
Abstract
Mental disorders are one of the leading causes of global health burden, while food flavors play a significant role in promoting the appetite and mood of people. This study aimed to investigate the intervention effects of two kinds of pleasant food odorants on depressed mice induced by reserpine. After 6-week exposure, beef odorant and milk odorant could effectively reduced pro-inflammatory cytokines (IL-6, TNF-α), restored hippocampal structure, elevated neurotransmitters (5-HT, DA, NE), and upregulated BDNF/GFAP expression. 16S rRNA sequencing analysis revealed that both odorants could ameliorate the gut microbiota dysbiosis, increasing the relative abundance of Bacteroidetes and reducing Firmicutes. Overall, milk and beef odorants showed reletively positive effect on depressed mice through gut-brain axis, where milk odorant was more pronounced. GC-MS analysis identified oleic acid in milk odorant as a potential active component. These findings highlighted food-derived odors as promising nutritional interventions for depression through neuroinflammation modulation and microbiota-gut-brain axis regulation.

Similar content being viewed by others


Introduction
Mental disorders are one of the leading causes of global health burden. According to the 2019 Global Burden of Disease Study, depression and anxiety are the two mental health disorders with the highest disability rates, both ranking among the top 25 global causes of disease burden in 20191. Altogether, major depressive disorder caused 49.4 million DALYs and anxiety disorders caused 44.5 million DALYs globally in 20202. Pharmacological and psychological interventions are commonly employed in the treatment of depression. The most commonly used clinical antidepressants are tricyclic antidepressants, selective serotonin reuptake inhibitors, and monoamine oxidase inhibitors3. However, traditional orthodox antidepressant drugs generally have disadvantages, such as a narrow spectrum, numerous side effects, and high cost, resulting in a non-ideal depression treatment4.
Aromatherapy is a therapeutic modality in which odorants or essential oils are absorbed into the body through inhalation, diffusion, bathing, or massage, to enhance both physical and mental well-being5. Currently, aromatherapy is extensively utilized in the treatment of various conditions, including chronic pain, depression, anxiety, insomnia, cognitive enhancement, stress relief, and other disorders associated with psychological and physiological imbalances6. Moreover, inhalation therapy is regarded as one of the safest methods of drug delivery. Small molecules can enter the blood circulation through the absorption of the nasal respiratory mucosa into the capillaries and the blood circulation through the lamina propria in the olfactory region7. For the moment, aromatherapy is widely recognized and used in clinical practice as an important alternative therapy8,9. Picea mariana Essential Oil with γ-terpene and β-pinene as main components has been shown to be effective in improving reserpine-induced anorexia and weight loss in Kunming mice10. Studies have shown that inhalation of navel orange essential oil can significantly mitigate depressive behaviors induced by chronic unpredictable mild stress11. At present, the majority of research on aromatherapy for depression focuses on essential oils, with relatively limited studies exploring the effects of food odors.
Food flavor is one of the important indexes of food sensory quality and consumption. Recent studies have shown that consumers are increasingly concerned about the potential health impacts of food odorants12. Many countries recommend reducing sodium intake according to the population and improving food flavor with herbs and flavor is a sodium reduction strategy recommended in many national dietary guidelines13. Food and beverages with milk flavor are very popular with consumers, and products with milk flavor can be found everywhere in the market14. Furthermore, milk flavor represents one of the largest categories of food odorants and flavoring agents. Beef flavor is one of the fastest-growing odorants in the 21st century, primarily used to enhance the taste and odorants of meat products, snacks, and baked goods, meeting the demand for richer color, taste and odorants in food15. At present, there are few studies on the health effects of beef and milk odorants. Therefore, this study focuses on two highly accepted food odorants with substantial development potential, investigating their effects on emotional health.
Besides, depression is closely related to intestinal microbiota homeostasis, which mainly refers to the changes in microbial diversity caused by the imbalance of microbiota and corresponding functional changes16. In recent years, with the in-depth study of the regulation of gut microbiota, it has been found that there is a network relationship of bidirectional information regulation pathways between the brain, gastrointestinal tract and microbiota related to depression, namely the “brain-gut-microbe” axis of gut microbiota17. The bidirectional regulation of the “brain-gut-microbiome” axis means that the physiological activities of the intestine can not only be regulated by the intestinal self-nerve based on physiological functions, but also receive mandatory signals from the central nervous system and make various physiological responses18. As well as, various physiological responses generated by changes in the structure of intestinal flora can also be transmitted to the brain. Stimulation to it leads to the occurrence of various physiological and pathological phenomena19. Food flavor may degrade the bacterial cell wall and destroy the protein structure of the cell membrane and then exert antibacterial effect by acting on the cell membrane and entering the cell interior20. Once the permeability of the cell membrane changes, its function will also change.
Therefore, this study aimed to investigate the effects of milk and beef odorants on the reserpine-induced depression model through sniffing. The composition of of milk and beef odorants were detected firstly and the depressive-like behavior, neurotransmitters, pro-inflammatory cytokine levels, tissue morphology, neurotrophic factors, and gut microbiota of mice sniffing to food odorants were determined. This study will give reference and broaden the application of food odorants in food research and human health.
Results
Composition analysis of beef and milk odorants
The volatile compounds in beef and milk odorants were analyzed by GC/MS and the results were shown in Table 1. Seventeen volatile compounds were identified in the beef odorant, with alkanes constituting the predominant class (77.46% of total relative peak area). The key flavor compound was identified as ethyl 1,3-dithiolane-2-carboxylate, representing 21.56% of the relative peak area. Meanwhile, the milk odorant contained ten volatile compounds, with cis-11-eicosenoic acid (50.70%) and oleic acid (39.31%) being the predominant flavor components.
Effects of beef and milk odorants on the bodyweight and food intake of mice
The mice was exposed to different food odors and treated as shown in Fig. 1. The changes on bodyweight and food intake of mice in each group were presented in Fig. 2. During the initial 4 weeks, all groups exhibited continuous weight gain. Subsequent to reserpine injection, significant weight reduction was observed in all groups except the BC group. Notably, the odorant-exposed groups (B: beef odorant; E: milk odorant) demonstrated slower rates of weight loss compared to the MC group, suggesting both odorants might alleviate the weight loss of depressed mice. Concomitant with bodyweight changes, reserpine injection induced significant appetite suppression in the groups, with the exception of BC group (Fig. 2B). The MC group exhibited the most rapid decline in food intake, whereas odorant-exposed groups maintained relatively higher feeding rates. It showed that reserpine successfully induced anorexia as a depression-associated phenotype, while chronic exposure to beef and milk odorants might significantly attenuate this hypophagic response.

Experimental schedule of animal treatments.

A Body weight. B Food intake. Data are presented as means ± SEM (n = 8). The different letters represent significant differences between different groups, p < 0.05.
Effect of beef and milk odorants exposure on the depression-like behavior of mice
As illustrated in Fig. 3A and B, the open field test (OFT) revealed the movement distance of mice in MC group was significantly reduced and their activity duration was slightly shorter, compared with those in BC group (p < 0.05). It’s confirmed that reserpine successfully induced depression-associated motor retardation. Notably, odorant intervention might partially reverse these effects. Compared to MC group, milk odorant-exposed mice (E group) demonstrated a significant prolongation of activity duration (p < 0.05), while beef odorant-exposed mice (B group) showed a slightly increase. Both odorant-treated groups exhibited significant recovery in locomotion distance (p < 0.05). These data suggested chronic exposure to milk odorant could significantly ameliorate the depression-associated motor retardation induced by reserpine, with beef odorant showing a non-significant trend toward improvement.

A Movement time of OFT. B Movement distance of OFT. C Immobility time of TST. D Immobility time of FST. E Sucrose preperence index. Data are presented as means ± SEM (n = 8). The different letters represent significant differences between different groups, p < 0.05.
As shown in Fig. 3C, the tail suspension test TST results revealed that the duration of immobility in MC group induced by reserpine was significantly prolonged compared to BC group (p < 0.05). After exposed to beef and milk odorants, this effect was significantly attenuated (p < 0.05). Notably, the E group (milk odorant) achieved immobility levels statistically indistinguishable from BC group, indicating that milk odorant could restore the survival desire of mice to near-complete normalization.
Meanwhile, the FST results were shown in Fig 3D. There was a significant increase of floating immobility time in the mice of MC group compared with BC group (p < 0.05). It suggested that reserpine significantly reduced the desire for survival of mice under severe conditions. Compared with the MC group, the floating immobility durations of mice in two sniffing groups were significantly reduced (p < 0.05).
In addition, the sucrose preference index of mice in MC group was significantly lower than that in BC group (p < 0.05), indicating that reserpine caused the decrease of mice’s desire to explore sugar water (Fig. 3E). The sucrose preference in the E group was significantly higher than that in model group (p < 0.05). It suggested that exposure to milk odorant could significantly restore the level of sucrose preference in depressed mice, while there was no significant effect by beef odorant. These results showed that a depression model of mice was successfully induced by reserpine. Both milk and beef odorant may improve the depression-like behavior of mice, where milk odorant may work more significantly.
Exposure to beef and milk odorants reduced the levels of inflammatory cytokines in serum
The concentration of serum pro-inflammatory factors were determined and shown in Fig. 4. Compared with the BC group, the levels of IL-6 and TNF-α in MC group mice were significantly increased (p < 0.05), indicating that reserpine could increase the level of proinflammatory factors in depressed mice and then produce inflammatory reaction. By sniffing at two food odorants, the levels of pro-inflammatory factors were decreased in different degrees compared with the MC group. Compared with the model group, the level of IL-6 and TNF-α in the E group was significantly reduced (p < 0.05), which was not significant to that in BC group. These results further indicated that both milk odorant and beef odorant could suppress inflammation in depressed mice, where milk odorant may have a reletively strong effect.

A IL-6. B TNF-α. Data are presented as means ± SEM (n = 8). The different letters represent significant differences between different groups, p < 0.05.
Exposure to beef and milk odorants enhanced the concentrations of key neurotransmitters in the hippocampus of mice
The neurotransmitter levels including 5-hydroxytryptamine (5-HT), norepinephrine (NE) and dopamine (DA) in hippocampus tissue of mice were shown in Fig. 5. Compared with the BC group, the 5-HT and DA levels in MC group were significantly decreased (p < 0.05). When exposure to both odorants (B and E group), the levels of 5-HT and DA in the hippocampus of mice were significantly restored compared with those in the MC group (p < 0.05). However, there’s no significant difference of NE level were observed statistically among BC group, MC group and E group (p > 0.05), while it was significantly increased in the B group. The results further indicated that the milk and beef odorants could effectively restore the decrease of 5-HT and DA content in depressed mice.

A 5-HT. B DA. C NE. Data are presented as means ± SEM (n = 8). The different letters represent significant differences between different groups, p < 0.05.
Exposure to beef and milk odorants ameliorated the structural alterations of hippocampus tissue
According to the hematoxylin-eosin (HE) staining results (Fig. 6), the hippocampal cells in the BC group mice were abundant, dense, orderly and compactly arranged with multiple cell layers. In the MC group induced by reserpine, the hippocampus tissue were observed pronounced damage with decreased number of nerve cells, larger intercellular space, smaller nuclei became, darker color, irregular arrangement, increased vacuoles. Compared with the MC group, the number and volume of nerve cells in the odorant groups were reduced, and nuclear shrinkage was alleviated. The number of sparse and disordered arrangements and incomplete structures was reduced. Among them, milk odorant has a better effect on the improvement of hippocampus structure in depressed mice.

a BC group, b MC group, c E group, d B group (200×). Data are presented as means ± SEM (n = 5). The different letters represent significant differences between different groups, p < 0.05.
Exposure to beef and milk odorants regulated the expression level of BDNF and GFAP in the hippocampus of mice
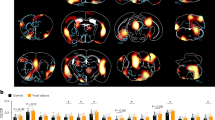
The effects of two odorants on brain neurotrophic factor brain-derived neurotrophic factor (BDNF) in mice were shown in Fig. 7A and B. The immunofluorescence intensity of BDNF in the MC group was significantly lower than in other groups (p < 0.05). It suggested that reserpine induction can reduce the expression of BDNF to hinder the plasticity of nerve cells, promote neuronal atrophy and reduce the number of synapses. When exposure to the two odorants, the expression levels of BDNF were upregulated significantly than the MC group (p < 0.05). There was no significant difference between the two odorants. It further demonstrated that both beef odorant and milk odorant could ameliorate the decline of BDNF expression in depressed mice.

A immunofluorescence image of BDNF (red) (400×). B BDNF immunofluorescence intensity. C Immunofluorescence image of GFAP (red) (400×). D Quantitative analysis of GFAP immunofluorescence intensity. Data are presented as means ± SEM (n = 8). The different letters represent significant differences between different groups, p < 0.05.
In addition, the immunofluorescence analysis on glialcellline-derived neurotrophic factor (GDNF) of astrocytes in the hippocampus of mice were also determined (Fig. 7C, D). The astrocytes in MC group were observed to be enlarged and polysynapse with the expression of GFAP increased compared with the BC group (p < 0.05). It indicated that the astrocytes in depressed model mice were reactive hyperplasia by reserpine. Compared with the model group, the immunofluorescence intensity of GFAP in the B and E groups were decreased significantly (p < 0.05). There’s no significant difference beween the two food odorants. Theses results indicated that beef odorant and milk odorant were effective in restoring the GAFP in depressed mice induced by reserpine.
Exposure to beef and milk odorants ameliorate the gut microbiota of depressed mice
To analyze the effect of two food odorants on the gut microbiota of depressed mice, the V3-V4 hypervariable region of 16S rRNA of microbiota in colon was sequenced and shown in Fig. 8. The rarefaction curve showed that the amount of sequencing data was progressive and reasonable, and the bacterial species of the microbiome reached the saturation stage. It reflect the vast majority of microbial diversity information in the samples. Then the specie numbers of OTUs in each group were shown in Fig. 8B. The common OUTs number of the four groups was 334. The unique OTUs numbers were found remarkably different in each group, including 77 unique OTUs in BC group, 224 in MC group, 64 in E group and 130 in B group, respectively. Among them, the number of OTUs in the MC group was extremely higher than that in BC group, indicating that reserpine induction could change the diversity or composition of gut microbiota. When exposure to milk and beef odorants, the increase of OTUs caused by reserpine was alleviated. Moreover, the LEfSe analysis in Fig. 8C showed that 14 species were significantly different in the four groups under the effect size LAD > 3 threshold which could be potential biomarkers. There were two species significant in B group and E groups, while only one species were significant in MC group. Overall, milk odorant and beef odorant significantly modulated the composition of intestinal microbiota in depressed mice.

A Rarefaction curves. B OTU cluster analysis and species annotations. C LEfSe analysis. D The relative abundances of the top 10 at the phylum level. E The relative abundances of the top 10 at the genus level. Data are presented as means ± SEM (n = 5). The different letters represent significant differences between different groups, p < 0.05.
Furthermore, the distribution of microbial communities in the four groups of mice was shown in Fig. 8D, E. At the phylum level, Bacteroidetes, Firmicutes and Proteobacteria bacteroidetes took the highest proportion over 90% of the microbial compositions in all samples. The rate of Firmicutes/Bacteroidetes were 0.55 in BC group, which was increased to 1.01 in MC group and then decreased to 0.65 in E group. It indicated that reserpine could decrease the relative abundance of Bacteroidetes and increase the relative abundance of Firmicutes, leading to the imbalance of bacterial community in depressed mice. When exposure to milk odorant, this trend can be changed and restored to normal direction as control group.
In addition, the microbial composition of the colon contents was analyzed at the genus level. Three genera were observed to be different significantly between the BC group and the MC group. The relative abundance of Akkermansia and Alloprevotella in MC group was higher than that in BC group, while the relative abundance of Lachnospiraceae_NK4A136_group in the MC group was reduced. It revealed that reserpine could upregulate the relative abundance of Akkermansia and Alloprevotella and downregulated the Lachnospiraceae_NK4A136_group. And these changes could be reversed by exposure of milk odorant (E group). Besides, the beef odorant exposure (B group) significantly increased the relative abundance of Ligilactobacillus and decreased the relative abundance of Akkermansia. In a word, these results suggest that exposure to beef odorant and milk odorant was capable of modulating the dysregulation of the bacterial ecosystem in colon contents caused by reserpine.
Discussion
A total of 17 compounds were identified by GC-MS analysis from beef odorant and 10 compounds were identified from milk odorant. The two odorants have simple flavor components and strong flavor. The ratio of Ethyl 1,3-Dithiolane-2-carboxylate in beef odorant was the highest, and the threshold value of sulfur-containing compounds was small, which contributed the most to beef odorant21. Homolysis of alkoxy groups of fatty acids, the main source of hydrocarbon compounds in beef essence. Long chain alkanes mainly come from the degradation of fatty acids or the fat deposited in the animal’s body by the animal’s feed. Normal alkanes may be derived from the oxidation of fatty acids, and short-chain alkanes are secondary products of the automatic oxidation of fats. Various alkanes (C5–C17) substances, because of their generally high aroma threshold, weak aroma or no odor, do not contribute much directly to the overall flavor22. However, they are important intermediates in the formation of heterocyclic compounds that contribute to the flavor of beef flavor. It can be inferred that these substances have a fundamental role in the formation of beef flavor. Oleic acid in milk odorant has the odor of animal oil or vegetable oil, oleic acid can regulate blood lipid levels23, reduce cholesterol24, effectively reduce the occurrence of hypercholesterolemia and cardiovascular diseases, and reduce the chance of coronary heart disease25. Stearic acid microstrip smell of butter. Fatty acids mainly come from the oxidative degradation of fats, mainly long-chain fatty acids, such as palmitic acid, oleic acid, etc. These esters have a milky smell.
The cause of depression is still unknown, but modern medicine has proposed various hypotheses. Among them, the monoamine neurotransmitter hypothesis, hypothalamic-pituitary-adrenal axis hypothesis, cytokine hypothesis and brain-derived neurotrophic factor hypothesis have been widely recognized26. Animal models on depression include drug and chronic stress models. The drug-induced model is easy to administer, has a short duration, and effectively replicates the onset of depression in humans. It is currently widely used27.
Reserpine is a substance widely used in depression drug models. Reserpine interferes with the reuptake of neurotransmitters into presynaptic vesicles, leading to their degradation by monoamine oxidase, which results in the depletion of neurotransmitters and the onset of depressive symptoms28. Currently, there is no unified standard dose of reserpine29. The dose of reserpine for inducing chronic and acute depression in rats is between 0.1‒0.5 mg/kg and 1–5 mg/kg, respectively30. Shehata A. et al.31 developed a depression model by injecting 1.0 mg/kg reserpine solution into rats intraperitoneally daily. Antkiewicz-Michaluk L. et al.32 developed a depression model by injecting 0.2 mg/kg reserpine solution into the abdominal cavity of rats for 14 consecutive days. All of these injections can induce depression, but long-term injections at low doses are more closely related to chronic depression in clinical practice. Thus, in this study, 0.4 mg/kg reserpine solution was injected intraperitoneally for 14 days to induce depression in mice.
In the research of depression, behavioral experiments can be used as basic evaluation indicators, mainly including OFT, TST, FST and SPT. OFT is widely used to assess anxiety-like behaviors in rodents, where animals are allowed to explore an open arena surrounded by walls. Typically, rodents tend to avoid the center of the arena and remain in the periphery, a behavior known as thigmotaxis. An increase in central activity suggests an anxiolytic effect, while a decrease indicates anxiety-like behavior33. TST and FST are well-established methods for evaluating antidepressant effects and have been extensively used in neuropharmacological research and the preliminary screening of antidepressants34. SPT is the best way to detect reward activity in rodents and can be used as an important indicator to evaluate animal models of depression35. In this study, reserpine-treated mice exhibited prominent depression-like behaviors, such as increased immobility during the forced swimming test, reduced sucrose preference, and prolonged tail suspension time, confirming the successful establishment of the depression model. Preliminary results showed that milk odorant and beef odorant can improve depression behavior. Based on this, we further explored several potential mechanisms underlying the antidepressant effects of odorants in reserpine-treated depressed mice.
Monoamine neurotransmitters, such as 5-HT, DA, and NE, play an important regulatory role in the development of mood disorders. Brain-derived neurotrophic factor (BDNF) is essential for promoting neuronal survival, growth, differentiation, and development, and is involved in the structural and functional plasticity of neurons. The synaptic plasticity of BDNF can affect the release and transmission of DA, 5-HT and adrenergic neurons, thus affecting depression36. Increased BDNF levels indicate the repair of nerve function and reflect the degree of nerve damage and prognosis. Clinical study found that patients with cerebrovascular accident of neural function damage can use BDNF levels to reflect and relate to the prognosis37. Loss of BDNF in the hippocampus can induce neuronal apoptosis, eventually leading to depression. In this study, the contents of 5-HT, DA and NE in the hippocampus and the expression of BDNF in the hippocampus increased after sniffing the two food odorants, indicating that the two food odorants can further improve depressive symptoms by regulating the expression of neurotrophic factors and the level of monoamine neurotransmitters.
Neuroinflammation is considered one of the key contributors to the onset and progression of depression. Excessive secretion of pro-inflammatory cytokines, especially IL-1β, IL-6 and TNF-α, may lead to neuroinflammation and brain damage, leading to depressed mood, anxiety, and impaired memory and attention, which can lead to depression38. A study showed that anti-inflammatory cytokines may exert antidepressant effects through various mechanisms39. For example, Apelin-13 can modulate the shift of microglia from pro-inflammatory to anti-inflammatory cytokine production, inhibit the release of pro-inflammatory cytokines, and promote the synthesis of anti-inflammatory cytokines, thereby alleviating depression-like behavior in mice. Depression has recently been considered a neuroinflammatory disease, with inflammatory changes in the cerebral cortex and hippocampus, in which inflammatory cytokines released by the activation of microglial cells play a key role in its pathogenesis40. Proinflammatory cytokines such as IL-1β, IL-6, and TNF-α promote inflammation by activating multiple types of immune cells, and overactive proinflammatory cytokines in the brain can interfere with many neuronal functions, thereby affecting emotional expression41. Microglia are immune cells in the central nervous system and mediators of inflammation. GFAP, one of the main components of astrocytes, is often used to specifically label astrocytes. Under pathological conditions, astrocytes can be rapidly activated, resulting in a dense distribution, enlargement of cell bodies, and numerous thick processes42. Overactivated astrocytes secrete various inflammatory mediators that can induce depression-like behavior. The results in present study indicated that the both milk odorant and beef odorant could ameliorate reserpine-induced depression of mice by inhibiting GFAP expression and reducing proinflammatory factor levels of IL-6 and TNF-α.
As the largest and most complex microecology in human body, intestinal microecology plays an important role in the occurrence and development of many diseases43. The diversity of gut microbiome in depressed patients is different from that in normal people44. In the process of human evolution, intestinal flora is interdependent and mutually restricted with the body to form a stable overall internal environment. Changes in homeostasis can lead to the overrelease of bacterial substrates, fermentation products, and gut hormones, which affect hormones, immunity, and the vagus nerve in the gut-brain pathway45. This bidirectional regulatory process between the brain and gut involves a number of single mechanisms proposed before. Using 16S rRNA gene sequencing, the community diversity and abundance of the gut microbiome were analyzed in this study.
GC/MS analysis identified two bioactive components in the milk odorant, oleic acid and cis-13-eicosenoic acid, both demonstrating gut microbiota-modulating potential. Previous research has shown that oleic acid-derived compounds reduced the Firmicutes/Bacteroidetes ratio, a microbial signature associated with metabolic dysregulation46. Mechanistically, long-chain monounsaturated fatty acids (carbon chain lengths ≥18) altered bile acid metabolism and improved cardiovascular risks of mice, by inhibiting Firmicutes proliferation while promoting Bacteroidetes growth47. As a polyunsaturated fatty acid (PUFA), cis-13-eicosenoic acid shared structural homology with immunomodulatory PUFAs that reverse gut dysbiosis in respiratory disease models, which could reverse gut microbial imbalance in asthmatic mice and reduce the relative abundance of Akkermansia48. In this study, chronic exposure to milk odorant significantly reduced the Firmicutes/Bacteroidetes ratio and the relative abundance of Akkermansia. It was suggested that milk odorant might modulate gut microbiota to alleviate depressive symptoms (Fig. 3), potentially mediated by its high oleic acid content and cis-13-eicosenoic acid.
To further underly the mechanism of the two food odorants on depressed mice, Spearman’s analysis for the above results were performed (Fig. 9). The relatively abundance of Bacteroides is positively correlated with immobility time in TST, floating immobility time in SFT and the expression of GAFP, which indicating that the increase of Bacteroides may contribute to the development of depressive symptoms. Besides, the concentrations of DA and 5-HT were positively correlated with Candidatus_Saccharimonas, Lachnospiraceae_NK4A136_group and Roseburia. BDNF expression levels were significantly positively correlated with Muribaculum, suggesting that a reduction in these bacteria may contribute to the development of depressive symptoms. Mucispirillum, a member of the phylum Deferribacteres, can cross the mucosal barrier, disrupt intestinal wall integrity, and induce inflammation, leading to colitis49,50. The results of these studies further suggest that Lachnospiraceae and Muribaculum play an important role in the development of depression through a mechanism involving damage to the intestinal barrier. The composition of the two odorants is mostly small molecule substances, which might be absorbed through the skin, meridian or blood and then spread to the whole body through the circulation of blood. It was inferred that both beef oaorant and milk odorant might regulate the brain function and behavior by regulating the gut microbiome through gut-brain axis to ameliorate reserpine-induced depression mice. However, the specific mechanism of food odorant on gut microbiota were needed to be claried in future research.

Red indicates a positive correlation, blue indicates a negative correlation, and the depth of the color indicates the strength of the correlation. *means significance p < 0.05, **means significance p < 0.01.
This study provided preliminary evidences supporting the neuroprotective potential of beef and milk odorants in a reserpine-induced depression model. The observed behavioral improvements appear mediated through multi-level modulation of the gut-brain axis, including neurochemical regulation (partial restoration of monoamine neurotransmitters), neuroplasticity enhancement (upregulation of BDNF expression and astrocyte activation of GFAP in hippocampal regions), downregulating of pro-inflammatory cytokines and microbial remodeling by correction of dysbiosis indices of Firmicutes/Bacteroidetes ratio and Akkermansia. The coordinated effects on neurotransmitter, neurotrophic, inflammatory, and microbial domains suggested a systems-level intervention mechanism. Notably, milk odorant exhibited superior efficacy than beef odorant, potentially attributable to its higher PUFAs content. However, several limitations were needed to be considered, such as model specificity, expsure dose and olfactory absorption mechanisms and so on. Future studies should validate these effects in transgenic models and explore clinical applications through controlled inhalation trials. This work established a foundational framework for developing odorant-based neuromodulatory strategies in nutritional psychiatry.
Methods
Materials and reagents
Injectable reserpine (1 mg/1 mL) was purchsed from Tianjin Jinyao Pharmaceutical Co., Ltd (Tianjin, China). Jojoba oil was purchased from Xinsen Natural Vegetable Oil Co. Ltd (Jiangxi province, China). Milk essence and beef essence were purchased from Qingdao Food Banquet Big Kitchen Food Co., Ltd. (Shandong province, China). All the essences were of food grade. The ELISA kits for 5-hydroxytryptamine (5-HT), norepinephrine (NE), dopamine (DA), Interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) were phurchaed from Shanghai ELISA Biotechnology Co., Ltd. (Shanghai, China). Brain-derived neurotrophic factor (BDNF) antibody and glial fibrillary acidic protein (GFAP) antibody were purchased from Wuhan Elabscience Biotechnology Co., Ltd.(Hubei, China).
GC-MS analysis
The volatile compounds in beef odorant and milk odorant were analyzed by gas chromatography-mass spectrometry (GC-MS), employing with solid-phase microextraction (SPME) for the extraction of volatile flavor components51. Specifically, 6 mL sample was placed in a 20 mL headspace bottle and 10 μL of 2-methyl-3-heptanone was added as an internal standard. The mixture was thoroughly homogenized and thermally equilibrated at 60 °C for 25 min using a magnetic stirrer. Then a 75 μm carboxen/polydimethylsiloxane (CAR/PDMS)-coated SPME fiber was inserted and exposed to the headspace phase for analyte adsorption at 60 °C. After adsorption for 45 min, the fiber was immediately inserted into the GC-MS injection port for thermal desorption at 250 °C for 15 min to ensure complete release. The GC-MS instrument (Thermo Scientific ISQ 7000 GC/MS, USA) was equiped with an Rtx-5MS column (30 m × 0.25 mm × 0.25 μm). Helium (purity >99.999%) was used as carrier gas at a flow rate of 2 mL/min. GC oven temperature was maintained at 5 °C for 1 min, increased to 180 °C at 3 °C/ min for 1 min, and then increased to 250 °C at 8 °C/min for 3 min. For MS detection, the ion source temperature was maintained at 200 °C, the injector temperature at 220 °C, and the desorption unit at 250 °C. Mass spectra were acquired in full-scan mode across a mass-to-charge (m/z) range of 30.0–500.0 at a scan speed of 1000 amu/s. The volatile compounds were identified based on linear retention Index (LRI), genuine criteria, and mass spectrometry matching the National Institute of Standards and Technology (NIST) library. Only the compounds with a matching degree over 85% were selected. Besides, the compounds were quantified based on the peak area and normalized by the concentration of internal standard.
Animals and experimental design
Thirty-two 6-week-old SPF ICR mice (weighing 18–22 g) were purchased from Beijing Spafu Laboratory Animal Technology Co., Ltd. (Beijing, China) (License No. SCXK 2015-0015). The mice were placed in a 10 cm × 10 cm × 20 cm EVC cage using wood shavings as bedding and maintained under standard experimental conditions (temperature, 22–25 °C; humidity 55 ± 5%, 12 h lighting /12 h darkness and noiselessness).
After adaptation for one week, the mice were randomly divided into 4 groups (8 mice of each group) with exposure to different odorants and treatment: blank control group (BC), model group (MC), beef-odorant group (B) and milk-odorant group (E). The animal experiment was designed as previous study with modifications51. Briefly, the milk essence and beef essence were respectively dilluted by jojoba oil with a dilution of 103 and dropped onto a sterilized cotton wool. Then these cotton wools were respectively filled in a sealed hollow stainless box and placed into the EVC cages of B group and E group. Meanwhile, jojoba oil was used as control odorant exposed to the mice of BC and MC group. The mice were allowed to sniff the odorants for 8 h per day for a continuous period of 6 weeks. To ensure the continuous odor and less microbial contamination, the cotton wool absorbed with essence were replaced every day. In the last 2 weeks, the mice in MC group, B group and E group were injected intraperitoneally with reserpine solution to establish a depression model at the dose of 0.4 mL/kg mice bodyweight every day, while the mice of BC group were injected with an equal dose of normal saline (Fig. 1). During the experimental period, all mice were free to drink and eat with regular food. The bodyweight and food intake of mice were recorded weekly. After behavioral evaluation, one mice was humanely euthanized by neck broken with no pain in 5 s and the other mouse was euthanized in turn. None of the mice were allowed to observe the euthanize process. Blood samples were collected by orbital puncture and centrifuged at 3000 r/min for 15 min at 4 °C. The serum was collected and stored at −80 °C for further analysis.
In addition, the brain tissue, hippocampus tissue and colonic contents of mice were collected, where part of them was fixed in 4% paraformaldehyde and the others were imediately frozen by liquid nitrogen and stored at −80 °C. All animal procedures were carried out in accordance with the Guidelines for the Care and Use of Laboratory Animals of Tianjin University of Science and Technology and the Institute of Radiation Medicine. The animal study was approved by the Animal Ethics Committee of Institute of Radiation Medicine [IRM-DWLL-201228].
Open field behavioral test (OFT)
The depressed-like behaviors and locomotor activities were assessed using a standardized open field experimental instrument (625 mm × 740 mm × 510 mm) constructed with matte-white polyvinyl chloride. During testing, individual mice were gently placed in the central quadrant of sanitized arena (disinfected with 70% ethanol between trials) under uniform illumination (300 lux at arena center). The movement time and distance of mice were recorded for 5 min using a camera in a quiet and light-free environment52.
Tail suspension test (TST)
Depressive-like behaviors of mice were quantified using an automated tail suspension system under standardized laboratory conditions. The mouse tail (2 cm away from tip) was wrapped with medical adhesive tape (approximately 2 cm width × 8 cm length) and hung upside down to a horizontal metal hook with controlled tension of head away from the ground. After a 2-min habituation period to eliminate acute stress artifacts, the behavior of mice was recorded by a digital video tracking for 4 min. The total duration of mice immobility was automatically calculated. An immobile state means that the animal gives up active struggle and its body is in a state of overhanging and not twisting53.
Forced swimming test (FST)
Forced swimming test were conducted as described by Khan Muhammad Imran et al.54 with slight modifications. The mice were placed in a cylindrical glass container (20 cm height, 10 cm diameter) filled with water at 20–25 °C. Before the day of formal test, the mice were acclimated for 15 min to induce behavioral adaptation. During the test, first 2 min was excluded for habituation. Then the total immobile swimming time of mice during the next 4 min was recorded. Mice were considered immobile when they floated on the water surface or moved only minimally to maintain breathing.
Sucrose preference test (SPT)
sucrose preference test test were performed by the literature with some improvements55. Before the formal test, the mice were acclimated two bottles of 1% sucrose solution. After drinking for 24 h, one bottle of sucrose solution was replaced with pure water and the mice continued to drink for another 24 h. After fasting and water restriction for 24 h, the mice were subjected to a sucrose preference test. Briefly, 200 mL of 1% sucrose water and 200 mL of pure water were weighed in two bottles and placed in mice cage. In the test, the positions of the bottles were changed at intervals during the test. After free water and food intake for 12 h, the bottles were removed and weighed. The sucrose preference index were caculated as equation (1).
Determination of neurotransmitters in the hippocampus of mice
The hippocampus of mice was homogenated with saline at a ratio of 1:10 (w/v) at 4 °C for 10 min and centrifugation at 3500 r/min for 20 min. The supernatant were collected for determination. The concentrations of 5-hydroxytryptamine (5-HT), norepinephrine (NE) and dopamine (DA) in the hippocampus tissue of mice were quantitatively measured by enzyme-linked immunosorbent assay (ELISA) kit strictly in accordance with the manufacturer’s instructions (Shanghai ELISA Biotechnology Co., Ltd).
Determination of inflammatory cytokines in the serum of mice
The levels of interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) in mice serum were detected by ELISA kits (Shanghai ELISA Biotechnology Co., Ltd). According to the manufacture’s protocols, the serum samples and standards were placed in the 96-well microtiter plates. After four-step incubations with antibodies and substrates and washing procedures, the reaction was terminated with termination solution. Then the absorbance values were determined to evaluate the concentration of cytokines by Multiskan FC (Thermo scientific).
Histopathological analysis on the hippocampus tissue of mice
The hippocampus tissue of mice was observed by hematoxylin-eosin (HE) staining. After cut into 5 mm thickness, the tissues were immediately fixed in 10% formalin neutral fixative for 30 min and incubated overnight with 0.1 M PBS at 4 °C. Then the tissues were dehydrated with gradient concentration ethanol: 95% ethanol twice each for 5 min → 80% ethanol for 5 min → 70% ethanol for 5 min → 100% xylene twice each for 5 min. Afterwards the hippocampus tissues were stained with hematoxylin and eosin, and gradient eluted by anhydrous ethanol again, followed by 100% xylene twice for 5 min. Finally, the specimen was sealed by neutral resin and the histology of thymus and spleen was observed under light microscope (Axio Lab. A1 pol, Germany).
Immunofluorescence analysis on the neurotrophic factor
The levels of brain-derived neurotrophic factor (BDNF) and glialcellline-derived neurotrophic factor (GDNF) secreted by hippocampal neurons were determined by immunofluorescence analysis. The brain tissue was fixed in 4% paraformaldehyde solution for 24 h, and the sample was dehydrated by routine continuous treatment with graded alcohol and xylene. The slices were dried and incubated overnight at 4 °C with rabbit anti-mouse antibody of GDNF and BDNF (Elabscience Biotechnology Co., Ltd.). Goat anti-rabbit antibody (1:300, dilution) was used as a secondary antibody and then dropped on the slice at 4 °C for 1 h. The tissue sections were then washed 3 times with phosphate-buffered saline (PBS). 4’ 6-diamino-2-phenylindole (DAPI) was added into the section and observed under a fluorescence microscope after 1 h.
Gut microbiota analysis
The gut microbiota from colonic contents was analyzed by 16S rDNA high-throughput sequencing based on 16S rDNAV3-V4 region sequence. Briefly, the total bacterial genomic DNA was extracted using the Fast DNA Stool Kit (Novogene, Beijing, China). Then the hypervariable regions (V3–V4) of the bacterial 16S rRNA gene were amplified based on universal primers (338F: ACTCCTACGGGAGGCAGCAG; 806R: GGACTACHVGGGTWTCTAAT). The thermal cycling conditions was: initial denaturation at 95 °C for 3 min, followed by 25–30 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 45 s, with a final extension at 72 °C for 10 min. After purified by magnetic beads and gel electrophoresis, the PCR amplicons were sequenced on an Illumina MiSeq platform (Illumina, San Diego, California, USA) to generate 2 × 300 bp paired-end reads. Afterwards, bioinformatics analysis was performed, where raw reads were demultiplexed and quality-filtered using Trimmomatic. The filtered reads were clustered into operational taxonomic units (OTUs) at 97% similarity using QIIME. Taxonomic classification was conducted against reference databases and the alpha/beta diversity metrics, along with statistical analyses, were computed using R packages such as phyloseq and vegan. Finally, LEfSe analysis of inter group biomarkers was performed with the screening value of linear discriminant analysis (LDA) score setting at 2.
Statistical analysis
All experimental data were repeated at least three times and expressed as mean ± standard error of means (SEM). The statistical analysis was conducted with SPSS statistical software 23 (SPSS Inc., Chicago, IL, USA), while Origin 2019 (OriginLab Corporation, Northampton, MA, USA) were used for data plotting. The data were firstly analyzed through normality and lognormality test. If it fitted the normal distribution, odinary one-way ANOVA test was carried out and Tukey’ test was performed for multiple comparison. If not, Kruskal–Wallis test was performed. p < 0.05 represented the difference between groups were significant at level of 0.05.
Data availability
The authors declare that the data supporting the findings of this study are available within the paper. Any raw data files in another format are available from the corresponding author upon reasonable request.
References
GBD 2019 Diseases and Injuries Collaborators. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet 396, 1204–1222 (2020).
COVID-19 Mental Disorders Collaborators. Global prevalence and burden of depressive and anxiety disorders in 204 countries and territories in 2020 due to the COVID-19 pandemic. Lancet 398, 1700–1712 (2021).
Fang, J. L., Luo, Y., Jin, S. H., Yuan, K. & Guo, Y. Ameliorative effect of anthocyanin on depression mice by increasing monoamine neurotransmitter and up-regulating bdnf expression. J. Funct. Foods 66, 103757 (2020).
Elashmony, S. M. et al. Knowledge, attitude, and practice of saudi medical, nursing, and pharmacy students and interns regarding antidepressant drugs and drug-induced serotonin syndrome. Cureus 15, e51335 (2023).
Perry, N. & Perry, E. Aromatherapy in the management of psychiatric disorders: clinical and neuropharmacological perspectives. CNS Drugs 20, 257–280 (2006).
Jafari-Koulaee, A., Elyasi, F., Taraghi, Z., Sadat Ilali, E. & Moosazadeh, M. A systematic review of the effects of aromatherapy with Lavender essential oil on depression. Cent. Asian J. Glob. Health 9, e442 (2020).
Agatonovic-Kustrin, S., Chan, C. K. Y., Gegechkori, V. & Morton, D. W. Models for skin and brain penetration of major components from essential oils used in aromatherapy for dementia patients. J. Biomolecular Struct. Dyn. 38, 2402–2411 (2020).
Fang, C. S., Tu, Y. K., Chou, F. H., Fang, C. J. & Chang, S. L. Effect of inhaled aromatherapy on sleep quality in critically ill patients: a systematic review and network meta-analysis. J. Clin. Nurs. 34, 1000–1012 (2025).
Zhang, Y. et al. Natural volatile oils derived from herbal medicines: a promising therapy way for treating depressive disorder. Pharmacol. Res. 164, 105376 (2021).
Wang, Y. et al. Antidepressant effect and mechanism of Picea mariana essential oil on reserpine-induced depression model mice. J. Microbiol. Biotechnol. 34, 1778–1788 (2024).
Zhang, L. L. et al. Antidepressant-like effect of Citrus sinensis (L.) osbeck essential oil and its main component limonene on mice. J. Agri. Food Chem. 67, 13817–13828 (2019).
Ayseli, M. T. & Ipek Ayseli, Y. Flavors of the future: health benefits of flavor precursors and volatile compounds in plant foods. Trends Food Sci. Technol. 48, 69–77 (2016).
Petersen, K. S. et al. Using herbs/spices to enhance the flavor of commonly consumed foods reformulated to be lower in overconsumed dietary components is an acceptable strategy and has the potential to lower intake of saturated fat and sodium: a national health and nutrition examination survey analysis and blind tasting. J. Acad. Nutr. Dietetics 124, 14 (2024).
Anastasiou, R. et al. Omics approaches to assess flavor development in cheese. Foods 11, 188 (2022).
O’Quinn, T. G. et al. A current review of U.S. beef flavor II: Managing beef flavor. Meat Sci. 209, 109403 (2024).
Wang, Y. & Kasper, L. H. The role of microbiome in central nervous system disorders. Brain Behav. Immun. 38, 1–12 (2014).
Shen, Y., Yang, X., Li, G., Gao, J. & Liang, Y. The change of gut microbiota in MDD patients under SSRIs treatment. Sci. Rep. 11, 14918 (2021).
Bi, B. et al. The impact and mechanism of arabinoxylan on behavioral regulation in PSD rats via gut microbiota modulation. Int. J. Biol. Life Sci. 7, 48–53 (2024).
Chen, C. Y., Wang, Y. F., Lei, L. & Zhang, Y. Impacts of microbiota and its metabolites through gut-brain axis on pathophysiology of major depressive disorder. Life Sci. 351, 122815 (2024).
Fung, T. K. H., Lau, B. W. M., Ngai, S. P. C. & Tsang, H. W. H. Therapeutic effect and mechanisms of essential oils in mood disorders: interaction between the nervous and respiratory systems. Int. J. Mol. Sci. 22, 4844 (2021).
Zhang, Z., Meng, F., Wang, B. & Cao, Y. Effects of antioxidants on physicochemical properties and odorants in heat processed beef flavor and their antioxidant activity under different storage conditions. Front. Nutr. 9, 966697 (2022).
Sha, K. et al. Changes in lipid oxidation, fatty acid profile and volatile compounds of traditional kazakh dry‐cured beef during processing and storage.J. Food Process. Pres. 41, 13059 (2017).
Fariña, C. A., Lavandera, J. & González, A. M. Effect of conjugated linoleic acids on nutritional status and lipid metabolism in rats fed linoleic‐acid‐deprived diets. Eur. J. Lipid Sci. Technol. 121, 1800362 (2019).
Shao, F. & Ford, D. A. Differential regulation of ABCA1 and macrophage cholesterol efflux by elaidic and oleic acids. Lipids 48, 757–767 (2013).
Nielsen, M. H. et al. Linoleic acid in adipose tissue and the risk of myocardial infarction: a case-cohort study. Eur. J. Nutr. 60, 3639–3646 (2021).
Liu, L. et al. Gut microbiota and its metabolites in depression: from pathogenesis to treatment. EBioMedicine 90, 104527 (2023).
Park, B. K. et al. Antidepressant-like effects of Gyejibokryeong-hwan in a mouse model of reserpine-induced depression. BioMed. Res. Int. 2018, 5845491 (2018).
Strawbridge, R., Javed, R. R., Cave, J., Jauhar, S. & Young, A. H. The effects of reserpine on depression: a systematic review. J. Psychopharmacol. 37, 248–260 (2023).
Farzin, D. & Mansouri, N. Antidepressant-like effect of harmane and other beta-carbolines in the mouse forced swim test. Eur. Neuropsychopharmacol. 16, 324–328 (2006).
Ikram, H. & Haleem, D. J. Repeated treatment with a low dose of reserpine as a progressive model of Parkinson’s dementia. Pak. J. Pharm. Sci. 32, 555–562 (2019).
Shehata, A. M., Aziz, A., Diab, A., Elmenofi, G. A. & Marwa, A. A. Regional brain amino acids imbalances in reserpine rat model of depression: an antidepressive effect of pomegranate juice. J. Am. Sci. 11, 18–26 (2015).
Antkiewicz-Michaluk, L., Wąsik, A., Możdżeń, E., Romańska, I. & Michaluk, J. Antidepressant-like effect of tetrahydroisoquinoline amines in the animal model of depressive disorder induced by repeated administration of a low dose of reserpine: behavioral and neurochemical studies in the rat. Neurotox. Res. 26, 85–98 (2014).
Kuniishi, H. et al. Early deprivation increases high-leaning behavior, a novel anxiety-like behavior, in the open field test in rats. Neurosci. Res. 123, 27–35 (2017).
Shewale, P. B., Patil, R. A. & Hiray, Y. A. Antidepressant-like activity of anthocyanidins from Hibiscus rosa-sinensis flowers in tail suspension test and forced swim test. Indian J. Pharmacol. 44, 454–457 (2012).
Liang, M. et al. SuHeXiang essential oil inhalation produces antidepressant- and anxiolytic-like effects in adult mice. Biol. Pharm. Bull. 41, 1040–1048 (2018).
Fahmy, H. M. et al. Thymoquinone-encapsulated chitosan nanoparticles coated with polysorbate 80 as a novel treatment agent in a reserpine-induced depression animal model. Physiol. Behav. 222, 112934 (2020).
Lasek-Bal, A. et al. Low concentration of BDNF in the acute phase of ischemic stroke as a factor in poor prognosis in terms of functional status of patients. Med. Sci. Monit. 21, 3900–3905 (2015).
Khadrawy, Y. A., Sawie, H. G., Hosny, E. N. & Mourad, H. H. Assessment of the antidepressant effect of caffeine using rat model of depression induced by reserpine. Bull. Natl Res. Cent. 42, 1–9 (2018). 2018.
Zhou, S., Chen, S., Xie, W., Guo, X. & Zhao, J. Microglia polarization of hippocampus is involved in the mechanism of Apelin-13 ameliorating chronic water immersion restraint stress-induced depression-like behavior in rats. Neuropeptides 81, 102006 (2020).
Gong, W. et al. Involvement of the microglial NLRP3 inflammasome in the anti-inflammatory effect of the antidepressant clomipramine. J. Affect. Disord. 254, 15–25 (2019).
Hu, X. et al. The novel and potent anti-depressive action of triptolide and its influences on hippocampal neuroinflammation in a rat model of depression comorbidity of chronic pain. Brain Behav. Immun. 64, 180–194 (2017).
Shu, X. et al. The effect of fluoxetine on astrocyte autophagy flux and injured mitochondria clearance in a mouse model of depression. Cell Death Dis. 10, 577 (2019).
Wang, Y. et al. Research on mechanism of charred hawthorn on digestive through modulating “brain-gut” axis and gut flora. J. Ethnopharmacol. 245, 112166 (2019).
Fung, T. C., Olson, C. A. & Hsiao, E. Y. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 20, 145–155 (2017).
Simpson, C. A. et al. Bugs and brains, the gut and mental health study: a mixed-methods study investigating microbiota composition and function in anxiety, depression and irritable bowel syndrome. BMJ Open, 11, e043221 (2021).
Mujico, J. R., Baccan, G. C., Gheorghe, A., Díaz, L. E. & Marcos, A. Changes in gut microbiota due to supplemented fatty acids in diet-induced obese mice. Br. J. Nutr. 110, 711–720 (2013).
Tsutsumi, R. et al. Long-chain monounsaturated fatty acids improve endothelial function with altering microbial flora. Transl. Res. J. Lab. Clin. Med. 237, 16–30 (2021).
Jiang, T. et al. Long-chain polyunsaturated fatty acids improve airway pathological features and gut microbial imbalances in balb/c mice with ovalbumin-induced asthma. J. Funct. Foods 81, 104465 (2021).
Umu, Ö. C. O., Mydland, L. T., Øverland, M., Press, C. M. & Sørum, H. Rapeseed-based diet modulates the imputed functions of gut microbiome in growing-finishing pigs. Sci. Rep. 10, 9372 (2020).
Zhai, X., Lin, D., Zhao, Y., Li, W. & Yang, X. Effects of dietary fiber supplementation on fatty acid metabolism and intestinal microbiota diversity in C57BL/6J mice fed with a high-fat diet. J. Agric. Food Chem., 66, 12706–12718 (2018).
Wang, Y. et al. Peanut oil odor enhances the immunomodulatory effect on immunosuppressed mice by regulating the cAMP signaling pathway via the brain-spleen axis. Food Funct. 15, 1994–2007 (2024).
Abbasi-Maleki, S. & Gholamreza Maleki, S. Antidepressant-like effects of foeniculum vulgare essential oil and potential involvement of dopaminergic and serotonergic systems on mice in the forced swim test. Pharma Nutr. 15, 100241 (2021).
Cryan, J. F., Mombereau, C. & Vassout, A. The tail suspension test as a model for assessing antidepressant activity: review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 29, 571–625 (2005).
Khan, M. I. et al. Antidepressant-like effect of ethanol in mice forced swimming test is mediated via inhibition of NMDA/nitric oxide/cGMP signaling pathway. Alcohol 92, 53–63 (2021).
Boyko, M. et al. Establishment of an animal model of depression contagion. Behav. Brain Res. 281, 358–363 (2015).
Acknowledgements
This work was supported by the Tianjin Innovation Platform Project (24PTLYHZ00020).
Author information
Authors and Affiliations
Contributions
Y. Jin and H. Wang wrote the main manuscript text and prepared all figures. Z. Zhang and E. Jin did the formal analysis. C. Yang did the investigation and data curation. J. Meng and T. Wu provided guidance on the methodology. Y. Jin and M. Zhang supposed the idea and supervision. T. Wu and M. Zhang supported the funding acquisition. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Jin, Y., Wang, H., Zhang, Z. et al. Chronic exposure to milk odorant might ameliorate the depressive-like behavior of mice through gut-brain axis. npj Sci Food 9, 108 (2025). https://doi.org/10.1038/s41538-025-00455-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41538-025-00455-2
