
Abstract
The rise of drug-resistant fungal pathogens, including Candida auris, highlights the urgent need for innovative antifungal therapies. We have developed a cost-effective platform combining microbial extract prefractionation with rapid mass spectrometry-bioinformatics-based dereplication to efficiently prioritize previously uncharacterized antifungal scaffolds. Screening C. auris and Candida albicans reveals coniotins, lipopeptaibiotics isolated from Coniochaeta hoffmannii, which are undetectable in crude extracts. Coniotins exhibits potent activity against critical priority fungal pathogens listed by the World Health Organization, including C. albicans, Cryptococcus neoformans, multidrug-resistant Candida auris, and Aspergillus fumigatus, with high selectivity and low resistance potential. Coniotin A targets beta-glucan, compromising fungal cell wall integrity, remodelling, and sensitizing C. auris to caspofungin. Identification of its hybrid polyketide synthase–nonribosomal peptide synthetase biosynthetic gene cluster facilitates discovering structurally diverse lipopeptaibiotics. Here, we show that natural product prefractionation enables the discovery of previously hidden bioactive scaffolds and introduces coniotins as candidates for combating multidrug-resistant fungal pathogens.
Similar content being viewed by others
Introduction
Fungal diseases represent a significant threat to public health, affecting over a billion people globally and resulting in more than 2.5 million deaths annually1,2, surpassing mortality rates from tuberculosis and malaria3. Developing antifungal therapies is particularly challenging due to the overlapping cell components, as well as conserved metabolic and biochemical pathways between fungi and their human hosts, leading to a limited repertoire of available treatments for invasive fungal infections4. The emergence of drug-resistant fungal pathogens, such as Candida auris, which has caused recent outbreaks in healthcare settings, further exacerbates this issue5,6. C. auris is recognized as a critical priority pathogen by the World Health Organization (WHO)7 and has been classified as an urgent threat by the US Centers for Disease Control and Prevention (CDC)8. C. auris isolates resistant to all existing drugs are increasingly common9. Unlike other Candida species, C. auris efficiently colonizes the skin, leading to rapid nosocomial transmission and systemic infections with mortality rates of 40–60%6,10. The urgent need for next-generation antifungal drugs is critical to prevent further failures in controlling fungal infections within hospitals and healthcare facilities.
Natural products and their derivatives have been an invaluable source of therapeutic agents, ranging from antibiotics to anticancer agents, thanks to their structural novelty, chemical complexity, and intrinsic bioactivity; consequently, natural products hold promise as leads for antifungal drug discovery11,12. The traditional compound-first discovery strategy using phenotypic cell growth inhibition screens of crude extracts of bacteria and fungi contributed to over half of the antibiotics and antifungal drugs in everyday use today13. However, the rediscovery of well-known chemical scaffolds, including antifungal classes such as the polyenes, is a challenge given the phenotypic dominance of highly expressed common scaffolds in natural product extracts14,15.
Owing to the rapid advancement of DNA sequencing technology16,17, genome sequences of natural product producers have revealed large numbers of untapped biosynthetic gene clusters (BGCs) of metabolites, predicting that traditional extract screens vastly undersample the available chemical space of natural products18. A genes-first genome mining strategy, coupled with advanced molecular technologies, is leading to the discovery of novel chemical entities19,20,21. However, predicting the biological activities of the natural products discovered based on bioinformatic analyses is difficult, even with known compound classes, which limits their application in drug development.
An orthogonal approach to access untapped natural product chemical space entails prefractionating crude natural product extracts before biological testing, thereby uncovering bioactive compounds that are often low in abundance or masked by other activities within complex mixtures22,23,24. This approach typically improves the hit rate in phenotypic screens and streamlines dereplication by concentrating minor components, reducing sample viscosity for automated platforms, and separating ubiquitous nuisance compounds from rare metabolites23,25. To further broaden accessible chemical diversity prior to fractionation, the one strain, many compounds (OSMAC) approach was integrated, which systematically modulates cultivation parameters to enhance microbial metabolite diversity26. By altering media composition, aeration, and culture formats, OSMAC enables the activation of silent or poorly expressed BGCs, facilitating the discovery of cryptic metabolites27,28. Although semipreparative HPLC-based fractionation approaches remain widespread, their scalability in academic settings is challenging due to the substantial resources required for the dedicated equipment and personnel to prepare the libraries29. In contrast, reversed-phase flash chromatography offers a scalable alternative compatible with resource‑constrained laboratories23,24.
Building on these insights, we developed a cost-effective prefractionation library (PFL) using medium-pressure reverse-phase separation, optimized for broad deployment in academic labs26. Here, we report a pilot application of this platform for antifungal discovery targeting C. auris and Candida albicans, coupling high-resolution mass spectrometry and tandem mass spectrometry (HRMS/MS) with genome-mining of the relevant BGCs for rapid dereplication. This workflow efficiently identified and triaged known antifungal families (e.g., enniatins, surfactins, and tunicamycins) and, critically, enabled the prioritization and discovery of a lipopeptaibiotic antifungal coniotin, which exhibits broad activity against multidrug-resistant fungal pathogens and is phenotypically undetectable in the crude extract. Unlike channel-forming lipopeptides, coniotin targets the fungal cell wall by binding β-glucan, disrupting cell wall remodeling, and sensitizing resistant C. auris to caspofungin, with promising selectivity and a low potential for resistance development. The identification of the linear NRPS-PKS hybrid gene cluster and the proposed coniotin biosynthetic pathway facilitate the exploration of structurally diverse lipopeptaibiotics from related clusters. This work highlights the utility of the PFL platform coupled with rapid HRMS/MS dereplication for the discovery of previously undetectable antifungal scaffolds and identifies coniotins as structurally unique candidates that target fungal cell wall integrity in important fungal pathogens.
Results
Antifungal screening of a fractionated natural product library
The PFL was derived from the medium-pressure reverse phase separation of fermentation methanolic extracts of our in-house collection of bacteria and fungi, resulting in eight fractions of metabolites sorted by hydrophilicity for each strain26. A total of 3048 fractions and the corresponding 381 crude extracts were screened against C. auris CBS10913 in duplicate, identifying 43 hits that showed growth inhibition from fractions, while only 12 hits were from crude extracts (Fig. 1a). Similarly, a parallel screen against C. albicans ATCC90028 yielded 28 active fractions and 9 crude hits. To identify broad-spectrum antifungal agents, nine hits shared across the two PFL screens were selected for further validation (Fig. 1b). Among these, antifungal bioactivity from WAC11084, WAC11113, and WAC11161 was exclusively observed in the fractionated samples, whereas their corresponding crude extracts showed minimal activity and were not identified during the cross-species hit screening (Fig. 1b and Supplementary Fig. 1a). These results highlight the advantage of screening fractionated extracts to improve hit detection.

a Overview of a rapid and risk-minimized approach for discovering novel antifungal natural products. The flowchart outlines a streamlined pipeline, starting from the construction of a PFL, followed by high-throughput screening against target organisms (C. albicans and C. auris), rapid dereplication using tandem mass spectrometry (MS2) fingerprinting and bioinformatics analysis, structure determination, off-target assessment in mammalian cells (HEK human embryonic kidney cells + RBC: red blood cell), broad-spectrum bioactivity evaluation, rapid therapeutic assessment using high-throughput animal models, and characterization of the mechanism of action (MOA). Created with BioRender.com and published under a BioRender CC-BY publication license. b Scatter plots illustrating high-throughput screening results of crude methanolic extracts and PFL against C. albicans (y-axis) and C. auris (x-axis). Colored circles within the red box represent active hits: crude extracts (blue) or fractions (red) that inhibit Candida growth by at least 75% compared to the untreated control. Active hits with their WAC and fraction identities are shown in Supplementary Fig. 1a, with validation results presented in Supplementary Fig. 1b. The source data is provided as a Source Data file.
Verification and rapid identification of known scaffolds
The WAC isolates that produced the nine anti-Candida hits were successfully re-grown, extracted, and re-fractionated to confirm their activity. The crude extracts were separated on a C18 Combiflash column, and 24 fractions were collected from each run, resulting in 216 fractions generated in approximately 4 h of instrument time (Supplementary Fig. 1b). All fractions were tested against C. auris CBS10913, but anti-Candida activity could not be reproduced for three of the nine isolates (WAC10997, WAC11024, and WAC11113), a common occurrence when re-evaluating wild actinomycete isolates from high throughput screening (Supplementary Fig. 1b).
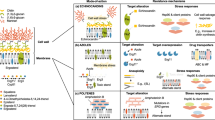
To rapidly identify known compound scaffolds and prioritize novel structures, HRMS coupled with tandem mass spectrometry (MS2) was employed for the tentative structural assignment of active metabolites. The MS and MS2 fragmentation spectra served as molecular fingerprints, enabling the construction of molecular networks that link to known compound classes by calculating spectral similarity relationships within the Global Natural Products Social molecular networking (GNPS, https://gnps.ucsd.edu/) platform (Fig. 2a)30,31. Spectral alignment of precursor and fragment ions identified a series of candidate enniatin analogs from the active fractions of isolate WAC11175, which was taxonomically assigned to Fusarium proliferatum (Fig. 2a). The identified analogs included enniatin A1 ([M + H]+ at m/z 668.4519), enniatin B ([M + H]+ at m/z 640.4207), and enniatin B1 ([M + H]+ at m/z 654.4369) (Fig. 2a), which are well-characterized nonribosomal peptide mycotoxins32,33 now found to exhibit significant activity against C. auris (Supplementary Fig. 1b). In support of this MS-based identification, genome analysis of WAC11175 uncovered a BGCs encoding a nonribosomal cyclic peptide synthetase with 98% sequence identity to a characterized enniatin synthetase (Fig. 2b and Supplementary Table 1)33. The predicted adenylation domain specificities correspond to D-2-hydroxyisovaleric acid and L-valine, as found in enniatin B, although the synthetase is known to accommodate a broader range of L-amino acid substrates, thereby enabling the production of diverse enniatin analogs (Supplementary Table 7)33,34,35. To validate this proof-of-concept approach for coupling HRMS2 with bioinformatics-based rapid identification of active compounds, we purified enniatin B and confirmed its structure through 1-D and 2-D-NMR analysis, thereby verifying the reliability of the method (Supplementary Table 2 and Supplementary Data 1).

a Identification of enniatin from active fractions of WAC11175 through GNPS based on HRMS and MS/MS data. (i) The enniatin GNPS molecular network, with MS/MS fragment ion matches between the sample (blue) and reference enniatin B1 (black)(inset). (ii) HR-LCMS analysis for enniatins detected in WAC11175 active fractions, including structures and representative HRMS spectra of enniatin A1 (iii), enniatin B (v), and enniatin B1 (vii). Corresponding MS/MS spectra acquired at 20 eV and fragmentation analyses are shown for enniatin A1 (iv), B (vi), and B1 (viii). N–Me–Val, N–Me–Ile, and Hiv represent N-Methyl-L-valine, N-Methyl-L-isoleucine, and 2-hydroxyisovaleric acid, respectively. b Identification of the enniatin BGC in WAC11175 genome and depiction of its biosynthesis. The enniatin synthetase Esyn1 contains the following domains: C (condensation), A (adenylation), P (phosphopantetheine attachment site), and nMT (N-methyltransferase). L-valine and D-hydroxycarboxylic acid precursors are activated at the A domain, and the building blocks are transferred between modules via P-domains. The final condensation, cyclization, and product release are catalyzed by the C-domains. c Identification of surfactin from active fractions of WAC11084. (i) GNPS molecular network, constructed from HR-MS/MS data, showing MS/MS fragment ion matches (inset panel) between the identified entity (blue) and the reference surfactin C (black). MS/MS fragmentation patterns and structures are shown below: (ii) Surfactin A ([M + H]+, m/z 1008.6565) fragments into main ions at 667.4, 568.3, and 455.3; (iii) Surfactin B ([M + H]+, m/z 1022.6731) into 909.5, 681.4, and 582.4; (iv) Surfactin C ([M + H]+, m/z 1036.6907) into 923.6, 695.4, and 596.4. Diagnostic fragment ions corresponding to specific amino acid sequences were observed, including m/z 227.1750 [Leu + Leu + H]+, 229.1145 [Asp + Leu + H]+, and 441.2699 [Leu + Asp + Val + Leu + H]+. d Genetic organization of the surfactin BGC in WAC11084 and proposed biosynthesis. The surfactin synthetase complex comprises SrfA, SrfB, and SrfC, which assemble the heptapeptide core, while SrfD serves as a key regulatory component. Functional domains include C (condensation), A (adenylation), T (thiolation), E (epimerization), and TE (thioesterase), the latter mediating cyclization and product release.
Using this strategy, we identified the cyclic lipopeptides surfactin A ([M + H]+ at m/z 1008.6565), surfactin B ([M + H]+ at m/z 1022.6731), and surfactin C ([M + H]+ at m/z 1036.6907) in the active fraction from isolate WAC11084, identified as Bacillus velezensis, (Fig. 2c, d and Supplementary Table 1), whose antifungal activity had been masked in crude extracts (Supplementary Fig. 1a). The nonribosomal peptide synthetase (NRPS) cluster was identified in the genome sequence of WAC11084 and directs the synthesis of the amphiphilic cyclic lipopeptide, which imparts strong surfactant properties and broad-spectrum biological activities36.
Similarly, we observed that Streptomyces microflavus strains WAC1325 and WAC1490 produce tunicamycins, displaying anti-Candida activity in the active fractions (Supplementary Figs. 1b and 2a). Tunicamycins are well-established antifungal agents that inhibit the unfolded protein response by blocking protein N-glycosylation, thereby inducing ER stress in fungi37. Genome sequencing of WAC1325 and WAC1490 confirmed the presence of a tunicamycin BGC, supporting this discovery (Supplementary Fig. 2b and Supplementary Table 1)38. We also detected the guanidinopolyol cyclic macrolides niphimycin C ([M + H]+ at m/z 1142.7335) and D ([M + H]+ at m/z 1228.7386) from active fractions produced by Streptomyces violaceusniger WAC5858 and confirmed the presence of the expected BGC in the sequenced genome (Supplementary Fig. 2c–e and Supplementary Table 1)39.
Isolation and characterization of lipopeptaibiotic coniotins
In contrast to known compounds, the antifungal metabolites present in the active fractions of Coniochaeta fungal species WAC11161 could not be identified using HRMS data analysis in GNPS, suggesting the presence of unique structural features not previously characterized. Activity-guided isolation identified the active compound 1 ([M + H]+ at m/z 2057.2609) with a molecular formula of C98H170N21O26, as determined by HRMS (Fig. 3a). 1H NMR analysis revealed a peptide structure rich in highly methylated amino acids (Supplementary Table 3 and Supplementary Data 2), with further structural confirmation achieved through HR mass fragmentation and Collision-Induced Dissociation (CID, including MS2 and in-source CID, MS3)40,41. CID-MS2 of the parent ion [M + H]+ at m/z 2057.26 and the doubly charged ion [M + 2H]2+ at m/z 1029.13 (Fig. 3a, b and Supplementary Fig. 3i) identified characteristic fragment ions ([M–H2O + H]+ at m/z 85.05), indicative of rare non-proteinogenic α-aminoisobutyric acid (Aib), a hallmark of fungal peptaibiotics essential for their stable α-helical structures42. 1H-, 13C-, and 2-D NMR spectra (COSY, HSQC, HMBC, NOESY, and 1H-15N-HSQC) identified six 2-Aib residues, five Ala, four Iva, three β-Ala, one Pro, one Leu, and one Asp residue, the latter being rare among peptaibiotics, with the 2-methyl-3-oxotetradecanoyl linked to the Pro residue (Fig. 3b, Supplementary Table 3, and Supplementary Data 2).

a HR mass spectrum of compound 1 obtained with QToF mass spectrometry, showing the [M + H]+ ion at m/z 2057.2609. In-source fragmentation produced ions at m/z 506.36, 602.31, 690.48, 843.45, 931.62, 1126.65, and 1215.81. b MS/MS analysis of compound 1 using CID combined with a product ion scan (MS/MS) of nominal m/z 2057.26. The precursor ion is indicated with a blue square. The structure of the lipopeptide 1 (termed coniotin A) is displayed above, along with its collision-induced fragmentation pattern, which corresponds to the detected b ions in the MS/MS spectrum. The b ions are labeled in blue, and the y ions are labeled in red. c The structures of antifungal lipopeptaibiotics analogs 1, 2, 3, and 4, identified from the Coniochaeta fungus WAC11161, termed coniotin A, B, C, and D. d HR mass spectrum of coniotin B obtained with QToF mass spectrometry, showing the [M + H]+ ion at m/z 2056.2781 (calculated for C98H171N22O25, 2056.2780). e HR mass spectrum of coniotin C obtained with QToF mass spectrometry, showing the [M + H]+ ion at m/z 2043.2464 (calculated for C97H168N21O26+, 2043.2464) and the [M+Na]+ ion at m/z 2065.2255. f HR mass spectrum of coniotin D obtained using QToF mass spectrometry, showing the [M + H]+ ion at m/z 2042.2689 (calculated for C97H169N22O25+, 2042.2624), the [M+Na]+ ion at m/z 2064.2536, and the [M + K]+ ion at m/z 2080.2316.
In-source fragmentation produced ions at m/z 506.36, 602.31, 843.45, 931.62, 1126.65, and 1215.81 (Fig. 3a), with m/z sums of 931.62 + 1126.65 and 843.45 + 1215.81 matching the molecular weight of 1, identifying them as N-terminal and C-terminal fragments, respectively. The fragment at m/z 336 was assigned to the prolyl-2-methyl-3-oxo-tetradecanoic acid moiety (Pro-MOTDA) (Fig. 3b), and the presence of the β-keto acid, 2-methyl-3-oxo-tetradecanoic acid (MOTDA), was further confirmed by analyzing an ethyl acetate extract of hydrolyzed 140,43,44. CID-MS2 analysis of the parent ion (MS2), along with N-terminal, C-terminal, and intermediate peptide fragments (MS3)42 identified the amino acid sequence as MOTDA-Pro–Aib–Aib–Aib–Iva–βAla–Ala–Iva–Ala–Leu–Iva–βAla–Ala–Iva–βAla–Ala–Aib–Aib–Aib–Ala–Asp–OH, a unique lipopeptaibiotic (Fig. 3b and Supplementary Fig. 3a–d). While α-cleavage of β-Ala ions was significantly suppressed, digestion with the non-specific protease papain generated peptide fragments ([M + H]+ at m/z 1303.85, 843.45, and 614.38), significantly facilitating sequence resolution (Supplementary Fig. 3b–d)45. Additionally, increasing CID to 50 eV enabled cleavage at βAla6–Ala7 and Ala9–Leu10, confirming the amino acid sequence at the single-residue level (Supplementary Fig. 3d–e). Amino acid stereochemistry was confirmed by Marfey’s analysis, with HPLC separation of the modified, hydrolyzed peptide revealing seven amino acids (Pro, Aib, βAla, Ala, Iva, Leu, and Asp) in 1 and identifying the absolute configurations as L-Pro, L-Ala, L-Leu, L-Asp, and D-Iva (Supplementary Fig. 3f)46.
A second active analog, compound 2 ([M + H]+ at m/z 2056.2781, [M + 2H]2+ at m/z 1028.64), was purified with a molecular formula of C98H170N22O25 (cal [M + H]+ at m/z 2056.2780) (Fig. 3c, d and Supplementary Fig. 3i). The mass difference of 0.983 between compounds 1 and 2, characteristic of –NH2 vs –OH, suggests an Asn at the N-terminus instead of Asp. CID-MS2 analysis confirmed an identical amino acid sequence to 1, with Asp replaced by Asn at the C-terminus (Supplementary Fig. 3g), which was further confirmed by Marfey’s analysis. Additionally, a non-methylated analog, compound 3 ([M + H]+ at m/z 2043.2464, [M + 2H]2+ at m/z 1028.64) was identified (Fig. 3c, e and Supplementary Fig. 3j), featuring a 3-oxotetradecanoic acid (OTDA) at the N-terminus in place of 1’s MOTDA (Fig. 3c). Each b fragment displayed an ion loss of 14 (–CH3 + H), including the terminal b1 fragment, identified as Pro-OTDA, at m/z 322 (Supplementary Fig. 3h). Similarly, analog 4, featuring an Asn at the N-terminus in place of Asp as in compound 3, was also identified (Figs. 3c, f and Supplementary Fig. 3j). Accordingly, we designated this group of lipopeptaibiotics, 1, 2, 3, and 4, derived from the Coniochaeta fungi as coniotin A, B, C, and D.
Coniotins exhibits selective antifungal activity
Lipopeptides exhibit a wide range of biological activities due to their unique structures47,48. We assessed the antifungal activity of coniotin A in comparison to first-line antifungals (caspofungin, amphotericin B, and fluconazole), revealing broad-spectrum activity against Candida species (C. albicans, C. parapsilosis, and C. tropicalis), Cryptococcus neoformans, Nakaseomyces glabratus, and Saccharomyces cerevisiae (Table 1). Notably, coniotin A demonstrated potent activity against multidrug-resistant C. auris and the mold Aspergillus fumigatus, both identified as critical threats on the WHO Fungal Priority Pathogens List7, surpassing the efficacy of caspofungin and fluconazole. Interestingly, coniotin A also enhanced the efficacy of caspofungin, significantly reducing its MIC against refractory C. auris to the CLSI breakpoint of 2 μg/mL (Fig. 4a)49.

a Coniotin A (CNA) synergizes with caspofungin (CAP) in Candida species. Checkerboard assays depicted as heatmaps show the average growth of biological duplicates, normalized to controls without compounds. The potentiation of coniotin A and caspofungin was evaluated against C. auris CBS12775 and C. albicans ATCC90028. Relative growth is depicted by color, as indicated by the scale bar in the bottom right. Fractional Inhibitory Concentration Index (FICI) values, calculated as described in the Methods, are shown in the top right corner of each checkerboard. FICI values below 0.5 denote synergistic interactions. b Rapid assessment of the therapeutic potential of coniotin A using a high-throughput Caenorhabditis elegans–C. albicans infection model. C. elegans were infected with C. albicans ATCC90028 and treated with various concentrations of coniotin A. Representative images show worms treated with DMSO (i, negative control), 1 µg/mL coniotin A (ii), and 8 µg/mL coniotin A (iii). Scale bar = 0.2 mm. Experiments were independently repeated three times with similar results. c Survival of C. elegans infected with C. auris CBS 12775 and treated with amphotericin B (AMB), coniotin A (CNA), or vehicle (DMSO). Twenty-five worms per condition were monitored over 48 h in three biologically independent experiments. Survival curves were analyzed using the Kaplan–Meier method. Statistical significance was assessed by a two-sided Log-rank (Mantel–Cox) test (χ² = 39.48, df = 1, p < 0.0001), with confirmation by the Gehan–Breslow–Wilcoxon test (χ² = 28.64, p < 0.0001), comparing CNA (1× MIC) treatment to the DMSO control group. d Proteolytic stability of coniotin A compared to LL-37 in the presence of trypsin (Try), neutrophil elastase (Ela), and human serum (HS). Peptides (40 μM) were incubated with trypsin, elastase, or 50% human serum at 37 °C for up to 12 h. Residual intact peptide was quantified by HR LC-MS and normalized to the untreated control (0 h). Data represent mean ± SD from three independent experiments. Figure 4c, d, Source data and full statistical results are provided as a Source Data file.
The therapeutic potential of coniotin A was assessed using the non-mammalian model C. elegans, a system well-suited for studying Candida interactions due to its intestinal similarities to mammals and ease of infection50. At a concentration of 8 μg/mL, coniotin A significantly reduced C. albicans infections in C. elegans (Fig. 4b). Building upon these results, further in vivo efficacy was evaluated against multidrug-resistant C. auris51. Coniotin A effectively extended the lifespan of C. elegans pre-infected with C. auris CBS 12775, a strain resistant to caspofungin and fluconazole, by approximately 30% over two days, while untreated nematodes succumbed to infection within 40 h (Fig. 4c). Transmission electron microscopy (TEM) demonstrated the interactions between the pathogen and host during infection, revealing that C. auris invaded through the intestinal wall, resulting in the disintegration of the brush border of the gastrointestinal tract (Supplementary Fig. 4a)52. Unlike membrane-perturbing lipopeptides such as iturin A53, coniotin A exhibited no hemolytic or antibacterial activity, indicating a different mode of action, with its target being absent in prokaryotes and human erythrocytes (Table 1, Supplementary Table 4, and Supplementary Fig. 4b).
Interestingly, amidation of the C-terminal Asp slightly reduced the antifungal efficacy of coniotin B against C. albicans ATCC 90028, C. neoformans H99, and C. auris strains CBS12766 and CBS12776 (Table 1). Furthermore, coniotin B demonstrated greater hemolytic and cytotoxic activity compared to coniotin A (Table 1 and Supplementary Fig. 4b, c), highlighting the critical role of Asp in influencing its bioactivity, despite the rarity of Asp residues in peptaibiotics due to their unfavorable effects on α-helix stabilization54.
Proteolytic degradation remains a key limitation for peptide-based therapeutics; however, the incorporation of unnatural and D-amino acids can substantially enhance resistance to proteolysis55,56. To further assess the proteolytic stability and potential bioavailability of coniotin A, we evaluated its susceptibility to endogenous human proteases such as trypsin and neutrophil elastase, and pooled human serum. Both trypsin and elastase (10 ng/μL) rapidly hydrolyzed 400 μM Nα-Benzoyl-L-arginine 4-nitroanilide and N-Succinyl–Ala–Ala–Ala–p-nitroanilide within 1 h (Supplementary Fig. 4d), confirming robust enzymatic activity. Under these conditions, coniotin A displayed markedly improved stability compared to the human antimicrobial peptide LL-37 (Fig. 4d). Proteolytic degradation was more pronounced in 50% human serum, where approximately 27% of coniotin A remained after 12 h of incubation, reflecting the higher protease complexity of this biological matrix.
Coniotin A targets β-glucan, impairing cell wall integrity
Fungal lipopeptaibols uniquely contain the nonstandard amino acid Aib, which confer an α-helix structure that enhances bioactivity and metabolic stability, allowing them to form ion channels, permeabilize cell membranes, and act as active agents57,58. To elucidate the mechanism of action (MOA) for coniotin A, a serial passage assay was performed to select C. albicans ATCC90028 and C. neoformans H99 mutants resistant to coniotin A, as resistance mutations typically arise in target genes59. However, after 20 serial passages under sub-MIC conditions, no resistance emerged, and all colonies remained susceptible to coniotin A, suggesting a low mutation rate or unstable resistance. Similarly, screening over 5000 C. albicans heterozygous (HET) deletion mutants revealed no strains resistant to coniotin A (Supplementary Fig. 5a), suggesting it, like amphotericin B, may target essential cellular components rather than specific protein targets, making resistant mutants exceedingly rare and its precise target(s) unclear60.
To investigate the target location of coniotin A and determine whether it requires intracellular entry to exert its activity, we first assessed its intracellular accumulation61. The membrane- and cell wall-perturbing lipopeptide iturin A53 and β-(1,3)-glucan synthase inhibitor caspofungin62 were used as controls, both demonstrating measurable accumulation, with caspofungin displaying significantly lower levels (Fig. 5a). In contrast, no intracellular accumulation of coniotin A was detected in either pathogen, indicating that it exerts its activity at the cell surface rather than intracellularly (Fig. 5a). Chitin is an essential component of the fungal cell wall located in the inner layer and has tightly regulated synthesis that can be induced in response to β-glucan damage, aiding in survival against cell wall stressors63,64. The levels of chitin were significantly elevated in response to coniotin A in C. albicans, C. auris, and C. neoformans, similar to the response observed with the β-(1,3)-glucan synthase inhibitor caspofungin (Fig. 5b–d). In addition to increased chitin production and thickened septa, treatment with coniotin A altered the morphology of C. albicans, leading to clusters of enlarged and elongated cells (Supplementary Fig. 5b).

a Intracellular accumulation of coniotin A (CNA), caspofungin (CAP), and iturin A (ITA) in C. auris CBS10913 (Cau) and C. albicans ATCC90028 (Cal) after 10 min treatment. Data represent mean ± SD from four biological replicates. b Chitin levels in fungal cells after 4 h treatment with ½ MIC of CNA, CAP, or vehicle control (DMSO). Calcofluor white (CFW)-stained cells were analyzed by epifluorescence microscopy; bars represent the mean, with individual cell values (from three independent biological replicates) overlaid as dots. Statistical significance was determined using unpaired two-tailed t-tests with Welch’s correction; all comparisons between CNA and DMSO yielded P < 0.000001. c Representative CFW-stained images corresponding to (b). Scale bars, 10 µm. d Schematic of the Candida cell wall. Glycosylphosphatidylinositol-anchored mannoproteins form the outer layer linked to a β-glucan–chitin core. Echinocandins target the glucan synthase Fks1, compromising wall integrity. Created with BioRender.com. e Mannoprotein staining in C. albicans (ConA–Alexa Fluor 647) after treatment. Confocal 3D projections showing cell wall disruption (blue arrows). Scale bars, 5 µm. f Quantification of cell perimeter from treatments as in (e), based on ~150 cells (ImageJ). Significance was determined by a two-tailed unpaired Student’s t-test: exact P values are shown in the figure. g CNA binding to β−1,3-glucan and chitin assessed by mass spectrometry after pull-down assay. Relative abundance shown for bound (Glu-B, Chi-B) and unbound (Glu-S, Chi-S) fractions. Data represent mean ± SD of three biological replicates; each dot corresponds to one replicate. h CNA inhibits β−1,3-glucanase (GCase) digestion. Hexa-glucose oligosaccharides released from laminarin (a β−1,3-glucan) after 0.5 h hydrolysis with or without CNA (64, 128, or 256 µg/mL) were quantified by mass spectrometry. Data from three independent replicates are shown as dots; values are mean ± SD. Significance was determined using two-tailed unpaired t-tests with Welch’s correction. Exact P values are shown in the figure. i CNA inhibits β−1,3-glucan activation of Limulus factor G in a dose-dependent manner. β−1, 3-glucan (Glu) was incubated without/with CNA (0.625, 5, or 40 µg/mL; 1×, 8×, and 64×) and assayed using Glucatell®. Reactions were monitored at 405 nm for 1 h. Data represent mean ± SD from n = 3 independent biological replicates. j TEM of C. auris and C. neoformans ± CNA (½ MIC). CNA induces membrane detachment (blue arrowheads), loss of integrity (orange), and cell wall thickening. G + C β-glucan/chitin, M mannoprotein, N nucleus, m mitochondria. Scale bars, nm. Figures 5b, f–i, Source data and full statistical results are provided as a Source Data file.
Further visualization of the impaired cell surface was achieved by staining the outermost mannoprotein layer with Alexa647-ConA (Fig. 5d)65. The compromised cell wall in C. albicans treated with coniotin A and caspofungin was evident, as characterized by a collapsed cell surface and the simultaneous production of multiple daughter cells that failed to complete division (Fig. 5e), deviating from the normal unipolar budding where a single mother cell generates one daughter cell at a time66. Cells treated with coniotin A exhibited a wide neck and morphological heterogeneity, aligning with characteristics and phenotypes commonly observed in cells exposed to cell wall-targeting agents, such as caspofungin (Fig. 5e)67. These observations indicate impaired cell wall remodeling, accompanied by disruptions in the structure and composition of the cell wall. Furthermore, treatment with coniotin A induced a 1.5-fold increase in the cell perimeter, based on analysis of over 150 stained cells (Fig. 5e, f); similar to caspofungin, this suggests cell wall softening, likely due to reduced β−1,3 glucan content, which compromises C. albicans cell shape, mechanical rigidity, and osmotic resistance, resulting in swollen cells68.
Given similar physiological effects with caspofungin, we evaluated the impact of coniotin A on β−1,3 glucan levels using aniline blue staining65. This analysis revealed a significant reduction in the staining of cell surface glucan, especially in characteristic cell wall regions (Supplementary Fig. 5c). This reduction was accompanied by increased diffusion of the stain into the cells, indicating enhanced osmotic fragility68, suggesting that coniotin A primarily targets glucan fibrils, destabilizing polysaccharides and ultimately leading to cell wall damage. The direct interaction between coniotin A and β−1,3 glucan was confirmed through a pull-down assay. Over 50% of coniotin A bound to β−1,3 glucan within a 1-h incubation, whereas chitin did not bind coniotin A, leaving a greater amount of it in the supernatant (Fig. 5g). To further validate this interaction, a BODIPY fluorescent moiety was conjugated to the terminal Asp of coniotin A, enabling visualization of its binding to β−1,3-glucan via a pull-down assay, with glucan particles analyzed through aniline blue staining (Supplementary Fig. 5d).
The binding of coniotin A to glucan further hindered the interaction of other enzymes or factors with glucan, as evidenced by its dose-dependent inhibition of glucanase-mediated glucan digestion, resulting in decreased production of hydrolyzed short-chain oligosaccharides (Fig. 5h). Similarly, coniotin A inhibited the activation of limulus coagulation factor G, which is highly sensitive to (1,3)-β-D-glucan (Fig. 5i). Typically, glucan-bound factor G initiates the coagulation cascade, generating the detectable chromophore p-nitroaniline (pNA) from the chromogenic substrate Boc–Leu–Gly–Arg–pNA. However, incubating with 5 μg/mL coniotin A significantly reduced reaction dynamics, suggesting a decrease in free, intact glucan. This effect likely results from the interaction of coniotin A with glucan, which may disrupt its single helical conformation, a key contributor to the activation of limulus coagulation factor G (Fig. 5i)69. Collectively, these findings suggest that the binding of coniotin A to glucan interferes with its enzymatic modification and remodeling, ultimately leading to a functionally compromised cell wall.
TEM was used to further examine the morphological changes in cell wall structure under treatment with coniotin A. Candida cells typically display characteristic two-layered cell walls63,70, as seen in C. auris CBS 12766 control cells: an electron-dense, mannan-rich outer layer (M) and a glucan-rich inner layer of lower electron density (G + C) (Fig. 5j, i–ii), both continuous with the plasma membrane71,72. In cells treated with coniotin A, a predominance of a thicker, more electron-dense layer was observed in place of the translucent inner layer, with the cell wall detaching from the membrane (Fig. 5j, iii–iv), likely due to upregulated chitin production. Chitin appeared in the outer and inner wall layers, with cell wall proteins increasingly linked to chitin rather than β−1,3-glucan, as seen in caspofungin-treated, glucan-compromised cells63. C. neoformans cells have distinct cell wall structures, with an exopolysaccharide capsule (C) anchored to the outer layer, which contains both α-glucan and β-glucan. In contrast, the inner layer is primarily composed of β-glucans and chitin. These two layers are tightly interwoven, forming a dense, thin cell wall (W) closely adjacent to the membrane (Fig. 5j, v–vi)71. Despite capsule protection, coniotin A induced severe cell wall damage, along with a much thicker, highly electron-dense cell wall structure, suggesting activation of chitin salvage pathways to withstand cell wall stress (Fig. 5j, vii–viii). Abnormal multilayered cell walls and focal enlargements were also observed in both C. auris and C. neoformans under treatment with coniotin A, suggesting that cell wall stress led to aberrant thickening as a survival response (Supplementary Fig. 5e, i–iv). Although C. neoformans possesses robust mechanical barriers that resist caspofungin, maintaining an intact cell wall and capsule even when killed by amphotericin B, cells killed by coniotin A exhibited an aberrantly thickened cell wall with clear signs of disintegration (Supplementary Fig. 5e, v–vii). TEM analysis of cell wall damage morphology reveals that coniotin A disrupts the dynamic physiological activity of the cell wall, leading to structural compromise, likely due to its targeting of glucan.
Coniotins are generated by the hybrid PKS-NRPS BGC
Understanding the biosynthesis of coniotin is crucial for exploring, harnessing, and optimizing fungal lipopeptaibiotics, whose bioactivity and stability are closely tied to key structural features such as N- and C-terminal modifications, amino acid composition and charge, and peptide length73,74. Lipopeptide biosynthesis in fungi, primarily mediated by multimodular non-ribosomal peptide synthetases (NRPSs)75, has emerged as a rapidly advancing field, driven by growing interest and deepening mechanistic understanding76,77,78. Coniotins are rare, linear lipopeptaibiotics composed of 21 amino acids, produced by Coniochaeta hoffmannii, a plant-pathogenic ascomycete with largely uncharacterized secondary metabolism73. The structural features, including multiple noncanonical amino acids and an N-terminal MOTDA moiety, strongly suggest a biosynthetic origin from a hybrid polyketide synthase–NRPS (PKS-NRPS) system75,78.
To investigate the genetic basis underlying coniotin biosynthesis, we sequenced the genome of the producing strain C. hoffmannii WAC11161. AntiSMASH analysis predicted 30 BGCs, including three hybrid PKS–NRPS clusters located in genomic regions 15, 31, and 52 (designated Clusters 1, 2, and 3, respectively)35. Clusters 1 and 3 each encode only a single-module NRPS with limited domain architecture and lack key biosynthetic features, rendering them unlikely to be involved in coniotin production (Supplementary Fig. 6a–d and Supplementary Table 6). In contrast, Cluster 2 spans ~182 kb and encodes three NRPS genes (conB, conC, and conD) comprising a total of 21 modules, corresponding precisely to the number of amino acid residues in coniotin (Fig. 6 and Supplementary Table 5). Substrate specificity predictions for individual adenylation domains closely match most of the amino acid composition of coniotin, supporting the proposed NRPS assembly line (Supplementary Table 7). Notably, an intervening open reading frame, orf17, encodes a predicted L-aspartate decarboxylase that may supply the nonproteinogenic precursor β-alanine. Additionally, the cluster harbors a highly reducing, iterative type I polyketide synthase (ConA), which shares 100% query coverage and 67% sequence identity with TexQ—the PKS responsible for MOTDA synthesis in the recently reported lipopeptaibol texenomycin and lipohexin BGCs—suggesting its functional role in assembling the N-terminal MOTDA acyl moiety (Supplementary Fig. 6e)43,79,80. Adjacent to conA, the acyl-CoA synthetase ConE shows high sequence similarity to characterized fungal lipopeptide ligases, including LcsD (A0A179HJB8.1), EasD (Q5BA81.1)81, GloD (S3DB78.1), Cm3C (G3J455.1), and EcdI (K0E2F3.1). Together with the predicted transferase ConF, these enzymes likely mediate the essential transfer of the linear polyketide intermediate that is released from ConA to the initiating thiolation (T₀) domain of the NRPS ConB81.

Organization of the coniotin BGC and proposed pathway in C. hoffmannii WAC11161. Open reading frames (ORFs) involved in coniotin biosynthesis are color-coded as follows: pink for the polyketide synthase (PKS) gene (conA), blue for the non-ribosomal peptide synthetase (NRPS) genes (conB-D), dark purple for the acyl-CoA ligase gene (conE), light purple for the transferase gene (conF), light yellow for the ABC transporter, and green for potential functional genes. For ORF annotations, see also Supplementary Table 5. The PKS domains in ConA are labeled as follows: AT (acyltransferase), KS (keto synthase), ACP (acyl carrier protein), KR (ketoreductase), DH (dehydratase), ER (enoylreductase), and MT (methyltransferase). The NRPS domains in ConB-D are labeled as follows: C (condensation domain), A (adenylation domain), T (thiolation domain), and TD (terminal domain).
Building on gene annotations within the con BGC (Supplementary Table 5), we propose a biosynthetic pathway for coniotin A (Fig. 6). The iterative PKS ConA initiates polyketide assembly by condensing malonyl-CoA extender units through its ketoacyl synthase (KS) domain, with acyltransferase (AT) loading and acyl carrier protein (ACP) tethering. Stepwise two-carbon elongation, coupled with selective β-keto processing by canonical dehydratase (DH), enoyl reductase (ER), and ketoreductase (KR) domains, generates a fully reduced 3-oxotetradecanoyl scaffold. During the final extension cycle, β-C-methylation by the C-methyltransferase (cMT) domain generates methylated polyketides, yielding the major congeners coniotin A and B; while unmethylated intermediates give rise to coniotin C and D. These polyketides (OTDA/MOTDA) are subsequently converted into CoA thioesters by the acyl-CoA synthetase ConE and transferred to the T₀ domain of the NRPS ConB81. The first condensation domain (C1) mediates amide bond formation between proline and (M)OTDA, initiating nonribosomal peptide assembly. Sequential incorporation of 20 additional amino acids proceeds according to the predicted domain organization, and the mature lipopeptide is ultimately released by the terminal thioesterase domain (TD).
Discussion
Natural products and their derivatives have long served as an invaluable reservoir of therapeutic agents, contributing to nearly half of all approved anticancer drugs, due to the structural novelty, diversity, and complexity of their metabolites. Microorganisms play a pivotal role in antibiotic biosynthesis to gain a competitive advantage for survival. The availability of extensive microbial genome sequence databases has unveiled a vast reservoir of untapped BGCs in microorganisms, revealing an immense, largely unexplored chemical space. However, the frequent rediscovery of known compounds poses a significant challenge to identifying previously uncharacterized bioactive molecules.
To overcome these challenges, we developed a cost-effective and resource-efficient platform to uncover active agents masked within crude extracts that are often overlooked using traditional screening methods, achieving over a 50% increase in hit rate. The prefractionated library effectively separates growth-enhancing and antibiotic molecules, as well as major and minor components, facilitating the detection of minimal or negligible secondary metabolite production. By integrating PFL screening with a rapidly expanding database of annotated MS/MS fragmentation spectra and characterized biosynthetic BGCs, we developed an efficient dereplication pipeline to prioritize structurally novel scaffolds from complex natural product mixtures. Application of this strategy led to the discovery of coniotins, a previously uncharacterized family of lipopeptaibiotics composed of 21 amino acid residues and an N-terminal MOTDA moiety.
Unlike classical lipopeptaibols, coniotins terminate in a polar amino acid residue rather than the canonical C-terminal amino alcohol, representing a structural deviation that helps define this family as a distinct subclass within the broader peptaibiotic lineage57,78. Moreover, microbial lipopeptaibiotics with extended peptide chains (>20 residues) remain relatively rare, likely due to the high metabolic cost associated with their biosynthesis under standard laboratory conditions75. Their identification often depends on genome-guided mining, heterologous expression, or advanced analytical workflows82. The recovery of such a compound directly from native fungal fermentation highlights the effectiveness of our integrative discovery platform.
Coniotin A exerts antifungal activity through a non-canonical mechanism that targets β-glucan, an essential polysaccharide in the fungal cell wall that is absent in mammalian cells. This selective interaction underlies its low hemolytic and cytotoxic profile relative to amphotericin B. By compromising cell wall integrity, coniotin A facilitates caspofungin access to its target, resulting in robust synergistic activity. This mechanism is particularly relevant for combating multidrug-resistant C. auris, a WHO-prioritized pathogen with high clinical concern. Notably, other peptaibiotics have also demonstrated cell wall-directed antifungal activity, suggesting a broader role for such structural motifs in targeting the fungal surface40,83,84.
Structurally, coniotin A is enriched in non-proteinogenic residues, including six α-aminoisobutyric acid (Aib) and four D-isovaline (Iva) units, predominantly located at the termini. These residues stabilize helical conformations, such as α- and 3₁₀-helices58,78, that are characteristic of peptaibiotics and associated with proteolytic stability and durable bioactivity55,56. While the precise three-dimensional structure of coniotin A remains to be determined, its sequence composition and preliminary safety profile support its potential as a pharmacologically resilient antifungal agent.
Although no lipopeptaibol has yet advanced to clinical use, coniotins represent a rare class of antifungals that act selectively on fungal cell wall components, offering both reduced host toxicity and a low likelihood of resistance emergence48. Their potent activity against resistant pathogens and synergy with existing therapies position coniotins as candidates for further development. Despite violating Lipinski’s rules, their pharmacological behavior echoes that of approved lipopeptides such as daptomycin and echinocandins, including high serum stability and low resistance potential. Given their physicochemical properties, coniotins are most likely suited for parenteral or topical delivery. The hydrophobic tail may facilitate dermal penetration47, making them attractive for superficial fungal infections such as cutaneous or mucosal candidiasis. Future studies will aim to define their in vivo efficacy, therapeutic window, and optimal formulation strategies.
More broadly, peptaibiotic is an increasingly recognized class of bioactive peptides with potent antifungal, antibacterial, antiviral, antitumor, and neuroactive activities, representing a versatile and expanding scaffold for therapeutic innovation75,78,85. The discovery of coniotins further exemplifies the potential of natural peptides as drug leads. Peptide-based antimicrobials often act through multiple mechanisms of action, a feature that helps delay or prevent the emergence of resistance48,86,87,88. In the case of coniotins, which target essential microbial structures such as the cell wall, the development of resistance appears limited, potentially due to the evolutionary constraint that pathogens are disincentivized to extensively remodel cell wall architecture in ways that would compromise viability. Structural elucidation of coniotin A via NMR or crystallography, coupled with detailed analyses of its β-glucan interactions, will provide a foundation for rational design of next-generation lipopeptaibiotics.
The identification of the coniotin BGC provides a genetic framework for the modular assembly of these unusually large, Aib-rich lipopeptaibols. The cluster encodes a hybrid highly reducing polyketide synthase (HR-PKS, ConA) and multiple large NRPSs (ConB–D), collectively predicted to incorporate over 21 amino acids, including several non-proteinogenic residues such as α-aminoisobutyric acid (Aib) and isovaline (Iva). Recent work on texenomycins, linear Aib-rich lipopeptaibols featuring the same N-terminal MOTDA moiety, highlights convergent biosynthetic strategies among filamentous fungi43. Both the texenomycin and coniotin BGCs share a PKS–NRPS hybrid architecture. However, comparative BGC analysis reveals key distinctions: while the PKSs responsible for MOTDA biosynthesis are highly conserved (67% identity, 100% coverage), the remaining NRPS modules display low sequence similarity (<30%), indicating divergence in both peptide backbone construction and potential tailoring logic. These differences suggest that, despite structural parallels, coniotins and texenomycins have evolved independently, pointing to distinct biosynthetic solutions for producing large lipopeptaibols in fungi and expanding the mechanistic repertoire of this underexplored class of natural products.
Targeted gene deletions, particularly within the loading modules of the PKS and NRPS systems, will be instrumental in defining the contributions of polyketide and peptide moieties to bioactivity, and may facilitate the rational design of analogs with improved pharmacological profiles. Collectively, our findings underscore the underexplored potential of large fungal lipopeptaibiotics and provide a foundation for future genome-guided discovery and synthetic biology approaches aimed at expanding the antifungal lipopeptide repertoire.
Methods
Ethics statement
All experiments involving human blood were conducted in accordance with relevant institutional and national ethical regulations. Commercially obtained, fully anonymized red blood cells (RBCs) (BioIVT) were used. As the study did not involve identifiable data or direct interaction with human participants, ethics approval was not required under TCPS2 (Article 12.3B) and McMaster University policies.
Cultivation and fermentation conditions of bacterial strains
Streptomyces sp
Strains WAC1325, WAC1490, and WAC5858 (this study) were initially cultured on Mannitol Soya Flour (MS) agar (2% mannitol, 2% soya flour, 2% agar) at 30 °C for 7 days to promote sporulation. A single colony from the sporulated MS agar culture was then transferred to Bennett’s agar plates and incubated under the same conditions for an additional 7 days for fermentation.
Bennett’s medium was prepared with the following composition per litre: 10 g potato starch, 2 g casamino acids, 1.8 g yeast extract, and 2 mL Czapek mineral mix. The Czapek mineral mix contained 10 g KCl, 10 g MgSO₄·7H₂O, 12 g NaNO₃, 0.2 g FeSO₄·7H₂O, 200 µL concentrated HCl, and was adjusted to 100 mL with double-distilled water (ddH₂O). The final pH was adjusted to 6.8, and the medium was autoclaved at 121 °C for 45 min.
B. velezensis
WAC11084 strains (this study) were revived from cryopreserved stocks and cultured in Luria–Bertani (LB) broth or on LB agar plates, with incubation overnight at 37 °C. For fermentation, the strain was transferred to Bennett’s medium and incubated at 37 °C for 5 days.
Culture conditions for fungal and yeast strains
C. hoffmannii
WAC11161 (this study), C. neoformans strain H99 (ATCC MYA-4901), C. auris (CBS10913, CBS12766, CBS12775, CBS12776; obtained from CBS-KNAW Fungal Biodiversity Centre), C. albicans (ATCC 90028, ATCC 200955), C. parapsilosis ATCC22019, C. tropicalis ATCC200956, C. glabrata CBS138 (formerly N. glabratus; CBS-KNAW), S. cerevisiae BY4741 (ATCC 201388), and A. fumigatus (Af293, 1478)89 were cultured under standard eukaryotic conditions. Cultures were grown in YPD medium (1% yeast extract, 2% peptone, and 2% dextrose) or Sabouraud Dextrose Broth (SDB, BD Difco) at 30–37 °C. When required, strains were maintained on corresponding agar plates. To ensure cell viability, sterile techniques were employed, and cells were regularly passaged.
Fermentation of coniotins
To produce coniotin A, C. hoffmannii WAC11161 cultures were grown on Bennett’s agar plates and incubated for 8 days at 30 °C.
High-throughput screening of natural product library and PFL
A high-throughput screening of a natural product library and a prefractionated library was performed to assess the advantages of fractionated libraries over crude extracts. The screening encompassed 379 crude extracts and 3032 corresponding fractions, each tested in duplicate. The screening was performed against C. albicans ATCC 90028 and C. auris CBS 10913 using the Biomek Fxp Integrated Liquid Handler. Candida cultures were streaked on YPD agar for single colonies and incubated at 30 °C for 48 h. Cultures were then prepared to a final concentration of 103 cells/mL in RPMI 1640 medium. In 384-well plates, 1 μL of crude extract, conditioned media, or fraction was mixed with 49 μL of the yeast culture using the Formulatrix Tempest Liquid Handler. Amphotericin B (8 μg/mL) served as a positive control. After 48 h of incubation, plates were read at OD530 on a Biotek Neo microtiter plate reader, and luminescence and absorbance were measured using BioTek Gen5 software (v3.14). Data were normalized using control-based normalization, and hits were defined as wells exhibiting a minimum of 75% growth reduction for both C. albicans and C. auris90.
Hit verification
Nine hits were selected for follow-up verification based on the initial screening data. The corresponding strains were revived from cryopreserved stocks, plated on Bennett’s agar, and incubated at 30 °C for 7 days. The agar cultures were then crushed and extracted three times with methanol. Methanol extracts were pooled and evaporated using a rotary evaporator. The dry samples were resuspended in DMSO and loaded onto a pre-packed C18 sample load cartridge. Fractionation was performed using a CombiFlash system (Teledyne ISCO, Inc.) equipped with REDISEP GOLD® C18 reversed-phase columns. Separation was achieved at a flow rate of 12 mL/min using a water-acetonitrile (CH₃CN) gradient. A total of 24 fractions were collected per run and dried using Genevac Evaporators (Canadawide Scientific). The dried fractions were then dissolved in 200 μL DMSO with sonication, and their antifungal activity was evaluated against C. albicans and C. auris.
Susceptibility test of antifungal agents
Minimum inhibitory concentration (MIC) determinations were performed following the National Committee for Clinical Laboratory Standards (NCCLS) protocol M27 (Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts). Several colonies from two-day-old cultures were resuspended in 0.85% saline to an initial OD530 of 0.11–0.14 and then diluted 1:2000 in RPMI 1640 medium. A two-fold serial dilution of test agents was prepared and added to the diluted culture in 96-well U-bottom plates. Column 11 served as the growth control (inoculum without drug), and Column 12 served as the sterile control (sterile media only). Both controls contained the same vehicle (e.g., DMSO) as the test wells. The sterile control readings were labeled “bkgd” (background), and the growth control readings were labeled “growth.” Growth inhibition was calculated as: % growth = [(OD530 – mean bkgd)/(mean growth − mean bkgd)] × 100. For the bioactivity testing of fractions, 4 μL of DMSO-dissolved fractions were added to the diluted culture. After 48 h of incubation at 30 °C, optical density (OD) at 530 nm was measured using a BioTek Synergy Microplate Reader. MIC for fluconazole was defined as the lowest concentration that caused an 80% reduction in growth, while MIC for other drugs was set as the lowest concentration that completely inhibited growth.
HRMS and chromatographic analysis
HRMS analysis of active fractions was performed using a qTOF LC/MS/MS system. An Agilent 1290 Infinity II LC system coupled to a qTOF 6550 mass detector (Agilent Technologies) was used to acquire mass spectra with Agilent MassHunter Workstation LC/MS Data Acquisition software (version 10.1). The instrument operated in positive electrospray ionization (ESI) mode with the following parameters: capillary voltage, 3500 V; nozzle voltage, 1000 V; fragmentor voltage, 380 V; drying gas flow, 14 L/min at 200 °C; nebulizer pressure, 35 psig; sheath gas flow, 11 L/min at 350 °C. Data were acquired over an m/z range of 100–3000 with a scan rate of 1 spectrum/s.
Targeted MS/MS analyses were performed on a list of specific precursor ions to confirm the structure of coniotins. The instrument settings were as follows: MS range was set to 50–3000 m/z, and MS/MS scans from 50 m/z to 3000 m/z, with fixed collision energies of 20 eV, 30 eV, 40 eV, and 50 eV. For GNPS-based untargeted analysis, auto MS/MS mode was used with an m/z range of 50–1700, a scan rate of 1 spectrum/sec, and an isolation width of 4 amu. Collision energies of 10 eV, 30 eV, and 60 eV were applied.
Chromatographic separation was achieved using an Eclipse SDB-C8 column (2.1 mm × 100 mm, 3.5 µm; Agilent) with a flow rate of 0.4 mL/min. The mobile phase consisted of H₂O (0.1% formic acid, v/v) and acetonitrile (0.1% formic acid, v/v). For targeted analysis of coniotins, the gradient started at 25% B for 0.5 min and increased to 100% B over 6.5 min. For GNPS acquisition, the gradient started at 10% B for 2 min, then ramped linearly to 100% B over 15 min.
Agilent Mass Hunter 10.0. Qualitative analysis software was used for data analysis. Compounds purified from different batches of fermentation were dissolved in DMSO, tested for bioactivity, and the active fractions were examined on the Qtof at least three times. For the coniotins, over ten times independent analysis of the different batches of fractions of fermentations were conducted.
A total of more than ten independent fermentation batches were analyzed. Bioactive fractions from each batch were subjected to repeated HRMS analysis (at least three independent runs per batch). Inactive fractions were included as negative controls for dereplication and specificity assessment, and DMSO was used as a blank control.
Data were processed using Agilent MassHunter Qualitative Analysis software (version 10.0). GNPS molecular networking was performed via the GNPS online platform (https://gnps.ucsd.edu) using the Feature-Based Molecular Networking (FBMN) workflow. MS/MS spectra were matched against public spectral libraries with a cosine similarity score threshold of ≥0.7.
The raw MS and MS/MS datasets have been deposited in the MassIVE/GNPS public repository under accession number MSV000098505 (https://doi.org/10.25345/C5057D541). Relevant peak assignments and spectral annotations are provided in the main figures and Supplementary Information.
Identification of known antifungals by GNPS via HRMS/MS
Raw HRMS/MS data were converted to mzXML format and analyzed using the GNPS platform (Global Natural Product Social Molecular Networking, https://gnps.ucsd.edu)30 via its online workflow (https://ccms-ucsd.github.io/GNPSDocumentation/). Data were filtered to exclude fragment ions within ±17 Da of the precursor m/z, retaining the top six fragment ions within ±50 Da throughout the spectrum. Mass tolerances were set to 1.0 Da for precursor ions and 0.5 Da for MS/MS fragment ions. A molecular network was generated with edges requiring a cosine score > 0.7 and at least six matched peaks. Nodes were connected only if they appeared in each other’s top ten most similar nodes. Molecular families were capped at 100 nodes by removing the lowest-scoring edges. Spectra in the network were searched against GNPS spectral libraries using the same filtering criteria. Matches required a cosine score > 0.7 and at least six matched peaks. This approach enabled the identification of known antifungal compounds.
Genome isolation
Genomic DNA was extracted from WAC1325, WAC1490, WAC5858, WAC11084, WAC11161, and WAC11175 for sequencing. Prokaryotic genomes were isolated from 48-h Tryptic Soy Broth (TSB) cultures. Cells were harvested and treated with 1 mg/mL lysozyme, followed by 1% SDS and 0.5 mg/mL proteinase K at 55 °C for 2 h. Proteins were removed via chloroform extraction and centrifugation. DNA was precipitated using cold isopropanol, washed with 70% ethanol, and dissolved in TE buffer. Residual RNA was eliminated using 100 μg/mL RNase.
Fungal DNA was isolated using a CTAB-based method91. Freeze-dried fungal cells were disrupted with glass beads and extracted with CTAB buffer (100 mM Tris-HCl, 0.7 M NaCl, 10 mM EDTA, 1% CTAB, 1% 2-mercaptoethanol, pH 7.5) at 65 °C for 30 min. Proteins were removed by chloroform extraction and centrifugation. DNA was precipitated with isopropanol, washed with 70% ethanol, and dissolved in TE buffer.
Genome sequencing and assembly
Genomic DNA was prepared for Illumina sequencing (MiSeq 2 × 300 bp reads) using the NEB Next Ultra V2 kit (New England Biosciences) with 500 ng of input DNA sonicated to 600 bp and size-selected with AMPure XP beads (Beckman Coulter). Sequencing was performed by the McMaster Genomics Facility, and reads were trimmed with Skewer v0.2.2 (-q 25, -Q25) and merged using FLASH v1.2.1192. De novo assembly was carried out with SPAdes v3.15.2 or SPAdes v3.15.4.
For WAC11161, the draft genome assembly consisted of 210 contigs with a total length of 35.4 Mb and an N50 of 778,487. The 18S rRNA gene sequence, retrieved with RNAmmer v1.2, was identified via BLASTN searches93. The top hits were Coniochaeta prunicola (99.88% identity over 93% query coverage) and C. hoffmannii (99.39% identity over 100% query coverage). Genome quality was assessed with BUSCO v5.4.7, confirming 97.9% completeness of conserved orthologs from the Sordariomycetes database (odb10)94. Illumina reads re-mapped using BWA MEM yielded 121X average coverage, a mean mapping quality of 59.89, and a mean base quality of Q33.89 (>99.95% base accuracy)95. Breseq v0.37.0 showed 99.0% of reads mapped to the assembly95. Fungismash (antiSMASH 8.035) identified a 182,160 bp BGC within a 444,462 bp contig associated with coniotin A. These results demonstrate a high-quality draft assembly of the Coniochaeta genome with excellent coverage and completeness, enabling further analysis.
Purification of active compound 1 from WAC11161
Single colonies of C. hoffmannii WAC11161 were picked from YPD agar plates after two days of growth and incubated on 20 Bennett’s agar plates (30 × 42 × 3 cm, 500 mL/plate) at 30 °C for 8 days. The fermented agar was blended and extracted with an equivalent volume of methanol under shaking. The methanol extract was concentrated under reduced pressure and resolubilized in 200 mL of methanol. Following centrifugation, the crude supernatant was combined with 5 g of C18 resin, dried by rotary evaporation, and loaded onto a RediSep C18 Gold column (86 g) for purification using a CombiFlash system (Teledyne ISCO, Inc.) at a flow rate of 66 mL/min. Partially purified compound 1 was eluted with a linear gradient of 10%–100% acetonitrile (0.1% formic acid). Active fractions were identified through bioactivity testing and LC-MS analysis, pooled, lyophilized, and further purified on an LH20 column (3 × 40 cm) with methanol as the eluent (fraction size: 10 mL).
Bioactive fractions (fractions 7–11) were concentrated to dryness using Genevac Evaporators (Canadawide Scientific) and subjected to HPLC purification (1260 Agilent Technologies) on an Eclipse SDB-C8 column (4.6 × 250 mm, 5 µm). The compound was eluted with 70% acetonitrile (0.1% formic acid) and assessed by HR-ESI-MS in positive ion mode. Compound 1: calculated mass for C98H170N21O26 [M + H]+: 2057.2620; observed 2057.2614. Approximately 10 mg of compound 1 was obtained. NMR data were acquired on a Bruker AVIII 700 MHz instrument equipped with a cryoprobe.
Chemical hydrolysis
Acid hydrolysis was performed by resuspending 1 mg of the compound in 500 μL of 6 N HCl and incubating the mixture at 100 °C for 20 h46. After the reaction cooling, 1 mL of ethyl acetate was added to the mixture. The organic phase was separated, dried, and subjected to analysis, while the aqueous phase was concentrated under nitrogen and used for subsequent derivatization with Marfey’s reagent.
Partial hydrolysis was conducted using 3 M HCl at 90 °C for 5 h. Aliquots were collected at defined time points and analyzed using HR-LC-MS. Selected cleaved peptides were further characterized using targeted MS/MS analysis.
Marfey’s reagent chemical modification
Approximately 0.2 mg of each amino acid standard was dissolved in 50 μL of H₂O, followed by the addition of 20 μL of 1 M NaHCO₃ and 100 μL of 1% Marfey’s reagent (Nα-(2,4-dinitro-5-fluorophenyl)-L-alaninamide, Acros Organics) in acetone46. The mixtures were agitated at 40 °C for 1 h, and the reactions were quenched with 10 μL of 2 N HCl. The resulting reaction products were dried under nitrogen, redissolved in ~1.7 mL methanol, and individually injected (0.5 μL) into a UPLC-MS for analysis.
To digest and derivatize hydrolyzed compounds: approximately 0.2–0.3 mg of compounds 1–3 were separately hydrolyzed in 500 μL of 6 N HCl at 90 °C for 24 h. The hydrolysates were dried under nitrogen and then treated with 25 μL of H₂O, 25 μL of 1 M NaHCO₃, and 50 μL of 1% Marfey’s reagent in acetone. Reactions were agitated at 40 °C for 1 h and quenched with 5 μL of 2 N HCl. The products were dried under nitrogen, dissolved in ~200 μL methanol, and analyzed by UPLC-MS under the same conditions as the standards.
Papain hydrolysis
Compound 1 (1 mg dissolved in 10 µL DMSO) was hydrolyzed in a reaction mixture containing 200 µL of 0.05 M Tris-HCl buffer (pH 6.8), 20 mM 2-mercaptoethanol, 0.5 mM EDTA, and 7 mg of papain45. The reaction was incubated at 37 °C with shaking for 4 days to ensure complete hydrolysis. Hydrolysis progress was monitored daily using LC-MS. At each time point, a 20 μL aliquot was collected from the reaction mixture, centrifuged, and the supernatant removed. The resulting pellet was resuspended in 20 µL of methanol, and 5 µL of the resuspension was injected for LC-MS analysis. Prominent peptide molecular ions were selected for further fragmentation analysis.
Hemolysis testing
Human blood collected in K2-EDTA tubes was obtained from BioIVT (New York, USA). The blood was centrifuged at 500×g for 5 min, and the plasma was removed. RBCs were washed twice with 150 mM NaCl in a volume equal to the removed plasma. After the second wash, RBCs were resuspended in phosphate-buffered saline (PBS, pH 7.4) at a volume equivalent to the plasma to maintain hematocrit levels. Compound solutions (1 μL) were added to 96-well V-bottom plates using a Labcyte Echo acoustic dispenser (Beckman Coulter). DMSO was included at a constant 1% (v/v) final concentration, with DMSO-only controls as negative controls. Triton X-100 (10 μL, starting at 20% and serially diluted 2-fold to 0.02%) served as a positive control. RBCs were diluted 1:50 in PBS (pH 7.4), and 99 μL of this suspension was added to each well. Plates were incubated at 37 °C for 1 h, followed by centrifugation at 500×g for 5 min to pellet intact RBCs. A 65 μL aliquot of the supernatant was transferred to a clear, flat-bottom 96-well plate, and absorbance was measured at 540 nm. Coniotin A and B were tested at a starting concentration of 128 μg/mL, while Iturin A, Caspofungin, and Amphotericin B were tested starting at 256 μg/mL. Compounds were diluted 2-fold to create an 11-point dose-response curve. Each concentration was tested in duplicate.
Cytotoxicity testing
On Day 1, HEK293 cells (ATCC CRL-1573; generation 6) were seeded at 7500 cells/well in 384-well tissue culture-treated white plates with 50 μL of Dulbecco Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 2 mM L-glutamine, 100 units/mL penicillin, and 100 μg/mL streptomycin. Cells were incubated for 18 h at 37 °C under 5% CO₂. On Day 2, 500 nL of compound solutions and DMSO were added to the wells using a Labcyte Echo acoustic dispenser (Beckman Coulter) and a Combi nL dispenser (ThermoFisher), maintaining a final DMSO concentration of 1% across all wells. After 48 h of incubation, cell viability was assessed using Promega CellTiter-Glo 2.0 reagent (Fisher Scientific). A total of 50 μL of CellTiter-Glo was added directly to each well, plates were shaken for 2 min, and then incubated for 10 min at room temperature. Luminescence was measured using a Neo2 plate reader (BioTek) equipped with a luminescence fiber optic and operated with Gen5 software (v3.14, BioTek). Untreated cells and DMSO-only treated cells were used as controls. Compounds were tested in triplicate at each concentration. Coniotin A and B were tested starting at 128 μg/mL, while Iturin A, Caspofungin, and Amphotericin B were tested starting at 256 μg/mL. Compounds were serially diluted 2-fold to generate an 11-point dose-response curve.
Dose-response curves were fitted using a four-parameter logistic (4PL) non-linear regression model, constrained to a minimum response of 0 and a maximum response of 1. The 4PL Eq. (1) used was:
Where y = the sample response in relative luminescence units, x = the drug concentration, a = the maximum response for infinite standard concentration, b = the Hill slope (typically negative), c = inflection point, and d = the response at a standard concentration of 0.
Rapid C. elegans–C. albicans antifungal activity assay
The C. elegans glp-4(bn2); sek-1(km4) double mutant AU37 (Caenorhabditis Genetics Center) was used in a rapid co-infection antifungal assay50. Briefly, worms were age-synchronized to the first larval stage (L1) and grown on nematode growth medium (NGM) at 25 °C for 48 h until the fourth larval stage (L4). In 96-well clear flat-bottom plates, 70 µL of screening medium (30% BHI in M9 buffer containing 90 µg/mL kanamycin, 200 µg/mL ampicillin, and 200 µg/mL streptomycin), 450nL of test compounds or DMSO vehicle, ~15 L4 worms, and 10 µL of C. albicans ATCC90028 (2.5 × 10⁴ cells/mL in PBS) were added per well. Worms were dispensed using the Union Biometrica COPAS BIOSORT system, and plates were sealed with a porous film. Plates were incubated at 25 °C for 96 h prior to imaging using a Nikon Multizoom AZ100M microscope equipped with a 2× Plan Fluor objective. Images were captured with NIS-Elements AR software (v5.11, Nikon).
C. elegans survival assay
A population of the C. elegans double mutant strain AU37 (glp-4(bn2); sek-1(km4)) was synchronized to L1 and grown on NGM at 25 °C for 48 h prior to infection. The C. auris infection protocol was adapted and scaled for 96-well plates51. Worms were washed with M9 buffer and placed onto brain heart infusion (BHI) agar plates supplemented with 50 µg/mL kanamycin and seeded with a C. auris lawn. After feeding for 3 h, worms were washed off and allowed to crawl on unseeded NGM plates for 1 h to remove external yeast. Using the Union Biometrica COPAS BIOSORT system, ~25 worms were dispensed into each well of a 96-well plate. The final media composition was 20% BHI and 80% M9 buffer supplemented with 10 µg/mL cholesterol. Test conditions included DMSO vehicle, 1× MIC of coniotin A, and amphotericin B, each tested in triplicate. Plates were sealed with a porous film, incubated at 25 °C, and worm survival was monitored every 8 h over 48 h.
Proteolytic stability assay of coniotin A
Protease activity was first confirmed using chromogenic peptide substrates. Specifically, 400 μM Nα-Benzoyl-L-arginine 4-nitroanilide hydrochloride (BApNA; Sigma-Aldrich, B3133), a trypsin substrate, and 400μM N-Succinyl–Ala–Ala–Ala–p-nitroanilide (Suc-Ala–Ala–Ala-pNA; Sigma-Aldrich, S4760), a substrate for neutrophil elastase, were incubated with 1 μg of trypsin (Promega, V511A) or elastase (Sigma-Aldrich, E0258) in Tris-HCl buffer (pH 7.5) at 37 °C. Release of pNA was monitored at 410 nm every 1 min using a microplate reader (Synergy Neo2, BioTek) operated with Gen5 software (v3.14, BioTek).
Coniotin A (40 μM) was subsequently incubated with either trypsin, elastase, or 50% (v/v) pooled human serum (Innovative Research, IPLA-HSER) in Tris-HCl buffer (pH 7.5) at 37 °C. At predetermined time points (0, 10, 20, 30, 60, 120, 300, 450, and 720 min), 10 μL aliquots were collected and immediately quenched with an equal volume of DMSO. Samples were analyzed using an Agilent 1290 Infinity II LC system (Agilent Technologies) coupled to a 6550 iFunnel Q-TOF mass spectrometer to quantify the remaining intact peptide. LL-37, a human antimicrobial peptide, served as a control.
Haploinsufficiency (HIP) screening
Glycerol stock pools of C. albicans HET double-barcoded deletion mutants were thawed, diluted to an OD600 of 0.05 into a 60 mL YPD culture, and grown at 30 °C under shaking conditions for 1.5 h. Subsequently, 1 mL of the sub-cultured HET pool was aliquoted into triplicate culture tubes, each containing 1 mL YPD medium with coniotin A or a DMSO solvent control. These cultures were grown at 30 °C under shaking conditions for 18 h. Cells were pelleted by centrifugation, the supernatant was removed, and the cell pellets were stored at −80 °C. Cell pellets were digested with Zymolase in buffer (1 M sorbitol, 10 mM sodium EDTA, 14 mM β-mercaptoethanol, 15 units of Zymolase enzyme) prior to genomic DNA extraction using the PureLink Genomic DNA Extraction kit, as per the manufacturer’s instructions (Invitrogen). Genomic DNA was recovered from columns provided by the kit using 10 mM Tris-HCl pH 8.0 and quantified using the PicoGreen DNA quantification kit (Invitrogen). Barcodes were PCR amplified with Takara Ex-Taq (Clonetech) using 150 ng of genomic DNA. UP-TAG primers (UP-TAG U and UP-TAG INX) and DOWN-TAG primers (DOWN-TAG U and DOWN-TAG INX) were used (Table S8)96. Equal quantities of UP-TAG and DOWN-TAG pools were combined to form a sequencing library, which was sequenced on an Illumina Next-Seq500 instrument (Mid-Output, V2 Chemistry) using specific primers to sequence and index the UP- (UP-TAG S and UP-TAG SINX) and DOWN-TAG (DOWN-TAG S and DOWN-TAG SINX) pools for each sample (Table S8)96. Barcode-sequence reads were mapped to an artificial genome containing known UP-TAG and DOWN-TAG sequences of each strain and compiled for each indexed sample. If a specific UP-TAG or DOWN-TAG had more than one of its triplicate samples in the solvent control condition with read counts <20% of the median read per million mapped, these reads were filtered and omitted from further analysis. Log2 fold differences for each strain’s UP-TAG and DOWN-TAG were calculated.
Intracellular drug accumulation assay
Intracellular accumulation of caspofungin, iturin A, and coniotin A was assessed using a quantitative LC-MS-based approach performed in biological triplicates61. C. albicans ATCC90028 and C. auris CBS10913 were used for the experiments. Overnight cultures (OD530 = 1.6–1.8) were subcultured into fresh SDB and grown at 30 °C with shaking until reaching mid-log phase (OD₅₃₀ ≈ 0.6). Cells were pelleted, washed twice with PBS, and resuspended in 15 mL fresh PBS. Aliquots (875 μL) were transferred into ten 1.5 mL Eppendorf tubes, resulting in a final concentration of ~3.3 × 107 cells/mL. Samples were equilibrated at 30 °C for 5 min before adding test compounds to a final concentration of 20 µM. After 10 min of incubation, 800 μL of the culture was layered onto 700 μL of pre-cooled silicone oil (9:1 mixture of silicone oil AR200 and Sigma High-Temperature silicone oil with 13.3% hexane) and centrifuged at 13,000×g to pellet cells through the oil. Surface-bound compounds were removed by the silicone oil interface. The supernatant and oil layers were carefully removed by pipetting. Pellets were transferred to new tubes, washed twice with water, and extracted with 150 μL DMSO/MeOH (2:1, v/v). Extracts were analyzed and quantified using HR-ESI-MS on an Agilent 1290 Infinity II HPLC system coupled with a qTOF 6550 ESI/MS (Agilent, USA), equipped with an Eclipse SDB-C8 column (2.1 mm ID × 100 mm, 3.5 µm; Agilent, USA). The instrument was operated in positive ion mode with a flow rate of 0.3 mL/min. The mobile phase consisted of 0.1% formic acid in water (phase A) and 0.1% formic acid in acetonitrile (phase B). All compounds used in biological assays were of ≥95% purity. Error bars represent the standard error of the mean of three independent biological replicates.
Calcofluor white staining and widefield microscopy
Fresh overnight cultures of C. albicans ATCC90028, C. neoformans H99, and C. auris CBS10913 were sub-cultured in YPD broth to an OD600 of 0.1. Cultures were treated with DMSO or half the MIC of test agents and incubated at 30 °C with shaking for 4 h. Cells were washed with PBS, resuspended in PBS to an OD600 of 3, and stained with calcofluor white (10 μg/mL). Imaging was performed using a Nikon Eclipse Ti inverted microscope equipped with a 100× Plan Fluor Apo λ oil immersion objective. Micrographs were captured as raw 16-bit TIFF files using NIS-Elements AR software (v4.50, Nikon) with a 4′,6-diamidino-2-phenylindole (DAPI) hybrid filter, and a SpectraX LED fluorescence source (20% power). Probe exposure was set to 50 ms, with gain at 0, and minimum/maximum values held constant across all samples. Fluorophore Intensity Quantification: Image analysis was performed using ImageJ and CellProfiler97. An ImageJ macro was used to subtract the background with a rolling ball radius of 50 pixels. The processed images were analyzed in CellProfiler (v4.2.1) to identify cells as primary objects and quantify fluorescence intensities for whole cells and cell edges. Calcofluor intensity per cell was calculated by dividing the total fluorescence intensity by the number of cells.
Cell surface staining and super-resolution confocal imaging
Cell surface properties were analyzed by staining with concanavalin A conjugated to Alexa Fluor™ 647 (Invitrogen, C21421)65. An overnight culture of Candida albicans ATCC 90028 was subcultured into SDB medium at an initial OD₆₀₀ of 0.1 and incubated at 30 °C with shaking for 4 h in the presence of coniotin A or caspofungin (each at ½ MIC) or an equivalent volume of DMSO (vehicle control). Treated cells were harvested by centrifugation, washed twice with 1 mL PBS, and briefly sonicated to remove aggregated debris. Cells were then incubated with 50 μg/mL Alexa Fluor 647-ConA for 10 min at room temperature in the dark. After staining, cells were washed three times with PBS, resuspended to an OD₆₀₀ of 1.0, and mounted onto square coverslips (#1.5, 12-541-AP, Fisherbrand) using ProLong™ Diamond Antifade Mountant (Invitrogen, P36961).
Imaging was performed with a Zeiss LSM 980 inverted confocal microscope (Zeiss Axio Observer.Z1/7 stand), equipped with an Airyscan 2 detector and a 63x/1.4 oil-immersion objective. Image acquisition was carried out using ZEISS ZEN Blue software (v3.8.99, Carl Zeiss, Germany). Excitation was performed using a 639 nm laser, and emission was collected using a 528/29 + 697/38 nm multi-band-pass filter. Z-stacks comprising 40–60 slices were acquired with a 0.2 µm step size in Airyscan mode (superresolution: 7.6 (3D, Auto)) using a pixel size of 56 nm. Super-resolution images were reconstructed using 3D Airyscan processing in ZEISS ZEN Blue v3.8.99. Cell perimeter measurements were performed using ImageJ (64-bit Java 8 bundled version) software98.
Glucan pull-down assay
A glucan pull-down assay was conducted to investigate the interaction between Coniotin A and polysaccharides. β-Glucan (1 mg/mL; Millipore Sigma 346210) and chitin (1 mg/mL; Sigma C9752) were suspended in 1× PBS (pH 7.0). Coniotin A (2 μL, 3.2 mg/mL stock solution) was added to 200 μL of each suspension, followed by incubation with shaking at 30 °C for 1 h. After incubation, samples were centrifuged for 5 min to separate the supernatant and pellet fractions. The supernatant was collected to analyze unbound coniotin A. The polysaccharide pellets were washed three times with PBS and extracted with DMSO using sonication to release bound Coniotin A. Coniotin A was identified and quantified in both the supernatant and DMSO extracts using an Agilent 1290 Infinity II LC system (Agilent Technologies) coupled with a qTOF 6550 mass detector and an Eclipse SDB-C8 column (2.1 mm ID × 100 mm, 3.5 µm; Agilent, USA). Coniotin A was identified by its molecular ion and quantified by peak area comparison to a pure standard. All experiments were performed in three biologically independent replicates, each consisting of a separate incubation of polysaccharides with coniotin A. Each dot represents one independent sample. The unit of study is each independent incubation. No technical replicates were pooled for statistical analysis. Data are presented as mean ± standard deviation (SD).
Synthesis and application of coniotin A-BODIPY
Coniotin A (2 mg, 0.97 µmol, 1 eq) was dissolved in DMSO, and a solution of Bodipy-FL-ethylenediamine (3.6 µg, 9.7 µmol, 10 eq) in DMSO was added. Benzotriazol-1-yloxytripyrrolidinophosphonium hexafluorophosphate (PyBop) (10 µL of 1 M solution in 50% DMF/DMSO, excess) and N-methylmorpholine (1 µL of 1 M solution in 50% DMF/DMSO, excess) were then added to the mixture. The reaction was carried out at room temperature (RT) with stirring for 40 min, after which the reaction mixture was lyophilized. The product was purified by semi-preparative HPLC (1260 Agilent Technologies) using an Eclipse SDB-C8 reverse-phase column (4.6 × 250 mm, 5 µm) with a gradient elution of water (0.1% formic acid) and acetonitrile (0.1% formic acid). The flow rate was set to 2 mL/min, and the compound was eluted with 80% acetonitrile (0.1% formic acid) and assessed by HR-ESI-MS in positive ion mode. Calculated mass for C130H208B2F4N29O26 [M + 1H]2+ 1344.7981; observed: 1344.7984. Purified product was collected, lyophilized, and stored for subsequent use.
β−1,3-glucan particles (1 mg/mL) were incubated with 120 µM BODIPY-conjugated coniotin A in PBS for 1 h. After incubation, the particles were washed three times with PBS and imaged using a Zeiss LSM 980 Upright Confocal Microscope (Zeiss Axio Imager.Z2 stand) with a GaAsp-Pmt3 detector using a 63×/1.4 oil-immersion objective. BODIPY was excited with a 488 nm laser, and fluorescence was detected using a FITC channel (525/28 nm bandpass). Imaging was performed using ZEN Blue software (v3.8.99, Carl Zeiss, Germany). To observe the general morphology of the glucan particles, a transmitted confocal laser image was captured in brightfield mode. Merged images were then generated by overlaying the brightfield and fluorescence signals in ImageJ.
Glucanase digestion assay
A glucanase digestion assay was performed using laminarin as the substrate to evaluate the effects of coniotin A on β-(1,3)-D-glucanase activity. Laminarin (125 μg/mL, Sigma L9634) was digested with β-(1,3)-D-glucanase (250 μg/mL, Sigma 67138) in 100 μL of 1× PBS (pH 7.0) containing 31.2, 62.4, or 124.8 μM coniotin A, or an equivalent volume of DMSO as a vehicle control. Reactions were incubated at 30 °C for 30 min. A negative control was performed by omitting β-(1,3)-D-glucanase. The reactions were quenched by adding 100 μL methanol, and 5 μL of the resulting mixture was analyzed using an Agilent 1290 Infinity II LC System (Agilent Technologies) coupled with a qTOF 6550 mass detector on a Luna HILIC 200 Å column (4.5 mm ID × 100 mm, 5 μm; Phenomenex). The oligosaccharide product (hexa-glucose) generated from the laminarin digestion was identified by its mass signal and quantified based on peak area. All experiments were conducted in three biologically independent replicates, each consisting of a separately prepared and processed enzymatic digestion reaction, and presented as one data point. The unit of study is each independent enzymatic digestion. No technical replicates were pooled for statistical testing. Values are shown as mean ± SD.
Kinetic assay of β-glucan activation of Limulus coagulation factor G
The kinetic chromogenic assay specific to (1,3)-β-D-glucan was performed using the Glucatell® (1,3)-Beta-D-Glucan Detection Reagent Kit (Associates of Cape Cod) following the manufacturer’s instructions. A 100 pg/mL solution of the provided glucan standard was incubated at 30 °C for 1 h with coniotin A (0.625, 5, or 40 μg/mL) or with an equivalent volume of DMSO as a control. Following incubation, 25 μL of each sample was added to designated wells containing 100 μL of freshly reconstituted Glucatell reagent. LAL Reagent Water served as a negative control. Kinetic absorbance readings were recorded at 405 nm every 30 s for 1 h using a preheated plate reader (Synergy Neo2, BioTek) operated with Gen5 software (v3.14, BioTek). All experiments were performed in three biologically independent replicates, each consisting of a separate CNA-glucan incubation and Glucatell reaction. The unit of study is each independent incubation and measurement reaction. No technical replicates were pooled for statistical testing. Results are presented as mean ± SD.
Aniline blue staining and confocal microscopy
Visualization of (1,3)-β-glucan was performed using aniline blue staining65. C. albicans ATCC 90028 overnight cultures were subcultured into YPD at an OD₆₀₀ of 0.1 and incubated with shaking at 30 °C for 6 h in the presence of coniotin A (½ MIC) or an equivalent volume of DMSO. Following treatment, cells were washed three times with PBS, resuspended to an OD₆₀₀ of 2, and stained with aniline blue (100 μg/mL) for 20 min. After staining, the cells were washed three times with PBS, adjusted to an OD₆₀₀ of 0.8, and mounted on square coverslips (#1.5, 12-541-AP, Fisherbrand) for imaging. Confocal fluorescence microscopy was performed using a Zeiss LSM 980 Upright Confocal Microscope (Zeiss Axio Imager.Z2 stand) with a GaAsp-Pmt3 detector using a 63×/1.4 oil-immersion objective. Aniline blue was excited using the 405 nm laser, and fluorescence was collected in the full visible spectrum (585/172 nm bandpass) to assess (1,3)-β-glucan distribution on the cell surface. Images were captured with consistent excitation power, detector gain, scanning speed, and pixel size settings for comparability. β−1,3-glucan particles (Millipore Sigma 346210) were stained with aniline blue and visualized following the same protocol.
TEM
To investigate morphological alterations, C. auris CBS12766 and C. neoformans H99 were cultured in RPMI 1640 medium supplemented with either coniotin A or amphotericin B at half the MIC, or an equivalent volume of DMSO as a vehicle control, until visible growth was observed. Cells were harvested by centrifugation and resuspended in a fixative solution containing 4% paraformaldehyde (PFA) and 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.4) with 1% Triton X-100. The samples were fixed for 15 min at room temperature and stored overnight at 4 °C. Samples were dehydrated through a graded ethanol series, embedded in resin, and sectioned using a Leica UCT ultramicrotome99. Ultrathin sections were mounted onto copper grids and post-stained with uranyl acetate and lead citrate. Sections were visualized using a JEOL JEM 1200 EX TEMSCAN transmission electron microscope (JEOL, Peabody, MA, USA) operating at 80 kV. Images were acquired with an AMT 4-megapixel digital camera (Advanced Microscopy Techniques, Woburn, MA, USA) and an AMT_V700 software (version 7.0.0.265).
For the visualization of C. elegans–C. auris infection, nematodes were harvested after 20 h of infection and fixed for 4 h at room temperature in 3.2% formaldehyde and 0.2% glutaraldehyde in 0.15 M sodium cacodylate buffer (pH 7.2), then stored at 4 °C. Worms were then processed using the same dehydration, embedding, sectioning, and staining protocols described above100.
Statistics and reproducibility
All key experiments were independently repeated at least three times with similar results, unless otherwise stated. Biological replicates refer to independently prepared and treated samples (e.g., separate cultures, animals, or reaction mixtures), with each data point in the figures representing one biological replicate. No technical replicates were pooled for statistical analysis. Statistical analyses were conducted using GraphPad Prism (v10.5.0). Welch’s t-tests, unpaired two-tailed Student’s t-tests, and one-way ANOVA with multiple comparison corrections (e.g., FDR via the two-stage step-up method of Benjamini, Krieger, and Yekutieli) were used as appropriate. For survival data (Fig. 4c), Log-rank (Mantel–Cox) and Gehan–Breslow–Wilcoxon tests were performed. For effect size estimation, eta squared and hazard ratios with 95% confidence intervals were reported where applicable. All statistical tests were two-sided unless otherwise noted. Detailed statistical parameters and exact P values are provided in figure legends and Source Data. No data were excluded from the analyses. No statistical method was used to predetermine sample size. Randomization and blinding were applied where appropriate; detailed descriptions are provided in the relevant Methods subsections and the Reporting Summary.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The Whole Genome Shotgun (WGS) sequencing data generated in this study have been deposited in GenBank under BioProject accession code PRJNA1171131101. The genome assemblies are available under the following accession codes: JBILZD000000000 (WAC1325) JBILZF000000000 (WAC1490) JBMVFL000000000 (WAC5858) JBILZE000000000 (WAC11084) JBLVKJ000000000 (WAC11161) JBLVKK000000000 (WAC11175) The HR electrospray ionization mass spectrometry data (HR-ESI-MS and MS/MS) for coniotin A have been deposited in the GNPS/MassIVE repository under the accession code MSV000098505 [https://doi.org/10.25345/C5057D541]102. The dataset is licensed under CC0 1.0 Universal (CC0 1.0). All data generated or analyzed during this study are included in the main article and its Supplementary Information. The NMR spectra and compound annotation tables are provided in the Supplementary Information. Source data for graphs and quantitative figures are provided with this paper. No other data is subject to access restrictions. Requests for additional materials or protocols can be directed to the Lead Corresponding Author, Dr. Gerard D. Wright (wrightge@mcmaster.ca), and will be fulfilled in accordance with institutional and data-sharing guidelines. Source data are provided with this paper.
References
Iliev, I. D. et al. Focus on fungi. Cell 187, 5121–5127 (2024).
Denning, D. W. Global incidence and mortality of severe fungal disease. Lancet Infect. Dis. 24, e428–e438 (2024).
Brown, G. D. et al. Hidden killers: human fungal infections. Sci. Transl. Med. 4, 165rv13 (2012).
Fisher, M. C., Hawkins, N. J., Sanglard, D. & Gurr, S. J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 360, 739–742 (2018).
Lyman, M. et al. Worsening apread of Candida auris in the United States, 2019 to 2021. Ann. Intern. Med. 176, 489–495 (2023).
Ruiz-Gaitán, A. et al. An outbreak due to Candida auris with prolonged colonisation and candidaemia in a tertiary care European hospital. Mycoses 61, 498–505 (2018).
WHO. WHO fungal priority pathogens list to guide research, development and public health action (WHO, 2022) https://www.who.int/publications/i/item/9789240060241.
Kadri, S. S. Key takeaways from the U.S. CDC’s 2019 antibiotic resistance threats report for frontline providers. Crit. Care Med. 48, 939 (2020).
Sabino, R., Veríssimo, C., Pereira, ÁA. & Antunes, F. Candida auris, an agent of hospital-associated outbreaks: Which challenging issues do we need to have in mind?. Microorganisms 8, E181 (2020).
Lockhart, S. R. et al. Simultaneous emergence of multidrug-resistant Candida auris on 3 continents confirmed by whole-genome sequencing and epidemiological analyses. Clin. Infect. Dis. Publ. Infect. Dis. Soc. Am. 64, 134 (2016).
Newman, D. J. & Cragg, G. M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 83, 770–803 (2020).
Wright, G. D. Something old, something new: revisiting natural products in antibiotic drug discovery. Can. J. Microbiol. 60, 147–154 (2014).
Harvey, A. L., Edrada-Ebel, R. & Quinn, R. J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 14, 111–129 (2015).
Walsh, C. T. A chemocentric view of the natural product inventory. Nat. Chem. Biol. 11, 620–624 (2015).
Pye, C. R., Bertin, M. J., Lokey, R. S., Gerwick, W. H. & Linington, R. G. Retrospective analysis of natural products provides insights for future discovery trends. Proc. Natl Acad. Sci. Usa. 114, 5601–5606 (2017).
Shendure, J. & Ji, H. Next-generation DNA sequencing. Nat. Biotechnol. 26, 1135–1145 (2008).
Wang, Y., Zhao, Y., Bollas, A., Wang, Y. & Au, K. F. Nanopore sequencing technology, bioinformatics and applications. Nat. Biotechnol. 39, 1348–1365 (2021).
Cimermancic, P. et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 158, 412–421 (2014).
Kang, H.-S. & Brady, S. F. Arimetamycin A: improving clinically relevant families of natural products through sequence-guided screening of soil metagenomes. Angew. Chem. Int. Ed. Engl. 52, 11063–11067 (2013).
Yamanaka, K. et al. Direct cloning and refactoring of a silent lipopeptide biosynthetic gene cluster yields the antibiotic taromycin A. Proc. Natl. Acad. Sci. USA 111, 1957–1962 (2014).
Culp, E. J. et al. Evolution-guided discovery of antibiotics that inhibit peptidoglycan remodelling. Nature 578, 582–587 (2020).
Thornburg, C. C. et al. NCI program for natural product discovery: a publicly-accessible library of natural product fractions for high-throughput screening. ACS Chem. Biol. 13, 2484–2497 (2018).
Kildgaard, S. et al. A dereplication and bioguided discovery approach to reveal new compounds from a marine-derived fungus Stilbella fimetaria. Mar. Drugs 15, 253 (2017).
Hansen, E. & Andersen, J. H. Screening for marine natural products with potential as chemotherapeutics for acute myeloid leukemia. Curr. Pharm. Biotechnol. 17, 71–77.
Wagenaar, M. M. Pre-fractionated microbial samples-the second generation natural products library at Wyeth. Mol. Basel Switz. 13, 1406–1426 (2008).
Cook, M. A. et al. Lessons from assembling a microbial natural product and pre-fractionated extract library in an academic laboratory. J. Ind. Microbiol. Biotechnol. 50, kuad042 (2023).
Bode, H. B., Bethe, B., Höfs, R. & Zeeck, A. Big effects from small changes: possible ways to explore nature’s chemical diversity. ChemBioChem 3, 619–627 (2002).
Pan, R., Bai, X., Chen, J., Zhang, H. & Wang, H. Exploring structural diversity of microbe secondary metabolites using osmac strategy: a literature review. Front. Microbiol. 10, 294 (2019).
Grkovic, T. et al. National Cancer Institute (NCI) program for natural products discovery: rapid isolation and identification of biologically active natural products from the NCI prefractionated library. ACS Chem. Biol. 15, 1104–1114 (2020).
Wang, M. et al. Sharing and community curation of mass spectrometry data with global natural products social molecular networking. Nat. Biotechnol. 34, 828–837 (2016).
Petras, D. et al. GNPS Dashboard: collaborative exploration of mass spectrometry data in the web browser. Nat. Methods 19, 134–136 (2022).
Kratky, C. & Dobler, M. The crystal structure of enniatin B. Helv. Chim. Acta 68, 1798–1803 (1985).
Sy-Cordero, A. A., Pearce, C. J. & Oberlies, N. H. Revisiting the enniatins: a review of their isolation, biosynthesis, structure determination, and biological activities. J. Antibiot.65, 541–549 (2012).
Terlouw, B. R. et al. PARAS: high-accuracy machine-learning of substrate specificities in nonribosomal peptide synthetases. Preprint at https://doi.org/10.1101/2025.01.08.631717 (2025).
Blin, K. et al. antiSMASH 8.0: extended gene cluster detection capabilities and analyses of chemistry, enzymology, and regulation. Nucleic Acids Res. https://doi.org/10.1093/nar/gkaf334 (2025).
Koglin, A. et al. Structural basis for the selectivity of the external thioesterase of the surfactin synthetase. Nature 454, 907–911 (2008).
Travers, K. J. et al. Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 101, 249–258 (2000).
Wyszynski, F. J., Hesketh, A. R., Bibb, M. J. & Davis, B. G. Dissecting tunicamycin biosynthesis by genome mining: cloning and heterologous expression of a minimal gene cluster. Chem. Sci. 1, 581–589 (2010).
Hu, Y. et al. Identification and proposed relative and absolute configurations of niphimycins C–E from the marine-derived Streptomyces sp. IMB7-145 by genomic analysis. J. Nat. Prod. 81, 178–187 (2018).
Hegde, V. R. et al. Novel fungal metabolites as cell wall active antifungals fermentation, isolation, physico-chemical properties, structure and biological activity. J. Antibiot. 56, 437–447 (2003).
Johnson, A. R. & Carlson, E. E. Collision-induced dissociation mass spectrometry: a powerful tool for natural product structure elucidation. Anal. Chem. 87, 10668–10678 (2015).
Degenkolb, T., Berg, A., Gams, W., Schlegel, B. & Gräfe, U. The occurrence of peptaibols and structurally related peptaibiotics in fungi and their mass spectrometric identification via diagnostic fragment ions. J. Pept. Sci. 9, 666–678 (2003).
Jiao, Y. et al. Genome mining reveals biosynthesis of the antifungal lipopeptaibols, texenomycins, through a hybrid PKS-NRPS system, in the fungus Mariannaea elegans. J. Agric. Food Chem. 73, 226–236 (2025).
Perlatti, B., Nichols, C. B., Alspaugh, J. A., Gloer, J. B. & Bills, G. F. Sphaerostilbellins, new antimicrobial aminolipopeptide peptaibiotics from Sphaerostilbella toxica. Biomolecules 10, 1371 (2020).
Fredenhagen, A., Molleyres, L.-P., Böhlendorf, B. & Laue, G. Structure Determination of Neoefrapeptins A to N: peptides with insecticidal activity produced by the fungus Geotrichum candidum. J. Antibiot. 59, 267–280 (2006).
Fujii, K., Ikai, Y., Oka, H., Suzuki, M. & Harada, K. A nonempirical method using LC/MS for determination of the absolute configuration of constituent amino acids in a peptide: combination of Marfey’s method with mass spectrometry and its practical application. Anal. Chem. 69, 5146–5151 (1997).
Vicente-Garcia, C. & Colomer, I. Lipopeptides as tools in catalysis, supramolecular, materials and medicinal chemistry. Nat. Rev. Chem. 7, 710–731 (2023).
Lazzaro, B. P., Zasloff, M. & Rolff, J. Antimicrobial peptides: application informed by evolution. Science 368, eaau5480 (2020).
Wright, G. D. Antibiotic adjuvants: rescuing antibiotics from resistance. Trends Microbiol 24, 862–871 (2016).
Okoli, I. et al. Identification of antifungal compounds active against Candida albicans using an improved high-throughput Caenorhabditis elegans assay. PLoS One 4, e7025 (2009).
Revie, N. M. et al. Targeting fungal membrane homeostasis with imidazopyrazoindoles impairs azole resistance and biofilm formation. Nat. Commun. 13, 3634 (2022).
Cruz, M. R., Graham, C. E., Gagliano, B. C., Lorenz, M. C. & Garsin, D. A. Enterococcus faecalis inhibits hyphal morphogenesis and virulence of Candida albicans. Infect. Immun. 81, 189–200 (2013).
Maget-Dana, R. & Peypoux, F. Iturins, a special class of pore-forming lipopeptides: biological and physicochemical properties. Toxicology 87, 151–174 (1994).
Morikawa, M., Hirata, Y. & Imanaka, T. A study on the structure–function relationship of lipopeptide biosurfactants. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 1488, 211–218 (2000).
Du, Y. et al. Incorporation of non-canonical amino acids into antimicrobial peptides: advances, challenges, and perspectives. Appl. Environ. Microbiol. 88, e01617–e01622 (2022).
Ding, Y. et al. Impact of non-proteinogenic amino acids in the discovery and development of peptide therapeutics. Amino Acids 52, 1207–1226 (2020).
Toniolo, C. et al. Lipopeptaibols, a novel family of membrane active, antimicrobial peptides. Cell. Mol. Life Sci. CMLS 58, 1179–1188 (2001).
Karle, I. L. Controls exerted by the Aib residue: helix formation and helix reversal. Biopolymers 60, 351–365 (2001).
Anderson, J. B. Evolution of antifungal-drug resistance: mechanisms and pathogen fitness. Nat. Rev. Microbiol. 3, 547–556 (2005).
Xu, D. et al. Genome-wide fitness test and mechanism-of-action studies of inhibitory compounds in Candida albicans. PLoS Pathog. 3, e92 (2007).
Richter, M. F. et al. Predictive compound accumulation rules yield a broad-spectrum antibiotic. Nature 545, 299–304 (2017).
Chen, S. C.-A., Slavin, M. A. & Sorrell, T. C. Echinocandin antifungal drugs in fungal infections: a comparison. Drugs 71, 11–41 (2011).
Gow, N. A. R. & Lenardon, M. D. Architecture of the dynamic fungal cell wall. Nat. Rev. Microbiol. 21, 248–259 (2023).
Walker, L. A. et al. Stimulation of chitin synthesis rescues Candida albicans from Echinocandins. PLoS Pathog. 4, e1000040 (2008).
Okada, H. & Ohya, Y. Fluorescent labeling of yeast cell wall components. Cold Spring Harb. Protoc. https://doi.org/10.1101/pdb.prot085241 (2016).
Sudbery, P. E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 9, 737–748 (2011).
Okada, H., Ohnuki, S., Roncero, C., Konopka, J. B. & Ohya, Y. Distinct roles of cell wall biogenesis in yeast morphogenesis as revealed by multivariate analysis of high-dimensional morphometric data. Mol. Biol. Cell 25, 222–233 (2014).
El-Kirat-Chatel, S. et al. Nanoscale analysis of caspofungin-induced cell surface remodelling in Candida albicans. Nanoscale 5, 1105–1115 (2013).
Aketagawa, J., Tanaka, S., Tamura, H., Shibata, Y. & Saitô, H. Activation of limulus coagulation factor G by several (l→3)-β-D-glucans: comparison of the potency of glucans with identical degree of polymerization but different conformations. J. Biochem. 113, 683–686 (1993).
Bizerra, F. C. et al. Changes in cell wall synthesis and ultrastructure during paradoxical growth effect of caspofungin on four different Candida species. Antimicrob. Agents Chemother. 55, 302–310 (2011).
Garcia-Rubio, R., de Oliveira, H. C., Rivera, J. & Trevijano-Contador, N. The fungal cell wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 10, 2993 (2020).
Horton, M. V. et al. Candida auris cell wall mannosylation contributes to neutrophil evasion through pathways divergent from Candida albicans and Candida glabrata. mSphere. https://doi.org/10.1128/msphere.00406-21 (2021).
Zhao, P. et al. Fungi-derived lipopeptide antibiotics developed since 2000. Peptides 113, 52–65 (2019).
Hoste, A. C. R. et al. The structure of lipopeptides impacts their antiviral activity and mode of action against SARS-CoV-2 in vitro. Appl. Environ. Microbiol. 0, e01036–24 (2024).
Hou, X. et al. Peptaibols: diversity, bioactivity, and biosynthesis. Eng. Microbiol. 2, 100026 (2022).
Jiang, K. et al. Clarification of the biosynthetic gene cluster involved in the antifungal prodrug echinocandin B and its robust production in engineered Aspergillus pachycristatus. Microbiol. Res. 293, 128069 (2025).
Harwani, D., Barupal, S., Begani, J. & Lakhani, J. Chapter 2 - Genetic diversity of polyketide synthases and nonribosomal peptide synthetases in fungi. In New and Future Developments in Microbial Biotechnology and Bioengineering (eds. Singh, J. & Gehlot, P.) 11–21 (Elsevier, 2020); https://doi.org/10.1016/B978-0-12-821008-6.00002-5.
Pereira-Dias, L. et al. Peptaibiotics: Harnessing the potential of microbial secondary metabolites for mitigation of plant pathogens. Biotechnol. Adv. 68, (2023).
Ma, S. M. et al. Complete reconstitution of a highly reducing iterative polyketide synthase. Science 326, 589–592 (2009).
Gilchrist, C. L. M. & Chooi, Y.-H. clinker & clustermap.js: automatic generation of gene cluster comparison figures. Bioinforma. Oxf. Engl. 37, 2473–2475 (2021).
Chiang, Y.-M. et al. Molecular genetic mining of the Aspergillus secondary metabolome: Discovery of the emericellamide biosynthetic pathway. Chem. Biol. 15, 527–532 (2008).
Lai, H.-E. et al. Calcium-dependent lipopeptide antibiotics against drug-resistant pathogens discovered via host-dependent heterologous expression of a cloned biosynthetic gene cluster. Angew. Chem. 136, e202410286 (2024).
Hegde, V. R. et al. Novel fungal metabolites as cell wall active antifungals: fermentation, isolation, physico-chemical properties, structure and biological activity. J. Antibiot. 54, 74–83 (2001).
Lorito, M., Farkas, V., Rebuffat, S., Bodo, B. & Kubicek, C. P. Cell wall synthesis is a major target of mycoparasitic antagonism by Trichoderma harzianum. J. Bacteriol. 178, 6382–6385 (1996).
Lee, J. W., Collins, J. E., Wendt, K. L., Chakrabarti, D. & Cichewicz, R. H. Leveraging peptaibol biosynthetic promiscuity for next-generation antiplasmodial therapeutics. J. Nat. Prod. 84, 503–517 (2021).
Guilhelmelli, F. et al. Antibiotic development challenges: the various mechanisms of action of antimicrobial peptides and of bacterial resistance. Front. Microbiol. 4, 353 (2013).
Schein, J. E. et al. Physical maps for genome analysis of serotype A and D strains of the fungal pathogen Cryptococcus neoformans. Genome Res. 12, 1445–1453 (2002).
Yamamura, D., Rotstein, C., Nicolle, L. E., Ioannou, S. & Disease, S. F. D. R. of the C. I. Candidemia at selected Canadian sites: results from the Fungal Disease Registry, 1992–1994. CMAJ 160, 493–499 (1999).
Stewart, J. I. P. et al. Reducing Aspergillus fumigatus virulence through targeted dysregulation of the conidiation pathway. mBio 11, e03202-19 (2020).
Mangat, C. S., Bharat, A., Gehrke, S. S. & Brown, E. D. Rank ordering plate data facilitates data visualization and normalization in high-throughput screening. SLAS Discov. 19, 1314–1320 (2014).
Metin, B., Findley, K. & Heitman, J. The mating type locus (MAT) and sexual reproduction of Cryptococcus heveanensis: insights into the evolution of sex and sex-determining chromosomal regions in fungi. PLoS Genet. 6, e1000961 (2010).
Magoč, T. & Salzberg, S. L. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 27, 2957–2963 (2011).
Lagesen, K. et al. RNAmmer: consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 35, 3100–3108 (2007).
Manni, M., Berkeley, M. R., Seppey, M., Simão, F. A. & Zdobnov, E. M. BUSCO update: novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 38, 4647–4654 (2021).
Deatherage, D. E. & Barrick, J. E. Identification of mutations in laboratory-evolved microbes from next-generation sequencing data using breseq. In Engineering and Analyzing Multicellular Systems: Methods and Protocols (eds. Sun, L. & Shou, W.) 165–188 (Springer, New York, 2014). https://doi.org/10.1007/978-1-4939-0554-6_12.
Iyer, K. R. et al. Identification of triazenyl indoles as inhibitors of fungal fatty acid biosynthesis with broad-spectrum activity. Cell Chem. Biol. 30, 795–810.e8 (2023).
Stirling, D. R. et al. CellProfiler 4: improvements in speed, utility and usability. BMC Bioinforma. 22, 433 (2021).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Wright, R. Transmission electron microscopy of yeast. Microsc. Res. Tech. 51, 496–510 (2000).
Kovács, A. L. The application of traditional transmission electron microscopy for autophagy research in Caenorhabditis elegans. Biophys. Rep. 1, 99–105 (2015).
Chen, X. & Guitor, A. K. Genome sequencing of strains from a natural product fractionation library. GenBank: PRJNA1171131; https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1171131 (2025).
Chen, X. & Koteva, K. GNPS-coniotin high-resolution mass spectrometry data. MassIVE: MSV000098505; https://doi.org/10.25345/C5057D541 (2025).
Acknowledgements
This work was supported by the Canadian Institutes for Health Research [Foundation grant FRN-148463 to G.D.W.]. L.E.C. is supported by the Canadian Institutes of Health Research (CIHR) Foundation grant (FDN-154288) and a National Institutes of Health (NIH) R01 grant (R01AI127375). L.E.C. is a Canada Research Chair (Tier 1) in Microbial Genomics & Infectious Disease and co-director of the CIFAR Fungal Kingdom: Threats & Opportunities program. CIFAR Catalyst Grants: CP21-065; Cowen, L. (University of Toronto); Heitman, J. (Duke University); Wright, G. (McMaster University); and Boone, C (University of Toronto). This research was supported by a Tier 1 Canada Research Chair award, a Foundation grant from the Canadian Institutes of Health Research (CIHR; FRN 143215) to E.D.B., and a CIHR grant PJT-156067 to L.M. The authors wish to express their sincere gratitude to the Centre for Microbial Chemical Biology (CMCB), particularly Tracey Campbell, Susan McCusker, and Nicola Henriquez, as well as the Canadian Centre for Electron Microscopy (CCEM), with special thanks to Marcia Reid, and the McMaster Centre for Advanced Light Microscopy (CALM), with appreciation for the support of Mouhanad Babi and Joao Pedro Bronze de Firmino. We are deeply grateful to Dr. Joseph Heitman, Dr. Jarrod Johnson, Dr. Matt Surette, Dr. Alexander Anderson, Maya George, and Andrea Alexei for their insightful suggestions, discussions, and support.
Author information
Authors and Affiliations
Contributions
X.C. and G.D.W. conceived the study, designed experiments, interpreted data, and wrote the manuscript. D.P. conducted and analyzed the screen, while X.C. and M.A.C. performed additional statistical analysis. X.C. purified the compounds, K.K. synthesized the coniotin A-BODIPY, and K.K. and X.C. conducted structural elucidation and data analysis. X.C. prepared the genomic DNA, and A.G. performed genome sequencing and assembly. S.C. and X.C. conducted the C. elegans animal study. X.C., N.R., D.S., and Y.L. investigated the MOA, with Y.L. and N.R. conducting the HIP screen. S.F. conducted widefield microscopy. X.C. and D.H. performed the bioinformatics analysis. X.C. performed all other experiments. E.D.B., L.T.M., L.E.C., and G.D.W. provided resources.
Corresponding author
Ethics declarations
Competing interests
L.E.C. is a co-founder and shareholder in Bright Angel Therapeutics, a platform company for the development of novel antifungal therapeutics. E.D.B. is the CEO, and L.E.C. and G.D.W. are Science Advisors for Kapoose Creek, a company that harnesses the therapeutic potential of fungi. All other authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks Kenichi Yokoyama, Thomas Ostenfeld-Larsen, and Scott Jarmusch for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Chen, X., Koteva, K., Chou, S. et al. Coniontins, lipopetaibiotics active against Candida auris identified from a microbial natural product fractionation library. Nat Commun 16, 7337 (2025). https://doi.org/10.1038/s41467-025-62630-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-025-62630-z
