
Abstract
FoF1-ATP synthase (FoF1) interconverts the energy of the proton motive force (pmf) and that of ATP through the mechanical rotation. The H+/ATP ratio, one of the most crucial parameters in bioenergetics, varies among species due to differences in the number of H+-binding c-subunits, resulting in H+/ATP ratios ranging from 2.7 to 5. In this study, we seek to significantly enhance the H+/ATP ratio by employing an alternative approach that differs from that of nature. We engineer FoF1 to form multiple peripheral stalks, each bound to a proton-conducting a-subunit. The engineered FoF1 exhibits an H+/ATP ratio of 5.8, surpassing the highest ratios found in naturally occurring FoF1s, enabling ATP synthesis under low pmf conditions where wild-type enzymes cannot synthesize ATP. Structural analysis reveals that the engineered FoF1 forms up to three peripheral stalks and a-subunits. This study not only provides valuable insights into the H+-transport mechanism of FoF1 but also opens up possibilities for engineering the foundation of cellular bioenergetics.
Similar content being viewed by others
Introduction
FoF1-ATP synthase (FoF1) is a ubiquitous enzyme found in the membranes of mitochondria, chloroplasts and bacteria. It synthesizes ATP from ADP and inorganic phosphate (Pi) coupled with proton translocation across membranes along the proton motive force (pmf)1,2,3. FoF1 is a unique molecular motor complex composed of two rotary molecular motors: F1 and Fo1,2. Bacterial FoF1 exhibits the simplest subunit composition of a1b2cx (x varies among species) for Fo and α3β3γ1δ1ε1 for F14,5,6 (Fig. 1a). Fo is a membrane-embedded molecular motor driven by pmf. When protons are translocated through the proton pathway in Fo along pmf, the multimeric rotor ring composed of c-subunits (termed c-ring) rotates against stator ab2 (Fig. 1a, b). F1 is the catalytic core domain of FoF1 for ATP synthesis and hydrolysis7. When isolated from Fo, F1 acts as an ATP-driven molecular motor that rotates the γε rotor complex against the catalytic α3β3 stator ring coupled with ATP hydrolysis8. These two motors are coupled via two stalks: the peripheral stalk, composed of the b2 dimer stalk and the δ subunit. The central rotor stalk is composed of a γε complex and a c-ring. Under ATP synthesis conditions where pmf is sufficient and the rotational torque of Fo exceeds that of F1, Fo rotates the γε complex in F1 in the reverse direction of ATP hydrolysis, driving the ATP synthesis reaction on the α3β3 ring9,10. Conversely, when the torque of F1 exceeds that of Fo, F1 rotates the c-ring in Fo, forcing Fo to pump protons in the reverse direction, thereby generating pmf. Thus, FoF1 interconverts the pmf and chemical potential of ATP hydrolysis through mechanical rotation.

a Bacterial ATP synthase (from thermophilic Bacillus PS3) consists of F1 (α3β3γδε) and Fo (ab2c10) motors. As F1 has three catalytic sites, three ATP molecules are synthesized per turn of the rotor subunits (γεc10) against the stator subunits (α3β3δab2) during ATP synthesis. b Models of proton translocation through Fo coupled with the rotation of the c-ring. The highly conserved arginine residues of the a-subunit and glutamate (or aspartate) residues of c-subunits are depicted with pink and black open circles, respectively. Protons are depicted as light blue circles. In the models of c10-ring/single a-subunit and c15-ring/single a-subunit, 10 and 15 protons, equal to the number of the c-subunits, are transferred in one turn, respectively. Conversely, in the c10-ring and triple a-subunits model, a total of 30 protons, equal to the number of the c-subunits multiplied by the number of the a-subunits, are transferred in one turn.
Considering the Gibbs free energy of this coupling reaction (\(\Delta G^{\prime}\)),
\(\Delta G^{\prime}\) is givens:
where \(\Delta {G}_{{{{\rm{ATP}}}}}^{{\prime} }\) is the Gibbs free energy of ATP synthesis, \(F\) is Faraday’s constant, \(n\) is the H+/ATP ratio, which is defined as the number of protons translocated through Fo coupled with a single turnover of ATP synthesis on F1. Hence, the following conditions must be satisfied to drive ATP synthesis:
Thus, the H+/ATP ratio is the critical factor in determining the lower limit of pmf required for ATP synthesis, given that \(\Delta {G}_{{{{\rm{ATP}}}}}^{{\prime} }\) does not largely vary among species. The H+/ATP ratio is principally defined by the ratio of the reaction stoichiometry of Fo per turn of the rotor complex to that of F1, and the ratio of H+/turn of Fo to ATP/turn of F1. All the F1s studied so far, without exception, have three catalytic β subunits and couple three reactions of ATP hydrolysis/synthesis per turn, defining the ATP/turn ratio as 310. The reaction stoichiometry of Fo, H+/turn varies among species due to differences in the number of c-subunits in the c-ring. According to the half-channel model, supported by recent structural studies, the H+ pathway in Fo is formed by the c-ring and the a-subunit, which has two half-channels exposed to the periplasmic or cytoplasmic side of the membrane11,12,13. During ATP synthesis, H+ from the periplasmic solution enters the half-channel exposed to the periplasmic space and is transferred to one of the c-subunits in the c-ring. Following one revolution of the c-ring, H+ is released into the cytoplasmic solution through the opposite half-channel (Fig. 1b, Supplementary Fig. 1). Thus, the half-channel model assumes that the stoichiometry of H+/turn is determined by the number of c-subunits.
The number of c-subunits in the c-ring in F-type ATP synthases ranges from 8 to 15, depending on the species14,15,16. When assuming perfect energy coupling between F1 and Fo, the H+/ATP ratio should vary between 2.7 and 5.0 among the species. Various groups have attempted to experimentally determine the H+/ATP ratio from the biochemical measurements of the thermodynamic equilibrium point where the pmf and \({\Delta G}_{{{{\rm{ATP}}}}}^{{\prime} }\) are balanced. The Bacillus PS3 FoF1, with a c10-ring, has been reported to show good agreement with the structurally expected H+/ATP ratio of 3.317. FoF1s from E. coli and yeast mitochondria, both of which also have the c10-ring, exhibit slightly different H+/ATP ratios: 4.0 ± 0.318 and 2.9 ± 0.219, respectively. For spinach chloroplast FoF1 with the c14-ring, two independent studies reported smaller values for the H+/ATP ratio: 4.0 ± 0.218 and 3.9 ± 0.319, which are lower than the expected value of 4.7. Thus, the experimentally determined H+/ATP ratios are close to, but not always identical to, the structurally expected values, varying within a narrow range of 3 to 4.
The H+/ATP ratio of FoF1 is one of the most critical parameters in the bioenergetic system of cells, which defines the energy cost of ATP synthesis and the threshold pmf required for ATP synthesis (see Eq. 3)14,17. Since \(\Delta {G}_{{{{\rm{ATP}}}}}^{\prime}\) does not largely differ across organisms, FoF1 with a higher H+/ATP ratio can synthesize ATP even at a lower pmf. In fact, alkaliphilic bacteria living in highly alkaline environments and photosynthetic organisms that grow under light-limiting conditions have a c-ring with a large number of c-subunits20,21,22. This is thought to be an evolutionary adaptation that allows stable ATP synthesis under low and/or unstable pmf conditions22,23. Thus, organisms may have optimized the H+/ATP ratio through evolution by tuning the stoichiometry of the c-ring to meet their energetic requirements.
Conversely, when reconsidering the half-channel mechanism, we can assume that the stoichiometry of H+/turn in Fo is determined not only by the number of c-subunits but also by the number of a-subunits (Fig. 1b, bottom). In particular, the Fo structures solved so far show that a large portion of the c-ring is exposed to the lipid bilayer, suggesting the possibility of accommodating one or two additional a-subunits, although FoF1 with multiple a-subunits has not yet been identified.
Here, we explore the possibility of doubling or tripling the H+/ATP ratio of FoF1 by increasing the number of a-subunits in Fo, rather than increasing the stoichiometry of the c-ring as observed in nature.
Results
Design for multiple peripheral stalks
So far, all ATP synthases have a single copy of the a-subunit per FoF1 complex. Considering that the a-subunit is tightly bound to the membrane portion of the peripheral stalk, the key factor for multiplying the a-subunits should be the structural mechanism that limits the number of peripheral stalks. The peripheral stalk of the b2 dimer extends from the membrane to the upper surface of the α3β3 subcomplex of F1, binding to the δ subunit. The δ subunit binds to the top of the α3β3 subcomplex, associating with the N-terminal regions of the three α subunits. One α subunit interacts with the C-terminal region of the δ subunit, while the other two interact with the N-terminal region of the δ subunit, as indicated by the arrows in Fig. 2a. Since the N-terminal domain of the δ subunit is located in the central concavity of the α3β3 ring, occupying the pseudo-threefold symmetry axis of the α3β3 ring, it is reasonable to assume that the N-terminal domain of the δ subunit disrupts the pseudo-threefold symmetry, limiting the stoichiometry of the peripheral stalk to one (Fig. 2a). We hypothesized that by removing the N-terminal domain of the δ subunit (Fig. 2b), it becomes possible to accommodate a truncated δ subunit on each α subunit (Fig. 2c). A possible concern regarding the truncation is that the truncated δ subunit (δΔN) does not form a stable complex with the α3β3 ring. Therefore, we designed the δΔN-α fusion construct of Bacillus PS3 FoF1, where the C-terminus of the δ subunit was genetically fused to the N-terminus of the α subunit. In addition, the inhibitory C-terminal domain of the ε subunit was removed to enhance enzymatic activity17,24. In this study, Bacillus PS3 FoF1-εΔC was used as a wild-type FoF1 for comparison.

Top views of Bacillus PS3 FoF1 from the cytoplasm (PDB ID: 6N2Y). The α (red), β (yellow), γ (blue), δ subunits (orange), and the b2 stalk (pink) are shown. The non-transparent red parts indicated by the arrows represent the N-terminal region (residues 2–30) of the α subunit. a Asymmetric interactions between the δ and α subunits. The single δ subunit interacts with the three α subunits, occupying the central concavity of the α3β3 subcomplex. b Model structure after deletion of the N-terminal domain (residues 2–104) of the δ subunit. The central concavity is exposed, and two of three α subunits are unoccupied. c Model structure after each of the N-terminus of the three α subunits is fused to the C-terminal domain (residues 105–178) of the δ subunit.
SDS-PAGE analysis of subunit stoichiometry
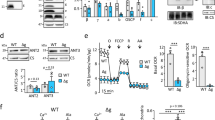
The δΔN-α fused FoF1 was purified following a previously reported procedure for the wild-type Bacillus PS3 FoF117. To estimate the subunit stoichiometry, the δΔN-α fused FoF1 was analyzed by SDS-PAGE with the wild-type FoF1 for comparison (Fig. 3a, b). The δΔN-α fused FoF1 lacked δ and α, and the δΔN-α fusion appeared above the band position for the α subunit (Fig. 3a). The δΔN-α fused FoF1 retained the complete set of subunits. Then, we estimated the subunit stoichiometry of the a- and b-subunits in the δΔN-α fused FoF1 by using the γ subunit as the internal reference in comparison with the wild-type (Fig. 3b, Supplementary Fig. 2). In the wild-type, the b-subunit exhibited a band intensity comparable to that of the γ subunit, whereas in the δΔN-α fused FoF1, the b-subunit showed higher signals relative to the γ subunit. Similarly, the δΔN-α fused FoF1 exhibited a higher band intensity for the a-subunit compared to the wild type, indicating that the δΔN-α fused FoF1 increases the stoichiometry of the a- and b-subunits. For a more quantitative estimation, we plotted calibration lines for the wild-type and mutant subunits and standardized the lines using the γ subunit calibration lines as the internal control (Fig. 3b). We then determined the stoichiometries of the a- and b-subunits in the δΔN-α fused FoF1 by comparing them to the wild-type. The estimated stoichiometries of the a- and b-subunits were 2.2 and 1.8 times higher than those of the wild-type FoF1, respectively (see Fig. 3b legend). Thus, it was confirmed that the δΔN-α fused FoF1 has multiple, two on average, peripheral stalks and a-subunits.

a SDS-PAGE analysis of the purified wild-type (WT) Bacillus PS3 FoF1 and the δΔN-α fused FoF1. Each sample was derived from a single purification batch. 3 μg of FoF1 was loaded in each lane. The molecular masses of the δΔN-α, α, β, γ, a, δ, b, εΔC, and c-subunits are 63, 55, 53, 32, 26, 20, 19, 9, and 7 kDa, respectively. The experiment was independently repeated three times with similar results. b The band intensity vs total protein amount. A single series of diluted FoF1 from the same purification batch was loaded into a gel and subjected to SDS-PAGE analysis. The band intensity of each subunit was plotted against the total amount of FoF1 loaded for SDS-PAGE analysis. The plots were fitted with a linear function. The slopes for γ (black), a- (red), and b-subunits (blue) were determined to be 2082, 502, and 2004 (arb. units/µg) for the WT FoF1, and 1521, 821, and 2675 (arb. units/µg) for the δΔN-α fused FoF1, respectively. By normalizing the slopes of the a- and b-subunits to the slope of the γ subunit of each FoF1, the stoichiometries of the a- and b-subunits of the δΔN-α fused FoF1 were estimated to be 2.2 ( = (821/1521)/(502/2082)) and 1.8 ( = (2675/1521)/(2004/2082)) times higher than those of the wild-type FoF1, respectively.
Functional analysis of the H+/ATP ratio
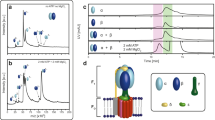
We attempted to determine the H+/ATP ratio of the δΔN-α fused FoF1 through biochemical measurements of the thermodynamic equilibrium between pmf and \(\Delta {G}_{{{{\rm{ATP}}}}}^{,}\), as previously reported17. Firstly, we prepared the FoF1-reconstituted proteoliposomes (PLs) and incubated them in an acidic buffer. The PLs were injected into the base assay medium to initiate ATP synthesis. The ATP synthesis/hydrolysis activity was monitored with the luciferin/luciferase assay system under various pmf conditions, with a given reaction quotient, Q \((=\left[{\mbox{ATP}}\right]/\left(\left[{\mbox{ADP}}\right]\cdot \left[{\mbox{Pi}}\right]\right))\). Figure 4a shows the time courses of the assay, in which the initial rate was determined. Figure 4b shows the reaction rates plotted against the pmf when Q = 2.5. The δΔN-α fused FoF1 was shown to catalyze the ATP synthesis reaction even at low pmf, at which wild-type FoF1 is unable to synthesize ATP. For a more quantitative analysis, the data points were fitted with an exponential function to determine the equilibrium pmf (pmfeq), where the torques of F1 and Fo are balanced and the net reaction rate is zero. At the condition of Fig. 4b where Q = 2.5, pmfeq was determined to be 68 mV for the δΔN-α fused FoF1 and 133 mV for the wild-type. Thus, the minimum pmf for ATP synthesis was halved for the δΔN-α fused FoF1, suggesting that the functional H+/ATP ratio of the δΔN-α fused FoF1 is also doubled, in agreement with SDS-PAGE analysis. For further confirmation, we determined the pmfeq at various Q values (Supplementary Fig. 3). Under all conditions, the pmfeq of the δΔN-α fused FoF1 was nearly half that of the wild-type.

a Time courses of the ATP synthesis/hydrolysis activity of the reconstituted proteoliposomes (PLs) at different pmf. ATP synthesis reaction was measured using the luciferin/luciferase system. The reaction quotient, Q was 2.5; [ATP] = 500 nM, [ADP] = 20 µM, [Pi] = 10 mM. The rate of catalysis was determined from the initial slopes (bold lines). b The ATP synthesis/hydrolysis rates determined from (a) were plotted against pmf. The data points were fitted with an exponential function for the determination of the equilibrium pmf, pmfeq, as the interception of the x axis. c Determination of the H+/ATP ratio. The mean (filled circles) and the SD of \(F\cdot {{pmf}}_{{eq}}\) values were determined from 3 to 4 independent biological replicates (open circles) at each Q condition using different batches of purified enzymes, and \(2.3{RT}\cdot \log Q\) values were plotted against the corresponding \(F\cdot {{pmf}}_{{eq}}\) values according to Eq. (5). Sample sizes (n) from left to right are: 3, 3, 3, 4, 3 (WT) and 4, 3, 3, 4, 3 (δΔN-α). Each line represents a linear regression fit to the dataset obtained from each FoF1.
For the comprehensive analysis of the functional H+/ATP ratio, Eq. (2) was transformed as below,
where \(\Delta {G}_{{ATP}}^{0,}\) is the Gibbs free energy of ATP synthesis under the biochemical standard state, R and T are the gas constant and absolute temperature, respectively. Here, pmfeq values were experimentally determined under defined Q conditions. The other values are constant. Therefore, when \(2.3{RT}\cdot \log Q\) is plotted against \(F\cdot {{pmf}}_{{eq}}\), n (= the H+/ATP ratio) is determined as the slope of the data points. In addition, \({G}_{{ATP}}^{0,}\) is determined as the interception of the y axis. As shown in Fig. 4c, the δΔN-α fused FoF1 exhibited a significantly steeper slope compared to the wild-type FoF1. From the linear fitting, the H+/ATP ratio was determined to be 5.8 ± 0.4 and 3.0 ± 0.2 (fitted value ± SE of the fit) for the δΔN-α fused FoF1 and wild-type, respectively. Although the H+/ATP ratio of the wild-type is slightly lower than the structurally expected value of 3.3 and the reported value (3.3 ± 0.1)17, δΔN-α fused FoF1 was shown to double the H+/ATP ratio, in close agreement with the subunit stoichiometry analysis from SDS-PAGE. This agreement suggests that the δΔN-α fused FoF1 has two functional a-subunits in the ensemble average. From the interception of the y axis, the \(\Delta {G}_{{{{\rm{ATP}}}}}^{0,}\) values are determined to be 38 ± 3 kJ mol−1 (fitted value ± SE of the fit) for both. This value shows a fine agreement with the reported values for Bacillus PS3 FoF1 (39 ± 1 kJ mol−1)17, E. coli (38 ± 3 kJ mol−1)18, yeast (36 ± 3 kJ mol−1)19, and chloroplasts (38 ± 3 and 37 ± 3 kJ mol−1)18,19, supporting the validity of the experiment.
Cryo-EM structural analysis
We determined the structure of the δΔN-α fused FoF1 by single-particle cryo-EM analysis. The purified δΔN-α fused FoF1 in detergent was applied to EM grids, frozen in liquid ethane, and imaged with 300 kV cryo-EM followed by single-particle analysis using cryoSPARC. The cryo-EM map was obtained by ab initio 3D reconstruction and classification, followed by refinement with C1 symmetry. Since the rotor complex in FoF is oriented at one of the three catalytic dwell angles relative to the stator ring and peripheral stalk, alignment against the central core complex, including the rotor complex, revealed three distinct positions of the peripheral stalk, each separated by 120°, as observed in previous reports25,26. Map structure classification was performed by masking the peripheral-stalk positions, confirming the presence or absence of the peripheral stalk at the masked position (Supplementary Fig. 4). This classification was conducted for each stalk position: Stalks 1, 2, and 3. Thus, the map structure was classified into eight sub-classes. The overall resolution was 2.5–3.2 Å (Supplementary Fig. 4). While the wild-type FoF1 contains only a single peripheral stalk, this structural classification confirmed that some fractions of the particles contained multiple peripheral stalks (Fig. 5 and Supplementary Fig. 4). A significant fraction of the particles, however, had either no peripheral stalk or only a single one. The percentages of FoF1 structures with 0, 1, 2, and 3 peripheral stalks, as determined from the 3D classification, were 15, 51, 26, and 8%, respectively. Because of the lower resolution of the Fo region, focused refinement with Fo was conducted by masking the Fo region. The refined structure achieved resolutions of 3.5–6.6 Å, confirming that the peripheral stalk always accompanies Fo a-subunits (Supplementary Fig. 4). Thus, the percentage of FoF1 with 0, 1, 2, and 3 a-subunits should correspond to that for peripheral stalks. The fractions of FoF1 with multiple a-subunits were small, and the ensemble average of peripheral stalk was only 1.26 per molecule, which is evidently lower than the expected value of ~2 per molecule based on the subunit stoichiometry analysis and the functional analysis of H+/ATP ratio. Although the exact reason for this discrepancy is unclear, it is highly likely that the peripheral stalk and the a-subunit dissociated due to the meniscus force and/or the interaction with the air/water interface during cryo-EM grid preparation27.

The composite cryo-EM maps of FoF1 with single, double, and triple peripheral stalks. The number represents the position of each peripheral stalk. Scale bar, 25 Å.
Interaction between the peripheral stalk and the δΔN-α fusion
Among the eight sub-classes, three structures contained a single copy of the peripheral stalk, each located at position Stalk 1, Stalk 2, or Stalk 3. These structures fit well with the reported three rotational state structure of the wild-type Bacillus PS3 FoF125, respectively (Supplementary Fig. 5), indicating the structural integrity of the binding of the b2 dimer to the F1 part via δΔN-α fusion. Slight differences were observed at the top of the F1 headpiece and on the side of the central stalk because the δΔN-α fused FoF1 lacks the N-terminal region of the δ subunit and the C-terminal helix of the ε subunit. The maps of the double- and triple-stalk FoF1 were compared with the corresponding maps of single-stalk FoF1, respectively (Supplementary Fig. 6). These maps were well fitted, indicating that multiple stalks have no significant structural constraints on the whole structure of FoF1.
The atomic models for triple-stalk FoF1 are shown in Fig. 6. The structure of the δΔN-α fusion region was well resolved, providing atomic details of its binding site with the b2 dimer (Fig. 6a, b). As designed, the three binding sites were almost identical, with structures similar to those found in wild type Bacillus PS3 FoF125 and other ATP synthases28 (Supplementary Fig. 7a). These observations reveal that the three b2 dimers are incorporated via the canonical interaction with δΔN-a fusion, suggesting the integrity of the peripheral stalks of the δΔN-α fused FoF1.

a Composite map and atomic models for the F1 and Fo regions of the triple-stalk FoF1. The middle regions of the b-subunits could not be modeled due to the lack of clear density. b The top view of the structure. The close-up view shows the side view of the interaction between δΔN-α fusion and the b2 dimer. c The view from the bottom of the structure shown in (a). d The superposition of the three a-subunits (green, purple, and blue) in the triple-stalk Fo.
Structure of the a-subunits
The structure and the position of the a-subunits of the triple-stalk FoF1 were investigated by comparing them with those in the wild-type Bacillus PS3 FoF125 (Fig. 6c and Supplementary Fig. 7b). The spatial intervals between the a-subunits are not perfectly symmetric because of the symmetry mismatch between the ring structures of F1 and Fo, that is, threefold versus tenfold. Each a-subunit interacted with three neighboring c-subunits, forming an a1c3 unit. Therefore, a total of nine c-subunits interacted with the a-subunits, leaving the remaining c-subunit at the open position (gap) between two a-subunits (Fig. 6c). As a result, the three a-subunits were not positioned exactly 120° apart from each other. The asymmetric positioning of the a-subunits is in good agreement with that suggested by the three states of the wild-type Bacillus PS3 FoF125. When the a-subunits in the triple-stalk FoF1 were compared to each other, Cα-RMSDs were 0.3–0.5 Å (Fig. 6d). In addition, the Cα-RMSDs estimated by superimposition of each a-subunit in the triple-stalk FoF1 with that in the corresponding state of the wild-type FoF1 were 0.9–1.0 Å. Moreover, superimposing the a1c3 units in the same manner yielded Cα-RMSDs of 1.0–1.2 Å (Supplementary Fig. 7c). Thus, at the current resolution, the overall structures of the three a-subunits in the triple-stalk FoF1 were essentially identical to each other and closely resembled that of the wild-type Bacillus PS3 FoF1. This suggests that all a-subunits are functional, which is consistent with the aforementioned biochemical analyses showing the enhanced H+/ATP ratio.
Structure of F1 part
The structure of the F1 portion of the triple-stalk FoF1 was investigated by comparing it with previously reported structures. The F1 structure was found to be very similar to that of the Bacillus PS3 FoF1-εΔC under uni-site catalysis conditions26, with one β subunit bound to ADP in a closed conformation (\({{{{\rm{\beta }}}}}_{{{{\rm{TP}}}}}^{C}\)) and two β subunits without bound nucleotide adopting open conformations (\({{{{\rm{\beta }}}}}_{{{{\rm{E}}}}}^{O}\), \({{{{\rm{\beta }}}}}_{{{{\rm{DP}}}}}^{O}\)) (Supplementary Fig. 8a, b). The purified δΔN-α fused FoF1 was prepared in nucleotide-free conditions. Therefore, it is highly likely that the bound ADP was endogenous. Notably, at a low-density threshold, weak map density was observed at the outer periphery of \({{{{\rm{\beta }}}}}_{{{{\rm{TP}}}}}^{C}\), which can be well fitted with the open β conformation of the nucleotide-depleted F1 in the Bacillus PS3 FoF1-εΔC26 (Supplementary Fig. 8c). This suggests that the cryo-EM map includes two conformations: closed and open. Other sub-class structures also exhibited similarly mixed maps.
Discussion
The present study provides insights into the design principles of ATP synthases using an engineering approach. Firstly, the δ subunit is the factor that defines the number of peripheral stalks. In this study, the N-terminal domain of δ, which breaks the structural pseudo-threefold symmetry by binding to the position on the symmetry axis, was deleted, and the C-terminal domain of δ was genetically fused to the N-terminal of the α subunit. As a result, up to three peripheral stalks were incorporated into the FoF1 complex. Structural analysis using cryo-EM revealed that the δΔN-α fusion and the b2 dimer adopted the canonical binding structure observed in the wild-type FoF1, except for the missing N-terminal domain of the δ subunit. This observation clearly shows that the N-terminal domain of the δ subunit determines the number of peripheral stalks per FoF1, restricting it to one by disrupting the structural symmetry.
Next, the number of a-subunits is determined based on the number of peripheral stalks. While cryo-EM analysis showed that some molecules lost peripheral stalks due to the detachment of the b2 dimer, the observed peripheral stalks always remained bound to the a-subunits, indicating stable binding between the a-subunit and the b2 dimer. Thus, the number of peripheral stalks determines the number of a-subunits. The structure of the triple-stalk FoF1 clearly showed that the c10 ring can accommodate up to three a-subunits, but not more than four due to spatial constraints. Considering that each a-subunit can interact with three c-subunits, accommodating four or more a-subunits in FoF1 would require the c-ring consisting of more than 12 c-subunits, along with further engineering on the F1 part, such as introducing a peripheral interaction site at the N-terminus of the β-subunit.
Another important finding of this study is the functional independence of the a-subunit, at least in terms of the coupling stoichiometry of H+ (see below). In the triple-stalk FoF1, all three a-subunits interacted with the c-ring. In addition, the structural features of the interaction agreed well with those observed in the wild-type FoF1 structures. This suggests that each of the three a-subunits is functional. SDS-PAGE analysis revealed that the samples used in this study had an average of two peripheral stalks and two a-subunits. Consistent with these results, analysis of the equilibrium pmf revealed an H+/ATP ratio that was doubled compared to that of the wild type. These results indicate that the coupling stoichiometry of the H+ ions is proportional to the number of a-subunits. The additivity in H+ stoichiometry means that each a-subunit tightly couples H+ translocation and rotation of the c-ring, regardless of the presence of other a-subunits. This is consistent with the half-channel model, which assumes that the probability of H+ translocation between the a and the c-subunits depends primarily on the relative position of these subunits, which explains well the functional independence of the a-subunits. Based on these considerations, we propose that the number of H+ ions transported coupled with rotation is determined not only by the number of c-subunits constituting the c-ring but also by the number of a-subunits as follows:
where \({N}_{{H}^{+}/{turn}}\), \({N}_{c}\), and \({N}_{a}\) represent the total number of H+ ions per turn, the number of c-subunit in the c-ring, and the number of a-subunit in Fo, respectively. However, one might point out the inconsistency between the biochemical results and the structural analysis with cryo-EM. The proportion of molecules with three peripheral stalks was extremely low, clearly lower than the average number suggested by the biochemical results. This is attributable to the dissociation of the peripheral stalks during sample preparation for cryo-electron microscopy. To confirm this point, it is necessary to develop multi-stalk FoF1 in which peripheral stalks stably bind to F1 and to more accurately analyze the correlation between the number of the a-subunit and the H+ stoichiometry.
As mentioned above, the increased H+/ATP ratio suggests the functional independence of the a-subunits. However, this does not guarantee kinetic independence among the a-subunits. Thus, an arising question is: ‘Can Fo with multiple a-subunits rotate the c-ring at the same rate as the wild-type Fo?’. In other words, ‘Do the a-subunits conduct H+ translocation without mutual interference?’ This is a reasonable question considering that, before each 36° rotation step of the c-ring, all a-subunits must complete the H+ translocation with the interacting c-subunits. Therefore, the time constant for each 36° rotation step is expected to be proportional to the number of a-subunits, meaning that the rate constant would decrease inversely.
Our biochemical experiments suggest that ATP hydrolysis-coupled proton pump activity was lower than that of the wild type (Supplementary Table 1). This aligns with the above expectation. However, other scenarios are possible: the engineered FoF1 may be more susceptible to the pmf progressively generated upon proton transport, and the engineered FoF1 may affect the catalytic activity of F1. Therefore, a quantitative and systematic analysis is necessary to verify this issue.
Regarding the lower activity of the mutant, a naive question could arise: ‘Why can the mutant still carry out the ATP synthesis reaction under lower pmf conditions despite its lower activity?’. The reason is as follows: the pmf required for FoF1 to synthesize ATP is determined by the pmfeq at which the synthesis and hydrolysis reaction rates are balanced. The present study shows that the engineered FoF1 with multiple a-subunits doubles the H+/ATP ratio, resulting in a halved pmfeq. Therefore, although the engineered ATP synthase has lower catalytic activity, the mutant enzyme can continue the ATP synthesis reaction under low pmf conditions where the wild-type enzyme is unable to synthesize ATP.
This study has shown that, in principle, ATP synthase has the capacity to expand its H+/ATP ratio to more than double. To date, experimentally confirmed H+/ATP ratios of FoF1 have only been ~3–4. Even structural estimates suggest that 5.0 is the maximum for photosynthetic bacteria. Regarding this point, an N-type ATPase—considered a subtype of the F-type—has been reported to have an exceptionally large c-ring composed of 17 c-subunits29. Although the function and overall structure of this enzyme remain unknown, its predicted H+/ATP ratio is 5.7. The present study demonstrates that the H+/ATP ratio can be significantly increased by genetically engineering ATP synthase to increase the number of a-subunits, without resorting to such a large c-ring. Such genetic mutations may have arisen over the course of evolution. Indeed, we found that the gene operon of ATP synthase from Acidaminococcus fermentans shows the gene fusion of the δ and the α subunits (Supplementary Fig. 9). In addition, the N-terminal domain of δ is missing. These features are well consistent with the δΔN-α fused FoF1 we designed. Thus, it is highly likely that A. fermentans FoF1 has a multi-stalk structure and can synthesize ATP under low pmf conditions. We also found that other species show similar features (UniProt ID: G4Q3K6, A0A1I2C5T3), suggesting more possibility of a multi-stalk FoF1 in nature. Thus, the present study suggests the unexpected diversity in the design principles of the FoF1 ATP synthase, which awaits further experimental verification.
From an engineering standpoint, the findings of the present study may provide future directions for cell engineering. So far, in microbial fermentation, attention has primarily been devoted to developing metabolic pathways and altering metabolic fluxes, while little effort has been made to optimize the intracellular concentrations and ratios of NAD(P)H and ATP, which are fundamental to cellular bioenergetics. When ATP synthesis is driven by oxidative phosphorylation, the amount of ATP produced depends on the extent of NADH oxidation. In such cases, an increase in the H+/ATP ratio translates into a higher NADH/ATP ratio. This could have a significant impact on microbial fermentations. In particular, it could enhance bioproduction in photosynthetic bacteria when ATP is the bottleneck factor22,30. To verify this possibility, it will be necessary to investigate how introducing ATP synthase with a modified H+/ATP ratio affects cell growth and product formation. Additionally, such insights are likely to shed light on the physiological role of naturally occurring multi-stalk FoF1, which may also exist in nature.
Methods
Preparation of FoF1
In this study, Bacillus PS3 FoF1-εΔC, which has a 10× His-tag at the N-terminus of the β subunit and lacks the inhibitory C-terminal domain of the ε subunit17,24 was used as the wild-type. The engineered δΔN-α fusion construct of Bacillus PS3 FoF1-εΔC lacks the full-length δ subunit and has a δΔN-α fused subunit in which the N-terminal domain (residues 2–104) of the δ subunit is deleted and its C-terminus is fused to the N-terminus (without Met) of the α subunit via a short linker. The wild-type and engineered FoF1s were expressed in E. coli DK8 cells, which lack endogenous FoF1 genes, by incubating in Super broth at 37 °C for 20 h. Cultured cells were suspended in a solution (10 mM HEPES, pH 7.5, 5 mM MgCl2, and 10% (v/v) glycerol) and disrupted by sonication. After removing the cell debris at 9100 × g for 45 min, membrane fraction was collected by centrifugation for 131,500 × g for 1 h at 4 °C. FoF1 was solubilized from the membrane fraction by adding 0.5% (w/v) LMNG (NG310; Anatrace, USA) and incubating for 30 min at 30°C. After centrifugation at 162,000 × g for 30 min, the solubilized fraction was applied to a Ni-Sepharose column pre-equilibrated with M buffer (20 mM potassium phosphate buffer and 100 mM KCl, pH 7.5) containing 0.005% LMNG. The column was washed with M buffer containing 20 mM imidazole and 0.005% LMNG, and FoF1 was eluted with M buffer containing 200 mM imidazole and 0.005% LMNG. The eluted FoF1 fractions were concentrated before being applied to a Superdex 200 Increase 10/300 column (Cytiva) equilibrated with gel filtration buffer (20 mM HEPES, pH7.5, 100 mM NaCl, and 0.005% LMNG). The peak fractions corresponding to FoF1 were collected and concentrated to 5–10 mg/mL, frozen with liquid nitrogen, and stored at −80 °C until use. The protein concentrations were determined using a BCA protein assay kit (Pierce) with bovine serum albumin as a standard. The molecular weight of the protein was calculated based on the sequence and subunit stoichiometry. For the δΔN-α fused FoF1, the molecular weight was calculated assuming an average of two peripheral stalks per FoF1 molecule.
Measurement of ATP synthesis/hydrolysis activity of FoF1
ATP synthesis/hydrolysis activity of FoF1 was measured using a luciferin-luciferase system at 25 °C, as described previously17. FoF1-reconstituted PLs were prepared as described17. Then, 300 µL of the PLs were mixed with 700 µL of acidic buffer containing 50 mM MES or HEPES buffer, 0.143–14.3 mM NaH2PO4, 6.7 mM KCl, 49 mM NaCl, 4 mM MgCl2, and 600 mM sucrose and NaOH to obtain the desired pH, and then ADP and valinomycin were added to a final concentration of 20-640 µM and 200 nM, respectively. After incubation for 10–24 h at 25 °C for acidification, base assay medium was prepared by mixing 25 µL of the luciferin/luciferase mixture (2× concentration of CLSII solution in ATP bioluminescence assay kit, Roche, and 5 mM luciferin), 800 µL of the base buffer (380 mM HEPES buffer, 0.1125–11.25 mM NaH2PO4, 5.63 mM KCl, 55 mM NaCl, 4 mM MgCl2, KOH to adjust K+ concentration and NaOH to adjust pH), 50-100 µL of ATP and ADP to obtain the desired concentration, and water to adjust the total volume to 900 µL and incubated for 10 min for equilibrium. Then, the 100 µL of acidified PLs was injected into the base assay medium to initiate the ATP synthesis reaction, and the ATP synthesis/hydrolysis activity was monitored with the luciferin/luciferase assay system using a luminometer (Luminescencer AB2200, ATTO). For calibrating luminescence light intensity to ATP concentration, 10 μL of 10 μM ATP was added. The rate was determined from the initial slope of the linear regression of the time courses. The ΔpH was obtained by subtracting pHin from pHout, which was determined by directly measuring the pH using a glass electrode. Transmembrane electrical potential, Δψ, was estimated from the Nernst equation.
Other assays
SDS-PAGE analysis was performed with 10–20% (w/v) gradient gels. The gels were stained with Coomassie Brilliant Blue (CBB Stain One Super, Nacalai Tesque, Japan) and imaged with a ChemiDoc Imaging System (BIORAD, USA). The band intensity of each subunit was measured using the gel analyzer tool of Fiji (ImageJ 1.54 f) software. ATPase activity measurements of PLs using an ATP regeneration system were performed at 25 °C in the ATPase assay solution (50 mM HEPES-KOH, pH 7.5, 100 mM KCl, 5 mM MgCl2, 2 mM ATP, 1 μg/ml Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone, 2.5 mM phosphoenolpyruvate, 100 µg/mL lactate dehydrogenase, 100 µg/mL pyruvate kinase and 0.2 mM NADH) as described previously31. ATP-driven H+-pumping activity was measured by quenching of ACMA (9-amino-6-chloro-2-methoxyacridine) fluorescence at 25 °C in PA4 buffer (10 mM HEPES-KOH, pH 7.5, 100 mM KCl, 5 mM MgCl2) supplemented with 0.3 μg/ml ACMA and 1.0 μg/ml FoF1-reconstituted PLs31.
Cryo-EM grid preparation and data collection
After adding 0.05% lysophosphatidylcholine (1-palmitoyl-2-hydroxy-sn-glycero-3-phosphocholine), 3.0 µL of purified protein (3–6 mg/mL) was loaded onto the glow-discharged Quantifoil R1.2/1.3 grids using a Vitrobot Mark IV (Thermo Fisher Scientific). Grids were blotted for 5 s with a blotting force of 15 under 100% humidity at 18 °C, and flash-frozen in liquid ethane. Data were collected using a 300 kV Titan Krios electron microscope (Thermo Fisher Scientific) with a Falcon 4i direct detector device camera with Selectris-X automated with EPU software. Images were recorded in electron counting mode by recording 50 movie frames with an exposure rate of 1.0 e−/Å2 per frame. The defocus range was 0.8–2.0 μm, and the original pixel size was 0.75 Å.
Cryo-EM data processing
All image processing steps were performed using cryoSPARC v4.3.0. Details of the image processing workflow are described in Supplementary Fig. 4. A total of 56,081 micrographs were first motion corrected, and the CTF was estimated by patch CTF estimation. Particles were manually picked, and templates for particle selection were generated from 2D classification. After template picking, the selected 4,316,627 particles were subjected to 2D classification. Then, further selections with ab initio 3D reconstruction, heterogeneous, homogeneous, and non-uniform refinements were performed. After non-uniform refinement, all particles were sequentially subjected to a focused 3D refinement using masks for each of the three peripheral stalks, including the N-terminal region of the α subunit, the C-terminal region of the δ subunit and the hydrophilic region of the b2-subunits. Each mask was generated from three rotational states of wild-type Bacillus PS3 FoF1 (PDBs 6N2Z, 6N30, and 6N2Y). Eight classes, including one to three peripheral stalks and three rotational states of FoF1, were identified. The three datasets were collected, merged and refined with non-uniform refinement, resulting in an overall resolution of 2.5–3.2 Å. Further refinement with the Fo mask resulted in a map of the Fo region with an improved resolution of 3.5–6.6 Å. The resolution was estimated using the FSC criterion of 0.143 threshold. The cryo-EM data collection and refinement statistics are shown in Supplementary Tables 2 and 3. The local resolution maps, FSC curves, orientation distribution plots and Model-to-map fits are shown in Supplementary Figs. 10–12.
Model building
Models were built and refined in COOT v0.9.8.93 and PHENIX v1.20.1-4487. using PDB 6N2Z, 6N30, and 6N2Y as the initial model. Validation statistics are shown in Supplementary Tables 2 and 3. A composite map was generated by combining the F1 region of the triple-stalk FoF1 map and the Fo map obtained through local refinement with the Fo mask, using UCSF ChimeraX v1.8 for illustration purposes only. Figures were prepared using PyMOL v2.4.0&2.5.2, UCSF Chimera v1.17.3, and UCSF ChimeraX v1.5&1.8. RMSD values for Cα-atoms were calculated using PyMOL 2.4.0 align command without outlier rejection.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The cryo-EM maps and models generated in this study were deposited to EMDB and PDB under the following accession codes: EMD-61339; EMD-61340; EMD-61341; EMD-61342; EMD-61343; EMD-61344; EMD-61345; EMD-61346; EMD-61347; EMD-61348; EMD-61349; EMD-61350; EMD-61351; EMD-61352; EMD-61353; EMD-61354, and 9JC1; 9JC2. Previously published structures used in this study are also available from PDB under the following accession codes: 6N2Z; 6N30, and 6N2Y. Source data are provided with this paper.
References
Walker, J. E. The ATP synthase: the understood, the uncertain and the unknown. Biochem. Soc. Trans. 41, 1–16 (2013).
Boyer, P. D. The ATP synthase-a splendid molecular machine. Annu. Rev. Biochem. 66, 717–749 (1997).
Yoshida, M., Muneyuki, E. & Hisabori, T. ATP synthase-a marvellous rotary engine of the cell. Nat. Rev. Mol. Cell Biol. 2, 669–677 (2001).
Guo, H. & Rubinstein, J. L. Cryo-EM of ATP synthases. Curr. Opin. Struct. Biol. 52, 71–79 (2018).
Kuhlbrandt, W. Structure and mechanisms of F-Type ATP synthases. Annu Rev. Biochem. 88, 515–549 (2019).
Stewart, A. G., Laming, E. M., Sobti, M. & Stock, D. Rotary ATPases-dynamic molecular machines. Curr. Opin. Struct. Biol. 25, 40–48 (2014).
Abrahams, J. P., Leslie, A. G., Lutter, R. & Walker, J. E. Structure at 2.8 A resolution of F1-ATPase from bovine heart mitochondria. Nature 370, 621–628 (1994).
Noji, H., Yasuda, R., Yoshida, M. & Kinosita, K. Jr. Direct observation of the rotation of F1-ATPase. Nature 386, 299–302 (1997).
Itoh, H. et al. Mechanically driven ATP synthesis by F1-ATPase. Nature 427, 465–468 (2004).
Rondelez, Y. et al. Highly coupled ATP synthesis by F1-ATPase single molecules. Nature 433, 773–777 (2005).
Vik, S. B. & Antonio, B. J. A mechanism of proton translocation by F1F0 ATP synthases suggested by double mutants of the a subunit. J. Biol. Chem. 269, 30364–30369 (1994).
Junge, W., Lill, H. & Engelbrecht, S. ATP synthase: an electrochemical transducer with rotatory mechanics. Trends Biochem. Sci. 22, 420–423 (1997).
Allegretti, M. et al. Horizontal membrane-intrinsic alpha-helices in the stator a-subunit of an F-type ATP synthase. Nature 521, 237–240 (2015).
Watt, I. N., Montgomery, M. G., Runswick, M. J., Leslie, A. G. W. & Walker, J. E. Bioenergetic cost of making an adenosine triphosphate molecule in animal mitochondria. Proc. Natl. Acad. Sci. USA 107, 16823–16827 (2010).
Ferguson, S. J. ATP synthase: From sequence to ring size to the P/O ratio. Proc. Natl. Acad. Sci. USA 107, 16755–16756 (2010).
Stock, D., Leslie, A. G. W. & Walker, J. E. Molecular architecture of the rotary motor in ATP synthase. Science 286, 1700–1705 (1999).
Soga, N., Kimura, K., Kinosita, K. Jr., Yoshida, M. & Suzuki, T. Perfect chemomechanical coupling of F(o)F(1)-ATP synthase. Proc. Natl. Acad. Sci. USA 114, 4960–4965 (2017).
Steigmiller, S., Turina, P. & Graber, P. The thermodynamic H+/ATP ratios of the H+-ATPsynthases from chloroplasts and Escherichia coli. Proc. Natl. Acad. Sci. USA 105, 3745–3750 (2008).
Petersen, J., Forster, K., Turina, P. & Graber, P. Comparison of the H+/ATP ratios of the H+-ATP synthases from yeast and from chloroplast. Proc. Natl. Acad. Sci. USA 109, 11150–11155 (2012).
Pogoryelov, D. et al. The oligomeric state of c rings from cyanobacterial F-ATP synthases varies from 13 to 15. J. Bacteriol. 189, 5895–5902 (2007).
Preiss, L. et al. The c-ring stoichiometry of ATP synthase is adapted to cell physiological requirements of alkaliphilic Bacillus pseudofirmus OF4. Proc. Natl. Acad. Sci. USA 110, 7874–7879 (2013).
Davis, G. A. & Kramer, D. M. Optimization of ATP synthase c-rings for oxygenic photosynthesis. Front. Plant. Sci. 10, 1778 (2019).
Cheuk, A. & Meier, T. Rotor subunits adaptations in ATP synthases from photosynthetic organisms. Biochem. Soc. Trans. 49, 541–550 (2021).
Masaike, T., Suzuki, T., Tsunoda, S. P., Konno, H. & Yoshida, M. Probing conformations of the beta subunit of F0F1-ATP synthase in catalysis. Biochem. Biophys. Res. Commun. 342, 800–807 (2006).
Guo, H., Suzuki, T. & Rubinstein, J. L. Structure of a bacterial ATP synthase. Elife 8, e43128 (2019).
Nakano, A., Kishikawa, J. I., Nakanishi, A., Mitsuoka, K. & Yokoyama, K. Structural basis of unisite catalysis of bacterial F(0)F(1)-ATPase. PNAS Nexus 1, pgac116 (2022).
D’Imprima, E. et al. Protein denaturation at the air-water interface and how to prevent it. Elife 8, e42747 (2019).
Spikes, T. E., Montgomery, M. G. & Walker, J. E. Structure of the dimeric ATP synthase from bovine mitochondria. Proc. Natl. Acad. Sci. USA 117, 23519–23526 (2020).
Schulz, S., Wilkes, M., Mills, D. J., Kuhlbrandt, W. & Meier, T. Molecular architecture of the N-type ATPase rotor ring from Burkholderia pseudomallei. EMBO Rep. 18, 526–535 (2017).
Nogales, J., Gudmundsson, S., Knight, E. M., Palsson, B. O. & Thiele, I. Detailing the optimality of photosynthesis in cyanobacteria through systems biology analysis. Proc. Natl. Acad. Sci. USA 109, 2678–2683 (2012).
Suzuki, T., Ueno, H., Mitome, N., Suzuki, J. & Yoshida, M. F. 0) of ATP synthase is a rotary proton channel. Obligatory coupling of proton translocation with rotation of c-subunit ring. J. Biol. Chem. 277, 13281–13285 (2002).
Acknowledgements
We thank all the members of our laboratory for their comments. We also thank N. Soga for technical advice on biochemical experiments. We also thank M. Kawasaki, A. Ikeda, S. Inaba, T. Moriya and the staff at the KEK Structural Biology Research Center for their assistance in collecting and analyzing cryo-EM data. We also thank T. Matsui for technical assistance with the biochemical experiments. This study was supported in part by a Grant-in-Aid for Scientific Research on Innovation Areas (JP21H00388 to H.U.), a Grant-in-Aid for Challenging Research (Exploratory; JP23K18092 to H.U.), a Grant-in-Aid for Scientific Research (B) (JP24K01987 to H.U.), and a Grant-in-Aid for Scientific Research (S) (JP19H05624 to H.N.) from JSPS, and a Research Grant from Human Frontier Science Program (Ref. No: RGP0054/2020 to H.N.), and a JST ASPIRE Program (JPMJAP24B5 to H.N.), and a Research Support Project for Life Science and Drug Discovery (Basis for Supporting Innovative Drug Discovery and Life Science Research (BINDS)) from AMED under Grant Number JP23ama121013 (support number 4318) to T.M. and JP21am0101071 (support number 3071) to T.S.
Author information
Authors and Affiliations
Contributions
H.U., K.Y. and R.M. performed the biochemical experiments. N.H-S. and N.A. performed the cryo-EM analysis. T.S. and T.M. provided the technical support and conceptual advice. H.U. and H.N. conceived and supervised the study and wrote the manuscript. All the authors discussed the results and commented on the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Yan Gao, Stephan Wilkens, and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ueno, H., Yasuda, K., Hamaguchi-Suzuki, N. et al. Engineering of ATP synthase for enhancement of proton-to-ATP ratio. Nat Commun 16, 5410 (2025). https://doi.org/10.1038/s41467-025-61227-w
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41467-025-61227-w
