
Abstract
Nutrient enrichment typically causes local plant diversity declines. A common but untested expectation is that nutrient enrichment also reduces variation in nutrient conditions among localities and selects for a smaller pool of species, causing greater diversity declines at larger than local scales and thus biotic homogenization. Here we apply a framework that links changes in species richness across scales to changes in the numbers of spatially restricted and widespread species for a standardized nutrient addition experiment across 72 grasslands on six continents. Overall, we find proportionally similar species loss at local and larger scales, suggesting similar declines of spatially restricted and widespread species, and no biotic homogenization after 4 years and up to 14 years of treatment. These patterns of diversity changes are generally consistent across species groups. Thus, nutrient enrichment poses threats to plant diversity, including for widespread species that are often critical for ecosystem functions.
Similar content being viewed by others

Introduction
Agricultural fertilization practices and atmospheric nutrient deposition have led to increased availability and redistribution of soil nutrients globally1,2,3. At local scales (i.e., α diversity), nutrient enrichment tends to reduce plant diversity4,5. This diversity decline is typically ascribed to disproportionate losses of rare species (i.e., species with relatively low cover) because small populations are more susceptible to extinction6,7,8. In addition, nutrient enrichment often leads to the removal of species with traits ill-suited for effectively competing in high nutrient conditions6. While species can vary widely in their nutrient requirements and tolerances, groups of species with similar (shared) characteristics can be lost from a flora. For example, native species are more likely to be lost than non-native species when nutrients are enriched because non-natives are often better-adapted to nutrient-rich conditions6,9,10. Similarly, nitrogen-fixing legumes may be more vulnerable than other species in high nutrient environments due to their decreased competitive advantage6,11.
Despite clear evidence that nutrient enrichment causes losses of α diversity in grasslands4,5, how these losses are reflected at larger spatial scales (i.e., γ diversity; calculated by aggregating local communities) is less straightforward12,13,14. Yet, it is diversity loss and change at larger spatial scales that is most often relevant for biodiversity conservation and management as well as for the provision of ecosystem functions and services15,16. It is often assumed that nutrient enrichment, like many other global changes, results in biotic homogenization (i.e., increasing similarity in composition among local communities, quantified as a decrease in β diversity)17,18,19,20,21. This is because local nutrient enrichment is expected to create homogeneous nutrient conditions among localities and consistently select for a smaller pool of species that are nutrient-demanding, fast-growing, and highly competitive for light22,23. Previous investigations of scale-dependent diversity change under nutrient enrichment have tended to be short term or limited in spatial extent24,25,26,27,28. These short-term or spatially-restricted studies have found mixed results, indicating that nutrient enrichment leads to biotic homogenization29,30,31, no changes in β diversity24,26,27,32 or even differentiation (i.e., increase in β diversity)25,28,33,34,35,36,37.
A recent extension to Whittaker’s multiplicative β diversity partition enables linking changes in average α diversity (\(\overline{\Delta \alpha }\)), γ, and β diversity (in log scale) to changes in the numbers of spatially restricted and widespread species (Fig. 1)12,38. This framework illustrates how nutrient enrichment could cause biotic homogenization if local communities gain widespread species (Fig. 1: scenario I), if spatially restricted species are replaced by widespread species (Fig. 1: scenario II), or if the number of spatially restricted species decreases (Fig. 1: scenario III). Conversely, nutrient enrichment could cause biotic differentiation if local communities lose widespread species (Fig. 1: scenario IV), if widespread species are replaced by spatially restricted species (Fig. 1: scenario V), or if the number of spatially restricted species increases (Fig. 1: scenario VI). Finally, if gains or losses of species at the α and γ scale are similar (i.e., approximately equal or proportional), then we would observe no change in β diversity (1:1 diagonal line in Fig. 1). However, clear links between changes in diversity across spatial scales and changes in the number of spatially restricted and widespread species under nutrient enrichment are yet to be made.

\(\overline{\Delta \alpha }\), ∆γ, and ∆β are log response ratios (LRR) of average α-, γ- and β- diversity under nutrient enrichment relative to that under ambient conditions (control). ∆β is equal to zero along the 1:1 diagonal line. Below the diagonal line, ∆γ < \(\overline{\Delta \alpha }\), ∆β < 0, nutrient enrichment causes biotic homogenization. Above the diagonal line, ∆γ > \(\overline{\Delta \alpha }\), ∆β > 0, nutrient enrichment causes biotic differentiation. Moreover, homogenization can be attributed to three scenarios including I: gain of widespread species (∆γ < \(\overline{\Delta \alpha }\), and ∆γ > 0, \(\overline{\Delta \alpha }\) > 0); II: spatially restricted species replaced by widespread species (∆γ < \(\overline{\Delta \alpha }\), ∆γ < 0 and \(\overline{\Delta \alpha }\) > 0); III: Loss of spatially restricted species (∆γ < \(\overline{\Delta \alpha }\), and ∆γ < 0, \(\overline{\Delta \alpha }\) < 0). Conversely, differentiation can be attributed to three scenarios that include IV: Loss of widespread species (∆γ > \(\overline{\Delta \alpha }\), and ∆γ < 0, \(\overline{\Delta \alpha }\) < 0); V: Widespread species replaced by spatially restricted species (∆γ > \(\overline{\Delta \alpha }\), ∆γ > 0 and \(\overline{\Delta \alpha }\) < 0); VI: gain of spatially restricted species (∆γ > \(\overline{\Delta \alpha }\), and ∆γ > 0, \(\overline{\Delta \alpha }\) > 0). Adapted from Blowes et al.12. https://doi.org/10.1126/sciadv.adj9395 under a CC BY license: https://creativecommons.org/licenses/by/4.0/.
Here, we use this framework to synthesize scale-dependent plant diversity change (for the entire community and groups of species) under nutrient enrichment using a long-term standardized experiment in 72 grasslands distributed across six continents (i.e., NutNet39; Fig. S1; Table S1). We use two treatments: Ambient (Control) and fertilization by nitrogen, phosphorus, and potassium together (i.e., NPK). Nutrients were added at a rate of 10 g m−2 annually. Treatments were randomly assigned to 5 m × 5 m plots and were replicated in three or more blocks. Species cover was recorded in one 1 m × 1 m permanent subplot using a standardized protocol. At each site, α diversity is determined as the number of species in each permanent subplot (i.e., species richness), and γ diversity as the total number of species occurring in three permanent subplots (for each treatment separately). We exclude additional blocks from sites that have more than three because γ and β diversity depend on the number of local communities used. We calculate ∆α as the richness difference in local communities (subplots) and ∆γ as the difference in the sum of the subplots under nutrient addition relative to that of control on the log scale. That is, ∆α = log(αNPK/αControl) and ∆γ = log(γNPK/γControl). We then calculate ∆β as ∆γ minus \(\overline{\Delta \alpha }\), where\(\overline{\,\Delta \alpha }\) is the average of ∆α over three blocks. Overall, we find proportionally similar species losses at local and larger scales, suggesting similar magnitudes of declines of spatially restricted and widespread species. Thus, we find no clear biotic homogenization or differentiation four years, and even up to 14 years, after nutrient additions began. Moreover, these overall patterns of little change in β diversity hold consistent across species groups.
Results and discussion
Changes in α-, γ-, and β diversity for the entire communities
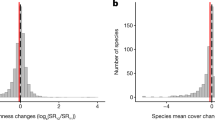
Overall, adding nutrients decreased α and γ diversity, but it had no significant effects on β diversity (∆β = 0.03; 95% credible interval: −0.02 to 0.08) (Fig. 2; Table S3), see also ref. 27. While we observed substantial variation in\(\,\overline{\Delta \alpha }\), ∆γ, and to a lesser extent, ∆β among sites (Table S4), we found no strong relationships between \(\overline{\Delta \alpha }\), ∆γ, and ∆β and distance among blocks within sites, site drought intensity, grazing intensity, productivity, or species pool that have been shown in previous literature to influence diversity change under nutrient enrichment in grasslands24,25,34,40 (Fig. S3). On a site level, we found biotic homogenization at 24 sites, differentiation at 47 sites, and no change in β diversity at one site. However, the site-level 95% credible intervals (see Methods) overlapped 0 for all sites, suggesting no significant change in β diversity with nutrient addition (Table S4). Importantly, the overall effects of nutrient addition on α, γ, and β diversity were similar when we used effective numbers of species based on either Shannon diversity or Simpson diversity that account for species relative covers40 (Fig. S4; Table S3). Because species richness is more strongly influenced by rare species, while Shannon and Simpson diversity increasingly weigh abundant species, this result suggests that relatively rare and abundant species responded similarly to nutrient addition.

LRR: log response ratio. The white 1:1 diagonal line indicates no effects of nutrient addition on β diversity. Numbers in the parentheses are the number of sites. When a site has\(\,\overline{\Delta \alpha }\) = 0, ∆γ = 0, or ∆β = 0, it was not counted into any of the six scenarios as shown in the framework. The small points represent site-level \(\overline{\Delta \alpha }\) and ∆γ at 72 sites. The large open point and error bars are the estimated mean and 95% credible intervals for \(\overline{\Delta \alpha }\), ∆γ, and ∆β across all sites. See Table S3 for model fit and estimated overall means and 95% credible intervals for \(\overline{\Delta \alpha }\), ∆γ, and ∆β. See Table S4 for site-level estimates and 95% credible intervals. Source data are provided as a Source Data file.
Changes in α-, γ-, and β diversity for species groups
The overall proportional species loss within the community at local and larger scales on average may result if different species groups have contrasting patterns of response to nutrient addition. For instance, this result could be the case if native species loss is greater at the larger spatial scale than at the local scale, while non-native species loss is lower at the larger than the local scale. To test this possibility, we investigated changes in α, γ, and β diversity for native and non-native species separately. Extending previous studies6,41, we found that nutrient addition decreased native species more than non-native species. Compared to non-native species, nutrient addition resulted in a 10% greater reduction of α diversity and a 16% greater reduction of γ diversity for native plant species, respectively (Table S5). The overall pattern of diversity change across spatial scales for native species largely followed that of the entire communities with similar magnitudes of decline in α and γ diversity and little change in β diversity (∆β = 0; 95% credible interval: −0.05 to 0.06; Fig. 3A). For non-native species, overall, nutrient addition decreased α diversity 6% more than γ diversity (Table S5). But nutrient addition had no significant effects on β diversity for non-native species (∆β = 0.04; 95% credible interval: −0.05 to 0.14; Fig. 3B).

A Native and B non-native species. LRR: log response ratio. The white 1:1 diagonal line indicates no effects of nutrient addition on β diversity. Numbers in the parentheses are the number of sites. When a site has \(\overline{\Delta \alpha }\,\)= 0, ∆γ = 0, or ∆β = 0, it was not counted into any of the six scenarios as shown in the framework. The small points represent site-level \(\overline{\Delta \alpha }\) and ∆γ. The large open point and error bars are the estimated mean and 95% credible intervals for \(\overline{\Delta \alpha }\), ∆γ, and ∆β across all sites. See Table S5 for model fit and estimated overall means and 95% credible intervals for \(\overline{\Delta \alpha }\), ∆γ, and ∆β. Source data are provided as a Source Data file.
We also separated species into forb, graminoid, legume, and woody species to investigate scale-dependent diversity change within species groups. Nutrient addition led to the greatest reduction of α diversity for forb species and of γ diversity for woody species (Table S6). Similar to that of entire communities, nutrient addition decreased α and γ diversity by similar magnitudes and it had no effects on β diversity for graminoid species (∆β = 0.01; 95% credible interval: −0.04 to 0.05; Fig. 4B; Table S6) and legume species (∆β = 0.00; 95% credible interval: −0.17 to 0.18; Fig. 4C; Table S6). Overall, nutrient addition decreased α diversity 8% more than γ diversity for forb species, while it decreased γ diversity 11% more than α diversity for woody species (Table S6). However, nutrient addition also did not have significant effects on β diversity for forb species (∆β = 0.09; 95% credible interval: −0.02 to 0.19; Fig. 4C; Table S6). Nutrient addition caused a weak biotic homogenization for woody species (∆β = −0.14; 95% credible interval: −0.30 to 0.003; Table S6), this was primarily linked to loss of spatially restricted species (Fig. 4D).

A Forb, B graminoid, C legume, and D woody species. LRR: log response ratio. The white 1:1 diagonal line indicates no effects of nutrient addition on β diversity. Numbers in the parentheses are the number of sites. When a site has \(\overline{\Delta \alpha }\,\)= 0, ∆γ = 0, or ∆β = 0, it was not counted into any of the six scenarios as shown in the framework. The small points represent site-level \(\overline{\Delta \alpha }\) and ∆γ. The large open point and error bars are the estimated mean and 95% credible intervals for \(\overline{\Delta \alpha }\), ∆γ, and ∆β across all sites. See Table S6 for model fit and estimated overall means and 95% credible intervals for \(\overline{\Delta \alpha }\), ∆γ, and ∆β. Source data are provided as a Source Data file.
Robustness and limitations
We tested the robustness of our results by performing multiple sensitivity tests. We redid the analyses for the effects of nutrient addition on α, γ, and β diversity for the entire communities as well as for species groups using a subset of 14 sites that had data 14 years after nutrient additions began (Figs. S5–S7). We found that the overall effects were largely similar in these longer-term sites to that of 72 sites that had data four years after nutrient additions began. Because three spatial blocks may be limited in spatial extent for estimating effects on β diversity, we redid the analyses for the effects of nutrient addition on α, γ, and β diversity for the entire communities as well as for species groups using 11 sites that had five spatial blocks (Figs. S8–S10). Again, the overall effects were largely similar to that we found for the full analysis of the 72 sites with three blocks.
Despite our evidence for little change in β diversity with nutrient addition across the entire experiment, we recognize limitations of extrapolating these results to the landscape scale (e.g., >1 km × 1 km). This is because many ecological processes that directly and indirectly influence plant diversity can be very different at the landscape scales14,42. The scale at which we inferred changes in the number of spatially restricted and widespread species, by examining how many local communities they were lost from, is a relatively small spatial scale. Linking estimates of species’ geographic range size and other key traits with changes in plant diversity across larger spatial scales19,42 will deepen our understanding of the mechanisms of diversity change.
To summarize, we provide a comprehensive synthesis of the impact of nutrient addition on scale-dependent plant diversity change in grasslands by applying a framework to a globally distributed long-term experiment. The framework links changes in species richness across scales to that changes in the numbers of spatially restricted and widespread species. Overall, we found similar proportional plant diversity declines at local and larger spatial scales with nutrient addition, and little evidence for either biotic homogenization or differentiation within sites. These overall patterns were largely consistent for diversity metrics that incorporate relative species covers, across species groups, and over long time periods. This demonstrates that nutrient enrichment poses a potential threat to all plant species groups, including widespread and native species that often drive ecosystem functions and services.
Methods
Experimental setup
The experimental sites used in this study are part of the Nutrient Network (NutNet, Fig. S1 and Table S1). The experimental design includes a factorial manipulation of nutrients (N, P, and K) plus two fences to exclude herbivores, see ref. 39 for more details. For the analyses here, we used plots under two treatments: Ambient (Control) and fertilization by nitrogen, phosphate, and potassium together (i.e., NPK). Treatments were randomly assigned to 5 m × 5 m plots and were replicated in three or more blocks. A micronutrient mix consists of Fe (15%), S (14%), Mg (1.5%), Mn (2.5%), Cu (1%), Zn (1%), B (0.2%), and Mo (0.05%) was added once only at the start of the experiment (i.e., year 1) for the nutrient addition plots, but not in subsequent years to avoid toxicity. Nitrogen, phosphate, potassium were added annually before the growing season of each treatment year at most sites. Nitrogen was added as 10 g m−2 yr−1 time-release urea [(NH2)2CO], phosphate was added as 10 g m−2 yr−1 triple-super phosphate [Ca(H2PO4)2], while potassium was added as 10 g m−2 yr−1 sulfate [K2SO4].
Data were retrieved from the NutNet database in November 2023. We analyzed data from 72 sites where 1) nutrients were applied for at least four years; and 2) each site had at least three blocks. These sites are distributed across six continents and include a wide range of grassland types. See Fig. S1 and Table S1 for details of geolocation, grassland types, and experimental years used.
Sampling protocol
Scientists at NutNet sites followed standard sampling protocols39. Specifically, a 1 m × 1 m subplot within each plot was permanently marked for annual recording of plant species composition. Species cover (%) was estimated visually for individual species in the subplots; thus the total cover of living plants may sometimes exceed 100% for multilayer canopies. At most sites, cover was recorded once per year at peak biomass. At some sites with strong seasonality, cover was recorded twice per year to include a complete list of species. For those sites, the maximum cover for each species and total biomass were used in the analyses. When taxa could not be identified to the species level, they were aggregated at the genus level but referred to as “species” for simplicity.
Quantifying changes in α, γ, and β diversity
We measured α and γ diversity using species richness (i.e., number of species) because it is the most commonly examined diversity metric43. At each site, α diversity was estimated as the number of species in each permanent subplot (1 m × 1 m), and γ diversity as the total number of species occurring in three permanent subplots (for each treatment separately). To standardize sampling effort, for sites with more than three blocks, we selected the first three blocks according to the block number recorded by site PIs. The framework relies on Whittaker’s multiplicative β diversity partition, and it quantifies β diversity using the effective number of communities12. As such, if all subplots share the same species, then β diversity would equal to one. In contrast, if each subplot has unique species, then β diversity would equal to three. We calculated ∆α as the richness difference in local communities (subplots) and ∆γ as the difference in the sum of the subplots under nutrient addition relative to that of control treatment on the log scale. That is, ∆α = log(αNPK/αControl) and ∆γ = log(γNPK/γControl). We calculated ∆β as ∆γ minus \(\overline{\Delta \alpha }\), where\(\,\overline{\Delta \alpha }\) is the average of ∆α over three blocks. A decrease in ∆β indicates nutrient addition causes species composition to be more similar among three subplots than that among control subplots. Because sites are not evenly distributed around the world, many sites are aggregated in North America, we checked spatial autocorrelation of diversity change under nutrient addition using Moran’s I44. We found that\(\,\overline{\Delta \alpha }\), ∆γ, and ∆β did not appear to be more similar for sites that are closer to each other (Table S2).
We fitted multilevel (also referred as mixed effects or hierarchical) models for ∆α, ∆γ, and ∆β (as the response variable; all on the log scale) separately. We included random intercept for each site, model was coded as: richness change ~ 1 + (1 |sites) to estimate site-level variation. We used Bayesian analysis because it yields full posterior distributions of parameters rather than point estimates and p-values, which provides a deeper understanding of the uncertainty and variability in the results45. Models described above were fitted using the Hamiltonian Monte Carlo (HMC) sampler in Stan and coded using the package ‘brms’ (version 2.21.0) in R (version 4.4.1)46,47. Models were fitted without explicitly specifying priors, allowing brms to assign its default priors. Models were fitted with 6 chains and 3000 iterations (1000 iterations for warm up). Visual inspection of the HMC chains and Rhat summaries showed model convergence (all Rhats <1.03; Tables S3, S5 and S6). We visually checked posterior predictive plots to determine how well models can reproduce the data (Fig. S2).
To examine whether diversity changes were sensitive to species relative covers, we redid the above analyses (i.e., based on species richness) using Shannon diversity and Simpson diversity (both converted to effective numbers)48 (Fig. S4). Species richness is most sensitive to rare species, followed by Shannon diversity, and Simpson diversity is more sensitive to the numbers of relatively abundant species. We calculated the exponential of Shannon diversity and the inverse form of Simpson diversity using the R package vegan (version 2.6-6.1)49. These three diversity metrics equal to diversity with order q = {0, 1, 2}, where increasing q decreases the influence of rare species, and Dq = \({\left({\sum }_{i=1}^{s}{p}_{i}^{q}\right)}^{1/\left(1-q\right)}\), where p is the relative cover of species i, s is the total number of species. These diversity metrics are also referred to as Hill numbers48,50.
Site covariates
We investigated whether the effects of nutrient addition on \(\overline{\Delta \alpha }\), γ, and β diversity based on species richness were mediated by site characteristics. We included site characteristics that have been shown in previous literature to influence ∆α, ∆γ, and ∆β in grasslands: site species pool, site productivity, drought intensity, and grazing intensity24,25,34,40. We quantified drought intensity as the sum of annual evapotranspiration/precipitation, and averaged it from year 0 to 4 at each site. Precipitation and potential evapotranspiration used were downloaded from https://crudata.uea.ac.uk/cru/data/hrg/cru_ts_4.07/. We quantified the site species pool as the total number of species and site productivity as the average aboveground biomass from year 0 to 4 under the control treatment in the three blocks at each site. Aboveground biomass was harvested within two 1 × 0.1 m strips (in total 0.2 m2), strips were moved from year to year to avoid resampling the same location. For subshrubs and shrubs occurring within strips, we collected all leaves and current year’s woody growth. All biomass was dried at 60 °C (to constant mass) before weighing to the nearest 0.01 g. We used published methods to quantify an integrated grazing intensity metric from vertebrate herbivores at each site. Specifically, herbivore species (>2 kg) that consume grassland biomass were documented at each site by site PIs, and each species was assigned an importance value from 1 (present but low impact and frequency) to 5 (high impact and frequency). An index value was calculated for each site as the sum of herbivore importance values for all herbivores following refs. 51,52. We also investigated relationships between change in diversity and distance among blocks, because species composition may become less similar as the distance between sampled communities increases. The average pairwise distance among the three blocks within sites ranged from 23.04 to 12538.09 m, with a mean of 513.01 m and a median of 118.7 m across 54 sites that have geolocation data for each block. We first calculated three Euclidean distances between pairs of blocks, we then used the mean of these pairwise distances as the average distance among blocks. We used the average distance among blocks instead of area, because blocks are arranged in parallel at some sites. We fitted linear regression models with\(\,\overline{\Delta \alpha }\), ∆γ, and ∆β as the response variable separately, and each of the site characteristics was used as a predictor variable.
Species groups
We then investigated the effects of nutrient addition on α, γ, and β diversity within groups of species with similar characteristics following the method for changes in α, γ, and β diversity in the entire communities. We eliminated sites where no species occurred in control, nutrient addition, or both plots for a particular group because the value of the log (0) is undefined. We ran the analyses separately for native and non-native species. Native and non-native species were classified by site PIs. Then, we investigated effects of nutrient addition on species richness for different life forms including forb, graminoid, legume, and woody species because previous studies have shown that different life forms may show distinct responses to nutrient addition6,11,53.
Sensitivity test
We tested whether effects of nutrient addition on species richness across spatial scales depend on experimental duration because a few single-site experiments have shown that the effects of nutrient additions on changes in diversity, especially β diversity, may take several years to emerge29,31. To that end, we used a subset of 14 sites that had data 14 years after nutrient additions began. Also, because three blocks may be limited in spatial extent, we tested whether combining more blocks to create the γ scale would alter our results. We redid the analyses using data from 11 sites that had five spatial blocks.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The species cover and species richness data, site abiotic and biotic environmental data used and generated in this study have been deposited in the Figshare database and are publicly available (https://doi.org/10.6084/m9.figshare.26412295.v4). The NutNet data are publicly available on the Environmental Data Initiative (EDI) (https://portal.edirepository.org/nis/advancedSearch.jsp). Source data are provided with this paper.
Code availability
The R codes used to produce results in this study have been deposited in the GitHub (https://github.com/chqq365/plant-diversity-and-biotic-homogenization.git) and archived through Zenodo (https://doi.org/10.5281/zenodo.14902812).
References
Sardans, J. & Peñuelas, J. Potassium: a neglected nutrient in global change. Glob. Ecol. Biogeogr. 24, 261–275 (2015).
Galloway, J. N., Bleeker, A. & Erisman, J. W. The human creation and use of reactive nitrogen: a global and regional perspective. Annu. Rev. Environ. Resour. 46, 255–288 (2021).
Yuan, Z. et al. Human perturbation of the global phosphorus cycle: changes and consequences. Environ. Sci. Technol. 52, 2438–2450 (2018).
Bobbink, R. et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol. Appl. 20, 30–59 (2010).
Seabloom, E. W. et al. Increasing effects of chronic nutrient enrichment on plant diversity loss and ecosystem productivity over time. Ecology 102, 1–11 (2021).
Suding, K. N. et al. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proc. Natl Acad. Sci. USA 102, 4387–4392 (2005).
Yang, Z., Hautier, Y., Borer, E. T., Zhang, C. & Du, G. Abundance- and functional-based mechanisms of plant diversity loss with fertilization in the presence and absence of herbivores. Oecologia 179, 261–270 (2015).
Zhang, P. et al. Space resource utilization of dominant species integrates abundance- and functional-based processes for better predictions of plant diversity dynamics. Oikos 2023, e09519 (2023).
MacDougall, A. S. et al. The neolithic plant invasion hypothesis: the role of preadaptation and disturbance in grassland invasion. N. Phytol. 220, 94–103 (2018).
Seabloom, E. W. et al. Plant species’ origin predicts dominance and response to nutrient enrichment and herbivores in global grasslands. Nat. Commun. 6, 7710 (2015).
Tognetti, P. M. et al. Negative effects of nitrogen override positive effects of phosphorus on grassland legumes worldwide. Proc. Natl Acad. Sci. USA 118, e2023718118 (2021).
Blowes, S. A. et al. Synthesis reveals approximately balanced biotic differentiation and homogenization. Sci. Adv. 10, eadj9395 (2024).
Keil, P. et al. Spatial scaling of extinction rates: theory and data reveal nonlinearity and a major upscaling and downscaling challenge. Glob. Ecol. Biogeogr. 27, 2–13 (2018).
Chase, J. M. et al. Species richness change across spatial scales. Oikos 128, 1079–1091 (2019).
Daleo, P. et al. Environmental heterogeneity modulates the effect of plant diversity on the spatial variability of grassland biomass. Nat. Commun. 14, 1809 (2023).
Le Provost, G. et al. The supply of multiple ecosystem services requires biodiversity across spatial scales. Nat. Ecol. Evol. 7, 236–249 (2022).
Daru, B. H. et al. Widespread homogenization of plant communities in the Anthropocene. Nat. Commun. 12, 6983 (2021).
Gossner, M. M. et al. Land-use intensification causes multitrophic homogenization of grassland communities. Nature 540, 266–269 (2016).
Staude, I. R. et al. Replacements of small- by large-ranged species scale up to diversity loss in Europe’s temperate forest biome. Nat. Ecol. Evol. 4, 802–808 (2020).
McKinney, M. L. & Lockwood, J. L. Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol. Evol. 14, 450–453 (1999).
Chen, Q. et al. Long‐term cross‐scale comparison of grazing and mowing on plant diversity and community composition in a salt‐marsh system. J. Ecol. 109, 3737–3747 (2021).
Huston, M. A general hypothesis of species diversity. Am. Nat. 113, 81–101 (1979).
Tilman, D. Plant Strategies and the Dynamics and Structure of Plant Communities (Princeton University Press, 1988).
Seabloom, E. W. et al. Species loss due to nutrient addition increases with spatial scale in global grasslands. Ecol. Lett. 24, 2100–2112 (2021).
Chalcraft, D. R. et al. Scale-dependent responses of plant biodiversity to nitrogen enrichment. Ecology 89, 2165–2171 (2008).
Lu, W. et al. Similar effects of seasonal nitrogen enrichment on plant species–area relationship in a temperate grassland. Ecol. Indic. 158, 111430 (2024).
Chen, Q. et al. Nutrients and herbivores impact grassland stability across spatial scales through different pathways. Glob. Change Biol. 28, 2678–2688 (2022).
Zhou, X., Liu, X., Zhang, P., Guo, Z. & Du, G. Increased community compositional dissimilarity alleviates species loss following nutrient enrichment at large spatial scales. J. Plant Ecol. 12, 376–386 (2019).
DeSiervo, M. H., Sullivan, L. L., Kahan, L. M., Seabloom, E. W. & Shoemaker, L. G. Disturbance alters transience but nutrients determine equilibria during grassland succession with multiple global change drivers. Ecol. Lett. 26, 1132–1144 (2023).
Eskelinen, A. & Harrison, S. Erosion of beta diversity under interacting global change impacts in a semi-arid grassland. J. Ecol. 103, 397–407 (2015).
Inouye, R. S. & Tilman, D. Convergence and divergence of old-field vegetation after 11 yr of nitrogen addition. Ecology 76, 1872–1887 (1995).
Zhang, Y. et al. Nitrogen addition does not reduce the role of spatial asynchrony in stabilising grassland communities. Ecol. Lett. 22, 563–571 (2019).
Foster, B. L. et al. Seed availability constrains plant species sorting along a soil fertility gradient. J. Ecol. 99, 473–481 (2011).
Koerner, S. E. et al. Nutrient additions cause divergence of tallgrass prairie plant communities resulting in loss of ecosystem stability. J. Ecol. 104, 1478–1487 (2016).
Lan, Z. et al. Testing the scaling effects and mechanisms of N-induced biodiversity loss: evidence from a decade-long grassland experiment. J. Ecol. 103, 750–760 (2015).
Xu, Z. et al. Antithetical effects of nitrogen and water availability on community similarity of semiarid grasslands: evidence from a nine-year manipulation experiment. Plant Soil 397, 357–369 (2015).
Yang, X. et al. Resource addition drives taxonomic divergence and phylogenetic convergence of plant communities. J. Ecol. 107, 2121–2132 (2019).
Whittaker, R. H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol. Monogr. 30, 279–338 (1960).
Borer, E. T. et al. Finding generality in ecology: a model for globally distributed experiments. Methods Ecol. Evol. 5, 65–73 (2014).
Chase, J. M. & Leibold, M. A. Spatial scale dictates the productivity-biodiversity relationship. Nature 416, 427–430 (2002).
Seabloom, E. W. et al. Predicting invasion in grassland ecosystems: is exotic dominance the real embarrassment of richness? Glob. Change Biol. 19, 3677–3687 (2013).
Staude, I. R. et al. Directional turnover towards larger-ranged plants over time and across habitats. Ecol. Lett. 25, 466–482 (2022).
Chase, J. M. et al. Embracing scale‐dependence to achieve a deeper understanding of biodiversity and its change across communities. Ecol. Lett. 21, 1737–1751 (2018).
Getis, A. A history of the concept of spatial autocorrelation: a geographer’s perspective. Geogr. Anal. 40, 297–309 (2008).
Kruschke, J. K. Bayesian estimation supersedes the t test. J. Exp. Psychol. Gen. 142, 573–603 (2013).
Bürkner, P.-C. Advanced Bayesian multilevel modeling with the R Package brms. R. J. 10, 395 (2018).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2022).
Jost, L. Entropy and diversity. Oikos 113, 363–375 (2006).
Oksanen, A. J. et al. Package ‘ vegan’. (2019).
Hill, M. O. Diversity and evenness: a unifying notation and its consequences. Ecology 54, 427–432 (1973).
Borer, E. T. et al. Nutrients cause grassland biomass to outpace herbivory. Nat. Commun. 11, 1–8 (2020).
Anderson, T. M. et al. Herbivory and eutrophication mediate grassland plant nutrient responses across a global climatic gradient. Ecology 99, 822–831 (2018).
Van Sundert, K. et al. Towards comparable assessment of the soil nutrient status across scales—review and development of nutrient metrics. Glob. Change Biol. 26, 392–409 (2020).
Acknowledgements
This work was generated using data from the Nutrient Network (http://www.nutnet.org) experiment, funded at the site-scale by individual researchers. We thank the Minnesota Supercomputer Institute for hosting project data and the Institute on the Environment for hosting Network meetings. We thank researchers from the NutNet who contributed data to our analysis, Table S7 lists these researchers. We thank Shuli Niu for constructive suggestions to improve our manuscript. Nitrogen fertilizer was donated to NutNet by Crop Production Services, Loveland, CO. Funding: German Center for Integrative diversity Research (iDiv) Halle-Jena-Leipzig (DFG FZT 118-202548816) for the contribution of J.M.C., S.A.B., E.L., Q.C., W.S.H., and other iDiv co-authors. ERC Advanced Grant (MetaChange) to J.M.C. for the contribution of J.M.C., S.A.B. Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union or the European Research Council. Neither the European Union nor the granting authority can be held responsible for them. National Science Foundation grant NSF-DEB-1042132 (E.T.B., E.W.S.; for NutNet coordination and data management) National Science Foundation grant NSF-DEB-1234162 (E.T.B., E.W.S.; for Long-Term Ecological Research at Cedar Creek). National Science Foundation grant NSFDEB-1831944 (E.T.B., E.W.S.; for Long-Term Ecological Research at Cedar Creek), and the Institute on the Environment (DG-0001-13; E.T.B., E.W.S.).
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
Q.C., J.M.C., S.A.B., E.L., and W.S.H. conceived the study; Q.C., J.M.C., and S.A.B. developed the methodology; Q.C. analyzed data with contribution from S.A.B.; W.S.H., E.T.B., A.M., J.P.M., J.D.B., P.M.T., E.W.S., P.D., S.A.P., C.R., P.B.A., I.D., G.W., C.S., G.F.V., A.C.R., G.M.W., Y.H., C.E., Y.N., P.L.P., A.E., D.S.G., H.O., C.D., M.W.C., S.H., N.E., J.C., R.V., J.W.M., M.T., S.B., M.C.C., M.B., J.M.H.K., C.R.D., N.H., A.J., P.M., B.B.O., and L.L. contributed data; Q.C. visualized the results; Q.C. wrote the original draft of the manuscript; Q.C., S.A.B., W.S.H., E.L., E.T.B., A.M., J.P.M., J.D.B., P.M.T., E.W.S., P.D., S.A.P., C.R., P.B.A., I.D., G.W., C.S., G.F.V., A.C.R., G.M.W., Y.H., C.E., E.H., Y.N., P.L.P., A.E., D.S.G., H.O., C.D., M.W.C., S.H., N.E., J.C., R.V., J.W.M., M.T., S.B., M.C.C., M.B., J.M.H.K., C.R.D., N.H., A.J., P.M., B.B.O., L.L., and J.M.C. reviewed and edited the manuscript. See Table S8 for more details for the contribution of each author.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewers for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, Q., Blowes, S.A., Harpole, W.S. et al. Local nutrient addition drives plant diversity losses but not biotic homogenization in global grasslands. Nat Commun 16, 4903 (2025). https://doi.org/10.1038/s41467-025-59166-7
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-59166-7
