
Abstract
Northern ecosystems (≥ 30° N) have been accumulating vegetation biomass carbon in recent decades, but increasing droughts and wildfires threaten this carbon sink. Here, we analyse annual changes in live vegetation biomass in northern ecosystems using low-frequency microwave satellite observations at 25 km spatial resolution from 2010 to 2022. We find that live biomass carbon stocks have undergone a reversal from a positive to a negative trend during the study period with 2016 marking the turning point. During 2016–2022, live biomass carbon stocks decreased at a rate of \({-{{\mathbf{0.20}}}}_{-{{\mathbf{0.26}}}}^{-{{\mathbf{0.11}}}}\) PgC yr−1 across northern ecosystems, primarily in temperate biomes (\({-{{\mathbf{0.26}}}}_{-{{\mathbf{0.33}}}}^{-{{\mathbf{0.17}}}}\) PgC yr−1). The annual mean gross loss of 4% of live biomass carbon in this region during 2016-2022 reflects high interannual variability, with significant losses associated with droughts and a further drop of \({-{{\mathbf{0.60}}}}_{-{{\mathbf{0.75}}}}^{-{{\mathbf{0.47}}}}\) PgC in the very dry year of 2022. Our findings highlight the vulnerability of live biomass carbon stocks to emerging climate-induced disturbances in northern ecosystems, challenging the sustainability of the current large terrestrial carbon sink in this key region for the global carbon balance.
Similar content being viewed by others
Introduction
Northern ecosystems (north of 30°N) hold about 41% of the world’s forest area1, and are critical for mitigating global warming2,3, contributing an estimated 1.4-2.0 PgC annually to the global terrestrial carbon sink4. In recent decades, rising CO2 levels and global warming have been reported to increase mid- and high-latitude vegetation productivity by lengthening the growing season5, which has resulted in a ‘greening’ trend as observed by optical satellite systems6. Concordantly, multiple lines of evidence from forest inventories7,8 and satellite-based studies9,10,11 suggest that biomass carbon has increased in northern ecosystems in recent decades.
Climate hazards, however, may have affected the biomass carbon budget of northern ecosystems as a result of increased wildfire frequency12, extensive drought-induced tree mortality13,14 and unprecedented scale of insect outbreaks15. Previous studies have indicated that threats to forest carbon sinks have increased in North America16,17,18 and Europe19,20,21 over recent decades due to tree mortality caused by very high temperatures and droughts. Wildfires, a main driver of Siberian and Canadian boreal forest disturbances, have been reported to cause large carbon emissions during recent extreme fire years12,22 and have even led to parts of Siberia turning into a carbon source13,23. Compound droughts-heat waves (typically characterized by low soil moisture and high air temperatures) and wildfires have affected the northern Hemisphere in recent years, such as in 201519,24, 201820, 202112,25 and 202221,26.
Improving our knowledge of the interannual variability and trends in biomass carbon is crucial for a timely understanding of how climate change and human management affect the terrestrial carbon balance. Even in regions and countries with dense forest inventory sampling, such as the United States and Europe, the spatial sampling design remains insufficient to assess regional scale changes27. Further, the practice of conducting inventory revisits in multi-year cycles omits unmanaged forests and relies on simple methods such as static regrowth curves in countries using the gain-loss inventory method, which may ultimately not capture recent climate impacts28. Large uncertainties have also been reported in the quantification of carbon fluxes and sinks, for which biomass carbon change represents a key component, whether estimated from inventories8, dynamic global vegetation models (DGVMs)29, or atmospheric inversion models30. Alternatively, satellite data from light detection and ranging (LIDAR), optical and synthetic aperture radar (SAR) have been combined to map biomass carbon31,32 or track its annual changes over large regions11,27. However, due to the lengthy process of acquiring, processing, and formatting new data, the use of methods based on multi-source remote sensing technologies poses challenges in achieving consistent and continuous biomass carbon assessments, particularly with respect to timeliness. Such significant delays impede a timely understanding of biomass carbon balance of northern ecosystems in the context of the recent increased frequency in compound droughts and heatwaves.
Here, we estimated the wall-to-wall annual changes in total (the sum of above- and below-ground) live biomass carbon in northern ecosystems from 2010 to 2022 using L-band (1.4 GHz) vegetation optical depth (L-VOD) data from the Soil Moisture and Ocean Salinity (SMOS) satellite. L-VOD relies only on passive microwave observations and therefore it can be rapidly updated to probe the biomass components of living woody vegetation33,34. Moreover, the high penetration of low-frequency radiation through the canopy makes L-VOD less prone to saturation, enabling its extensive use in tracking interannual changes in global vegetation carbon stocks10,13,35,36,37. Spatially explicit aboveground biomass carbon (AGC) at 25 km spatial resolution was first retrieved from L-VOD (Methods). Total live biomass carbon was then derived from AGC using the root-to-shoot ratio of below- and above-ground biomass from the IPCC Guidelines for National Greenhouse Gas Inventories38. From the L-VOD-derived biomass carbon estimates, we analyse the changes in live biomass carbon at the regional scale and identify factors that drive the spatial variability in biomass carbon density changes across different biomes. The L-VOD-derived biomass carbon density was also compared with the DGVM simulations from TRENDY v12 project39, particularly focusing on recent trends.
Results
Changes in total live biomass carbon
For the years 2010–2022, the L-VOD-derived live biomass carbon stocks showed an average net gain in the northern ecosystems at a rate of \({+0.25}_{+0.25}^{+0.26}\) PgC yr−1 (the range represents the uncertainties, derived from 12 calibrations of L-VOD against three benchmark maps; Methods). Among ecosystems, boreal regions had a net increase of \({+0.15}_{+0.16}^{+0.15}\) PgC yr−1 in live biomass carbon, while northern temperate regions remained nearly neutral with a net change of \({+0.03}_{+0.02}^{+0.04}\) PgC yr−1 (Fig. 1d, e). Our estimates, updated to 2022, are in line with previous global analyses of L-VOD until 201911, when comparing changes in biomass carbon over the same time period and under the same biome delineations (Supplementary Fig. 1). In addition, a comparison with the national inventories from the Food and Agriculture Organization (FAO) Global Forest Resources Assessment indicates that the L-VOD estimated biomass carbon stocks closely match those reported by countries (Supplementary Fig. 2). Moreover, the biomass carbon changes in managed forests reported by L-VOD show a strong spatial correlation with estimates from the national forest inventories across countries (R2 = 0.58, P < 0.01).

a, b Per-pixel linear trend in biomass carbon for the periods (a) 2010–2016 and (b) 2016–2022 (n = 49,914). c Annual variations in live biomass carbon stocks, expressed as the difference from 2010 values in northern ecosystems (n = 49,914) and in northern ecosystems of partly burned (n = 27,451) and unburned (n = 22,463) regions. The ‘unburned’ areas are pixels where the annual fire area fraction was consistently < 1% each year from 2005 to 2022 within a 25 km grid, while all other pixels are considered partly burned areas13. d, e Corresponding changes in biomass carbon are shown for two main biomes in northern ecosystems (d) Temperate (n = 26,880) and (e) Boreal (n = 18,721). The ranges represented by shading around the line denote the minimum and maximum of biomass carbon changes estimated by 12 calibrations (see “Method” and Supplementary Table 1). The black line in (a, b) represents the delineation of the boreal biome, south of which (but still north of 30° N) is the temperate biome (Supplementary Fig. 1a), whereas the black dots denote areas with P < 0.05. The turning point shown by the red arrow in c was determined by a piecewise linear regression (Methods). Source data are provided as a Source Data file.
The temporal variations in annual biomass carbon shows a shift from a positive to a negative trend after 2016 (Fig. 1c). The turning point occurring around 2016 (P < 0.05) detected by a piecewise linear regression (Methods) is confirmed by the analysis of two independent datasets derived from multiple satellites available up to 2019: one from a high-frequency C-band VOD product40 and the other from global annual biomass maps11 with native spatial resolutions of 25 km and 10 km, respectively. Both sources indicated that live biomass carbon in northern ecosystems peaked in 2016, representing the highest levels observed since 2000 (Supplementary Fig. 3). Spatially, the transition to biomass loss revealed by L-VOD predominantly occurred in the south-central United States, Kazakhstan, and western Russia, while both loss and gain were observed in Europe (Fig. 1a, b). L-VOD-derived results indicated that the largest decline in biomass carbon during the 2010-2022 period occurred in 2022, with a net change of \({-0.67}_{-0.69}^{-0.41}\) PgC across northern ecosystems, of which \({-0.60}_{-0.75}^{-0.47}\) PgC occurred in temperate regions and \({-0.14}_{-0.27}^{-0.13}\) PgC in boreal regions (Fig. 1c, d, e).
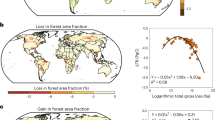
Between 2016 and 2022, net L-VOD-based biomass carbon estimates decreased at a rate of \({-0.20}_{-0.26}^{-0.11}\) PgC yr−1 across the entire northern ecosystems. This net decrease is the result of gross gains (\({+2.76}_{+2.54}^{+3.12}\) PgC yr−1) being offset by larger average gross losses (\({-2.96}_{-3.38}^{-2.65}\) PgC yr−1) (Supplementary Fig. 4). Over this period, 53.84% of the study area showed a net carbon loss in live biomass, which is reflected by the imbalance between gross gains and gross losses along the latitudinal band from 30° to 52° N (Fig. 2b, c). The majority of net losses occurred in temperate ecosystems (\({-0.26}_{-0.33}^{-0.17}\) PgC yr−1) including North America (\({-0.15}_{-0.18}^{-0.10}\) PgC yr−1) and Eurasia (\({-0.11}_{-0.16}^{-0.07}\) PgC yr−1), while boreal biomes in these two regions experienced a small net gain (\({+0.07}_{+0.07}^{+0.08}\) PgC yr−1) and loss (\({-0.03}_{-0.04}^{-0.02}\) PgC yr−1) of carbon, respectively (Fig. 2b). Notably, a gross live biomass carbon loss of \({-1.50}_{-1.77}^{-1.26}\) PgC yr−1 occurred in temperate biomes, which was partially offset by a gross gain of \({+1.24}_{+1.09}^{+1.44}\) PgC yr−1. This gross annual loss represents about 4% of the total live biomass carbon in northern temperate regions (\({35.05}_{28.65}^{42.79}\) PgC in 2016) (Supplementary Fig. 5), with hotspots in the south-central United States, southwestern Europe, and western Russia (Supplementary Fig. 4).

a Yearly net changes in forest-loss rates from the Global Forest Watch41 (n = 49,914). b Spatial patterns of net changes in L-VOD-derived biomass carbon density. c Latitudinal sums of gross gains and losses in L-VOD biomass estimates from 2016 to 2022, derived from per-pixel gain and loss maps by cumulating positive and negative changes for consecutive years. The shaded ranges around the line denote uncertainty based on the minimum and maximum L-VOD-derived biomass carbon values (Methods). d Yearly net changes in burned area estimated using moderate-resolution imaging spectroradiometer (MODIS) data. e Mean anomalies (expressed as z-score) in ERA5 soil moisture for 2016-2022 relative to 2010-2022. f Mean Standardized Precipitation Evapotranspiration Index (SPEI) at 12-month timescale for 2016–2022. Source data are provided as a Source Data file.
Annual gross losses in boreal biomes amounted to about 2% (\({-1.29}_{-1.47}^{-1.22}\) PgC yr−1) of the total live biomass carbon stock (\({54.26}_{48.88}^{61.69}\) PgC in 2016), with higher losses in the Siberian region, where the spatial loss patterns are quite consistent with burned areas (Fig. 2b, d). Fan et al.13 reported that forest disturbances, attributed to fires and droughts in recent years, have reduced the biomass accumulation in Siberia, bringing the biomass carbon balance of the region close to neutrality. Gross gains in boreal regions were \({+1.32}_{+1.26}^{+1.46}\) PgC yr−1, mainly located in northwestern Canada, Alaska, and northern Finland (Supplementary Fig. 4a). The fact that the reported values of gross fluxes were much larger than those of net fluxes suggests that changes in live biomass carbon stocks were highly dynamic during the study period, especially for the temperate biomes11.
Drivers influencing the recent decline in live biomass carbon over northern ecosystems
The determinants of the spatio-temporal variations in live biomass carbon changes are multifaceted and can be related to drought, mortality and fire events in northern ecosystems. Spatially, regions showing a decrease in live biomass from 2016 to 2022 closely matched those of forest area loss reported by the Global Forest Watch (GFW)41 and the areas of fire disturbance monitored by active fire data42 (Fig. 2a, b, d). We separately calculated the total biomass losses in relation to forest area loss and burned area for each subregion of the northern ecosystems defined in the IPCC Sixth Assessment Report (Supplementary Fig. 6). The extent of biomass losses increases with burned area (R2 = 0.86, P < 0.01) and forest loss area (R2 = 0.49, P < 0.01) within each 25 km L-VOD grid cell. The high R2 for the relationship between biomass carbon losses and burned area indicates that wildfires are a significant disturbance factor reducing live biomass accumulation13,43. To further explore the factors behind the spatial variability of net changes in biomass carbon across different ecosystems during 2016-2022, we employed an interpretable machine learning (RF-SHAP) framework to quantify the relative importance of more than 15 factors separately for temperate and boreal biomes (Methods). The factors we considered include climate, vegetation, edaphic, and anthropogenic disturbances (Supplementary Table 2). Altogether, these factors explained 51% and 61% of the spatial variation in live biomass carbon changes observed in temperate and boreal biomes, respectively (Fig. 3). Moreover, the RF models presented good predictive skills in reproducing the observed spatial patterns of biomass carbon changes (Supplementary Fig. 7).

a Relative importance of the variables explaining the spatial variability of biomass carbon changes in temperate biomes. b–e Partial dependence plots of the top four variables, namely, tree cover (b), surface solar radiation changes (ΔRadiation) (c), soil nitrogen (d) and soil moisture changes (ΔSM) (e). The Y-axis represents the SHAP value for the corresponding predictor (X-axis), where positive values contribute to higher predicted Δbiomass values, and negative values to lower ones. The red line indicates the median SHAP values across 50 bins of x-axis variables, plotted only for bins containing more than 20 samples explained at least 10 times by SHAP. The partial dependence plots illustrate how individual variables contribute to the predicted Δbiomass, capturing both their direct effects and interactions with other variables (Methods). f Similar to (a), but for boreal biomes. g–j Partial dependence plots of the top four explanatory variables in boreal biomes, namely, ΔRadiation (g), non-fire-induced forest losses changes (h), forest loss due to stand-replacing wildfires (i) and vapor pressure deficit changes (ΔVPD) ( j). Details on these variables are given in Supplementary Table 2. Source data are provided as a Source Data file.
The most important climate-related variable affecting live biomass carbon change (Δbiomass) in both temperate and boreal biomes was the change in surface solar radiation (ΔRadiation), accounting for 13% and 19% of the explained variability, respectively (Fig. 3a,f). However, partial dependence plots revealed that the effect of ΔRadiation on Δbiomass differed between the two biomes, with Δbiomass decreasing as ΔRadiation increased in temperate areas, while a positive relationship was found in boreal areas (Fig. 3c, g). The negative relationship between Δbiomass and Δradiation in temperate regions may be associated with reduced soil moisture (SM) and increased vapor pressure deficit (VPD) as radiation increases (Supplementary Fig. 8a, b). The Spearman rank correlation between L-VOD biomass estimates and SM at low-mid latitudes highlights the negative sensitivity of live biomass carbon stock changes to soil moisture anomalies in this area (Supplementary Fig. 9f). ΔSM contributed 10% of the explained variability, ranking among the higher climate factors, with soil moisture deficits (i.e., negative ΔSM) linked to negative biomass carbon changes (Fig. 3a, e). The most significant factor positively influencing Δbiomass in temperate biome was tree cover with an average contribution of 18%, followed by soil nitrogen content at 11% (Fig. 3a, b, d). For the boreal biome, ΔRadiation is ranked as the most important factor affecting Δbiomass, which may reflect the limitation of solar energy on biomass accumulation, consistent with its role in constraining vegetation photosynthesis (gross primary production) for this region22. The U-shaped relationship, with the lowest point around ΔRadiation ≈ 0 J/m², is likely influenced by interactions with other factors, particularly disturbances related to forest area (Supplementary Fig. 8c, d). The second-most influential climatic factor was ΔVPD, contributing 8%, with increasing VPD associated with decreasing biomass (Fig. 3f, j), reflecting the fact that rising VPD is associated with tree mortality, reduced growth44, and fire activity27. Notably, while forest loss due to stand-replacing wildfires ranked high (9%), non-fire forest disturbances such as logging and drought-induced mortality had a comparatively larger impact on Δbiomass in the boreal areas, accounting for 12% (Fig. 3f, h).
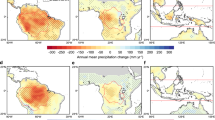
For the years 2016-2022, the northern lands experienced extensive drought events, as indicated by negative anomalies in soil moisture and SPEI values alongside elevated VPD, especially in recent years45, such as the 2022 global widespread heatwaves (Fig. 4a, e, f). Over this period, the net decrease in L-VOD live biomass carbon in the low-mid latitudes (30°N ~ 55°N) coincided with the negative trend in soil moisture (Fig. 4d). Moreover, these conditions of extreme drought were associated with large wildfires, which were particularly pronounced at high latitudes (> 60°N) (Fig. 4c). Correspondingly, the area of forest loss per year in the boreal biome reported by GFW showed a clear positive trend at a rate of 1.01 ×106 ha/year (Supplementary Fig. 10). Notably, the live biomass carbon changes revealed by L-VOD showed a decreasing trend around 60°N – 65°N during 2016-2022, which coincided with the highest average annual burned area recorded in this region over the same period (Fig. 4h). However, the TRENDY DGVMs with rapid update capabilities did not mirror this trend, partly due to limitations in their prognostic fire modules in realistically reflecting wildfire occurrence and intensity as well as the associated loss of live biomass. Yet, the three DGVMs models, JULES, OCN, and ORCHIDEE, which used prescribed burned areas from the ESA Climate Change Initiative (CCI-Fires) (Methods), revealed better consistency with L-VOD estimates compared to their prognostic fire versions (Supplementary Text 4).

a–d Anomalies for 2010–2022 in ERA5 soil moisture (a) L-VOD-derived live biomass carbon density (b) and MODIS burned area (c). d Net changes in L-VOD-derived live biomass carbon (summed by latitude) from 2016 to 2022 are shown for areas with negative (red) and positive (green) soil moisture trends (P < 0.05) over the same period. The background gray area plot represents the average SPEI values for the corresponding pixels. e–g Anomalies for 2010–2022 in SPEI values (e), VPD (f), and TRENDY DGVMs-simulated biomass carbon density (g). h Linear trends (P < 0.05) in live biomass carbon density observed by L-VOD and modeled by TRENDY DGVMs during 2016–2022. The background bars in h represent the annual average burned area from 2016 to 2022 for the pixels showing trends (P < 0.05), while the shaded ranges around the line indicate uncertainty (Methods). Source data are provided as a Source Data file.
During the summer drought in Siberia in 2021, when record wildfire emissions were reported12, the area of forest loss due to wildfires was 7.58×106 ha as monitored by stand-replacing wildfires data46. This event is consistent with a decline in live biomass carbon of \({-0.12}_{-0.14}^{-0.11}\) PgC that year, which suggests that disturbances in recent years have further reduced live biomass carbon stocks in Siberian forests, consistent with Fan et al.13. The severe drought of summer 2022 had swept widely across northern temperate regions (Supplementary Fig. 11), such as Europe and the contiguous United States45, coinciding with the pronounced decrease in biomass carbon of \({-0.25}_{-0.28}^{-0.25}\) PgC and \({-0.18}_{-0.21}^{-0.11}\) PgC observed in these two regions during that year. Latitudinally, the decrease was particularly strong at low to mid-latitudes (30°-50°N), where the biomass carbon anomalies showed a clear congruence with the ERA547 soil moisture anomalies and SPEI (Fig. 4a, b, e). The L-VOD-derived live biomass carbon decrease in this latitudinal band was generally reproduced by the TRENDY DGVMs (Fig. 4g) and the high frequency X-VOD (Supplementary Fig. 9a), suggesting that the compound effects of droughts and heatwaves in 2022 severely suppressed the vegetation carbon uptake in northern temperate biomes21,26.
Live biomass carbon losses from forest loss and degradation during 2016-2022
Live biomass carbon losses in northern ecosystems have been attributed to increased forest fires13,43, insect outbreaks48, logging27 and tree mortality due to climatic disturbances such as windstorms and drought18. Here, we separated the contributions of forest area loss and ‘degradation’ to live biomass carbon losses using the method of Harris et al.49 and Qin et al.36. Since forest loss and degradation cannot be explicitly separated as two distinct processes within a 25 km grid cell, we simplified our definition by categorizing all mechanisms that do not result in forest area loss as degradation13. To do so, we subtracted live biomass carbon losses due to forest loss events, identified by Landsat data41 at a spatial resolution of 30 m, from the total biomass carbon losses. This means that smaller scale (<30 m) forest area loss is implicitly considered as degradation (Methods).
For the period 2016-2022, the gross live biomass carbon losses were \({-2.96}_{-3.38}^{-2.65}\) PgC yr−1, of which ~38% (\({-1.12}_{-1.27}^{-1.02}\) PgC yr−1) was estimated to result from forest area loss and ~62% (\({-1.84}_{-2.12}^{-1.63}\) PgC yr−1) from degradation. Here, the degradation occurred mainly in temperate regions, accounting for 83% of the total biomass carbon loss in this biome, highlighting this as an important process driving biomass carbon loss in temperate biomes. In contrast, the forest area loss (\({-0.85}_{-0.96}^{-0.77}\) PgC yr−1) was the major factor (~66%) in reducing the live biomass carbon stocks in boreal biomes (Supplementary Fig. 12). The impacts of stand-replacing wildfires and other stand-replacing processes were further separated in each 25 km grid cell in forest loss areas (Methods). The latter, including processes such as timber harvest and insect outbreaks, resulted in ~26% (\({-0.76}_{-0.85}^{-0.68}\) PgC yr−1) of the gross live biomass losses across the northern ecosystems, with ~70% (\({-0.53}_{-0.60}^{-0.48}\) PgC yr−1) and ~29% (\({-0.22}_{-0.25}^{-0.20}\) PgC yr−1) occurring in boreal and temperate forests, respectively. The average annual carbon removed through timber harvest reported by FAO is less than half of the losses from non-fire stand-replacing disturbances (Supplementary Fig. 13), suggesting that harvesting may not be the main factor of these losses but rather other types of disturbances18,27, warranting further investigation. Stand-replacing fires (4.51×107 ha) corresponding to ~33% of the forest loss area caused a gross live biomass carbon loss of \({-0.37}_{-0.41}^{-0.33}\) PgC yr−1 in northern ecosystems, of which ~87% (\({-0.32}_{-0.36}^{-0.29}\) PgC yr−1) occurred in boreal forests (Supplementary Fig. 12). While this result confirms that wildfires are a key source of forest disturbance in boreal forests relative to other ecosystems50, other types of forest area disturbances within this biome were ~1.7 times larger than those caused by stand-replacing fires.
In addition to stand-replacing events associated with forest area loss, we further investigated the impacts of all wildfires on live biomass carbon stock changes in northern ecosystems. We considered ‘unburned’ areas defined as areas that include pixels with an annual fire area fraction < 1% within a 25 km grid between 2005 and 2022, while all other pixels were considered as partly burned areas, as in Fan et al.13. During 2016–2022, we found a net biomass carbon change of \({-0.17}_{-0.21}^{-0.12}\) PgC yr−1 in partly burned areas covering ~55% of the study area (Supplementary Fig. 14), resulting in a reduction in live biomass carbon stocks to levels close to those observed a decade ago (Fig. 1c). In contrast, live biomass carbon stocks over unburned areas showed a smaller decline at a rate of \({-0.03}_{-0.05}^{-0.00}\) PgC yr−1. Over temperate regions, partly burned and unburned areas showed similar net losses of \({-0.15}_{-0.20}^{-0.10}\) PgC yr−1 and \({-0.10}_{-0.14}^{-0.07}\) PgC yr−1, respectively. This similarity suggests that biomass carbon loss in this region is not directly driven by fire but by other factors (e.g., droughts), which is in line with the RF-based results (Fig. 3a). In contrast, there is a distinct pattern in the time variations in biomass carbon between fire-disturbed and non-disturbed areas in boreal regions (Fig. 1e). In the latter biome, the gross losses (\({-0.81}_{-0.88}^{-0.76}\) PgC yr−1) and gains (\({+0.79}_{+0.74}^{+0.86}\) PgC yr−1) in the partly burned areas were ~ 40 times larger than the net change (\({-0.02}_{-0.03}^{-0.02}\) PgC yr−1), suggesting that wildfires were an important driver of biomass dynamics in this region. L-VOD-derived gross live biomass carbon losses in partly burned areas of the boreal regions were 2.7 times higher than those estimated solely from stand-replacing fires (\({-0.30}_{-0.34}^{-0.27}\) PgC yr−1), implying that Landsat forest loss data may not capture all small-scale fire-burned forest loss patches and/or that disturbances such as drought and low-intensity understory wildfires may not always result in stand replacement but cause biomass carbon losses, consistent with the studies of Fan et al.13 and Rogers et al.51.
Discussion
Previous studies based on field inventories8, observations from optical11 and high-frequency microwave9 satellite observations have reported increases in live biomass carbon stocks in both temperate and boreal biomes during the first two decades of the 21st century. Recently, an L-VOD-based analysis has even shown that these two biomes represent the main live biomass carbon accumulation at the global scale during 2010–201910. Our study is consistent with these findings, showing that the northern region remained an area of net gain in live biomass carbon stocks for the period 2010-2022. However, we found a reversal of this trend from 2016 to 2022, with total live biomass carbon clearly shifting from an increasing trend to a decreasing trend from around 2016. The downward trend from 2016 to 2022 is reflected by a net loss of biomass carbon over the northern temperate biome (\({-0.26}_{-0.33}^{-0.17}\) PgC yr−1) and boreal Eurasia (\({-0.03}_{-0.04}^{-0.02}\) PgC yr−1).
While a fair comparison with most previous estimates is not straightforward due to differences in the time period studied, we compiled data from several studies providing live biomass carbon estimates over northern ecosystems, overlapping where possible, with the post-2016 decline period identified by L-VOD (Supplementary Table 3). The recent signs of net biomass carbon loss observed by L-VOD were comparable to those reported by other remote sensing products, particularly for the northern temperate biome, where L-VOD estimates ranged from −0.33 PgC yr−1 to -0.06 PgC yr−1, as estimated using the live biomass datasets from Xu et al.11 and the ESA CCI biomass project52. For the boreal biome, L-VOD and CCI showed biomass carbon neutrality, whereas data from Xu et al.11 indicated a significant net loss. The inventory-based integrated data53 also indicated a reduction in the biomass carbon sink of northern ecosystems during the 2010s compared to the 1990s and 2000s (Supplementary Table 3). However, while the overall results are generally consistent between the different data sets, L-VOD, with its rapid update capability, was able to capture the latest reversal in live biomass carbon stock trends in northern ecosystems during 2016-2022. The inventory-based integrated data differ from the remote sensing products due to the exclusion of unmanaged forests and non-forest categories, as well as the lack of consistent inventory data in some countries, such as Russia, since the early 2000s. The CCI biomass maps have inter-annual biases due to the use of different satellite sensors in different years52 and the dataset from Xu et al.11 presents saturation in dense forests due to the use of optical and high frequency observations, limiting their applicability for consistent multi-year change detection34.
We attributed the widespread L-VOD-derived live biomass carbon losses mostly to drought for the temperate biome and to forest disturbances such as wildfires and other stand-replacing processes for the boreal biome, with the former biome being particularly associated with recent frequent severe drought events. Notably, 2022, the year most affected by drought, showed the sharpest decline in live biomass carbon stocks over the 2010-2022 period, with a net change of \({-0.67}_{-0.69}^{-0.41}\) PgC. Of this reduction, 37% (\({-0.25}_{-0.28}^{-0.25}\) PgC) occurred in Europe, which is qualitatively consistent with the widespread impacts of drought in the region observed during the summer of 2022, including reduced net carbon uptake by forests21,26. These losses highlight the severity of the impacts of climate extremes on the regional carbon cycle54. Additionally, our interpretable machine learning analysis identified change in radiation and soil moisture as the two most important climatic contributors influencing the spatial variability of live biomass carbon changes in the northern temperate biomes. Knowing that increased incoming radiation can further exacerbate drought formation, the high ranking of the latter underscores the important role of soil moisture stress in vegetation growth and regulating carbon fluxes in the region55,56. However, it is noteworthy that although multiple variables were included to explain spatial variations in live biomass carbon changes, 49% and 39% of the variance remain unexplained in temperate and boreal biomes, respectively. This suggests that additional climatic and biotic factors, such as canopy structure57, fire radiative power58, and insect disturbances27, merit exploration in future research, together with enhanced L-VOD spatial resolution to better capture small-scale drivers.
A relatively large proportion (41%) of the live biomass carbon losses estimated from L-VOD in boreal biomes was attributed to non-fire forest loss (\({-0.53}_{-0.60}^{-0.48}\) PgC yr−1), which was 1.70 times larger than the losses (23%) from stand-replacing fires (\({-0.30}_{-0.34}^{-0.27}\) PgC yr−1). This suggests that other types of disturbances, such as logging, widespread insect outbreaks and permafrost thaw may have a significant impact on biomass carbon dynamics in the boreal biomes, supporting previous studies in North American boreal forests18,27. Future studies could apply the approach of this study to isolate live biomass carbon loss from timber harvest, provided spatial data defining harvest disturbances or reliable proxies (e.g., forest management boundaries) become available59. Moreover, live biomass carbon losses from non-fire disturbances may not be immediately emitted into the atmosphere in the year they occur but could be delayed (e.g., through the production of harvested wood products) or transferred to other carbon pools (e.g., from live biomass to dead wood and litter)35. In addition, forest fires can have low combustion completeness13,50,51, e.g., in Siberian larch, Siberian fir, or temperate broadleaved forests. Therefore, future studies should further quantify the fate of carbon to better contextualize net carbon stock changes within the framework of the global carbon budget and terrestrial carbon emissions and removals4,11. Importantly, our results showed that the DGVMs were not able to reproduce the effects of wildfires on the observed live biomass carbon evolution in boreal regions, possibly due to insufficient consideration of fire disturbance regimes in the models, particularly inaccurate representation of extreme fire occurrences60,61. Another possible reason is that DGVMs may insufficiently capture vegetation responses to environmental stress, notably the role of VPD in this region27, where increasing VPD was associated with decreasing biomass.
While it is difficult to conclude from the seven years of observations (2016-2022) whether the observed negative trend reflects only quasi-decadal changes or is indicative of a regime shift with consequences for the long-term dynamics of the region, the considerable live biomass losses we observed during this time period highlight the need to reassess the sustainability of this important terrestrial carbon sink under continued global warming. Moreover, the live biomass carbon turning point detected by L-VOD at around 2016 was also confirmed by a C-band VOD product and recent estimates of vegetation biomass changes based on optical and high-frequency satellite observations covering 2000–201911 (Supplementary Fig. 3). If dry years become more frequent, especially when associated with record temperatures such as in 202245, widespread losses of live vegetation biomass may accelerate carbon transfer from vegetation to the atmosphere, weakening the carbon sink function of northern ecosystems. The exceptionally severe Canadian wildfires in 2023 further cement the decreasing net carbon sink trend and the role of wildfires in driving it60. Our study, therefore, highlights the importance of timely monitoring of the trends and spatial distribution of live biomass carbon stock changes in the northern extra-tropics to reassess the role of the terrestrial biosphere and nature-based solutions in mitigating climate change.
Methods
Northern Ecosystems
Our study area is located above 30°N, excluding southern China and southeastern Europe, where L-VOD is susceptible to radio frequency interference (RFI) from human activities33. There are two main ecosystems included in our study area, namely the northern temperate biomes and the boreal biomes, and their boundaries are defined according to Dinerstein et al.62. Due to the potential underestimation of L-VOD in areas where the SMOS footprint contains significant open water bodies10, we have excluded 25 km pixels predominantly characterized by ‘wetland’ land cover from our analysis. Our analysis ultimately focused on pixels (Fig. 1a) that consistently present values for each year from 2010 to 2022, as discontinuous observations could affect the calculation of trends.
SMOS-IC L-VOD
We investigated changes in live biomass carbon over northern ecosystems using the L-VOD product derived from the Soil Moisture and Ocean Salinity (SMOS) satellite, the first L-band passive microwave satellite mission since 2010. L-VOD, which probes the biomass components of live woody vegetation, is a promising satellite data source for monitoring global vegetation carbon stocks in space and time10,33,34,35,36. While the combination of forest inventory plots and remotely sensed datasets (e.g., canopy height estimates obtained from the Geoscience Laser Altimeter System (GLAS) LiDAR and vegetation indices from optical imagery (MODIS)) can also produce spatial maps of aboveground forest biomass estimates, this combination mostly provides data for only a single or discrete year31,32. Most importantly, the integration of multiple satellite images results in delays in data updates, making it difficult to promptly assess regional biomass changes. In contrast, L-VOD, which relies only on passive microwave satellite observations, can be rapidly updated annually, thus allowing monitoring of interannual changes in global vegetation biomass. Moreover, compared to optical vegetation indices and other higher-frequency (> 5 GHz) VOD products9, L-VOD is more sensitive to the biomass of trunks, branches and leaves and does not saturate even in dense forests owing to the higher penetration of low-frequency L-band radiation within the tree canopy34,63. In this study, we utilized the daily 25 km L-VOD product retrieved from the SMOS satellite observations during 2010-2022 using the SMOS-IC algorithm in version 233, originally designed by Institut national de recherche pour l’agriculture, l’alimentation et l’environnement (INRAE) and Centre d’Etudes Spatiales de la Biosphère (CESBIO).
As in refs. 10,13,64,65, strict filtering of the SMOS data is necessary due to the effects of RFI, which disrupt the natural microwave signals from Earth’s surface as measured by passive microwave radiometers. Here, a triple-filtering method is used to filter the daily SMOS L-VOD data for both ascending and descending orbits by taking into account inter-orbit differences and outliers (Supplementary Text 1). The efficiency of this method has been demonstrated in a recent study of global biomass carbon estimation based on L-VOD10. As L-VOD is directly proportional to the total vegetation water content (VWC), daily values of L-VOD reflect not only changes in water mass due to changes in aboveground biomass (AGB) of live vegetation, but also to changes in water mass per unit of biomass (relative water content, RWC)66. Previous studies have assumed that RWC is relatively stable on an annual scale, or during the wet season, and does not show long-term (typically within a decade) increasing or decreasing trends, so that long-term changes in annual L-VOD can be directly related to changes in AGB10,34,35,36,58,66. A detailed analysis of these assumptions and a review of validation studies on the use of L-VOD for biomass monitoring is provided in Wigneron et al.34. In this study, to further eliminate a possible contribution of the RWC variations to changes in annual L-VOD, we developed multiple regression models based on the multiplicative relationship considering L-VOD scales with both dry biomass and RWC67 (Supplementary Text 2). Correction for water information in L-VOD showed that RWC had a minimal effect on annual L-VOD changes, changing the trend by less than 5% between corrected and uncorrected data (Supplementary Figs. 15-17). This is consistent with the conclusions of Yang et al.57 in a study covering the tropics.
Benchmark maps of vegetation carbon density
SMOS L-VOD was calibrated according to aboveground carbon density (AGC; Mg C ha−1) using previously published live biomass maps as a benchmark10,13. However, considering that none of the available live biomass maps can be considered completely reliable (because the true AGC map is unknown) and that they all contain uncertainties and biases, we focused on utilizing the spatial gradients of AGC derived from several existing maps. Any systematic errors in the AGC reference maps are also a major external source of uncertainty in the L-VOD-derived live biomass maps. While individual reference maps may have biases, using an averaged calibration from multiple maps across various regions as the final biomass carbon density mitigates potential biases associated with any single map, as outlined in the L-VOD biomass data production and uncertainty analysis. In a second step, the temporal dynamics of AGC was derived from L-VOD. Here, we used three of the most recent global static AGC benchmarks, including those provided by Saatchi et al.31 updated in 2015, Santoro et al.68 in 2010, as well as ESA CCI biomass map52 (https://climate.esa.int/en/projects/biomass/) in 2017, hereafter referred to as the “Saatchi”, “GlobBiomass” and “ESA CCI”, respectively. All these maps were aggregated by averaging to match the spatial resolution of SMOS (i.e., 25 km grid cell).
Total live biomass carbon derived from L-VOD
The method used here to obtain the calibration coefficients between the annual L-VOD and the reference AGC is the same as that used in ref. 13, where it is documented in detail. The AGC was calculated from the annual L-VOD based on the following empirical spatial calibration:
where a and b are best fit coefficients. Here, we used the annual L-VOD in 2011 to calibrate Eq. 1, and as shown by ref. 13, the year of L-VOD used for the calibration has a very little impact on the final calibration result. In contrast to previous studies, instead of creating a single spatial calibration function for the study area, where possible outliers could affect the fitting parameters, we roughly (not according to administrative regions) divided the northern ecosystems (> 30°N) into three parts: Russia (40°E-180°E), Europe (20°W-40°E), and North America (20°W-180°W) and then computed the optimal coefficients of the calibration equations for the three parts of the study area separately. Twelve calibrations of Eq. 1 were thereby obtained (Supplementary Table 1). They were generally consistent across subregions and throughout the entire northern ecosystems (Supplementary Fig. 18), suggesting that the statistical relationship between L-VOD and vegetation carbon stocks is robust across regions. To estimate the total biomass carbon (BC) from aboveground biomass, we estimated belowground biomass using the root-to-shoot ratio38 suggested by the IPCC Guidelines for National Greenhouse Gas Inventories (Supplementary Table 4). We produced twelve maps of BC stocks using all twelve calibrated L-VOD AGCs. As in refs. 10,35, the median of these twelve maps was finally used to calculate annual BC maps for the period 2010–2022.
Uncertainties and evaluation related to the L-VOD-derived live biomass carbon product
There are two main sources of uncertainty associated with the L-VOD-derived live BC estimates, including: i) external uncertainties, which are caused by the systematic errors in the reference biomass maps; ii) internal uncertainties corresponding to calibration and sampling strategy errors. Previous analyses by Fan et al.13 found a strong dominance of external errors over internal uncertainties (by an order of magnitude), and the total relative uncertainties relative for the carbon stock and carbon stock changes are on the order of 20–30%. Our bootstrap cross-validation results showed that all twelve BC maps exhibited high spatial correlation values and lower RMSE values compared to the corresponding reference dataset, suggesting that no obvious regional bias could be found between the reference biomass maps and the L-VOD-derived live BC over the different continents (Supplementary Table 5). In addition, the 95% bootstrap confidence intervals for the BC estimates obtained using each set of calibrations are small, suggesting that the internal uncertainty due to sampling/calibration errors is small. In consideration of the above, only external errors, as indicated by the range (or spread) of the twelve maps, are reported in the main text.
Wigneron et al.34 provided a review of evaluation studies using L-VOD to calculate BC. Here, to further evaluate the L-VOD-derived live BC data, we used biomass carbon data from the National Forest Inventory (NFI) reported by 43 countries to the Food and Agriculture Organization of the United Nations (FAO) and by 34 Annex I countries to the United Nations Framework Convention on Climate Change (UNFCCC), respectively, which are relatively accurate and independent sources. Spatially, the L-VOD-based live BC stocks were not only found to be similar to the carbon stock values reported by the FAO, but also to circumvent the underestimation problem of using a single reference dataset in some countries (Supplementary Text 3). Temporally, the total net carbon change in managed forests estimated by L-VOD for Annex I countries (Supplementary Fig. 19) between 2010 and 2021 correlated well with that provided by the UNFCCC (R2 = 0.58, P < 0.01) (Supplementary Fig. 2). This long-term comparison added confidence to the L-VOD estimates of biomass carbon changes. It should be noted that the UNFCCC NFI could not capture updated interannual biomass variations, as the NFI was limited by its revisit time rates (in most cases, in each country, several years were required before all plots of the NFI network can be revisited) (Supplementary Fig. 20).
Outputs of dynamic global vegetation models
We utilized the ‘cVeg’ outputs, which represent total live biomass as the sum of above- and below-ground biomass, from 17 DGVMs provided by the TRENDY v12 project39: CABLE-POP, CARDAMOM, CLASS, CLM5.0, DLEM, EDv3, IBIS, ISAM, JSBACH, JULES, LPJ-GUESS, JPLml, LPJwsl, LPX-Bern, OCN, ORCHIDEE and SDGVM. The TRENDY DGVMs performed three factorial simulations. S1 was driven only by CO2, using a pre-industrial climate and land-use mask. S2 included both CO2 and climate while retaining a pre-industrial land-use mask. S3 incorporated CO2, climate, and land use, all varying over time. We selected the S3 simulations, which were spun-up by recycling climate data from 1960 to 1969 until carbon pools were in equilibrium, and then applied varying CO2, climate and land-use conditions over the period 1960 to 2022. We used the mean of these DGVMs in the main text, marking uncertainty by the range between maximum and minimum values. It should be noted that these DGVMs, which have rapid update capabilities, handle emissions using a prognostic fire module. In addition, we analyzed three DGVMs (JULES, OCN, and ORCHIDEE), which were the only models available through 2020 at the time of analysis and used prescribed burned areas from the ESA Climate Change Initiative (CCI-Fires) for calculating emissions69. This allowed us to focus on their consistency with L-VOD estimates before and after incorporating fire diagnostics (Supplementary Text 4).
The RF- SHAP interpretation framework
We applied the Random Forest models-Shapley additive explanations (RF-SHAP) framework to quantify the relative importance of the different climatic, environmental and human-induced variables on the live BC density changes. Comprehensive details of these variables are cataloged in Supplementary Table 2. Here, we aim to understand the underlying processes driving net changes in live biomass carbon density (Δbiomass) in the northern region during 2016–2022, considering both recovery and disturbance factors. While this study focuses on net changes, a separate analysis of biomass carbon losses57 and gains58 may offer additional insights and merits more detailed investigation in future work. The RF model is a widely used supervised learning algorithm, known for reducing overfitting through ensemble learning and random feature selection57. It builds multiple decision trees on random subsets of the training data, and aggregates their predictions for the final result. Our modeling approach employed a hold-out validation strategy, using 75% of the dataset for training the RF model and reserving the remaining 25% for validation. To determine the optimal number of trees for the RF model, we used a trial-and-error approach, incrementally varying the ‘number of trees’ parameter from 25 to 1500 in steps of 100. We ultimately set the number of trees to 400, as the model performance on the test set, measured by R², improved with increasing number of trees and plateaued beyond 400 (Supplementary Fig. 21). We built separate RF models for temperate and boreal biomes, and compared the spatial distribution of predictions from these models with observed values (Supplementary Fig. 7). Finally, we used the state-of-the-art TreeExplainer-based SHAP framework to explain each RF model. SHAP, based on cooperative game theory, quantifies marginal contributions through SHAP values and captures complex interactions, offering a more nuanced view of variable importance than traditional metrics70.
The SHAP value represents each variable’s contribution to the model’s prediction, with positive values associated with higher predicted Δbiomass, and negative values with lower predictions. The importance of each feature was determined by the mean absolute SHAP values, which were scaled to percentage contributions for easier interpretation. We applied SHAP to each RF model 30 times and calculated the standard deviation to quantify uncertainty. To elucidate the nonlinear relationships between Δbiomass and its four key explanatory variables across the two ecosystems, SHAP-based partial dependence plots (PDPs) were generated, showing how SHAP values change with each variable while accounting for both its direct effects and interactions with other predictors70. By combining SHAP rankings with SHAP-based PDPs, we not only identify variable importance but also gain insights into how the top variables shape Δbiomass patterns. To ensure robustness and reliability, the scatter plot highlights points explained by SHAP at least 10 times while rendering others more transparent. The variable range was then divided into 50 bins based on its maximum and minimum values, and the median was computed only for bins containing more than 20 points to clearly illustrate the variable’s contribution pattern to biomass change (Fig. 3).
In addition to the RF model, we also employed a conventional least-squares-based multiple linear regression (MLR) to validate the robustness of the machine learning model. We calculated the variance inflation factor (VIF) for each variable and found that most predictors had a VIF below 3, indicating minimal multicollinearity. Although removing variables with high VIFs (e.g., soil organic matter (SOC) with a VIF ~ 4 in the northern temperate model) reduced the VIFs for the remaining variables, the Akaike Information Criterion (AIC) increased, indicating a reduction in the model’s explanatory power for the target variable. Consequently, the optimal MLR model retained all of the predictors listed in Supplementary Table 2. However, the MLR models could only explain 13% and 15% of biomass variation in northern temperate and boreal biomes, respectively, which is significantly lower than the 51% and 61% explained by our RF models (Supplementary Fig. 22). This suggests that the linear model is insufficient to explain variations in Δbiomass compared to the RF model.
Vegetation and climate variables
Forest loss data
The global forest change product generated from Landsat imagery by Hansen et al.41 was used to calculate the rate of forest loss. Forest loss provided in this product, called ‘lossyear’, is defined as a stand-replacing disturbance, or a change from forest to non-forest. Each 30 m pixel of the ‘lossyear’ data is labeled with a number between 0 and 22, where ‘0’ represents no loss and the others correspond to the loss of forest cover (defined as canopy closure for all vegetation above 5 m) in that year during 2001–2022. Here, the percentage forest loss rate for 2016–2022 at the SMOS resolution is calculated as the proportion of the summed area of forest loss for this period within each 25 km grid cell, as done by Fan et al.13.
Stand-replacing wildfires product
This product, updated from van Wees et al.46, estimates the areas where burned areas or active fires overlap with forest losses at a spatial resolution of 500 m. It defines fire-induced forest losses as encompassing various sequences of events involving wildfire and forest loss. This includes cases where fire (e.g., wildfires) and forest loss occur simultaneously, post-fire forest loss (e.g., tree mortality due to fire damage), and fire following forest loss (e.g., burning of felled trees after logging, which mostly occurs in the same year as logging). We ultimately computed the annual stand-replacing fire fraction at the SMOS resolution using the same approach as above for processing forest loss.
Non-fire induced forest loss
Non-fire related forest losses are defined as forest loss outside of the various sequences of events involving wildfires and forest loss described above. Similarly, the annual fraction of non-fire induced forest loss for the period 2010–2022 was calculated as the proportion of the summed area of non-fire related forest loss within each 25 km grid cell.
Annual burned fraction
The annual active fire maps were generated using active fire products from the latest version of the Terra MODIS Level 3 Thermal Anomalies/Fire 8-Day Global 1 km SIN Grid product (MOD14A2, V6.1). The MOD14A2 provides 8-day fire mask composites at 1 km resolution, which contains the maximum values for each pixel class during the composite period42. This widely used dataset provides global coverage, making it ideal for monitoring fire activity across diverse ecosystems, while its high temporal resolution enables timely detection over large areas12,13. The MOD14A2 active fire observations with a high confidence level were first identified using the associated quality information. We then calculated the annual burned fraction at the SMOS resolution for the study period as the proportion of the summed area of active fires assuming that each 1 km pixel was fully burned.
SPEI-12
The Standardized Precipitation-Evapotranspiration Index (SPEI)-12 refers to drought conditions measured over a 12-month timescale, obtained from the global SPEI database (https://spei.csic.es/). SPEI-12 is a multi-scalar index commonly used to quantify drought. Unlike some other drought indices, SPEI accounts for multiple climatological factors, including both precipitation and temperature, making it crucial for evaluating the impact of climate change on drought conditions71.
Additional data
Soil moisture and 2-m air temperature data resampled to the SMOS resolution were obtained from the European Centre for Medium-Range Weather Forecasts Reanalysis v.5 (ECMWF ERA5), which is an advanced and widely used climate reanalysis47. NDVI data from the MODIS Vegetation Indices 16-Day (MOD13C1) V6.1 product were used as a proxy for leaf biomass. Other high-frequency VOD products at the C-band for 2003–2018 (C-VOD) and X-band (X-VOD) for 2012–2022 are provided by the global long-term microwave VOD Climate Archive (VODCA)40 and the global land parameter data record (LPDR)63, respectively.
Net and gross gains/losses in BC
Mean annual net carbon changes were estimated from the difference in BC between the end year and the start year divided by the year interval. A positive value indicates a net gain (sink) in BC, while a negative value indicates a net loss (source) of carbon. Gross losses (gains) were estimated by accumulating negative (positive) differences between the consecutive years over each pixel, following the method of Brandt et al.55.
Identification of the turning point in live biomass carbon
We used the piecewise linear regression method to determine the turning point of total live BC across the northern ecosystems72. The Mann-Kendall nonparametric trend test was then used to calculate the temporal trends in total live BC after the turning point, while the Sen’s slope statistics were used to determine confidence intervals.
Contributions of forest loss and forest degradation to gross live biomass carbon losses
The gross losses of live BC in a SMOS grid cell can be attributed to loss of forest area, forest degradation and other mechanisms such as non-forest carbon loss. Here, we performed a simple separation of the contribution of forest area loss and degradation to live BC loss based on the method proposed by Harris et al.49 and Qin et al.36. It should be noted that forest loss and forest degradation are not independent processes and cannot be explicitly separated on a 25 km grid cell, and for simplicity we defined all mechanisms that do not result in loss of forest area as degradation. As in ref. 36, the contribution of forest loss to gross live BC loss (Gross BC lossforestloss) was first isolated by multiplying the total area of forest loss (calculated from the 30 m Landsat forest loss map41) during 2017-2022 by the BC density in 2016 (Eq. 2). Degradation was then roughly estimated as subtracting this forest loss from the gross L-VOD-derived BC losses (Eq. 3).
The contributions of stand-replacing wildfires (Gross BC lossstand-replacing fire) and other stand-replacing processes (e.g., clearcutting and widespread insect outbreaks) were further separated from the gross live BC losses due to forest loss (Gross BC lossforestloss). To do so, we multiplied the total area of forest loss due to stand-replacing fires (i.e., the sum of the area of forest loss overlapping with fire in each year during 2017-2022) by the live BC density in 2016.
Data availability
Source data are provided with the paper as a zipped folder, available at https://zenodo.org/records/15283697. The SMOS-IC L-VOD from this study can be freely downloaded from INRAE Bordeaux remote sensing lab website (https://ib.remote-sensing.inrae.fr/). The Saatchi biomass map is available upon request from Dr. S. Saatchi (sasan.s.saatchi@jpl.nasa.gov). The global map of ecoregions is available at https://ecoregions.appspot.com/. The ERA5 reanalysis data can be obtained from https://www.ecmwf.int/en/forecasts/dataset/ecmwf-reanalysis-v5. MOD13C1 NDVI data is available from https://lpdaac.usgs.gov/products/mod13c1v061/, while MOD14A2 is processed by the GEE platform. The model outputs from TRENDY are available upon request from Dr. S. Sitch (S.A.Sitch@exeter.ac.uk) or can be accessed at https://globalcarbonbudgetdata.org/. Additional data involved in this paper can all be found in the corresponding references.
Code availability
The main code used to produce the results can be accessed at https://zenodo.org/records/15283697.
References
Keenan, R. J. et al. Dynamics of global forest area: results from the FAO global forest resources assessment 2015. Ecol. Manag. 352, 9–20 (2015).
Ciais, P., Tans, P., Trolier, M., White, J. & Francey, R. A large northern hemisphere terrestrial CO2 sink indicated by the 13C/12C ratio of atmospheric CO2. Science 269, 1098–1102 (1995).
Tans, P. P., Fung, I. Y. & Takahashi, T. Observational contrains on the global atmospheric CO2 budget. Science 247, 1431–1438 (1990).
Friedlingstein, P. et al. Global carbon budget 2024. Earth Syst. Sci. Data 17, 965–1039 (2024).
Piao, S., Friedlingstein, P., Ciais, P., Viovy, N. & Demarty, J. Growing season extension and its impact on terrestrial carbon cycle in the Northern Hemisphere over the past 2 decades. Glob. Biogeochem. Cycles 21, GB3018 (2007).
Piao, S. et al. Characteristics, drivers and feedbacks of global greening. Nat. Rev. Earth Environ. 1, 14–27 (2020).
Pugh, T. A. et al. Role of forest regrowth in global carbon sink dynamics. Proc. Natl Acad. Sci. USA 116, 4382–4387 (2019).
Pan, Y. et al. A large and persistent carbon sink in the world’s forests. Science 333, 988–993 (2011).
Liu, Y. Y. et al. Recent reversal in loss of global terrestrial biomass. Nat. Clim. Change 5, 470–474 (2015).
Yang, H. et al. Global increase in biomass carbon stock dominated by growth of northern young forests over past decade. Nat. Geosci. 16, 886–892 (2023).
Xu, L. et al. Changes in global terrestrial live biomass over the 21st century. Sci. Adv. 7, eabe9829 (2021).
Zheng, B. et al. Record-high CO2 emissions from boreal fires in 2021. Science 379, 912–917 (2023).
Fan, L. et al. Siberian carbon sink reduced by forest disturbances. Nat. Geosci. 16, 56–62 (2023).
Zscheischler, J. et al. Extreme events in gross primary production: a characterization across continents. Biogeosciences 11, 2909–2924 (2014).
Kautz, M., Meddens, A. J., Hall, R. J. & Arneth, A. Biotic disturbances in Northern Hemisphere forests–a synthesis of recent data, uncertainties and implications for forest monitoring and modelling. Glob. Ecol. Biogeogr. 26, 533–552 (2017).
Van Mantgem, P. J. et al. Widespread increase of tree mortality rates in the western United States. Science 323, 521–524 (2009).
Liu, Q. et al. Drought-induced increase in tree mortality and corresponding decrease in the carbon sink capacity of Canada’s boreal forests from 1970 to 2020. Glob. Change Biol. 29, 2274–2285 (2023).
Ma, Z. et al. Regional drought-induced reduction in the biomass carbon sink of Canada’s boreal forests. Proc. Natl Acad. Sci. USA 109, 2423–2427 (2012).
Büntgen, U. et al. Recent European drought extremes beyond common era background variability. Nat. Geosci. 14, 190–196 (2021).
Bastos, A. et al. Direct and seasonal legacy effects of the 2018 heat wave and drought on European ecosystem productivity. Sci. Adv. 6, eaba2724 (2020).
Van Der Woude, A. M. et al. Temperature extremes of 2022 reduced carbon uptake by forests in Europe. Nat. Commun. 14, 6218 (2023).
Soja, A. J. et al. Climate-induced boreal forest change: predictions versus current observations. Glob. Planet. Change 56, 274–296 (2007).
Schaphoff, S., Reyer, C. P., Schepaschenko, D., Gerten, D. & Shvidenko, A. Tamm review: observed and projected climate change impacts on Russia’s forests and its carbon balance.Ecol. Manag. 361, 432–444 (2016).
Goulden, M. L. & Bales, R. C. California forest die-off linked to multi-year deep soil drying in 2012–2015 drought. Nat. Geosci. 12, 632–637 (2019).
Thompson, V. et al. The 2021 western North America heat wave among the most extreme events ever recorded globally. Sci. Adv. 8, eabm6860 (2022).
Hermann, M. et al. Meteorological history of low-forest-greenness events in Europe in 2002–2022. Biogeosciences 20, 1155–1180 (2023).
Wang, J. A., Baccini, A., Farina, M., Randerson, J. T. & Friedl, M. A. Disturbance suppresses the aboveground carbon sink in North American boreal forests. Nat. Clim. Change 11, 435–441 (2021).
Williams, C. A., Gu, H., MacLean, R., Masek, J. G. & Collatz, G. J. Disturbance and the carbon balance of US forests: a quantitative review of impacts from harvests, fires, insects, and droughts. Glob. Planet. Change 143, 66–80 (2016).
Tagesson, T. et al. Recent divergence in the contributions of tropical and boreal forests to the terrestrial carbon sink. Nat. Ecol. Evol. 4, 202–209 (2020).
Stephens, B. B. et al. Weak northern and strong tropical land carbon uptake from vertical profiles of atmospheric CO2. Science 316, 1732–1735 (2007).
Saatchi, S. S. et al. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl Acad. Sci. USA 108, 9899–9904 (2011).
Baccini, A. et al. Tropical forests are a net carbon source based on aboveground measurements of gain and loss. Science 358, 230–234 (2017).
Wigneron, J.-P. et al. SMOS-IC data record of soil moisture and L-VOD: historical development, applications and perspectives. Remote Sens. Environ. 254, 112238 (2021).
Wigneron, J.-P. et al. Global carbon balance of the forest: satellite-based L-VOD results over the last decade. Front. Remote Sens. 5, 1338618 (2024).
Bar-On, Y. M. et al. Recent gains in global terrestrial carbon stocks are mostly stored in nonliving pools. Science 387, 1291–1295 (2025).
Qin, Y. et al. Carbon loss from forest degradation exceeds that from deforestation in the Brazilian Amazon. Nat. Clim. Change 11, 442–448 (2021).
Fawcett, D. et al. Declining Amazon biomass due to deforestation and subsequent degradation losses exceeding gains. Glob. Change Biol. 29, 1106–1118 (2023).
Mokany, K., Raison, R. J. & Prokushkin, A. S. Critical analysis of root: shoot ratios in terrestrial biomes. Glob. Change Biol. 12, 84–96 (2006).
Friedlingstein, P. et al. Global carbon budget 2023. Earth Syst. Sci. Data 15, 5301–5369 (2023).
Moesinger, L. et al. The global long-term microwave vegetation optical depth climate archive (VODCA). Earth Syst. Sci. Data 12, 177–196 (2020).
Hansen, M. C. et al. High-resolution global maps of 21st-century forest cover change. Science 342, 850–853 (2013).
Giglio, L., Schroeder, W. & Justice, C. O. The collection 6 MODIS active fire detection algorithm and fire products. Remote Sens. Environ. 178, 31–41 (2016).
Curtis, P. G., Slay, C. M., Harris, N. L., Tyukavina, A. & Hansen, M. C. Classifying drivers of global forest loss. Science 361, 1108–1111 (2018).
Delpierre, N., Berveiller, D., Granda, E. & Dufrêne, E. Wood phenology, not carbon input, controls the interannual variability of wood growth in a temperate oak forest. N. Phytol. 210, 459–470 (2016).
Tripathy, K. P., Mukherjee, S., Mishra, A. K., Mann, M. E. & Williams, A. P. Climate change will accelerate the high-end risk of compound drought and heatwave events. Proc. Natl Acad. Sci. USA 120, e2219825120 (2023).
Van Wees, D. et al. The role of fire in global forest loss dynamics. Glob. Change Biol. 27, 2377–2391 (2021).
Hersbach, H. et al. The ERA5 global reanalysis. Q. J. R. Meteorol. Soc. 146, 1999–2049 (2020).
Kurz, W. A. et al. Mountain pine beetle and forest carbon feedback to climate change. Nature 452, 987–990 (2008).
Harris, N. L. et al. Global maps of twenty-first century forest carbon fluxes. Nat. Clim. Change 11, 234–240 (2021).
Gauthier, S., Bernier, P., Kuuluvainen, T., Shvidenko, A. Z. & Schepaschenko, D. G. Boreal forest health and global change. Science 349, 819–822 (2015).
Rogers, B. M., Soja, A. J., Goulden, M. L. & Randerson, J. T. Influence of tree species on continental differences in boreal fires and climate feedbacks. Nat. Geosci. 8, 228–234 (2015).
Santoro, M. & Cartus, O. ESA Biomass Climate Change Initiative (Biomass_cci): global datasets of forest above-ground biomass for the years 2010, 2015, 2016, 2017, 2018, 2019, 2020 and 2021, v5.01. NERC EDS Centre for Environmental Data Analysis https://doi.org/10.5285/bf535053562141c6bb7ad831f5998d77 (2024).
Pan, Y. et al. The enduring world forest carbon sink. Nature 631, 563–569 (2024).
Reichstein, M. et al. Climate extremes and the carbon cycle. Nature 500, 287–295 (2013).
Brandt, M. et al. Satellite passive microwaves reveal recent climate-induced carbon losses in African drylands. Nat. Ecol. Evol. 2, 827–835 (2018).
Kannenberg, S. A., Anderegg, W. R., Barnes, M. L., Dannenberg, M. P. & Knapp, A. K. Dominant role of soil moisture in mediating carbon and water fluxes in dryland ecosystems. Nat. Geosci. 17, 38–43 (2024).
Yang, H. et al. Climatic and biotic factors influencing regional declines and recovery of tropical forest biomass from the 2015/16 El Niño. Proc. Natl Acad. Sci. USA 119, e2101388119 (2022).
Feng, Y. et al. Global patterns and drivers of tropical aboveground carbon changes. Nat. Clim. Change 14, 1064–1070 (2024).
Lesiv, M. et al. Global forest management data for 2015 at a 100 m resolution. Sci. Data 9, 199 (2022).
Byrne, B. et al. Carbon emissions from the 2023 Canadian wildfires. Nature 633, 835–839 (2024).
Nurrohman, R. K. et al. Future projections of Siberian wildfire and aerosol emissions. Biogeosciences 21, 4195–4227 (2024).
Dinerstein, E. et al. An ecoregion-based approach to protecting half the terrestrial realm. BioScience 67, 534–545 (2017).
Li, X. et al. Global-scale assessment and inter-comparison of recently developed/reprocessed microwave satellite vegetation optical depth products. Remote Sens. Environ. 253, 112208 (2021).
Li, X. et al. The first global soil moisture and vegetation optical depth product retrieved from fused SMOS and SMAP L-band observations. Remote Sens. Environ. 282, 113272 (2022).
Ma, H. et al. Surface soil moisture from combined active and passive microwave observations: integrating ASCAT and SMAP observations based on machine learning approaches. Remote Sens. Environ. 308, 114197 (2024).
Frappart, F. et al. Global monitoring of the vegetation dynamics from the vegetation optical depth (VOD): a review. Remote Sens 12, 2915 (2020).
Momen, M. et al. Interacting effects of leaf water potential and biomass on vegetation optical depth. J. Geophys. Res. Biogeosci. 122, 3031–3046 (2017).
Santoro, M. et al. The global forest above-ground biomass pool for 2010 estimated from high-resolution satellite observations. Earth Syst. Sci. Data 13, 3927–3950 (2021).
Lizundia-Loiola, J., Otón, G., Ramo, R. & Chuvieco, E. A spatio-temporal active-fire clustering approach for global burned area mapping at 250 m from MODIS data. Remote Sens. Environ. 236, 111493 (2020).
Lundberg, S. M. et al. From local explanations to global understanding with explainable AI for trees. Nat. Mach. Intell. 2, 56–67 (2020).
Vicente-Serrano, S. M., Beguería, S. & López-Moreno, J. I. A multiscalar drought index sensitive to global warming: the standardized precipitation evapotranspiration index. J. Clim. 23, 1696–1718 (2010).
Toms, J. D. & Lesperance, M. L. Piecewise regression: a tool for identifying ecological thresholds. Ecology 84, 2034–2041 (2003).
Acknowledgements
This study received support from the European Space Agency Climate Change Initiative (ESA-CCI) RECCAP2 project 1190 (contract no. 4000123002/18/I-NB). It was also funded by the Centre National d’Études Spatiales (CNES, France) as part of its TOSCA program supporting research activities in the context of the SMOS mission. P.C. acknowledges support from the EU EYE-CLIMA project (Grant Agreement No. 101081395). X.X. was supported by a research grant from the U.S. National Science Foundation (#1946093). We also acknowledge Dr. Evgenii Churiulin for providing the DGVMs.
Author information
Authors and Affiliations
Contributions
X.J.L., J.-P.W. and P.C. conceived and designed the overall study. X.J.L. and J.-P.W. prepared the SMOS-IC L-VOD product. S.S. and A.B. processed the TRENDY data. X.J.L., J.-P.W., P.C., R.F., J. Chave, S.S., J.G.C., M.B., L.F., X.X., S.L.T., H.W., C.A. and F.F. contributed to the interpretation of the results. H.Y., M.J. Wang, P.M., Y.W.Q., Z.P.X., T.X.C., L.Y., L.H., Y.Z., X.Z.L., Y.Q.L. and A.D.T. provided comments and feedback on the discussion. X.J.L. performed the analysis and drafted the manuscript with input from all co-authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Wouter Peters, who co-reviewed with Marnix van de Sande, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, X., Ciais, P., Fensholt, R. et al. Large live biomass carbon losses from droughts in the northern temperate ecosystems during 2016-2022. Nat Commun 16, 4980 (2025). https://doi.org/10.1038/s41467-025-59999-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-59999-2
