
Abstract
H9N2 influenza virus infections represent a significant respiratory health concern, yet the functional role of gut microbiota during infection progression remains poorly understood. Here, we show that H9N2 infection causes dose-dependent alterations in gut microbial communities in a mammalian infection model, particularly the depletion of Prevotella species. Prophylactic administration of Prevotella copri improved survival and clinical outcomes in infected mice by restructuring the gut microbiome, promoting beneficial bacteria, and suppressing pathogens. Metabolomic profiling revealed increased isovaleric acid levels in the intestine and serum. Isovaleric acid pretreatment reduced pulmonary inflammation, alleviated tissue damage, and preserved epithelial integrity. Isovaleric acid pretreatment alleviates lung inflammation, reduces tissue damage, and maintains epithelial integrity. Additionally, isovaleric acid mitigates infection caused by the H1N1 influenza virus. These findings highlight the immunomodulatory role of gut commensals and their metabolites in antiviral defense, offering a new approach to influenza virus treatment.
Similar content being viewed by others


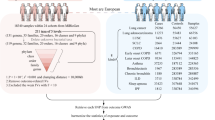
Introduction
H9N2 avian influenza virus has emerged as a significant public health concern due to increasing human infections1,2,3,4. This virus serves as a genetic donor for multiple human-infective avian influenza strains, including H7N9 and H10N8, contributing to generating novel influenza variants and facilitating zoonotic spillover events. Specifically, The H9N2 virus contributed its polymerase genes, PB2 and PA, to the H7N9 virus through reassortment, enhancing the replication capacity of H7N9 in human cells and thereby increasing its cross-species transmissibility and infectivity5. The virus demonstrates remarkable adaptability, exhibiting enhanced binding affinity to human receptors and improved replication efficiency in mammalian cells6. Clinical manifestations of H9N2 infection range from mild respiratory symptoms to severe pneumonia, with inflammatory responses pivotal in disease progression7. The virus has evolved diverse strategies to evade host immune responses, including suppression of interferon production and modulation of cell death pathways8. Current prevention and control measures, primarily based on vaccination and biosecurity, have shown limited effectiveness due to rapid viral evolution, with a primary focus on poultry hosts9. The emergence of antiviral-resistant strains further complicates treatment options. Recent studies have highlighted the emerging antiviral resistance among various influenza strains, including H9N2, which is complicating treatment strategies10. The complex interplay between viral pathogenesis and host immune responses suggests novel therapeutic approaches are needed11. Understanding the role of virus-host interactions in disease progression is crucial for developing effective interventions.
The gut microbiota has emerged as a critical orchestrator of host immunity and disease resistance12. Recent advances in microbiome research have unveiled extensive communication networks between intestinal bacteria and diverse physiological systems13. Disruption of microbial communities has been linked to increased susceptibility to respiratory infections through alterations in immune homeostasis14. Specific bacterial species have been identified as key modulators of antiviral immunity, capable of enhancing both innate and adaptive immune responses15,16,17. The gut-lung axis represents a crucial pathway through which intestinal microbes influence respiratory health. Microbiota-derived signals have been shown to regulate immune cell development and function, including T lymphocytes and innate lymphoid cells (ILCs)18. The composition of gut microbiota significantly influences vaccine responses and infection outcomes. Mounting evidence underscores the microbiota’s role in maintaining barrier integrity and modulating inflammation19. Through interactions with pattern recognition receptors and metabolite-sensing pathways, commensal bacteria shape local and systemic immune responses20. Advanced sequencing technologies have enabled detailed characterization of microbiome alterations during viral infections, revealing dynamic changes in bacterial communities that correlate with disease progression and resolution.
Short-chain fatty acids (SCFAs), primary metabolites produced by gut bacteria, have emerged as crucial mediators of host-microbe interactions21. Recent investigations have revealed diverse immunomodulatory functions of SCFAs, particularly in regulating T cell differentiation and function22. The widespread distribution and expression patterns of SCFAs receptors across tissues suggest their broad physiological impact23. Different SCFAs exhibit distinct biological activities, with butyrate, propionate, and acetate being the most extensively studied24,25,26. These metabolites modulate immune responses through multiple mechanisms, including histone deacetylase inhibition and G protein-coupled receptor activation27,28. SCFAs influence barrier function and inflammatory responses in both intestinal and respiratory tissues, with recent evidence suggesting tissue-specific effects mediated through distinct signaling pathways29. Bacteroides-derived isovaleric acid has been shown to enhance mucosal immunity by promoting intestinal IgA responses in chickens30. In addition, isovaleric acid induces relaxation of colonic smooth muscle via the cAMP/PKA pathway31. The therapeutic potential of SCFAs in various disease contexts has gained increasing attention.
Our study addresses a critical gap in understanding the role of microbiota-derived metabolites in viral infections. We provide novel insights into potential therapeutic strategies by investigating the protective effects of P. copri and its metabolite isovaleric acid against H9N2 and H1N1 infection. These findings have significant therapeutic implications. Ultimately, this research establishes a new paradigm for treating respiratory viral diseases by targeting microbiota-immune interactions.
Results
Gut microbiota composition significantly correlates with H9N2 infection severity
To investigate the role of gut microbiota during H9N2 infection, we established a mouse model using three viral titers (H9N2H, H9N2M, and H9N2L, corresponding to 104.5-106.5 EID50) (Fig. 1a). Survival analysis (Mice were euthanized if they experienced more than 20% weight loss or showed symptoms like lethargy, difficulty breathing, or unresponsiveness, according to the humane endpoints.) revealed a dose-dependent mortality pattern, with the H9N2H group showing the most severe outcome, while H9N2M and H9N2L groups maintained higher survival rates of 40% and 60%, respectively (Fig. 1b). This differential virulence was further reflected in body weight dynamics, where H9N2H-infected mice exhibited the most dramatic weight loss (approximately 20% reduction) during days 4-8 post-infection, followed by a gradual recovery phase in surviving animals (Fig. 1c). Evaluation of the relative viral load in lung tissues by TCID50 assay confirmed that the H9N2H group had the highest viral load, while the viral loads in the H9N2L and H9N2M groups decreased significantly (Fig. 1d). Histopathological examination revealed dose-dependent lung injury, with the H9N2H group showing the most severe tissue damage and inflammatory infiltration (Fig. 1e, f). Assessment of inflammatory markers in both lung tissue and serum revealed significantly elevated levels of pro-inflammatory cytokines (IL-6, TNF-α, and IL-1β) in the H9N2H group (Fig. 1g). Immunofluorescence analysis of H9N2-NA in lung tissue indicated significant viral protein expression in the H9N2H group, with lower levels observed in H9N2M and H9N2L groups (Fig. 1h, i). Intriguingly, analysis of barrier integrity markers revealed significant disruption of epithelial barrier function, evidenced by decreased expression of occludin and ZO-1 mRNA levels, particularly in H9N2H-infected mice (Fig. 1j). Next, we performed 16S rRNA sequencing of the gut microbiota. Following H9N2 infection, we observed significant perturbations in gut microbial diversity profiles in mice (Fig. 1k). Hierarchical clustering analysis revealed distinct microbial community structures among different infection groups, with Prevotella abundance notably decreasing in a dose-dependent manner with infection severity (Fig. 1l). Correlation analysis further demonstrated significant negative associations between Prevotella abundance and inflammatory markers, while showing positive correlations with barrier function markers, suggesting a potential link between Prevotella and H9N2 infection (Fig. 1m).

a Establishment of H9N2 infection model; H9N2H, 106.5 EID50, n = 5; H9N2M, 105.5 EID50, n = 5; H9N2L, 104.5 EID50, n = 5; NC, n = 5. b Survival rates of mice across different groups post-H9N2 infection. c Body weight loss percentage in different groups following H9N2 infection. d Viral load determination in lung tissue by TCID50 assay using MDCK cells; n = 3/group. e Comparison of histopathological changes; images taken at 100× and 200× magnifications. f Histopathological scoring of lung tissues; n = 3/group. g IL-6, TNF-α, and IL-1β levels in serum and lung tissue; n = 3/group. h Immunofluorescence analysis of lung tissue using H9N2-NA-FITC and DAPI; images taken at 200× and 400× magnifications. i Analysis of mean fluorescence intensity of H9N2-NA; n = 3/group. j Relative mRNA expression of occludin and ZO-1 in lung tissue; n = 3/group. k Analysis of alpha diversity of intestinal microbiota; Shannon, Chao1, Observed species, and Simpson. l 16S rRNA analysis of fecal samples shows bacterial genera’s relative abundance across groups; n = 3/group. m Correlation analysis between dominant bacterial genera and infection-related parameters, calculated using Pearson correlation coefficient. Samples for (d–m) were collected on day 6 after H9N2 infection. The data in (b–j) are representative of three independent experiments, yielding an identical pattern of results. Results are shown as means ± SD, with statistical significance determined by one-way ANOVA for three and four groups. *P < 0.05; **P < 0.01; ***P < 0.01; ****P < 0.0001.
Prevotella copri ameliorates H9N2 infection
We established a Prevotella pretreatment model to investigate its protective effects against H9N2 infection. To simulate extreme infection conditions and evaluate the protective efficacy of Prevotella, a high dose of H9N2 (106.5 EID50) was used for infection. Prior to infection, mice were administered P. copri (108 CFU) for 7 consecutive days (Fig. 2a), and effective colonization of P. copri was confirmed (Supplementary Fig. 1a). Remarkably, P. copri pretreatment (PRE + H9N2) significantly improved host survival compared to H9N2-only infection (50% versus 12.5% survival at day 14) (Fig. 2b). This protection was paralleled by attenuated body weight loss in the PRE + H9N2 group, while the H9N2-only group showed severe weight reduction reaching approximately 20% by day 8 post-infection (Fig. 2c). Relative evaluation of viral loads indicated that P. copri pretreatment reduced viral replication compared with infection with H9N2 alone (Fig. 2d). The protective mechanism of P. copri was further characterized through inflammatory profiling. P. copri pretreatment markedly suppressed both systemic and pulmonary inflammatory responses, evidenced by reduced levels of pro-inflammatory cytokines (IL-6, TNF-α, and IL-1β) in both serum and lung tissue (Fig. 2e). Notably, P. copri pretreatment preserved epithelial barrier integrity during H9N2 infection, demonstrated by maintained expression of tight junction proteins ZO-1 and occludin (Fig. 2f). Histopathological examination revealed substantially reduced lung injury in PRE + H9N2 mice, characterized by decreased inflammatory infiltration and preserved alveolar structure (Fig. 2g, h). Immunofluorescence analysis of H9N2-NA in lung tissues further confirmed that P. copri could reduce viral expression (Fig. 2i, j).

a The P. copri protection model was established, with mice receiving daily oral administration of 108 CFU P. copri from day -8 to day -1 before H9N2 infection; control groups received PBS. PRE + H9N2, n = 8; H9N2, n = 8; PRE, n = 8; NC, n = 8. b Survival rates across different groups following H9N2 infection. c Body weight loss percentage in different groups post-H9N2 infection. d Viral load quantification in lung tissue by TCID50 assay using MDCK cells. e IL-6, TNF-α, and IL-1β levels in serum and lung tissue; n = 3/group. f Relative mRNA expression of ZO-1 and occludin in lung tissue; n = 3/group. g Comparison of histopathological changes; images taken at 100× and 200× magnifications. h Histopathological scoring of lung tissues; n = 3/group. i Immunofluorescence analysis of lung tissue using H9N2-NA-FITC and DAPI; images taken at 200× and 400× magnifications. j Analysis of mean fluorescence intensity of H9N2-NA; n = 3/group. Samples for (d–j) were collected on day 6 after H9N2 infection. The data in (b–j) are representative of three independent experiments, yielding an identical pattern of results. Results are shown as means ± SD, with statistical significance determined by t tests for two groups and one-way ANOVA for four groups. *P < 0.05; **P < 0.01; ***P < 0.01; ****P < 0.0001.
P. copri ameliorates H9N2 infection through modulation of gut microbiota composition
We established a microbiota depletion model to investigate whether P. copri functions dependently on gut microbiota (Fig. 3a). After antibiotic treatment, P. copri was continuously administered for 7 days to ensure its effective colonization in the mice’s intestines (Supplementary Fig. 1b). Notably, P. copri lost its protective effects against H9N2 infection in the depleted gut environment, indicating that interactions between gut microbiota and P. copri are essential for its function (Fig. 3b, c). To investigate the impact of P. copri intervention on gut microbiota during H9N2 infection, we performed 16S rRNA sequencing analysis. Rarefaction curves demonstrated species richness and evenness (Supplementary Fig. 2a). Venn diagram analysis revealed distinct operational taxonomic unit (OTU) distributions among groups, with P. copri supplementation significantly reducing OTU numbers in both H9N2-infected and healthy mice. The number of shared OTUs between H9N2 groups was 401 (Fig. 3d). Principal coordinate analysis (PCoA) and NMDS plots demonstrated clear clustering patterns, indicating that both H9N2 infection and P. copri intervention significantly altered microbial community structures (Fig. 3e). Alpha diversity analysis revealed that P. copri pretreatment significantly modulated microbial diversity indices during H9N2 infection. The PRE + H9N2 group showed significantly lower Shannon, Simpson, and Pielou-e indices compared to the H9N2-only group (Fig. 3f). Taxonomic analysis at the phylum level showed that P. copri pretreatment significantly increased Firmicutes abundance while reducing Bacteroidota abundance. At the genus level, P. copri pretreatment enhanced the abundance of Lactobacillus, Dubosiella, and Ligilactobacillus while reducing Desulfovibrio abundance (Fig. 3g, h). Quantitative analysis of species contributions to differences between PRE + H9N2 and H9N2 groups revealed high contributions from commensal probiotics Lactobacillus and Ligilactobacillus following P. copri pretreatment (Fig. 3i). This bacterial contribution pattern was consistent between PRE and NC groups (Supplementary Fig. 2b). Linear discriminant analysis (LEfSe) identified key bacterial taxa distinguishing different groups. Notably, beneficial bacteria, including Lactobacillaceae, were enriched in the PRE + H9N2 group (Fig. 3j). Cladogram analysis further illustrated the phylogenetic distribution of these differential microbes (Fig. 3k). Correlation analysis showed that Lactobacillus and Ligilactobacillus were negatively correlated with inflammatory markers and positively correlated with barrier proteins ZO-1 and occludin (Fig. 3l), suggesting that P. copri-mediated microbiota may potentially regulate H9N2 infection.

a Establishment of gut microbiota depletion model; n = 6/group. b Survival rates of mice across different groups post-H9N2 infection. c Body weight loss percentage in different groups following H9N2 infection. d Comparative analysis of OTU numbers among groups; n = 6/group. e PCoA and NMDS analysis based on weighted UniFrac distances to identify microbial structural changes. f Comparative analysis of Simpson, Pielou, Shannon, and observed features indexes. g Relative abundance analysis at phylum level. h Relative abundance analysis at genus level. i Simper analysis shows the top 20 species with the highest contribution to differences between H9N2 and PRE + H9N2 groups and their abundance. j LEfSe analysis; LDA score > 3. k Bacterial phylogenetic branches, with different circle layers from inner to outer representing seven taxonomic levels: domain, phylum, class, order, family, genus, and species; each node represents a species classification at that level. l Correlation analysis between dominant bacterial genera and infection-related parameters, calculated using Pearson correlation coefficient. Six samples in each group were used for 16S rRNA analysis. Samples for d-l were collected on day 6 after H9N2 infection. The data in panels b, c are representative of three independent experiments, yielding an identical pattern of results. Results are shown as means ± SD, with statistical significance determined by one-way ANOVA for four groups. *P < 0.05; **P < 0.01; ***P < 0.001.
P. copri intervention enhances isovaleric acid and isobutyric acid production during H9N2 infection
SCFAs derived from gut microbiota play crucial roles in host health and immune-mediated diseases32. To explore the metabolic impact of P. copri intervention during H9N2 infection, we systematically analyzed SCFAs in both systemic and gut compartments, given their crucial role in host health and immune-mediated diseases. Remarkably, P. copri pretreatment (PRE + H9N2) led to significantly elevated isovaleric acid and isobutyric acid levels in both serum and fecal samples compared to the H9N2-only group (Fig. 4a, b). While other SCFAs, including acetic acid, butyric acid, propionic acid, and valeric acid, showed varying trends, the most pronounced and consistent change was observed in isovaleric acid concentrations across both compartments. We performed a correlation analysis between SCFAs profiles and bacterial taxa abundances identified from our previous 16S rRNA sequencing to decipher the microbial contributors to this metabolic shift. The correlation heatmap revealed that isovaleric acid and isobutyric acid levels from both serum and feces strongly positively correlated with several beneficial bacteria, particularly Lactobacillus, Prevotellaceae_UCG-001, and Ligilactobacillus, suggesting active microbial production of isovaleric acid in the gut environment (Fig. 4c). Moreover, analysis revealed inverse correlations between isovaleric acid, isobutyric acid, and potentially pathogenic bacteria such as Enterococcus and Mucispirillum, indicating a possible role of isovaleric acid in maintaining microbial homeostasis.

a Quantification of short-chain fatty acids in serum samples from PRE + H9N2 and H9N2 groups on day 6; n = 4/group. b Analysis of short-chain fatty acids in fecal samples from PRE + H9N2 and H9N2 groups on day 6; n = 4/group. c Correlation analysis between short-chain fatty acids in serum and feces with infection-related parameters, calculated using Pearson correlation coefficient. Samples for (a, b) were collected on day 6 after H9N2 infection. Results are shown as means ± SD, with statistical significance determined by t tests for two groups. *P < 0.05; **P < 0.01.
Isovaleric acid supplementation prevents H9N2 infection
To validate the protective role of isovaleric acid and isobutyric acid against H9N2 infection, we established a pretreatment model where mice received isovaleric acid (0.2% in drinking water) and isobutyric acid (0.2% in drinking water) for 7 days before the H9N2 challenge (Fig. 5a). Notably, isobutyric acid did not provide significant protection (Supplementary Fig. 3a, b), whereas isovaleric acid pretreatment (IA + H9N2) significantly improved survival rates compared to H9N2-only infection (40% versus 0% survival at day 14) (Fig. 5b). This protection was accompanied by markedly reduced weight loss in the IA + H9N2 group throughout the infection period (Fig. 5c). Furthermore, the relative viral load was assessed by TCID50 assay, and the results showed that isovaleric acid pretreatment modestly reduced the viral load compared with the H9N2 group (Fig. 5d). Inflammatory profiling revealed that isovaleric acid pretreatment markedly suppressed both systemic and pulmonary inflammatory responses, evidenced by reduced levels of pro-inflammatory cytokines (IL-6, TNF-α, and IL-1β) in both serum and lung tissue (Fig. 5e). Importantly, isovaleric acid maintained the integrity of the epithelial barrier during H9N2 infection and increased the expression of tight junction proteins ZO-1 and occludin (Fig. 5f). Histopathological examination revealed substantially attenuated lung injury in IA + H9N2 mice, characterized by preserved alveolar structure and reduced inflammatory infiltration (Fig. 5g, h). Moreover, isovaleric acid treatment resulted in a reduction in H9N2-NA viral expression in lung tissues (Fig. 5i, j). However, isovaleric acid pretreatment did not enhance the expression of IgG and IgM in serum or sIgA in the lungs of mice (Supplementary Fig. 4).

a Establishment of isovaleric acid protection model: mice received drinking water containing isovaleric acid from day -8 to day -1 prior to H9N2 infection; control groups received PBS. IA + H9N2, n = 8; H9N2, n = 8; IA, n = 8; NC, n = 8. b Survival rates across different groups following H9N2 infection. c Body weight loss percentage in different groups post-H9N2 infection. d Viral load quantification in lung tissue by TCID50 assay using MDCK cells. e IL-6, TNF-α, and IL-1β levels in serum and lung tissue; n = 3/group. f Relative mRNA expression of ZO-1 and occludin in lung tissue; n = 3/group. g Comparison of histopathological changes; images taken at 100× and 200× magnifications. h Histopathological scoring of lung tissues; n = 3/group. i Immunofluorescence analysis of lung tissue using H9N2-NA-FITC and DAPI; images taken at 200× and 400× magnifications. j Analysis of mean fluorescence intensity of H9N2-NA; n = 3/group. Samples for (d–j) were collected on day 6 after H9N2 infection. The data in (b–j) are representative of three independent experiments, yielding an identical pattern of results. Results are shown as means ± SD, with statistical significance determined by t tests for two groups and one-way ANOVA for four groups. *P < 0.05; **P < 0.01; ***P < 0.01; ****P < 0.0001.
Isovaleric acid reduces H1N1 infection in mice
To evaluate the broader applicability of isovaleric acid, we employed a mouse-adapted H1N1 influenza virus as a model pathogen and applied an experimental protocol identical to that described previously (Fig. 5a). Remarkably, isovaleric acid demonstrated comparable protective effects in the H1N1 infection model, significantly attenuating disease severity. Survival analysis revealed that mice in the H1N1 group began to succumb by day 5 post-infection, with survival dropping to 10% by day 8. In contrast, mice in the IA + H1N1 group exhibited a markedly improved survival rate of 50% at the same time point (Fig. 6a). Consistently, isovaleric acid treatment ameliorated body weight loss typically observed during infection (Fig. 6b). Histopathological examination further confirmed these protective effects. Lungs from H1N1-infected mice displayed extensive parenchymal damage, dense inflammatory cell infiltration, and pronounced alveolar wall thickening, whereas lungs from IA-treated mice showed substantially reduced inflammatory infiltration and only mild alveolar thickening (Fig. 6c). Collectively, these findings suggest that isovaleric acid confers cross-subtype protective effects against influenza virus infection.

a Survival rates across different groups following H1N1 infection; n = 10/group. b Body weight loss percentage in different groups post-H1N1 infection. Comparison of histopathological changes; images taken at 200× magnifications. Samples for (c) were collected on day 6 after H1N1 infection. The data in (a–c) are representative of three independent experiments, yielding an identical pattern of results.
Discussion
This study delineates a protective role for the gut commensal Prevotella copri and its metabolite isovaleric acid during infection with influenza virus (Fig. 7). Using a murine infection model, we demonstrate that P. copri pretreatment markedly enhances host survival, mitigates body weight loss, and suppresses viral replication. Most notably, we found that isovaleric acid is a key effector metabolite that not only reduces H9N2 infection but also provides protection against H1N1 influenza virus subtype infection. Our findings uncover a previously unrecognized mechanism by which a commensal microbe modulates antiviral immunity through a specific metabolic product, providing a foundation for the development of microbiota-based antiviral strategies that harness host–microbe–immune system interactions.

H9N2 infection induces distinct alterations in gut microbial communities, characterized by progressive depletion of Prevotella species with increasing infection severity. P. copri supplementation not only attenuates H9N2 infection but also restructures the gut microbiota composition and SCFA metabolite profiles. Notably, isovaleric acid supplementation mitigates H9N2/N1N1 infection.
The H9N2 virus primarily infects avian species, with relatively few human infections. In contrast, Influenza A viruses (such as H1N1, H3N2, H5N1, and H7N9) have circulated in human populations for extended periods, with these viral strains constantly mutating, leading to immune escape and posing a challenge for influenza control33,34,35,36. When H9N2 undergoes genetic reassortment with highly pathogenic avian influenza viruses like H7N9, the resulting novel viral strains may enhance their adaptability to humans through genetic exchange with human influenza viruses, thereby increasing their transmission potential37. Previous studies have demonstrated the critical role of various gut probiotics in viral defense and immune regulation. B. pseudolongum NjM1 or its metabolite acetate provided protection against influenza virus PR838, while Lactobacillus rhamnosus GG induced cGAS/STING-dependent type I interferon production and modulated responses to immune checkpoint blockade39. Recent research revealed that acetate derived from the commensal bacterium Blautia coccoides mitigated influenza virus infection by modulating virus-specific CD8+ T cells through GPR4340. Additionally, the gut commensal Akkermansia muciniphila was found to produce harmaline, which protected mice from SFTSV (Severe Fever with Thrombocytopenia Syndrome Virus) infection by suppressing NF-κB-mediated systemic inflammation41. In contrast to these studies, our work is the first to elucidate the unique role of Prevotella in antiviral immunity. Recent investigations have established P. copri as a crucial human gut commensal, playing vital roles in maintaining intestinal barrier integrity and regulating inflammatory responses42. While this bacterium has been implicated in bile acid metabolism43, its function during viral infections remained poorly understood. Studies have shown that Prevotella regulates immune responses by activating Toll-like receptor 2, leading to the production of Th17 polarizing cytokines IL-23 and IL-1 by antigen-presenting cells44. Furthermore, Prevotella in the airways promotes TLR2-dependent neutrophil activation and rapid clearance of S. pneumoniae45. Our findings not only expand the known functional repertoire of P. copri but also provide mechanistic insights into its immunomodulatory effects, establishing a new paradigm for probiotic therapeutic applications. Uniquely, we identified isovaleric acid as the key metabolite mediating P. copri’s antiviral effects, demonstrating a novel metabolite-mediated mechanism distinct from previously described probiotic interventions. This mechanistic understanding offers new opportunities for targeted therapeutic development in viral infections.
SCFAs play crucial roles in immune regulation and disease intervention. Recent studies highlight diverse immunomodulatory functions. Butyrate from R. intestinalis enhances anti-PD-1 efficacy by inducing functional CD8 T cells in colorectal cancer prevention46, while microbiota-derived propionate regulates intestinal cholesterol metabolism and immune responses47. Acetate facilitates metabolic adaptation of brain innate immunity and functions as a metabolic immunomodulator48, enhancing anti-tumor immunity in breast cancer through T cell effector modulation49. Isovaleric acid is primarily derived from leucine metabolism. After deamination, leucine is converted into α-ketoisocaproate, which is subsequently decarboxylated and reduced to form isovaleric acid50. Previous studies have shown that the microorganisms Propionibacterium freudenreichii TL 34 and ITGP23 can convert L-leucine into isovaleric acid51. In addition, Lactobacillus gasseri RW2014, isolated from a healthy infant, significantly increased the concentration of isovaleric acid in rat feces when used as a probiotic supplement52. Particularly relevant to our findings, Bacteroides-derived isovaleric acid has been shown to enhance mucosal immunity by promoting intestinal IgA responses in chickens30. Additionally, isovaleric acid supplementation stimulates rumen development by improving ruminal mucosal morphology and function, promoting calf growth53. Our study builds upon these findings by revealing a previously underappreciated role for isovaleric acid in antiviral immunity. This effect is not limited to the H9N2 subtype, as isovaleric acid also confers protection against H1N1 influenza virus infection, highlighting its broad-spectrum potential in combating influenza viruses. This discovery deepens our understanding of how microbial metabolites shape immune function and host defense mechanisms during infection.
The current treatment of influenza virus infections primarily relies on neuraminidase inhibitors, endonuclease inhibitors and M2 ion channel blockers54. However, viral resistance is becoming increasingly prevalent. Recent studies indicate a global rise in oseltamivir-resistant strains, while developing novel antiviral drugs faces numerous challenges55,56. In this context, microbiota-based therapeutic strategies offer promising alternatives. Recent clinical trials have demonstrated that certain probiotics can significantly alleviate flu-like symptoms and reduce viral loads57. Furthermore, microbial metabolites have gained increasing attention as therapeutic targets. Recent research has revealed beneficial effects of SCFAs in modulating host vaccine immunogenicity58. Our study introduces new candidate strains and identifies isovaleric acid as a specific metabolite that could serve as a target for novel antiviral drug development. Despite these significant findings, several limitations warrant consideration. First, our study primarily utilized mouse models, and the translatability of these findings to humans requires further validation. The notable differences between human and mouse gut microbiota may impact the translation of these research outcomes. Second, the pharmacokinetic properties of isovaleric acid in vivo and optimal dosing regimens require systematic investigation. Moreover, the variability in efficacy among different Prevotella strains and the potential roles of other metabolites they produce merit further exploration.
Methods
Ethics approval and consent to participate
The experimental procedures in this study were reviewed and authorized by the Animal Protection and Ethics Committee of Jilin Agricultural University (JLAU20220610001). The committee also monitored the adherence to established protocols throughout the process to ensure ethical compliance.
Bacteria, viruses and cells
The H9N2 (mouse-adapted A/duck/Xuzhou/07/2003) and H1N1 (A mouse-adapted, low-pathogenicity isolated of IAV H1N1 strain PR840) avian influenza virus was propagated in 9-day-old specific pathogen-free embryonated chicken eggs at 37 °C. After 48 h of incubation, allantoic fluid was harvested, clarified by centrifugation (3000 × g, 10 min, 4 °C), and filtered through a 0.22 μm filter. Virus stocks were aliquoted and stored at −80 °C. Virus titers were determined by 50% egg infectious dose (EID50) using the Reed-Muench method, with a titer of H9N2 106.5 EID5059.
Madin-Darby Canine Kidney (MDCK) cells were maintained in DMEM supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin at 37 °C with 5% CO2.
Prevotella copri (DSM 18205) was cultured anaerobically in DSMZ Medium 1669 supplemented with 5 μg/mL hemin and 1 μg/mL vitamin K1 at 37 °C in an anaerobic chamber (10% CO2, 10% H2, 80% N2). After 48 h of cultivation, bacterial cells were harvested by centrifugation (5000 × g, 10 min, 4 °C), washed twice with sterile PBS, and adjusted to 109 CFU/mL using McFarland standards and verified by plate counting.
Animal models
Six-week-old female C57BL/6 specific pathogen-free (SPF) mice were obtained from Changchun Yisi Laboratory Animal Technology Co., Ltd. The mice were housed in an SPF animal facility with free access to water and food. The viral infection experiments were performed under anesthesia to ensure animal welfare and minimize stress responses. Isoflurane was administered as the anesthetic agent using a sealed induction chamber, with 5% concentration for induction and 2–3% for maintenance. Mice were euthanized at the end of the experiment with CO2 asphyxiation with cervical dislocation as a physical secondary assurance method. All animal models were established with parallel experimental groups for sample collection, and feces, lung tissues, and serum were collected on day 6 for subsequent analysis.
H9N2 dose-response study
For the dose-dependent infection model, mice were randomly divided into four groups (n = 5 per group): H9N2H (106.5 EID50), H9N2M (105.5 EID50), H9N2L (104.5 EID50), and NC (Normal Control group). Mice were infected intranasally with 50 μL virus suspension (25 μL per nostril). Body weight changes and survival were monitored daily over 14 days. Fecal samples were collected on day 6 for 16S rRNA sequencing analysis.
P. copri administration
For the bacterial intervention study, mice were randomly divided into four groups (n = 8 per group): PRE (P. copri)+H9N2, H9N2, PRE (P. copri), and NC (Normal Control group). During the pretreatment phase, mice received daily oral gavage of either 100 μL bacterial suspension (108 CFU) or sterile PBS for 7 consecutive days before the H9N2 challenge (106.5 EID50) on day 0. Fecal and serum samples were collected on day 6 for 16S rRNA sequencing and short-chain fatty acid analysis.
Gut microbial depletion
For microbiota depletion studies, mice were randomly divided into five groups (n = 6/group): Abx+PRE (P. copri)+H9N2, PRE (P. copri)+H9N2, Abx+H9N2, H9N2, and NC (Normal Control group). Experimental groups received antibiotic cocktail treatment for 7 days, followed by P. copri supplementation (108 CFU) for 7 days. Mice were then challenged with H9N2 (106.5 EID50) and monitored for infection progression over the subsequent 14 days.
Isovaleric acid treatment
For metabolite intervention studies, mice were randomly divided into four groups (H9N2 infection, n = 8 per group; H1N1 infection, n = 10 per group): IA (Isovaleric acid)+H9N2/H1N1, H9N2/H1N1, IA, and NC (Normal Control group). Isovaleric acid (MedChemExpress, ≥97% purity) was dissolved in drinking water at 0.2% concentration, with solutions freshly prepared daily. Mice received isovaleric acid-supplemented water for 7 consecutive days before H9N2 (106.5 EID50) and H1N1 (106 TCID50 in 40 μL of PBS) challenge on day 0.
Antibiotic treatment
A cocktail of antibiotics consisting of ampicillin (1.0 g/L), neomycin (1.0 g/L), metronidazole (1.0 g/L), and vancomycin (0.5 g/L) was dissolved in sterile drinking water and administered to mice for 7 consecutive days. Bacterial culturing and colony counting methods were employed to validate the effectiveness of gut microbiota depletion. After antibiotic treatment, fecal samples from mice were collected and inoculated onto Columbia-CAN, MacConkey, Sabouraud, and Shaedler agar plates. After culturing, bacterial colonies were manually counted to determine the microbial load. The effectiveness of antibiotic treatment in depleting the gut microbiota was verified by comparing the colony count results with pre-treatment samples. The standard for successful gut microbiota depletion is defined as a reduction of at least 90% in bacterial load in fecal samples after antibiotic treatment (compared to pre-treatment colony counts), with certain bacterial species failing to form cultivable colonies on the agar plates.
Histopathological analysis
Lung tissues were fixed in 4% paraformaldehyde at 4 °C for 24 h, dehydrated through graded ethanol series, cleared in xylene, and embedded in paraffin. Five-micrometer sections were prepared for analysis. For H&E staining, sections were deparaffinized in xylene, rehydrated through a graded ethanol series, stained with hematoxylin for 5 min, differentiated in 1% acid alcohol, treated with 0.2% ammonia water, counterstained with eosin for 2 min, then dehydrated, cleared, and mounted.
Pathological scoring
Lung pathological scoring was conducted based on eight distinct histopathological parameters: alveolar rupture, edema, inflammatory cell infiltration, hemorrhage, fibrosis, connective tissue hyperplasia, capillary proliferation, and airway inflammation. Each parameter was evaluated on a five-point scale (0: no pathological changes; 1: mild changes; 2: moderate changes; 3: marked changes; 4: severe changes). The cumulative score, ranging from 0 (no pathological alterations) to 32 (maximum severity), was calculated by quantitatively summing individual parameter scores to assess the overall pulmonary pathological status. Lung sections were examined under light microscopy and independently scored by two experienced pathologists.
Immunofluorescence analysis
Lung tissue sections (3 μm) were deparaffinized in xylene and rehydrated through a graded ethanol series. Antigen retrieval was performed in citrate buffer (10 mM, pH 6.0) at 95 °C for 20 min. After cooling to room temperature, sections were washed with PBST (PBS containing 0.1% Tween-20) and blocked with 5% BSA in PBS for 1 h at room temperature. Sections were then incubated with primary antibody against H9N2 (SinoBiological, 1:1000 in PBS) overnight at 4 °C. After washing with PBST, sections were incubated with sheep anti-rabbit secondary antibody (Servicebio, 1:600 in PBS) for 1 h at room temperature in the dark. Nuclei were counterstained with DAPI (1:5000) for 5 min. Images were acquired using a fluorescence microscope (Leica Microsystems, Germany) with identical exposure settings across all samples.
ELISA analysis
Blood samples were collected and allowed to clot for 30 min at room temperature. Serum was separated by centrifugation (2000 × g, 15 min, 4 °C) and stored at −80 °C. Lung tissues were homogenized in PBS containing protease inhibitor cocktail. After centrifugation (8000 × g, 10 min, 4 °C), supernatants were collected for analysis. Levels of IL-6, TNF-α, IL-1β, IgG, IgM and sIgA were determined using commercial ELISA kits (MEIMIAN-0040M1, 0163M2, 0132M2, 0057M2, 0430M1, China) according to manufacturer’s instructions.
qPCR
Total RNA was extracted from mouse lung tissues and CTLs using TaKaRa MiniBEST Universal RNA Extraction Kit. Complementary DNA (cDNA) was synthesized from approximately 2 μg total RNA using SweScript All-in-One RT SuperMix for qPCR Kit (Servicebio). The mRNA expression levels of ZO-1 and occludin were analyzed using SYBR Green premix kit (Servicebio) on a real-time fluorescent quantitative PCR detection system. Gene expression was normalized to GAPDH using the 2-ΔΔCt method. The copy number of P. copri was based on previous studies60.Sequences are as follows: mouse ZO-1, forward 5′-GTCCCTCCTCTGATACCTTCCTC-3′, reverse 5′- CTGGCAGTGTCATTCACATCTTTCT-3′; mouse occludin, forward 5′-TACAGACCCAAGAGCAGCA-3′, reverse 5′- AGCCGTACATAGATCCAGAA-3′; mouse GAPDH, forward 5′-AACGTGTCAGTCGTGGACCTG-3′, reverse 5′-AGT GGGTGTCGCTGTFGAAGT-3′; P.copri 16S, forward 5′-CGAAAGCTTGCTTTTGATGG-3′, reverse 5′-CGCAAGGTTATCCCCAAGT-3′.
16S rRNA sequencing and analysis
Microbial DNA was extracted from fecal samples (n = 6/group) using QIAamp PowerFecal Pro DNA Kit, and the V3-V4 region of bacterial 16S rRNA was amplified using primers 515 F (5’-CCTACGGGNGGCWGCAG-3’) and 806 R (5’-GGACTACHVGGGTATCTAAT-3’). PCR was performed using Phusion High-Fidelity PCR Master Mix with the following conditions: 98 °C for 1 min; 30 cycles of 98°C for 10 s, 50 °C for 30 s, and 72 °C for 30 s, followed by 72 °C for 5 min. PCR products were purified using AMPure XP beads, quantified by Qubit, pooled in equimolar amounts, and sequenced on the Illumina NovaSeq platform with paired-end 250 bp reads. Raw sequences were processed using QIIME2 (version 2022.2), where paired-end reads were merged using FLASH (v1.2.11) after removing barcodes and primers. Quality filtering was performed using fastp (v0.23.1), and chimeric sequences were removed by comparison against the Silva database (release 138.1). Clean reads were clustered into ASVs using the DADA2 algorithm, and taxonomy was assigned using the Silva database for bacterial classification. For community analysis, alpha diversity indices were calculated using QIIME2, while beta diversity was analyzed using weighted UniFrac distances. Differential abundance analysis was performed using LEfSe, and statistical significance was determined using ANOSIM and PERMANOVA tests with P < 0.05 considered significant.
SCFA measurement
The levels of SCFAs in feces and serum samples of mice were measured on day 6 after infection (PRE + H9N2 and H9N2, n = 4/group). SCFAs were analyzed using a Thermo Trace 1310 gas chromatograph and an ISQ LT mass spectrometer (Thermo Fisher Scientific, USA). The separation was performed on an Agilent HP-INNOWAX capillary column (30 m × 0.25 mm ID × 0.25 μm). For sample preparation, samples were homogenized with 500 μL water and 100 mg glass beads, followed by centrifugation at 12000 rpm for 10 min at 4 °C. The supernatant (200 μL) was extracted with 100 μL of 15% phosphoric acid, 20 μL of internal standard (4-methylvaleric acid, 375 μg/mL), and 280 μL ether. After centrifugation, the supernatant was analyzed by GC-MS. The GC conditions were: split injection (10:1) with 1 μL injection volume; injector temperature 250 °C; ion source temperature 300 °C; and transfer line temperature 250 °C. The oven temperature was programmed from 90 °C, increased to 120 °C at 10 °C/min, then to 150 °C at 5 °C/min, and finally to 250 °C at 25 °C/min (held for 2 min). Helium was used as carrier gas at 1.0 mL/min. Mass spectrometric detection was performed in electron impact mode (70 eV) using single ion monitoring (SIM).
Statistical analysis
All data are expressed as the mean ± SD. Statistical analysis was performed using one-way ANOVA and t tests with GraphPad Prism 10.0 (GraphPad software). The P values are indicated as follows: *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001.
Data availability
Data Availability Sequencing data of 16s have been uploaded to the NCBI Sequence Read Archive database with registration numbers PRJNA1188328 and PRJNA1188327, respectively. All data supporting the results of this study can be found in the manuscript.
References
Tan, M. et al. Reported human infections of H9N2 avian influenza virus in China in 2021. Front. Public Health 11, 1255969 (2023).
Um, S. et al. Human Infection with Avian Influenza A(H9N2) Virus, Cambodia, February 2021. Emerg. Infect. Dis. 27, 2742–2745 (2021).
Jin, Y. et al. Evidence for human infection with avian influenza A(H9N2) virus via environmental transmission inside live poultry market in Xiamen, China. J. Med. Virol. 95, e28242 (2023).
Ali, S. et al. H9N2 influenza A viruses found to be enzootic in Punjab Pakistan’s bird markets with evidence of human H9N2 nasal colonization. Int. J. Infect. Dis. 146, 107146 (2024).
Pu, J. et al. Reassortment with dominant chicken H9N2 influenza virus contributed to the fifth H7N9 virus human epidemic. J. Virol. 95, e01578-20 (2021).
Zhang, J. et al. Mutational antigenic landscape of prevailing H9N2 influenza virus hemagglutinin spectrum. Cell Rep. 42, 113409 (2023).
Zhang, R. R. et al. Improved pathogenicity of H9N2 subtype of avian influenza virus induced by mutations occurred after serial adaptations in mice. Micro Pathog. 160, 105204 (2021).
Gao, W. et al. Prevailing I292V PB2 mutation in avian influenza H9N2 virus increases viral polymerase function and attenuates IFN-β induction in human cells. J. Gen. Virol. 100, 1273–1281 (2019).
Gu, M., Xu, L., Wang, X. & Liu, X. Current situation of H9N2 subtype avian influenza in China. Vet. Res 48, 49 (2017).
Song, E. J. et al. Inhibitory effects of aprotinin on influenza A and B viruses in vitro and in vivo. Sci. Rep. 11, 9427 (2021).
Hu, P. et al. Citric acid promotes immune function by modulating the intestinal barrier. Int. J. Mol. Sci. 25, 1239 (2024).
Adak, A. & Khan, M. R. An insight into gut microbiota and its functionalities. Cell Mol. Life Sci. 76, 473–493 (2019).
Wang J., Zhu N., Su X., Gao Y., Yang R. Gut-microbiota-derived metabolites maintain gut and systemic immune homeostasis. Cells 12, 793 (2023).
Marsland, B. J., Trompette, A. & Gollwitzer, E. S. The gut-lung axis in respiratory disease. Ann. Am. Thorac. Soc. 12, S150–S156 (2015).
Zhang, Q. et al. Influenza infection elicits an expansion of gut population of endogenous Bifidobacterium animalis which protects mice against infection. Genome Biol. 21, 99 (2020).
Baldridge, M. T. et al. Commensal microbes and interferon-λ determine persistence of enteric murine norovirus infection. Science 347, 266–269 (2015).
Xing, J. H. et al. Gut microbiota-derived LCA mediates the protective effect of PEDV infection in piglets. Microbiome 12, 20 (2024).
Budden, K. F. et al. Emerging pathogenic links between microbiota and the gut-lung axis. Nat. Rev. Microbiol. 15, 55–63 (2017).
Ng, S. C. et al. Gut microbiota composition is associated with SARS-CoV-2 vaccine immunogenicity and adverse events. Gut. 71, 1106–1116 (2022).
Ni, J., Wu, G. D., Albenberg, L. & Tomov, V. T. Gut microbiota and IBD: causation or correlation?. Nat. Rev. Gastroenterol. Hepatol. 14, 573–584 (2017).
Morrison, D. J. & Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 7, 189–200 (2016).
Yang, W. et al. Intestinal microbiota-derived short-chain fatty acids regulation of immune cell IL-22 production and gut immunity. Nat. Commun. 11, 4457 (2020).
Blaak, E. E. et al. Short chain fatty acids in human gut and metabolic health. Benef. Microbes 11, 411–455 (2020).
Stoeva, M. K. et al. Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes 13, 1–28 (2021).
Tian, G. X. et al. Propionic acid regulates immune tolerant properties in B Cells. J. Cell Mol. Med. 26, 2766–2776 (2022).
Hu, C. et al. Gut microbiota-derived short-chain fatty acids regulate group 3 innate lymphoid cells in HCC. Hepatology 77, 48–64 (2023).
Zhao, Y. et al. GPR43 mediates microbiota metabolite SCFA regulation of antimicrobial peptide expression in intestinal epithelial cells via activation of mTOR and STAT3. Mucosal Immunol. 11, 752–762 (2018).
Schilderink, R. et al. The SCFA butyrate stimulates the epithelial production of retinoic acid via inhibition of epithelial HDAC. Am. J. Physiol. Gastrointest. Liver Physiol. 310, G1138–G1146 (2016).
Bach Knudsen, K. E. et al. Impact of diet-modulated butyrate production on intestinal barrier function and inflammation. Nutrients 10, 1499 (2018).
Wang, X. et al. Bacteroides-derived isovaleric acid enhances mucosal immunity by facilitating intestinal IgA response in broilers. J. Anim. Sci. Biotechnol. 14, 4 (2023).
Blakeney, B. A., Crowe, M. S., Mahavadi, S., Murthy, K. S. & Grider, J. R. Branched short-chain fatty acid isovaleric acid causes colonic smooth muscle relaxation via cAMP/PKA pathway. Dig. Dis. Sci. 64, 1171–1181 (2019).
Martin-Gallausiaux, C., Marinelli, L., Blottière, H. M., Larraufie, P. & Lapaque, N. SCFA: mechanisms and functional importance in the gut. Proc. Nutr. Soc. 80, 37–49 (2021).
Sullivan, S. J., Jacobson, R. M., Dowdle, W. R. & Poland, G. A. 2009 H1N1 influenza. Mayo Clin. Proc. 85, 64–76 (2010).
Allen, J. D. & Ross, T. M. H3N2 influenza viruses in humans: viral mechanisms, evolution, and evaluation. Hum. Vaccin. Immunother. 14, 1840–1847 (2018).
Gambotto, A., Barratt-Boyes, S. M., de Jong, M. D., Neumann, G. & Kawaoka, Y. Human infection with highly pathogenic H5N1 influenza virus. Lancet 371, 1464–1475 (2008).
Wu, X., Xiao, L. & Li, L. Research progress on human infection with avian influenza H7N9. Front. Med. 14, 8–20 (2020).
Sun, Y. & Liu, J. H9N2 influenza virus in China: a cause of concern. Protein Cell 6, 18–25 (2015).
Niu, J. et al. Microbiota-derived acetate enhances host antiviral response via NLRP3. Nat. Commun. 14, 642 (2023).
Si, W. et al. Lactobacillus rhamnosus GG induces cGAS/STING- dependent type I interferon and improves response to immune checkpoint blockade. Gut 71, 521–533 (2022).
Qiu, J. et al. Microbiota-derived acetate is associated with functionally optimal virus-specific CD8(+) T cell responses to influenza virus infection via GPR43-dependent metabolic reprogramming. Gut Microbes 16, 2401649 (2024).
Xie, J. et al. Akkermansia muciniphila protects mice against an emerging tick-borne viral pathogen. Nat. Microbiol. 8, 91–106 (2023).
Fan, X. et al. Prevotella histicola ameliorates DSS-induced colitis by inhibiting IRE1α-JNK pathway of ER stress and NF-κB signaling. Int Immunopharmacol. 135, 112285 (2024).
Gong, J. et al. Effects of Prevotella copri on insulin, gut microbiota and bile acids. Gut Microbes 16, 2340487 (2024).
Larsen, J. M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 151, 363–374 (2017).
Horn, K. J., Schopper, M. A., Drigot, Z. G. & Clark, S. E. Airway Prevotella promote TLR2-dependent neutrophil activation and rapid clearance of Streptococcus pneumoniae from the lung. Nat. Commun. 13, 3321 (2022).
Kang, X. et al. Roseburia intestinalis generated butyrate boosts anti-PD-1 efficacy in colorectal cancer by activating cytotoxic CD8(+) T cells. Gut 72, 2112–2122 (2023).
Haghikia, A. et al. Propionate attenuates atherosclerosis by immune-dependent regulation of intestinal cholesterol metabolism. Eur. Heart J. 43, 518–533 (2022).
Erny, D. et al. Microbiota-derived acetate enables the metabolic fitness of the brain innate immune system during health and disease. Cell Metab. 33, 2260–2276.e2267 (2021).
Miller, K. D. et al. Acetate acts as a metabolic immunomodulator by bolstering T-cell effector function and potentiating antitumor immunity in breast cancer. Nat. Cancer 4, 1491–1507 (2023).
Britz, M. L. & Wilkinson, R. G. Partial purification and characterization of two enzymes involved in isovaleric acid synthesis in Clostridium bifermentans. J. Gen. Microbiol. 129, 3227–3237 (1983).
Thierry, A., Maillard, M. B. & Yvon, M. Conversion of L-leucine to isovaleric acid by Propionibacterium freudenreichii TL 34 and ITGP23. Appl. Environ. Microbiol. 68, 608–615 (2002).
Li, X. et al. Lactobacillus gasseri RW2014 ameliorates hyperlipidemia by modulating bile acid metabolism and gut microbiota composition in rats. Nutrients 14, 4945 (2022).
Liu, Q. et al. Effects of isovalerate supplements on morphology and functional gene expression of rumen mucosa in pre- and post-weaning dairy calves. Animal 12, 491–500 (2018).
Javanian, M. et al. A brief review of influenza virus infection. J. Med Virol. 93, 4638–4646 (2021).
Skog, E. et al. An oseltamivir-resistant avian H1N1 influenza A virus can transmit from mallards to chickens similarly to a wild-type strain: implications for the risk of resistance transmission to humans. J. Gen. Virol. 104, (2023).
Kayani, M. D. Oseltamivir-resistant h1n1 influenza virus: Case report. J. Pak. Med. Assoc. 72, 2112–2113 (2022).
Tran, T. T. et al. Efficient symptomatic treatment and viral load reduction for children with influenza virus infection by nasal-spraying Bacillus spore probiotics. Sci. Rep. 13, 14789 (2023).
Wu, Q. et al. Short-chain fatty acids alleviate vancomycin-caused humoral immunity attenuation in rabies-vaccinated mice by promoting the generation of plasma cells via Akt-mTOR pathway. J. Virol. 97, e0065623 (2023).
Yan, Q. et al. LysoPE mediated by respiratory microorganism Aeromicrobium camelliae alleviates H9N2 challenge in mice. Vet. Res. 55, 136 (2024).
Verbrugghe, P., Van Aken, O., Hållenius, F. & Nilsson, A. Development of a real-time quantitative PCR method for detection and quantification of Prevotella copri. BMC Microbiol. 21, 23 (2021).
Acknowledgements
We acknowledge the members of Jilin Provincial Engineering Research Center of Animal Probiotics, Jilin Provincial Key Laboratory of Animal Microecology and Healthy Breeding, Engineering Research Center of Microecological Vaccines (Drugs) for Major Animal Diseases, Ministry of Education, Jilin Agricultural University, Changchun, 130118, China. We acknowledge Biorender (www.biorender.com) for providing the mapping platform. This work was supported by the National Key R&D Program of China (2023YFD1800301), the National Natural Science Foundation of China (U21A20261, 32202819 and 32072888), the China Agriculture Research System of MOF and MARA (CARS-35), the Science and Technology Development Program of Jilin Province (YDZJ202301ZYTS326, YDZJ202102CXJD029, 20210202102NC, 20230202080NC, 20220202057NC and 20230101346JC).
Author information
Authors and Affiliations
Contributions
Author contributions C.F.W., W.T.Y. and G.L.Y. designed the experiments. J.H.X. wrote the paper. B.S.Z. organized and typeset the images. T.M.N., C.W.S. and T.Y. helped with sample collection and data presentation. S.H.F., D.X.Z. and H.B.H. performed the majority of the experiments and analyzed the data. J.Z.W. and D.Z. helped revise the manuscript. X.C., N.W. and Y.Z. supervised the study. Y.L.J. drafted the original paper. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Xing, J., Niu, T., Yu, T. et al. Gut microbiota-derived isovaleric acid ameliorates influenza virus infection via gut-lung axis. npj Biofilms Microbiomes 11, 116 (2025). https://doi.org/10.1038/s41522-025-00753-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41522-025-00753-3
