions using chrysophyte cysts in the Lake Redon RCA94 core51,52. Longer ice-cover periods may have favored parasite transmission as found in subarctic lake salmonids53.
At the beginning of the Little Ice Age, the sharpest decline in E. lamellatus remains occurred (Fig. 6). Using the traditional assessment of fish introduction based on cladocerans remains, this decline could have been identified as the moment of fish introduction. There is consensus that the larger the species, the higher the fish predation pressure on them8. However, E. lamellatus is the cladoceran that is less venturing into cold waters among those species in the lake (Supplementary Fig. 9); specifically, it shows warmer and more productive habitat preferences than A. affinis12,54. Therefore, the relative decline during the Little Ice Age of this species could be due to the colder conditions. In shallow mountain lakes, the fish impacts in the lake community are usually more apparent2,3,6. The independent assessment of the fish introduction using fish parasite molecular fingerprints may improve the evaluation of such impacts in the past, using a broad spectrum of fish prey indicators, either body remains (e.g., chironomids55) or DNA more specific barcoding56.
Overall, the study demonstrates that assessing fish introductions in high-mountain lakes using the DNA fingerprints of their natural parasites offers the possibility to develop a detailed survey across regions, ranges, and continents and investigate the cultural and natural conditions that stimulated the practice and their consequences for lake community structure. The historical patterns and impacts may be confronted with current practices of massive introduction in some ranges for sport fishing. Molecular evidence of fish parasites in Lake Redon sediments indicates fish introduction since at least the 7th century, much earlier than documentary evidence suggested, and coherent with the period of increased mountain land use in the area. Given that the culture of writing and preserving documents in Europe only underwent a revolution during the 12th and 13th centuries57, it is likely that European ranges and worldwide mountain lake areas could have experienced a similar early fish introduction when historical land use accelerated. The methodological approach used here is an example of how paleolimnology can help bypass archeology’s limitations to study some anthropogenic actions in highlands that leave almost no archeological traces.
Methods
Study site
This study is part of Lake Redon’s long-term limnological research program by the Center of High Mountain Research (CRAM) of the University of Barcelona (UB). No specific permissions for the study were required. Lake Redon (Fig. 1) is a headwater high mountain lake at 2240 m a.s.l. in the central Pyrenees (42° 38’ N, 0° 46’ E). In the context of alpine lakes, the lake is large (24 ha) and deep (maximum depth 73 m, mean depth 32 m). The watershed is relatively small (155 ha), with granodioritic bedrock and barely developed soil. It is located above the tree line, with vegetation consisting of alpine meadows and a large percentage of bare rock. The lake has a retention time of about four years and extremely low watershed sediment export23. The lake is dimitic and ice-covered from December to May–June. The lake is ultraoligotrophic, with an average total phosphorus concentration of 0.2 µmol l–1, and the soluble reactive fraction is generally lower than 0.01 µmol l–1 58. The water column is well-oxygenated except in deep layers ( > 60 m) that are hypoxic during winter and lake summer stratification. The study included three differentiated activities: (1) Attempts to detect trout in sediment using several approaches, considering Lake Redon top sediment and, as references, trout muscle, and Lake Llebreta top sediment. (2) Lake Redon RC18 core analyses, including dating, 18S rRNA gene in sedDNA, pigments, and cladoceran remains analyses. (3) Water column and sediment trap 18S rRNA gene analyses for detecting current fish parasites.
Tests of fish detection in sediments
Several species-specific primers for Salmo trutta Linnaeus, 1758 and one set of general fish primers were used aiming to identify the fish presence in the sedDNA samples (Supplementary Table 2). Although positive signals were obtained using fish tissue and Lake Llebreta samples, which is a shallow lake with abundant fish in a nearby valley, no specific signals were obtained for Lake Redon sediment samples, including top and near-surface sediment samples corresponding to periods in which the fish presence in the lake is known. Therefore, only the 18S rRNA gene was analyzed across the RC18 sediment core.
Core description and age-depth model
The selection of the coring site (Fig. 1) was a compromise between a persistent accumulation of fine sediment and the influence of the littoral zone where trout tend to be, with the largest specimens located deep20. The analyzed sediment core was collected at 47-m depth (42°38’ 22.0” N 0°46’ 36.3” E) using a gravity corer59 in July 2018. The core (RC18), 29.4 cm long, was vertically extruded, sliced into 2 mm sections (146 samples), and stratigraphically described (Supplementary Fig. 1). Upper core samples were dated with 210Pb (Edith Cowan University, Australia), and, given the rare presence of plant macrofossils, ten bulk sediment samples were dated using AMS-14C (Beta Analytic, Florida), including some samples overlapping with 210Pb dating to correct for aging effects (Supplementary Table 1). The age-depth model was established based on the two types of dating and using Bacon v 2.4.360 to combine them in a single model (Supplementary Fig. 2). The concentrations of 210Pb were determined through the analysis of 210Po by alpha spectrometry after adding 209Po as an internal tracer and digestion in acid media using an analytical microwave61. Some samples were analyzed by gamma spectrometry to determine the concentrations of 226Ra and 137Cs. The concentrations of excess 210Pb used to obtain the age models were determined as the difference between total 210Pb and 226Ra (supported 210Pb).
The concentrations of 210Pb decreased with depth from the surface (1590 Bq·kg−1) to a relatively constant concentration below 4 cm (average 67 ± 7 Bq·kg−1), consistent with the 226Ra concentrations and taken as supported 210Pb. Using the CF:CS model62, we established two different average sedimentation rates: 0–1.2 cm: 0.190 ± 0.007 mm·yr-1 (or 0.0043 ± 0.0002 g·cm−2·yr−1); 1.2–4.2 cm: 0.44 ± 0.04 mm·yr−1 (or 0.0071 ± 0.0007 g·cm−2·yr−1).
The change in the sedimentation rate would have occurred during the late 1950s. We considered applying the CRS model63, which would require assuming that the flux of excess 210Pb had been constant, a condition that may be compromised given the apparent change in the sedimentation rate. However, if we dismiss this potential issue, we obtained geochronology using the CRS model compatible with the CF:CS model results.
The concentrations of 137Cs decreased with depth from the surface and down to 2.9 cm, below which it was not detected, and with a relative maximum at 2.5 cm depth. The 137Cs data did not allow for confirming the 210Pb dating. One would expect to identify three specific markers: i) first appearance during the 1950s, corresponding to the beginning of the nuclear tests in the atmosphere; ii) a relative maximum in 1963, corresponding to the peak of nuclear detonations; and iii) a relative maximum in 1986, due to the nuclear accident in Chornobyl. This pattern was not identified, which could be attributed to the penetration of 137Cs to deeper layers once deposited on top sediments, likely due to these sediments’ elevated water content.
Organic bulk sediment (gyttja) samples were radiocarbon AMS dated. Samples were pretreated with acid digestion, and the conventional radiocarbon ages were corrected for total isotopic fractionation effects. The lake is in a granodiorite basin; however, existing faults with calcite precipitation provide small amounts of old carbon that strongly influence the 14C fraction due to the water’s low alkalinity ( ~ 50 µeq/L). Samples with 210Pb and 14C data were used to estimate a reservoir effect (1189 ± 79), which was considered in developing the Bacon age-depth model. IntCal20 was used for 14C calibration64. Finally, the age-depth model was based on nineteen 210Pb and seven 14C dates (Supplementary Table 1).
Core sedDNA analysis
Two subsamples for each core slice were collected from the inner part to avoid contamination from the core walls. The samples were stored with 0.23 ml lysis buffer (40 mM EDTA, 50 mM Tris pH 8.3; 0.75 M sucrose) at −80 °C until the DNA extraction of one of them, keeping the other as backup. Subsampling, DNA extraction, and PCR were carried out in different laboratories to prevent contamination. We processed 146 core sediment samples and 18 blanks. DNA extraction was performed using ~0.8 g of wet sediment and the DNeasy® PowerSoil® kit according to the manufacturer’s instructions (Qiagen, Hilden, Germany). DNA concentration was quantified using a Qubit® 3.0 fluorometer (Thermo Fisher Scientific Inc.) and Qubit dsDNA HS Assay kit, which simultaneously shows a high sensitivity and broad dynamic range (0.2 to 100 ng/mL).
We use the hypervariable region V9 of the 18S rRNA gene to assess the protist fish parasite diversity. The primers used were 1391 f (5’-GTACACACCGCCCGTC-3’) and EukBr (5’-TGATCCTTCTGCAGGTTCACCTAC-3’), which are based on Amaral-Zettler et al.65 and recommended by the Earth Microbiome Project (https://earthmicrobiome.org/protocols-and-standards/18s/). The primer has been highlighted as showing a low interference by bacterial sequences and applied for parasite detection66,67. Library preparation, amplification, and sequencing on a MiSeq PE300 (Illumina, San Diego, CA, USA) were performed at AllGenetics & Biology SL (A Coruña, Spain).
PCRs were carried out in a final volume of 25 µL, containing 2.5 µL of template DNA, 0.5 µM of the primers, 12.5 µL of Supreme NZYTaq 2x Green Master Mix (NZYTech) supplemented with bovine serum albumin (BSA) at a final concentration of 0.1 µg/µL, and ultra-pure water up to 25 µL. The reaction mixture was incubated as follows: an initial denaturation at 95 oC for 5 min, followed by 35 cycles of 95 oC for 30 s, 54 oC for 30 s, 72 oC for 30 s, and a final extension step at 72 oC for 10 min.
The oligonucleotide indices required for multiplexing different libraries in the same sequencing pool were attached in a second PCR round with identical conditions but only 10 cycles and 60 oC as the annealing temperature. A negative control that contained no DNA (BPCR) was included in every PCR round to check for contamination during library preparation. The libraries were run on 2% agarose gels stained with GreenSafe (NZYTech) and imaged under UV light to verify the library size. Two samples failed to yield visible bands in the gel or yielded multiple bands. For the sample that failed to yield a visible band, the optimization consisted of using 5 µL of DNA (instead of 2.5), lowering the annealing temperature, and raising the final concentration of BSA in the mix. The optimization of the sample that yielded multiple bands consisted of raising the annealing temperature, lowering the final concentration of BSA, and keeping the amount of DNA at 2.5 µL. The results of these samples did not differ from the neighboring ones. Libraries were purified using the Mag-Bind RXNPure Plus magnetic beads (Omega Biotek), following the instructions provided by the manufacturer. After amplification, samples were pooled in DNA equimolar proportions based on the quantification data provided by the Qubit dsDNA HS Assay (Thermo Fisher Scientific). This procedure results in a comparable sequencing depth for each sample because the pool was sequenced in a fraction of a MiSeq PE300 v3 run (Illumina). Only a few samples could not be pooled in equimolar amounts due to the low yield after PCR, but they were included in the pool nevertheless. Except in these few samples, the detection effort was therefore equivalent among samples, independently of the initial DNA content in the sediment. For this reason, the results are reported in the number of reads of exact sequence variants (ESVs).
The eukaryotic 18S rRNA gene sequences were processed using USEARCH v1168 and the open-source alternative Vsearch v2.13.469. The paired-end reads were merged, controlling the number of mismatches in the alignment and the minimum length for the merged sequences. Low-quality reads were stringently filtered, discarding reads with an expected number of errors >0.25. After dereplication, 453,005 unique sequences were obtained. The UNOISE70 algorithm was used to denoise reads with a reduced min size of 4 and remove chimeras71, obtaining 8,658 ESVs and an average of 68,542 reads per sample. Blanks were not all absolutely clean but showed very low reads (median 11, min 0, max 182) corresponding to a few ESVs (median 5, min 0, max 30); their taxonomical assignment indicated that none corresponded to targeted organisms.
ESV taxonomy was assigned using the SINTAX algorithm71, and SILVA_18S_12372, and PR2 4.13.073 reference databases. The algorithm predicts taxonomy by using k-mer similarity to identify the top hit in the reference database and provides bootstrap confidence for all ranks in the prediction. Then, we followed a supervised procedure to obtain a unique ESV taxonomic assignment and identify those ESVs of interest (i.e., potential fish parasites) in taxonomic clusters (TCs) of similar taxonomic resolution and life form, which could facilitate the presentation and discussion of the results. The main steps were: (1) The two databases usually provided different taxonomic resolutions and bootstrap confidence for the same taxonomic rank taxa because they use different reference sequences. Therefore, we merged the two sources of information into a single taxonomic assignment, prioritizing the highest bootstrap support in the databases. (2) Because SILVA and PR2 do not follow the same nomenclature for the taxonomic categories, we harmonized the taxonomy following NCBI nomenclature. ESVs with low taxonomic resolution and taxonomic inaccuracy were discarded. 2,468 ESVs were kept, corresponding to a total of 1,151,243 reads. (3) We checked for taxa with potential fish parasite life forms. There were cases with the highest possible bootstrap support to the genus level (e.g., Ichthyobodo, Aphanomyces), others with high support of higher rank taxonomic levels that include mainly fish parasites but also parasites of other organisms (e.g., Dermocystida) and, finally, others with low support to a genus or group that are fish parasites (e.g., Trichodina). All the ESVs (46) with a potential indication of fish parasitism were part of either Kinetoplastea, Oomycota, Ichthyosporea, or Ciliophora. (5) For each of these main clades, all ESVs assigned to the clade were compared using neighbor-joining trees74 based on an identity matrix calculated with function dist.alignment from the seqinr R package75. This procedure allowed evaluation of how similar the potential parasite ESVs were among them and how isolated they were from other ESVs of the main clade, an aspect that was relevant in cases of low support to the genus level. (6) The ESVs of potential parasites were merged into TCs, joining ESVs with similar taxonomic assignments and clustered closely in the tree. This taxonomic clustering facilitated the exposition of results and discussion of the reliability of the attribution to suspected fish parasites. However, they did not condition the main results in terms of the chronology of fish introduction and persistence of the signal, which could be similarly supported without ESVs clustering. (7) Finally, the ESVs of each main clade suspected of indicating a fish parasite were aligned to evaluate the variation within the TCs and discuss the characteristics of taxonomic assignments with poor bootstrapping confidence. Alignments were performed using DECIPHER76.
Water and sediment trap eDNA
Sampling was conducted every four weeks, from July 2017 to November 2018, to assess the current fish parasites in Lake Redon. Two sample types were collected: an integrated water column sample (from 0 to 60 m depth) and a sediment trap sample recovered from a 12 cm diameter sediment trap placed at 60 m depth. Subsamples (1300–3000 ml for water and 10–80 ml for the sediment trap) were filtered through a 3 μm pore size polycarbonate filter (47 mm Ø) until saturation and preserved with lysis buffer (40 mM EDTA, 50 mM Tris, pH 8.3, and 0.75 M sucrose) at −80 °C until DNA extraction. The DNA extraction and the amplification of the V9 region of the 18S rRNA gene were performed together with the core samples.
Pigments analysis
Core slice sub-samples for pigment analysis were kept at −20 oC in darkness and freeze-dried before extraction, which was performed using sonication of ~200 mg of dry sediment in acetone (90%). The extract was filtered through 0.2 μm hydrophilic polytetrafluoroethylene (PTFE) filters (MILLEX LG Merck) and analyzed by HPLC. The HPLC system (1100, Agilent Technologies) was equipped with a C18 column (dimensions: 250 x 4.6 mm, particle size: 5 µm; Waters Spherisorb ODS1) and an online pre-column filter (Microfilter PEEK 0.5 µm, Supelco – Merck). Detection was performed using a diode-array Agilent 1100/1200+. The detector was set at 440 and 660 nm wavelengths for carotenoid and phorbin peak integration, respectively. After sample injection (40 µl of acetone extract), pigments were eluted by a 35 min gradient from 100% solvent A (0.3 M ammonium acetate in methanol: acetonitrile: MilliQ water, 51:36:13 (v/v/v)) to 100% B (ethyl acetate: acetonitrile, 7:3, (v/v)). Solvent A ran 10 min between samples. The flow was constantly set at 0.6 ml/min. The pigment identification was based on the HPLC retention time and pigment absorption spectrum using a commercial mixture of phytoplankton pigments (mix-123, DHI Laboratory Products, Hoersholm, Denmark) and from a house-built library of pigment spectra when the standards were unavailable. Pigment main patterns were assessed by Principal Component Analysis using R package stats. About 72% of the total variation in the data was explained by the first component (PC1), which indicates a robust linear relationship among most of the pigments. Therefore, there were no great substitutions in algal group dominance. We could have used the PC1 scores as a reference for lake productivity. Still, to make it less abstract and more comparable to other studies, we used total a-phorbins (chlorophyll-a and its related degradation forms), which makes little difference than considering PC1 scores because of their high correlation.
Consequently, the ratio between some pigments representative of major algal groups (i.e., alloxanthin for cryptophytes, diatoxanthin for diatoms, and lutein for chlorophytes) and total a-phorbins, representative of total algal biomass, was used to evaluate the relative deviation of these algal groups respect the main trend. The pigments with the highest load in the PC2 were those known to be less stable in the sediments. Primarily, fucoxanthin, a carotenoid in chrysophytes and diatoms, quickly decomposes to colorless forms. The compounds with quick degradation show a typical log-decaying pattern over time. As indicated by the amount of variation explained by PC2 (about 15%), the degradation process had a minor influence on the long-term pigment record compared to lake productivity. Consequently, the ratio between some highly stable pigments and representative of major algal groups (i.e., alloxanthin for cryptophytes, diatoxanthin for diatoms, and lutein for chlorophytes) and total a-phorbins, representative of total algal biomass, was used to evaluate the relative deviation of the primary algal groups with respect of the main trend.
Cladoceran analysis
Approximately 1 cm3 of each sediment slice was weighed and stored in plastic vials. The samples were washed with tap water and filtered through a 60-µm sieve to remove fine inorganic and organic particles. The material left on the mesh was transferred to a vial with 10 ml volume. Before the stereo microscope analysis, the sample was well shaken, and 1 ml of suspension was transferred to a Petri dish and screened for crustacean remains at 40x magnification. The remains were classified as head shields, valves, and postabdomens and were taxonomically identified. Each species’ most frequent body part was selected to report the concentrations of the remains. Eggshells from hatched neonates of Daphnia, postabdomens, and filtering combs were present in the upper sediment samples but markedly declined in lower sections. So, Daphnia was mainly evaluated by the ephippia remains. In contrast, benthic species, which show a more compact armor, provided a consistent record throughout the core.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All the data in this study are provided in supplementary files. The parasite sequences generated in this study are available in the NCBI GenBank (https://www.ncbi.nlm.nih.gov/genbank/), accession codes OR539619 to OR539672, the code complete list can be found in Supplementary Data 4. Source data for Fig. 2 can be found in Supplementary Data 1. Source data for Fig. 4 can be found in Supplementary Data 2. Source data for Fig. 5 can be found in Supplementary Data 3. Source data for Fig. 6 can be found in Supplementary Data 1, 2, and 3. No core samples remained available after processing.
References
Knapp, R. A., Corn, P. S. & Schindler, D. E. The introduction of nonnative fish into wilderness lakes: good intentions, conflicting mandates, and unintended consequences. Ecosystems 4, 275–278 (2001).
Ventura, M. et al. Why should we preserve fishless high mountain lakes? In: High mountain conservation in a changing world (eds Catalan J., Ninot J. M., Aniz M. M.) (Springer, 2017).
Brancelj, A., Šiško, M., Rejec Brancelj, I., Jeran, Z. & Jaćimović, R. Effects of land use and fish stocking on a mountain lake-evidence from the sediment. Period Biol. 102, 259–268 (2000).
Miro, A. & Ventura, M. Historical use, fishing management and lake characteristics explain the presence of non-native trout in Pyrenean lakes: implications for conservation. Biol. Conserv 167, 17–24 (2013).
Pechlaner, R. Historical evidence for the introduction of Arctic charr into high-mountain lakes of the Alps by man. In: Proceedings of the International Symposium on Arctic Charr (eds Johnson J., Burns B. L.). University of Manitoba Press (1984).
Brancelj, A. The extinction of Arctodiaptomus alpinus (Copepoda) following the introduction of charr into a small alpine lake Dvojno Jezero (NW Slovenia). Aquat. Ecol. 33, 355–361 (1999).
Knapp, R. A. Effects of nonnative fish and habitat characteristics on lentic herpetofauna in Yosemite National Park, USA. Biol. Conserv. 121, 265–279 (2005).
Jeppesen, E., Leavitt, P., De Meester, L. & Jensen, J. P. Functional ecology and palaeolimnology: using cladoceran remains to reconstruct anthropogenic impact. Trends Ecol. Evol. 16, 191–198 (2001).
Brancelj, A. Shifts in zooplankton communities in high-mountain lakes induced by singular events (fish stocking, earthquakes): evidence from a 20-year survey in Slovenia (Central Europe). Aquat. Ecol. 55, 1253–1271 (2021).
Jimenez-Seinos, J. L., Alcocer, J. & Planas, D. Food web differences between two neighboring tropical high mountain lakes and the influence of introducing a new top predator. PLoS One 18, e0287066 (2023).
Štillová, V. et al. Tracking fish introduction in a mountain lake over the last 200 years using chironomids, diatoms, and cladoceran remains. Water 15, 1372 (2023).
Catalan, J. et al. Ecological thresholds in European alpine lakes. Freshw. Biol. 54, 2494–2517 (2009).
Catalan, J. et al. Lake Redo ecosystem response to an increasing warming in the Pyrenees during the twentieth century. J. Paleolimnol. 28, 129–145 (2002).
Richard Albert, M. et al. Phosphorus and land-use changes are significant drivers of cladoceran community composition and diversity: an analysis over spatial and temporal scales. Can. J. Fish. Aquat. Sci. 67, 1262–1273 (2010).
Salo, J. et al. Fish predation and reduction in body size in a Cladoceran population: palaeoecological evidence. Freshw. Biol. 21, 217–221 (1989).
Turner, C. R., Uy, K. L. & Everhart, R. C. Fish environmental DNA is more concentrated in aquatic sediments than surface water. Biol. Conserv 183, 93–102 (2015).
Huston, G. P. et al. Detection of fish sedimentary DNA in aquatic systems: a review of methodological challenges and future opportunities. Environ. DNA 5, 1449–1472 (2023).
Capo, E. et al. How does environmental inter-annual variability shape aquatic microbial communities? A 40-year annual record of sedimentary DNA from a boreal lake (Nylandssjön, Sweden). Front. Ecol. Evolut. 7 (2019).
Barber, I., Hoare, D. & Krause, J. Effects of parasites on fish behaviour: a review and evolutionary perspective. Rev. Fish. Biol. Fish. 10, 131–165 (2000).
Encina, L. & Rodriguez-Ruiz, A. Abundance and distribution of a brown trout (Salmo trutta, L.) population in a remote high mountain lake. Hydrobiologia 493, 35–42 (2003).
Zamora, F. Respuestas al cuestionario de Francisco Zamora sobre el Corregimiento del Valle de Arán. Biblioteca del Palacio Real de Madrid (1789).
Garcia-Casas, D., Rodríguez-Antón, D., Costa-Badia, X. & Gassiot-Ballbè, E. Arqueología del paisaje en la alta montaña. Una primera aproximación al estudio de las ocupaciones ganaderas de época medieval en el Parque Nacional de Aigüestortes i Estany de Sant Maurici (Pirineo Occidental de Cataluña). Cuadernos de Arqueología, 30, 211–238 (2022).
Camarero, L. et al. Historical variations in lead fluxes in the Pyrenees (northeast Spain) from a dated lake sediment core. Water Air Soil Pollut. 105, 439–449 (1998).
Catalan, J. Primary production in a high mountain lake: an overview from minutes to kiloyears. Atti della Associazione Ital. di Oceanologia e Limnologia 13, 1–21 (2000).
Lluis, I., Ginovart, J., Ugalde-Blázquez, I. & Lluis-Teruel, C. Gisemundus and the orientation of the romanesque churches in the Spanish Pyrenees (11th-13th centuries). Mediterr. Archaeol. Archaeom. 21, 205–214 (2021).
Capo, E. et al. Lake sedimentary DNA research on past terrestrial and aquatic biodiversity: overview and recommendations. Quaternary 4, 6 (2021).
Leavitt, P. R. & Hodgson, D. A. Sedimentary pigments. In: Tracking Environmental Change Using Lake Sediments. Volume 3: Terrestrial, Algal, and Siliceous Indicators (eds Smol J. P., Birks H. J. B., Last W. M.). Kluwer Academic Publishers (2006).
Hofreiter, M., Jaenicke, V., Serre, D., von Haeseler, A. & Pääbo, S. DNA sequences from multiple amplifications reveal artifacts induced by cytosine deamination in ancient DNA. Nucleic Acids Res. 29, 4793–4799 (2001).
Iberahim, N. A., Trusch, F. & Van West, P. Aphanomyces invadans, the causal agent of Epizootic Ulcerative Syndrome, is a global threat to wild and farmed fish. Fungal Biol. Rev. 32, 118–130 (2018).
Glockling, S. L., Marshall, W. L. & Gleason, F. H. Phylogenetic interpretations and ecological potentials of the Mesomycetozoea (Ichthyosporea). Fungal Ecol. 6, 237–247 (2013).
Felip, M., Bartumeus, F., Halac, S. & Catalan, J. Microbial plankton assemblages, composition and biomass, during two ice-free periods in a deep high mountain lake (Estany Redó, Pyrenees). J. Limnol. 58, 193–202 (1999).
Felip, M. A., Camarero, L. & Catalan, J. Temporal changes of microbial assemblages in the ice and snow cover of a high mountain lake. Limnol. Oceanogr. 44, 973–987 (1999).
Leavitt, P. R. A review of factors that regulate carotenoid and chlorophyll deposition and fossil pigment abundance. J. Paleolimnol. 9, 109–127 (1993).
Reuss, N. & Conley, D. J. Effects of sediment storage conditions on pigment analyses. Limnol. Oceanogr. Methods 3, 477–487 (2005).
Reynolds, C. S. Ecology of phytoplankton. Cambridge University Press (2006).
Zufiaurre, A. et al. Episodic nutrient enrichments stabilise protist coexistence in planktonic oligotrophic conditions. J. Ecol. 109, 1717–1729 (2021).
Buchaca, T. & Catalan, J. Nonlinearities in phytoplankton groups across temperate high mountain lakes. J. Ecol. 112, 755–769 (2024).
Barouillet, C. et al. Paleoreconstructions of ciliate communities reveal long-term ecological changes in temperate lakes. Sci. Rep. 12, 7899 (2022).
Pietramellara, G. et al. Extracellular DNA in soil and sediment: fate and ecological relevance. Biol. Fertil. Soils 45, 219–235 (2009).
Kino, K., Hirao-Suzuki, M., Morikawa, M., Sakaga, A. & Miyazawa, H. Generation, repair and replication of guanine oxidation products. Genes Environ. 39, 21 (2017).
Prescott, D. M. The DNA of ciliated protozoa. Microbiol Rev. 58, 233–267 (1994).
Isaksen, T. E., Karlsbakk, E., Repstad, O. & Nylund, A. Molecular tools for the detection and identification of Ichthyobodo spp. (Kinetoplastida), important fish parasites. Parasitol. Int. 61, 675–683 (2012).
Gassiot Ballbè, E. & Pèlachs Mañosa, A. The husbandry occupation of the western Pyrenees of Catalonia in Roman. Begin. Middle Ages. Treb. d.’Arqueologia 21, 287–306 (2017).
Minvielle Larousse, N. The exploitation of silver deposits in early medieval Europe: some documentary, economic and social problems. Archaeometry. halshs-04757361, https://doi.org/10.1111/arcm.13035 (2024).
Catalan, J., Pla-Rabés, S., García, J. & Camarero, L. Air temperature-driven CO2 consumption by rock weathering at short timescales: Evidence from a Holocene lake sediment record. Geochim. Cosmochim. Acta 136, 67–79 (2014).
García-Ruiz, J. M. et al. Transhumance and long-term deforestation in the subalpine belt of the central Spanish Pyrenees: an interdisciplinary approach. Catena 195, 104744 (2020).
Moreno, A. et al. The Medieval climate anomaly in the Iberian Peninsula reconstructed from marine and lake records. Quat. Sci. Rev. 43, 16–32 (2012).
Peregrine, P. N. Social resilience to climate-related disasters in ancient societies: a test of two hypotheses. Weather Clim. Soc. 10, 145–161 (2018).
Oliva, M. et al. The Little Ice Age in Iberian Mountains. Earth-Sci. Rev. 177, 175–208 (2018).
Moore, J. W. The crisis of feudalism: an environmental history. Organ Envir 15, 301–322 (2002).
Catalan, J., Pla, S., Garcia, J. & Camarero, L. Climate and CO2 saturation in an alpine lake throughout the Holocene. Limnol. Oceanogr. 54, 2542–2552 (2009).
Pla, S. & Catalan, J. Chrysophyte cysts from lake sediments reveal the submillennial winter/spring climate variability in the northwestern Mediterranean region throughout the Holocene. Clim. Dynam. 24, 263–278 (2005).
Prati, S., Henriksen, E. H., Knudsen, R. & Amundsen, P. A. Seasonal dietary shifts enhance parasite transmission to lake salmonids during ice cover. Ecol. Evol. 10, 4031–4043 (2020).
Stoch, F., Vagaggini, D. & Margaritora, F. G. Macroecological and spatial patterns in the distribution of cladocerans in Alpine lakes. Limnetica 38, 119–136 (2019).
Smol, J. P., Birks, H. J. B. & Last, W. M. Tracking environmental change using lake sediments: zoological indicators. Kluwer Academic Publishers (2002).
Capo, E., Barouillet, C. & Smol, J. P. Tracking environmental change using lake sediments. Volume 6: Sedimentary DNA. Springer (2023).
Clanchy, M. T. From memory to written record: England 1066-1307, 3rd edn. Wiley-Blackwell (2012).
Catalan, J. et al. Seasonal ecosystem variability in remote mountain lakes: implications for detecting climatic signals in sediment records. J. Paleolimnol. 28, 25–46 (2002).
Glew, J. R. Miniature gravity corer for recovering short sediment cores. J. Paleolimnol. 5, 285–287 (1991).
Blaauw, M. & Christen, J. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 6, 457–474 (2011).
Sanchez-Cabeza, J. A., Masque, P. & Ani-Ragolta, I. Pb-210 and Po-210 analysis in sediments and soils by microwave acid digestion. J. Radioanal. Nucl. Chem. 227, 19–22 (1998).
Krishnaswamy, S., Lal, D., Martin, J. M. & Meybeck, M. Geochronology of lake sediments. Earth Planet Sc. Lett. 11, 407–414 (1971).
Appleby, P. G. & Oldfield, F. The calculation of lead-210 dates assuming a constant rate of supply of unsupported 210Pb to the sediment. Catena 5, 1–8 (1978).
Reimer, P. J. et al. The IntCal20 Northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757 (2020).
Amaral-Zettler, L. A., McCliment, E. A., Ducklow, H. W. & Huse, S. M. A method for studying protistan diversity using massively parallel sequencing of V9 hypervariable regions of small-subunit ribosomal RNA genes. PLOS ONE 4, e6372 (2009).
Kounosu, A., Murase, K., Yoshida, A., Maruyama, H. & Kikuchi, T. Improved 18S and 28S rDNA primer sets for NGS-based parasite detection. Sci. Rep. 9, 15789 (2019).
Maritz, J. M. et al. An 18S rRNA workflow for characterizing protists in sewage, with a focus on zoonotic trichomonads. Micro. Ecol. 74, 923–936 (2017).
Edgar, R. C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461 (2010).
Rognes, T., Flouri, T., Nichols, B., Quince, C. & Mahé, F. VSEARCH: a versatile open source tool for metagenomics. PeerJ 4, e2584 (2016).
Edgar, R. C. UNOISE2: improved error-correction for Illumina 16S and ITS amplicon sequencing. bioRxiv, 081257 (2016).
Edgar, R. C. SINTAX: a simple non-Bayesian taxonomy classifier for 16S and ITS sequences. bioRxiv, https://doi.org/10.1101/074161 (2016).
Quast, C. et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res 41, D590–D596 (2013).
Guillou, L. et al. The Protist Ribosomal Reference database (PR2): a catalog of unicellular eukaryote Small Sub-Unit rRNA sequences with curated taxonomy. Nucleic Acids Res 41, D597–D604 (2012).
Paradis, E. & Schliep, K. ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35, 526–528 (2019).
Charif, D. & Lobry, J. SeqinR 1.0-2: a contributed package to the R project for statistical computing devoted to biological sequences retrieval and analysis. In: Structural approaches to sequence evolution: Molecules, networks, populations (ed Bastolla U. P. M., Roman H., Vendruscolo M). Springer verlag (2007).
Wright, E. S. DECIPHER: harnessing local sequence context to improve protein multiple sequence alignment. BMC Bioinforma. 16, 322 (2015).
Ota, S. & Li, W.-H. NJML: a hybrid algorithm for the neighbor-joining and maximum-likelihood methods. Mol. Biol. Evol. 17, 1401–1409 (2000).
Acknowledgements
The research was funded by grants from the Spanish Government (Transfer, MCIN/ AEI/ CGL2016–80124-C2-1-P) and Biodivrestore Cofund 2020 (FishMe, PCI2022-132949). E.F. acknowledges her predoctoral fellowship (BES-2017-081553) and A.B. additional funding through the national program P1-0255 of the Slovenian Ministry of Science and Education (ARRS). The authors thank Lluís Camarero, Sergi Pla-Rabès, and Meritxell Batalla for support at different stages of the study; the Servei d’Anàlisi Química (SAQ) from the Universitat Autònoma de Barcelona for chemical analysis advice, and HPLC facilities for pigment analyses; and AllGenetics & Biology SL (A Coruña, Spain) for sequencing.
Author information
Authors and Affiliations
Contributions
J.C. and M.F. conceived and designed the research and coordinated data acquisition and analyses. E.F. and M.F. carried out fieldwork, core subsampling, and description. E.F., M.F., and J.C. performed eDNA analyses. E.F. and J.C. performed pigment analyses. A.B. performed the analysis of cladocerans. P.M. performed 210Pb dating. E.F., M.F., and J.C. analyzed data. J.C. drafted the original manuscript, with contributions and final approval by all co-authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interest.
Peer review
Peer review information
Nature Communications thanks Cécilia Barouillet, and the other, anonymous, reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Fagín, E., Felip, M., Brancelj, A. et al. Parasite sedimentary DNA reveals fish introduction into a European high-mountain lake by the seventh century. Nat Commun 16, 3081 (2025). https://doi.org/10.1038/s41467-025-57801-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-57801-x
This article is cited by
-
Comparative analyses of 18S rDNA regions for assessing the eukaryotic community in ancient lake sediment samples
Journal of Paleolimnology (2025)
