
Abstract
Particular attention has been paid to the nutritional potential of edible insects as well as the health benefits associated with their bioactive compounds. This paper focused on an in-depth review compiling the most recent information on health benefits of insect bioactive metabolites as well as their purification and identification, in addition to consumer attitudes towards edible insects. It was found that, insect bioactive metabolites, including marcocarpal, grandinol, trolline, pancratistatin, narciclasin, ungeremin, cantharidin, cordycepin, roseoflavin, lecithin, reblastatin, chitin, chitosan and desmosterol deemed to have biological activities, such as tumor suppression, anticancer, antihypertensive, anti-inflammatory, antioxidant, immunomodulator, neuroprotective, glycemic and lipid regulation, blood pressure reduction, regulation of intestinal bacterial flora and cardiovascular protection among others. Furthermore, proper sample preparation and extraction is the first step in the purification of bioactive metabolites from edible insects. After concentration, bioactive metabolites are purified using chromatographic and separation techniques including High-Performance Liquid Chromatography (HPLC), Gas Chromatography (GC), Thin-Layer Chromatography (TLC), Size-Exclusion Chromatography (SEC). Finally, their nutritional potential, health benefits, environmentally friendly, great taste, traditions, taboo, safety concerns, unpleasant past experiences, allergies, and unnaturalness are among the main factors influencing attitudes towards insect consumption.
Similar content being viewed by others
Introduction
With a rapidly growing world population1 and the goal of promoting healthier as well as sustainable food systems2, there is a growing demand for alternative proteins3. This situation is exacerbated by the scarcity of essential arable land4, environmental pressures linked to the uncertainties of climate change5,6. Edible insects are thus seen as a formidable alternative to address the issues of global food insecurity7 for their nutritional potential8,9, taste10, economic benefits11,12, environmental benefits13, as well as their potential health benefits14.
In many parts of the world, entomotherapy is used as medicine and is an important alternative to modern therapy through their bioactive metabolites including pancratistatin, narciclasin, ungeremin, cantharidin, cordycepin, roseoflavin, lecithin, reblastatin, chitin, and chitosan15,16. These bioactive compounds present important physiological effects on living organisms through their physiological properties encompassing anti-obesity, antihypertensive, antithrombotic, antioxidant, hypocholesterolemic, antimicrobial, opioid, cytomodulatory, anti-inflammatory, cardioprotective, immunomodulatory, antiangiogenic, and immunomodulatory activities14,17.
Given their diverse functions, high bioavailability and efficacy even at low concentrations, bioactive compounds attract a great deal of attention, although some bioactive compounds are naturally present in isolation, many are hidden within the intact structure18. Even though effort is being made, consumer attitudes and willingness to consume insects remain a major challenge in many societies19, due to traditions, superstitions and taboos as well as familiarity with insect20, their appearance and great taste8,21.
Considering the attention paid to insects as food and feed, this review compiled the most recent information focusing on health benefits of insect bioactive metabolites as well as their purification and identification, and finally a particular attention was paid to sensory attributes and consumer attitudes towards edible insects.
Potential health benefits of insect bioactive metabolites
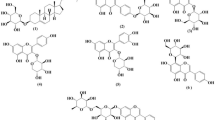
Insects are characterized by several bioactive metabolites, including marcocarpal, grandinol, trolline, pancratistatin, narciclasine, ungeremine, cantharidin, cordycepin, roseoflavin, lecithin, reblastatin, chitin, chitosan and desmosterol (Fig. 1), which confer a variety of beneficial biological activities to human health, including tumor suppression, anti-cancer, anti-hypertensive, anti-inflammatory, antioxidant, immunomodulatory, neuroprotective, blood sugar and lipid regulation, blood pressure reduction, regulation of intestinal bacterial flora and cardiovascular protection (Table 1). While Fig. 2 illustrates the biological activities of bioactive insect metabolites and their mechanisms of activity, detailed information associating insect species to their bioactive metabolites is depicted in Table 1.

Health benefits of insect bioactive metabolites and their potential mechanisms.

Chemical structures of selected bioactive metabolites found in edible insects.
Anti-cancer and tumor suppressive effects have been observed for bioactive metabolites such as actinomycin-D, isocoumarins periplatins A-D, (R)-(+)-palasonin, palasonimide, cantharimide, palasonin, cantharidin, norcantharidin, pederin, pancratistatin, narciclasin and ungeremin found in insect species including Macrotermes natalensis22, Periplaneta americana23, Hycleus oculatus24, Hycleus lunata25, Paederus sp26 and Brachystola magna17 by inducing apoptosis and inhibiting cancer cell growth, in addition to inhibiting tumor metastasis and affecting cancer cell energy metabolism.
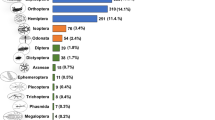
Hypertension is one of the main risk factors for cardiovascular disease, affecting millions of people every year. Angiotensin-converting enzyme (ACE) plays a key role in the regulation of blood pressure, and its efficacy in the treatment of hypertension has been proven27. Protein hydrolysates from insect species belonging to the orders Coleoptera, Diptera, Hymenoptera, Lepidoptera and Orthoptera have demonstrated ACE inhibitory activity. In other research, specific ACE inhibitory peptides from Bombyx mori, Tenebrio molitor, Spodoptera littoralis and Oecophylla smaragdina have been identified27,28. It has been reported that protein synthesis can release certain amino acids with important physiological regulatory functions including inhibiting severe hypertension and lowering blood pressure as a result of combination of lysine and methionine, and histidine29.
Pessina and collaborators30 reported anti-hypertensive effects of defatted Tenebrio molitor exhibiting strong ACE inhibitory activity with a dose-devendent reduction of svstolic blood pressure in hypertensive rats. Additionally, biactive metabolites such as macrotermycin A-D, natalenamides A-C, blapsols A-D, molossusamides A-C, photoinduced, melanins and ommochromes, aspongopusamides A-D, polyrhadopamines A-E, troline found in Macrotermes natalensis31,32, Blaps japanensis17,33, Hermetia illucens34, Polyrhachis dives35 were highlighted to have anti-inflammatory properties through effective inhibition of LOX and COX-2 activity, regulation of expression of inflammation- and immunity-related factors to improve the inflammation pathology. By boosting probiotic production and reducing pro-inflammatory cytokines and plasma lipids, immune response and function in humans, particularly in the gastrointestinal tract, would be linked to insect chitin content.
Chitin is the second most abundant biopolymer found in the exoskeletons of arthropods including insects36. It has some potential antinutritional properties, particularly when consumed in large quantities. The antinutritional aspects of chitin includes digestibility issues, inhibition of nutrient absorption and interference with protein digestion. Moreover, Chitin is largely indigestible to humans due to a lack of the enzymes needed to break it down, as its structure is resistant to digestive enzymes such as amylase, protease and lipase37. Additionally, chitin can bind to certain nutrients, potentially reducing their bioavailability. Furthermore, due to its rigid, fibrous nature, large quantities of chitin can lead to reduced protein absorption or incomplete digestion by interfering with the digestion of proteins and other macromolecules in the stomach38.
Glycosaminoglycan, a polysaccharide found in Gryllus bimaculatus, showed a significant anti-inflammatory effect against chronic arthritis in mice by inhibiting C-reactive protein (CRP) through suppression of a number of inflammatory biomarkers in vitro39. Furthermore, in combination with indomethacin, glycosaminoglycan was more effective than either agent alone in suppressing paw edema39. Furthermore, in rats fed a high-fat diet, glycosaminoglycan reduced CRP levels, abdominal and epididymal fat mass and various serobiochemical parameters (phospholipids, aspartate transaminase (AST), alanine transaminase (ALT), total cholesterol and glucose)40. Another study in diabetic mice revealed that glycosaminoglycan supplementation reduced blood glucose and LDL cholesterol levels, and increased the activity of antioxidant enzymes, notably catalase, superoxide dismutase and glutathione peroxidase41.These findings indicate that the glycosaminoglycan present in Gryllus bimaculatus may help reduce the risk of cardiovascular disease.
The A-D blapsols contained in Blaps japanensis demonstrated antioxidant properties42. Similarly, higher antioxidant activity was found in peptide43, photoinduced, melanins and ommochromes44 by analyzing DPPH and hydroxyl radical scavenging activities. Moreover, carminic acid found in Dactylopius coccus also exhibited antioxidant activity17. Insect hydrolysates and peptide fractions have demonstrated antioxidant properties, by contributing to reduce inflammation and oxidative stress by lowering the level of free radicals present in the body45,46. Di Mattia and collaborators37 reported that water-soluble extracts found in grasshoppers, silkworms and crickets have an antioxidant capacity around five times greater than that of fresh orange juice in vitro, due to their higher protein/peptide content.
Cantharidin17 and defensins-DLP2 and DLP447 found in Hycleus lunata exhibited immunomodulatory effects by promoting the expression of immune-related factors, enhancing natural killer cell activity as well as stimulating and activating the innate immune system. Furthermore, protein-enriched fraction from Musca domestica also showed immunomodulatory effects48. Moreover, immunomodulatory hexapeptide from alcalase hydrolysate of ultramicro-pretreated present in Bombyx mori pupae protein has potential therapeutic value as an immunomodulatory bioactive metabolite49. Purified polypeptide components (BPP-21 and BPP-22) found in Apis mellifera pupae revealed immunomodulatory activity in vivo and in vitro by increasing the phosphorylation of ERK and p38, and modulating the expression of intranuclear transcription factors (EIK-1, MEF-2 and CREB) in the MAPK signaling pathway50.
Pessina and collaborators30 observed neuroprotective effects in defatted Tenebrio molitor larvae. Additionally, bioactive compounds such as Coprismycin A-B and Collismycin A contained in Paederus sp have shown neuroprotective effects26. Moreover, polybioside51 and tetraponerins52 present respectively in Polybia paulista and Tetraponera rufonigra revealed neuroprotective effects.
Anti-microbial effects have been observed in several insect bioactive metabolites, such as Shellolic acid A found in Kerria lacca53, macrocarpal and grandinol found in Amauronematus amplus, Arge sp, Dineura pullior, Nematus brevivalvis, Nematus pravus, Nematus viridescens, Nematus viridis, Perga affinis, Pristiphora alpestris, Trichiosoma scalesii54, Desmosterol, (3β, 5α) cholesta-8, 14, 24-trien-3-ol, 4, 4-dimethyl, (3β, 20 R) cholesta-5, 24-dien-3, 20-diol found in Schistocerca gregaria55.
The antimicrobial effect of Tenebrio molitor and Zophobas morio has been proven in reducing E. coli and Salmonella infections in broilers56 due to their chitin content, which is a polymer of b-1, 4N-acetylglucosamine and is the primary component of the insect exoskeleton57,58. The latter and its degraded products, such as chitosan, exert antimicrobial, antioxidant, anti-inflammatory, anticancer and immunomodulatory activity59. Moreover, Nino et al.60 and Torres-Castillo et al.61 reported potential bioactivity of insect phenolics including tricin, luteolin, apigenin, orientin, iso-orientin, vitexin, iso-vitexin, kaempferol, quercetin, isorhamnetin, myricetin, ferulic acid, sinapic acid, gallic acid, 4-hydroxybenzoic acid, syringic acid, p-coumaric acid, caffeic acid, ferulic acid, sinapic acid, linked to chronic diseases such as antioxidant, anti-inflammatory, and anticancer, among others. Chitooligosaccharides, depolymerized products of chitin and chitosan, taken orally for eight weeks significantly reduced the level of the pro-inflammatory cytokine TNF-α and interleukin (IL)-1β in elderly people62.
Antibacterial activity was observed for actinomycin-D, macrotermycin A-D and pseudoxyallemycin-B present in Macrotermes natalensis22,31,63, 1-(2,5-Dihydroxyphenyl)-3-hydroxybutan-1-one, Roseoflavin and 8-methylamino-8-demethyl-d-riboflavin found in Odontotermes formosanus64,65, roseoflavin, 8-methylamino-8-demethyl-D-riboflavin, natalamycin and termisoflavones A-C present in Macrotermes spp65,66, molossusamides A-C found in Catharsius molossus67, lenzimycins A-B found in Onthophagus lenzii68, α-pyrone, diketopiperazine, pyrone derivatives, diketopiperazine, photoinduced, melanins and ommochromes found in Hermetia illucens69,70, and papilistatin found in Byasa polyeuctes71.
Bioactive metabolites found in insects such as 5-Hydroxyramulosin and biatriosporin-M found in Odontotermes formosanus64, natalamycin-A, geldanamycin, reblastatin, banegasin, cyclo-NMe-L-3,5-dichlorotyrosine-Dhb and rubrominin A-B found in Macrotermes natalensis22, efomycin K, efomycin L, efomycin M, efomycin G, elaiophylin, roseoflavin, 8-methylamino-8-demethyl, D-riboflavin, natalamycin, termisoflavones A-C present in Macrotermes spp65,72, tricin, palmitinic acid and eicosane found in Holotrichia diomphalia73 have shown antifungal effects.
Insect-based feeding is associated with the production of short-chain fatty acids (SCFAs), in terms of increasing the abundance and diversity of beneficial bacteria in the gut. One study showed that chitin is broken down into propionate and butyrate SCFAs by the gut microbiota58, followed by a reduction in blood cholesterol and triglyceride levels in chickens fed insect meal, with an increase in energy58. An increase in white blood cells, haemoglobin and red blood cells, followed by improved immune function was observed in fish supplemented with chitin and chitosan74. A reduction in triglyceride and cholesterol levels and an increase in blood calcium levels were observed in chickens supplemented with H. illucens larvae. This is explained by the fact that chitin’s positive charge enables it to bind negatively charged free fatty acids and bile acids75.
There are a variety of hypoglycemic bioactive metabolites in insects and their products, including proteins, peptides, polysaccharides, unsaturated fatty acids, alkaloids, and flavonoids76. Silkworm hydrolysate and fibroin are said to be ideal blood sugar regulators53. In addition, silkworm larvae, honey and chrysalises contain a large number of polysaccharides with hypoglycemic effects77. Removing the acetyl group, chitin is transformed into soluble chitosan. The oligosaccharides obtained by enzymolysis or acid hydrolysis of chitosan also have hypoglycemic effects in humans78. Insect fat is rich in unsaturated fatty acids79, including linoleic acid, which can improve glucose tolerance, with effects on insulin and reduces the incidence of cardiovascular and retinal complications in diabetic patients80.
Studies on trace elements show that magnesium, zinc, calcium, iron, copper, chromium, nickel, selenium among others are linked to human blood sugar metabolism with hypoglycemic effects81. Additionally, edible insects contain high levels of linolenic acid, which can prevent the synthesis of fatty acids and glycyrrhizin and accelerate the β-oxidation of fatty acids. Linolenic acid functions to reduce triacylglycerides, prolong clotting time and combat thrombosis, and is widely present in lepidopterous larvae, such as Clanis bilineata tsingtauica Mell, Tenebrio molitor, Zophobas atratus82. Moreover, chitin and chitosan present in Tenebrio molitor larvae can reduce blood pressure, blood lipids, and promote cholesterol metabolism78,83.
Moreover, Teixeira et al. 84 reported 177 peptides with predicted bioactivities and 61 peptides with bioactivity assessed In vitro and 3 peptides with bioactivity assessed In vivo from Gryllodes sigillatu, Gryllus assimilis, Schistocerca gregaria, Alphitobius diaperinus, Tenebrio molitor,Polyphylla adspersa, Apis mellifera, Oecophylla smaragdina, Bombyx mori, Spodoptera littoralis, Hermetia illucens, and Musca domestica.
Purification and identification of bioactive metabolites found in edible insects
The purification of bioactive metabolites from edible insects
Several key techniques and methodologies are being used to isolate, identify and purify bioactive metabolites present in the tissues of edible insects. These bioactive metabolites are of growing interest due to their potential health benefits, including antimicrobial, antioxidant, anti-inflammatory and anticancer properties. The general process for purifying bioactive metabolites from edible insects is described below.
Sample preparation and extraction
Proper sample preparation and extraction is the first step in the purification of bioactive metabolites from edible insects. The insect species selected can vary according to the bioactive compounds sought85. Depending on the solubility of the target metabolites, bioactive compounds can be extracted using a variety of solvents, including methanol and ethanol for extracting polar compounds like polyphenols and peptides86, hexane for lipid-soluble compounds such as fatty acids and sterols87, water for hydrophilic bioactive compounds, especially antioxidants88, and acetone is also used for both lipid and protein extractions89. Once extraction is complete, the resulting solution is usually concentrated using techniques such as rotary evaporation to remove the solvent. In addition, filtration is performed to remove insoluble solids, leaving a clear extract ready for further purification.
Purification Techniques
Once extraction is complete, techniques such as rotary evaporation are used to remove the solvent and concentrate the solution. In addition, insoluble solids are removed by filtration, leaving a clear extract ready for further purification. After concentration, bioactive metabolites are purified using chromatographic and separation techniques including High-Performance Liquid Chromatography (HPLC), one of the most common methods for separating and purifying bioactive metabolites from insect extracts90, Gas Chromatography (GC) which is particularly particularly useful for purifying volatile compounds, such as fatty acids and terpenoids91, Thin-Layer Chromatography (TLC), this one can be used as a preliminary purification step for lipophilic compounds such as sterols and antioxidants, even though not as advanced as HPLC; Size-Exclusion Chromatography (SEC): SEC is beneficial technique for separating compounds based on their molecular size. Very useful when purifying large molecules like proteins or polysaccharides from insect exoskeletons92, and Ion-Exchange Chromatography which is a method particularly used for isolating charged compounds, such as bioactive peptides93.
Characterization of purified bioactive metabolites in edible insects
After purification, isolated metabolites are characterized to confirm their identity as well as their bioactivity using several techniques including mass spectrometry (MS): a powerful tool for identifying the molecular weight and structure of bioactive metabolites94, nuclear magnetic resonance (NMR): often used for detailed structural characterization of purified metabolites, particularly to identify complex molecules such as fatty acids and peptides; and UV-Vis spectrophotometry, which is frequently used to identify and quantify light-absorbing bioactive compounds, including polyphenols and flavonoids95.
Identification of bioactive metabolites in edible insects
The identification of bioactive metabolites in edible insects has garnered much attention due their potential health benefits, such as antimicrobial, antioxidant, anti-inflammatory and even anticancer properties, due to their wealth of bioactive compounds, including peptides, lipids, polyphenols, vitamins, minerals and chitin derivatives96. Insects are rich in proteins which can be hydrolyzed to release bioactive peptides with potential health-promoting properties including antimicrobial, antihypertensive by inhibiting angiotensin-converting enzyme (ACE), and antioxidant effects97.
Moreover, edible insects are caracerized by a variety of lipids, including essential fatty acids important for human health. Insects such as crickets, mealworms and grasshoppers contain polyunsaturated fatty acids (PUFAs), notably omega-3 and omega-6 fatty acids98. Furthermore, many edible insects are rich in polyphenolic compounds, particularly phenolic acids and flavonoids, known for their antioxidant in cells and tissues, free radical scavenging activity and potentially anti-cancer properties61.
Additionally, edible insects contain essential vitamins and minerals that support various bodily functions including metabolism, immune function, wound healing, bone health, red blood cell production, and maintaining a healthy nervous system8. Other bioactive metabolites such as sterols and triterpenoids are found in the lipids of insects and are known to contribute cardiovascular health by lowering cholesterol, reduce inflammation, and exhibit anticancer properties99. In addition to chitin, other polysaccharides such as glucans found in the hemolymph of insects have been studied for their potential bioactivity including anticancer and immunomodulatory properties by stimulating the immune system and improving resistance to infections98.
Consumer attitudes toward edible insects
Consumer attitudes toward edible insects have been a subject of interest and debate in recent years19. As the world grapples with the challenges of sustainable food production and environmental concerns, edible insects have emerged as a potential solution to address these issues14. However, the acceptance and adoption of edible insects as a mainstream food source largely depends on consumer attitudes and perceptions21.
One of the primary factors influencing consumer attitudes toward edible insects is cultural and societal norms100. In many Western countries, insects are not traditionally part of the culinary landscape and are often associated with disgust or considered as pests101. This deeply ingrained cultural bias leads to a significant barrier to acceptance. However, in other cultures, such as parts of Asia, Africa, and Latin America, insects have long been consumed and are even considered delicacies102. Cultural exposure and familiarity with edible insects play a crucial role in shaping consumer attitudes and acceptance103.
Many people are concerned about the safety of consuming insects, particularly regarding potential allergenic reactions or contamination104. However, numerous studies have shown that edible insects are safe for human consumption when sourced from reliable and regulated suppliers105. In fact, insects are often rich in protein, vitamins, and minerals, making them a nutritious and sustainable food option9. Education and awareness campaigns highlighting the nutritional benefits and safety standards associated with edible insects can help reshape consumer attitudes.
Traditional livestock production, such as cattle farming, is resource-intensive and contributes to greenhouse gas emissions and deforestation. In contrast, insects require minimal resources, emit fewer greenhouse gases, and can be reared on organic waste, making them an environmentally friendly alternative106. Consumers who are conscious of these environmental issues may be more open to incorporating insects into their diet as a sustainable choice5,107,108.
The way edible insects are marketed and presented to consumers can significantly impact their perception and willingness to try them109. Manufacturers and retailers should focus on creating appealing and visually appealing products that align with consumers’ taste preferences and dietary habits110. Clever marketing strategies that emphasize the novelty, sustainability, and health benefits of edible insects can help overcome initial resistance and spark curiosity among consumers111.
Furthermore, taste preferences are often developed through exposure and personal experiences. Offering opportunities for consumers to sample and taste insect-based products in a non-threatening and controlled environment can help overcome the initial resistance and foster positive experiences110. Social influences, such as peer recommendations and endorsements from influential figures, can also sway consumer attitudes and drive acceptance. Overcoming cultural biases, addressing safety concerns, and raising awareness about the nutritional and environmental benefits of edible insects are crucial steps in reshaping consumer attitudes. By actively engaging consumers, providing appealing product options, and dispelling misconceptions, edible insects have the potential to become a viable and sustainable food source in the future.
It can be concluded that some of the main positive factors influencing attitudes towards insects include nutritional potential, health benefits, environmentally friendly, great taste, and traditions; on the other hand, the main factors underlining negative attitudes towards insects are, among others, taboo, safety concerns, unpleasant past experiences, allergies and unnaturalness as summarized in Fig. 3.

Attitudes towards edible insects as food.
Conclusion and future perspectives
It can be generally concluded that insect bioactive metabolites, including marcocarpal, grandinol, trolline, pancratistatin, narciclasin, ungeremin, cantharidin, cordycepin, roseoflavin, lecithin, reblastatin, chitin, chitosan and desmosterol play a crucial role in conferring several beneficial biological activities, such as tumor suppression, anticancer, antihypertensive, anti-inflammatory, antioxidant, immunomodulator, neuroprotective, glycemic and lipid regulation, blood pressure reduction, regulation of intestinal bacterial flora and cardiovascular protection among others. However, proper sample preparation and extraction is the first step in the purification of bioactive metabolites from edible insects. After concentration, bioactive metabolites are purified using chromatographic and separation techniques including High-Performance Liquid Chromatography (HPLC), Gas Chromatography (GC), Thin-Layer Chromatography (TLC), Size-Exclusion Chromatography (SEC). It is noteworthy that nutritional potential, health benefits, environmentally friendly, great taste, traditions, taboo, safety concerns, unpleasant past experiences, allergies, and unnaturalness are among the main factors influencing attitudes towards insects.
Given the immense insect biodiversity, more in-depth investigations should focus on undiscovered bioactive metabolites, for more information on their potential as a sustainable therapeutic source. Particular attention should be paid to increasingly describing the therapeutic benefits and modes of action of insect bioactive metabolites. Additionally, as many human experiments as possible to explore the biological activities of these bioactive metabolites should also be carried out. Studies focusing on cross-reactivity of edible insects, as well as novelty, smart marketing, and good education can further influence attitudes towards insect consumption.
Data Availability
The datasets generated or analyzed in the current study are available from the corresponding author upon reasonable request.
References
Desa, U. N. World population prospects 2019. Highlights N. Y. U. Nations Dep. Econ. Soc. Aff. 11, 125 (2019).
McClements, D. J. et al. Building a resilient, sustainable, and healthier food supply through innovation and technology. Annu. Rev. Food Sci. Technol. 12, 1–28 (2021).
Godfray, H. C. J. et al. Food security: the challenge of feeding 9 billion people. Sci. 327, 812–818 (2010).
Dickie, F., Miyamoto, M. & Collins, C. M. T. The potential of insect farming to increase food security. Edible Insects 1–10 (2019).
Ishara, J., Ogunyiola, A., Matendo, R., Kiyala, J. C. K. & Karume, K. Climate Change and Its Implications on Food Security in the Great Lakes Region. In Climate Change and Socio-political Violence in Sub-Saharan Africa in the Anthropocene: Perspectives from Peace Ecology and Sustainable Development 113–140 (Springer, 2024).
Tabari, H. Climate change impact on flood and extreme precipitation increases with water availability. Sci. Rep. 10, 13768 (2020).
Lange, K. W. & Nakamura, Y. Edible insects as future food: chances and challenges. J. Futur. foods 1, 38–46 (2021).
Ishara, J. et al. The contribution of commonly consumed edible insects to nutrition security in the Eastern DR Congo. Sci. Rep. 14, 16186 (2024).
Van Huis, A., Rumpold, B., Maya, C. & Roos, N. Nutritional qualities and enhancement of edible insects. Annu. Rev. Nutr. 41, 551–576 (2021).
Mishyna, M., Chen, J. & Benjamin, O. Sensory attributes of edible insects and insect-based foods–Future outlooks for enhancing consumer appeal. Trends Food Sci. Technol. 95, 141–148 (2020).
Dobermann, D., Swift, J. A. & Field, L. M. Opportunities and hurdles of edible insects for food and feed. Nutr. Bull. 293–308. https://doi.org/10.1111/nbu.12291. (2017)
Tanga, C. M. et al. Edible insect farming as an emerging and profitable enterprise in East Africa. Curr. Opin. insect Sci. 48, 64–71 (2021).
Panassiti, B. et al. Insects benefit from agri-environmental schemes aiming at grassland extensification. Agric. Ecosyst. Environ. 356, 108613 (2023).
Nowakowski, A. C., Miller, A. C., Miller, M. E., Xiao, H. & Wu, X. Potential health benefits of edible insects. Crit. Rev. Food Sci. Nutr. 62, 3499–3508 (2022).
Koutsos, E., Modica, B. & Freel, T. Immunomodulatory potential of black soldier fly larvae: applications beyond nutrition in animal feeding programs. Transl. Anim. Sci. 6, txac084 (2022).
Bingqian, N. et al. Insect bioactive compounds and their potential use in animal diets and medicine. Entomol. Res. (2023).
Seabrooks, L. & Hu, L. Insects: an underrepresented resource for the discovery of biologically active natural products. Acta Pharm. Sin. B 7, 409–426 (2017).
Akbarian, M., Khani, A., Eghbalpour, S. & Uversky, V. N. Bioactive peptides: Synthesis, sources, applications, and proposed mechanisms of action. Int. J. Mol. Sci. 23, 1445 (2022).
Megido, C. R. et al. Edible insects acceptance by B elgian consumers: promising attitude for entomophagy development. J. Sens. Stud. 29, 14–20 (2014).
Payne, C. L. R., Scarborough, P., Rayner, M. & Nonaka, K. A systematic review of nutrient composition data available for twelve commercially available edible insects, and comparison with reference values. Trends Food Sci. Technol. 47, 69–77 (2016).
Wendin, K. M. E. & Nyberg, M. E. Factors influencing consumer perception and acceptability of insect-based foods. Curr. Opin. food Sci. 40, 67–71 (2021).
Benndorf, R. et al. Natural products from Actinobacteria associated with fungus-growing termites. Antibiotics 7, 83 (2018).
Luo, S.-L. et al. Isocoumarins from American cockroach (Periplaneta americana) and their cytotoxic activities. Fitoterapia 95, 115–120 (2014).
Mebs, D., Pogoda, W., Schneider, M. & Kauert, G. Cantharidin and demethylcantharidin (palasonin) content of blister beetles (Coleoptera: Meloidae) from southern Africa. Toxicon 53, 466–468 (2009).
Wang, J. et al. Functional study on the mutations in the silkworm (Bombyx mori) acetylcholinesterase type 1 gene (ace 1) and its recombinant proteins. Mol. Biol. Rep. 41, 429–437 (2014).
Kellner, R. L. L. & Dettner, K. Allocation of pederin during lifetime of Paederus rove beetles (Coleoptera: Staphylinidae): evidence for polymorphism of hemolymph toxin. J. Chem. Ecol. 21, 1719–1733 (1995).
Dai, C., Ma, H., Luo, L. & Yin, X. Angiotensin I-converting enzyme (ACE) inhibitory peptide derived from Tenebrio molitor (L.) larva protein hydrolysate. Eur. Food Res. Technol. 236, 681–689 (2013).
Cito, A., Botta, M., Francardi, V. & Dreassi, E. Insects as source of angiotensin converting enzyme inhibitory peptides. J. Insects Food Feed 3, 231–240 (2017).
Hadi, J. & Brightwell, G. Safety of alternative proteins: Technological, environmental and regulatory aspects of cultured meat, plant-based meat, insect protein and single-cell protein. Foods 10, 1226 (2021).
Pessina, F. et al. Antihypertensive, cardio-and neuro-protective effects of Tenebrio molitor (Coleoptera: Tenebrionidae) defatted larvae in spontaneously hypertensive rats. PLoS One. 15, e0233788 (2020).
Beemelmanns, C. et al. Macrotermycins A–D, glycosylated macrolactams from a termite-associated Amycolatopsis sp. M39. Org. Lett. 19, 1000–1003 (2017).
Lee, S. R. et al. Natalenamides A–C, cyclic tripeptides from the termite-associated Actinomadura sp. RB99. Molecules 23, 3003 (2018).
Yan, Y.-M. et al. Compounds from the insect Blaps japanensis with COX-1 and COX-2 inhibitory activities. Bioorg. Med. Chem. Lett. 25, 2469–2472 (2015).
Richter, H., Gover, O. & Schwartz, B. Anti-inflammatory activity of black soldier fly oil associated with modulation of tlr signaling: A metabolomic approach. Int. J. Mol. Sci. 24, 10634 (2023).
Tang, J.-J. et al. Constituents from the edible Chinese black ants (Polyrhachis dives) showing protective effect on rat mesangial cells and anti-inflammatory activity. Food Res. Int. 67, 163–168 (2015).
Wijesekara, T. & Xu, B. New insights into sources, bioavailability, health-promoting effects, and applications of chitin and chitosan. J. Agric. Food Chem. 72, 17138–17152 (2024).
Eunice Boko, A. C., Blei, S. H., Koko, A. C. & Angaman, D. M. Evaluating Nudaurelia dione (Saturniidae), an Edible Insect, for Sustainable Nutrition: Composition, Benefits, and Antinutritional Insights. J. Food Biochem. 2024, 5559567 (2024).
Mu, L., Wu, L., Wu, S., Ye, Q. & Zhong, Z. Progress in chitin/chitosan and their derivatives for biomedical applications: Where we stand. Carbohydr. Polym. 122233 (2024).
Ahn, M. Y., Han, J. W., Hwang, J. S., Yun, E. Y. & Lee, B. M. Anti-inflammatory effect of glycosaminoglycan derived from Gryllus bimaculatus (a type of cricket, insect) on adjuvant-treated chronic arthritis rat model. J. Toxicol. Environ. Heal. Part A. 77, 1332–1345 (2014).
Ahn, M. Y., Hwang, J. S., Kim, M.-J. & Park, K.-K. Antilipidemic effects and gene expression profiling of the glycosaminoglycans from cricket in rats on a high fat diet. Arch. Pharm. Res. 39, 926–936 (2016).
Ahn, M. Y. et al. Anti-diabetic activity of field cricket glycosaminoglycan by ameliorating oxidative stress. BMC Complement. Med. Ther. 20, 1–10 (2020).
Yan, Y.-M., Luo, Q., Li, J.-J., Tu, Z.-C. & Cheng, Y.-X. Novel spirooxindole alkaloid derivatives from the medicinal insect Blaps japanensis and their biological evaluation. Bioorg. Chem. 141, 106845 (2023).
Lu, J. et al. Probing the antioxidant activity of functional proteins and bioactive peptides in Hermetia illucens larvae fed with food wastes. Sci. Rep. 12, 2799 (2022).
Dontsov, A. E. et al. Ommochromes from the compound eyes of insects: physicochemical properties and antioxidant activity. Biochem. 85, 668–678 (2020).
Zielińska, E., Baraniak, B. & Karaś, M. Antioxidant and anti-inflammatory activities of hydrolysates and peptide fractions obtained by enzymatic hydrolysis of selected heat-treated edible insects. Nutrients 9, 970 (2017).
Di Mattia, C., Battista, N., Sacchetti, G. & Serafini, M. Antioxidant activities in vitro of water and liposoluble extracts obtained by different species of edible insects and invertebrates. Front. Nutr. 6, 106 (2019).
Li, Z. et al. Antibacterial and immunomodulatory activities of insect defensins-DLP2 and DLP4 against multidrug-resistant Staphylococcus aureus. Sci. Rep. 7, 12124 (2017).
Ai, H., Wang, F., Zhang, N., Zhang, L. & Lei, C. Antiviral, immunomodulatory, and free radical scavenging activities of a protein-enriched fraction from the larvae of the housefly, Musca domestica. J. Insect Sci. 13, 112 (2013).
Li, Z. et al. Purification and characterization of a novel immunomodulatory hexapeptide from alcalase hydrolysate of ultramicro-pretreated silkworm (Bombyx mori) pupa protein. J. Asia. Pac. Entomol. 22, 633–637 (2019).
Chen, Y. et al. Purification of novel polypeptides from bee pupae and their immunomodulatory activity in vivo and in vitro. J. Insects Food Feed 8, 1117–1132 (2022).
Saidemberg, D. M., da Silva-Filho, L. C., Tognoli, L. M. M. C., Tormena, C. F. & Palma, M. S. Polybioside, a neuroactive compound from the venom of the social wasp Polybia paulista. J. Nat. Prod. 73, 527–531 (2010).
Bosque, I., Gonzalez-Gomez, J. C., Loza, M. I. & Brea, J. Natural tetraponerines: A general synthesis and antiproliferative activity. J. Org. Chem. 79, 3982–3991 (2014).
Lu, J. et al. Sesquiterpene acids from Shellac and their bioactivities evaluation. Fitoterapia 97, 64–70 (2014).
Vihakas, M. A., Kapari, L. & Salminen, J.-P. New types of flavonol oligoglycosides accumulate in the hemolymph of birch-feeding sawfly larvae. J. Chem. Ecol. 36, 864–872 (2010).
Cheseto, X. et al. Potential of the desert locust Schistocerca gregaria (Orthoptera: Acrididae) as an unconventional source of dietary and therapeutic sterols. PLoS One 10, e0127171 (2015).
Islam, M. M. & Yang, C.-J. Efficacy of mealworm and super mealworm larvae probiotics as an alternative to antibiotics challenged orally with Salmonella and E. coli infection in broiler chicks. Poult. Sci. 96, 27–34 (2017).
Stull, V. J. et al. Impact of edible cricket consumption on gut microbiota in healthy adults, a double-blind, randomized crossover trial. Sci. Rep. 8, 1–13 (2018).
Borrelli, L. et al. Insect-based diet, a promising nutritional source, modulates gut microbiota composition and SCFAs production in laying hens. Sci. Rep. 7, 16269 (2017).
Liaqat, F. & Eltem, R. Chitooligosaccharides and their biological activities: A comprehensive review. Carbohydr. Polym. 184, 243–259 (2018).
Nino, M. C., Reddivari, L., Osorio, C., Kaplan, I. & Liceaga, A. M. Insects as a source of phenolic compounds and potential health benefits. J. Insects Food Feed 7, 1077–1088 (2021).
Torres-Castillo, J. A. & Olazarán-Santibáñez, F. E. Insects as source of phenolic and antioxidant entomochemicals in the food industry. Front. Nutr. 10, 1133342 (2023).
Kim, J.-K. et al. Anti-inflammatory effect of roasted licorice extracts on lipopolysaccharide-induced inflammatory responses in murine macrophages. Biochem. Biophys. Res. Commun. 345, 1215–1223 (2006).
Kreuzenbeck, N. B. et al. Isolation, (bio)synthetic studies and evaluation of antimicrobial properties of drimenol-type sesquiterpenes of Termitomyces fungi. Commun. Chem. 6, 79 (2023).
Xu, X. et al. Diversity, bacterial symbionts, and antimicrobial potential of termite-associated fungi. Front. Microbiol. 11, 300 (2020).
Zhou, Y. et al. Nutritional Composition, Health Benefits, and Application Value of Edible Insects: A Review. Foods 11, 3961 (2022).
Kang, C. K. et al. Antibacterial cotton fibers treated with silver nanoparticles and quaternary ammonium salts. Carbohydr. Polym. 151, 1012–1018 (2016).
Lu, J. et al. Identification of N-acetyldopamine dimers from the dung beetle Catharsius molossus and their COX-1 and COX-2 inhibitory activities. Molecules 20, 15589–15596 (2015).
An, J. S. et al. Lenzimycins A and B, metabolites with antibacterial properties from Brevibacillus sp. associated with the dung beetle Onthophagus lenzii. Front. Microbiol. 11, 599911 (2020).
Correa, Y. et al. Identification and characterization of compounds from Chrysosporium multifidum, a fungus with moderate antimicrobial activity isolated from Hermetia illucens gut microbiota. PLoS One 14, e0218837 (2019).
Mudalungu, C. M., Tanga, C. M., Kelemu, S. & Torto, B. An overview of antimicrobial compounds from African edible insects and their associated microbiota. Antibiotics 10, 621 (2021).
Pettit, G. R., Ye, Q., Herald, D. L., Hogan, F. & Pettit, R. K. Antineoplastic agents. 573. isolation and structure of papilistatin from the papilionid butterfly Byasa polyeuctes termessa. J. Nat. Prod. 73, 164–166 (2010).
Klassen, J. L., Lee, S. R., Poulsen, M., Beemelmanns, C. & Kim, K. H. Efomycins K and L from a termite-associated Streptomyces sp. M56 and their putative biosynthetic origin. Front. Microbiol. 10, 1739 (2019).
Dong, Q.-F. et al. Antifungal activity of crude extracts and fat-soluble constituents of Holotrichia diomphalia larvae. Bioresour. Technol. 99, 8521–8523 (2008).
Harikrishnan, R., Kim, J.-S., Balasundaram, C. & Heo, M.-S. Immunomodulatory effects of chitin and chitosan enriched diets in Epinephelus bruneus against Vibrio alginolyticus infection. Aquaculture 326, 46–52 (2012).
Marono, S. et al. Productive performance and blood profiles of laying hens fed Hermetia illucens larvae meal as total replacement of soybean meal from 24 to 45 weeks of age. Poult. Sci. 96, 1783–1790 (2017).
Liu, Y. et al. Hormonal and nutritional regulation of insect fat body development and function. Arch. Insect Biochem. Physiol. Publ. Collab. Entomol. Soc. Am. 71, 16–30 (2009).
Zhao, P. et al. Genome-wide identification and immune response analysis of serine protease inhibitor genes in the silkworm, Bombyx mori. PLoS One 7, e31168 (2012).
Hahn, T. et al. Current state of chitin purification and chitosan production from insects. J. Chem. Technol. Biotechnol. 95, 2775–2795 (2020).
Finke, M. D. Complete nutrient content of four species of feeder insects. Zoo. Biol. 32, 27–36 (2013).
Belluco, S. et al. Edible insects in a food safety and nutritional perspective: a critical review. Compr. Rev. food Sci. food Saf. 12, 296–313 (2013).
Kulma, M., Kou, L., Homolková, D. & Plachý, V. Effect of developmental stage on the nutritional value of edible insects. A case study with Blaberus craniifer and Zophobas morio. J. Food Compos. Anal. 92, (2020).
Kong, B. & Xiong, Y. L. Antioxidant activity of zein hydrolysates in a liposome system and the possible mode of action. J. Agric. Food Chem. 54, 6059–6068 (2006).
da Silva Lucas, A. J., de Oliveira, L. M., Da Rocha, M. & Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chem. 311, 126022 (2020).
Teixeira, C. S. S., Villa, C., Costa, J., Ferreira, I. M. & Mafra, I. Edible insects as a novel source of bioactive peptides: A systematic review. Foods 12, 2026 (2023).
Psarianos, M., Aghababaei, F. & Schlüter, O. K. Bioactive compounds in edible insects: Aspects of cultivation, processing and nutrition. Food Res. Int. 115802 (2025).
Gullón, P., Gullón, B., Romaní, A., Rocchetti, G. & Lorenzo, J. M. Smart advanced solvents for bioactive compounds recovery from agri-food by-products: A review. Trends Food Sci. Technol. 101, 182–197 (2020).
Karrar, E. et al. Lipid-soluble vitamins from dairy products: extraction, purification, and analytical techniques. Food Chem. 373, 131436 (2022).
Farooq, S., Abdullah, Zhang, H. & Weiss, J. A comprehensive review on polarity, partitioning, and interactions of phenolic antioxidants at oil–water interface of food emulsions. Compr. Rev. Food Sci. Food Saf. 20, 4250–4277 (2021).
Rose, A., Jaczynski, J. & Matak, K. Extraction of lipids from insect powders using a one-step organic solvent extraction process. Futur. Foods 4, 100073 (2021).
An, R. et al. Isolation, purification and identification of biological compounds from Beauveria sp. and their evaluation as insecticidal effectiveness against Bemisia tabaci. Sci. Rep. 11, 12020 (2021).
Giovannoni, S. et al. Determination of variability of terpenes and terpenoids in Cannabis sativa by gas chromatography-flame ionization detection and gas chromatography-mass spectrometry. J. Chromatogr. A 1687, 463669 (2023).
da Silva Lucas, A. J. et al. Extraction, physicochemical characterization, and morphological properties of chitin and chitosan from cuticles of edible insects. Food Chem. 343, 128550 (2021).
Yea, C. S., Ahmadi, R., Zarei, M. & Muhialdin, B. J. Fractionation and Purification of Bioactive Peptides. in Bioactive Peptides from Food 267–298 (CRC Press, 2022).
Gupta, P. et al. Mass spectrometry-based technology and workflows for studying the chemistry of fungal endophyte derived bioactive compounds. ACS Chem. Biol. 16, 2068–2086 (2021).
Shakir, N. et al. Impact of NaCl stress on phytoconstituents and bioactivity of Matricaria chamomilla: a multi-analytical approach. Sci. Rep. 14, 19717 (2024).
Rivas-Navia, D. M. et al. Bioactive compounds of insects for food use: Potentialities and risks. J. Agric. Food Res. 14, 100807 (2023).
Zhang, J., Li, M., Han, Z. & Shao, J.-H. A review of insect peptides: From extraction and preparation to bioactivities. Curr. Opin. Food Sci. 101301 (2025).
Stull, V. J. & Weir, T. L. Chitin and omega-3 fatty acids in edible insects have underexplored benefits for the gut microbiome and human health. Nat. Food 4, 283–287 (2023).
Mudalungu, C. M., Mokaya, H. O. & Tanga, C. M. Beneficial sterols in selected edible insects and their associated antibacterial activities. Sci. Rep. 13, 10786 (2023).
Siddiqui, S. A. et al. Legal situation and consumer acceptance of insects being eaten as human food in different nations across the world–A comprehensive review. Compr. Rev. Food Sci. Food Saf. (2023).
Sidali, K. L., Pizzo, S., Garrido-Pérez, E. I. & Schamel, G. Between food delicacies and food taboos: A structural equation model to assess Western students’ acceptance of Amazonian insect food. Food Res. Int. 115, 83–89 (2019).
Kelemu, S. et al. African edible insects for food and feed: inventory, diversity, commonalities and contribution to food security. J. Insects Food Feed 1, 103–119 (2015).
Ishara, J. et al. Inventory reveals wide biodiversity of edible insects in the Eastern Democratic Republic of Congo. Sci. Rep. 12, 1576 (2022).
Imathiu, S. Benefits and food safety concerns associated with consumption of edible insects. NFS J. 18, 1–11 (2020).
Ng’ang’a, J. et al. Microbial quality of edible grasshoppers Ruspolia differens (Orthoptera: Tettigoniidae): From wild harvesting to fork in the Kagera Region, Tanzania. J. Food Saf. 39, e12549 (2019).
Borges, M. M., da Costa, D. V., Trombete, F. M. & Câmara, A. K. F. I. Edible insects as a sustainable alternative to food products: An insight into quality aspects of reformulated bakery and meat products. Curr. Opin. Food Sci. 46, 100864 (2022).
Van Huis, A. and Oonincx D. G. A. B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. https://doi.org/10.1007/s13593-017-0452-8 (2017)
Ishara, J. et al. Insights into the effects of geographical sourcing area on nutrient composition and sensory attributes of nine edible insects. Sci. Rep. 15, 11610 (2025).
Hopkins, I., Farahnaky, A., Gill, H., Newman, L. P. & Danaher, J. Australians’ experience, barriers and willingness towards consuming edible insects as an emerging protein source. Appetite 169, 105832 (2022).
Mishyna, M., Chen, J. & Benjamin, O. Sensory attributes of edible insects and insect-based foods – Future outlooks for enhancing consumer appeal. Trends Food Sci. Technol. https://doi.org/10.1016/j.tifs.2019.11.016 (2019)
Melgar-lalanne, G., Hernandez-Alvarez, A.-J. & Salinas-Castro, A. Edible Insects Processing: Traditional and Innovative Technologies. Compr. Rev. Food Sci. Food Saf. 18, (2019).
Jiang, N. et al. Design, synthesis and antiproliferative activity of novel 2-substituted-4-amino-6-halogenquinolines. Molecules 17, 5870–5881 (2012).
Zhu, H.-J., Xu, T., Yan, Y.-M., Tu, Z.-C. & Cheng, Y.-X. Neolignans and norlignans from insect medicine Polyphaga plancyi and their biological activities. Nat. Products Bioprospect. 11, 51–62 (2021).
Kim, K. H. et al. Natalamycin A, an ansamycin from a termite-associated Streptomyces sp. Chem. Sci. 5, 4333–4338 (2014).
Kang, H. R. et al. Termisoflavones A–C, isoflavonoid glycosides from termite-associated Streptomyces sp. RB1. J. Nat. Prod. 79, 3072–3078 (2016).
Wyche, T. P. et al. Linear peptides are the major products of a biosynthetic pathway that encodes for cyclic depsipeptides. Org. Lett. 19, 1772–1775 (2017).
Carr, G. et al. Microtermolides A and B from termite-associated Streptomyces sp. and structural revision of vinylamycin. Org. Lett. 14, 2822–2825 (2012).
von Bonin, A. et al. Efomycine M: an inhibitor of selectins?. Nat. Med. 12, 873 (2006).
Twu, N.-F., Srinivasan, R., Chou, C.-H., Wu, L.-S. & Chiu, C.-H. Cantharidin and norcantharidin inhibit caprine luteal cell steroidogenesis in vitro. Exp. Toxicol. Pathol. 64, 37–44 (2012).
Kim, S.-H. et al. Tripartin, a histone demethylase inhibitor from a bacterium associated with a dung beetle larva. Org. Lett. 15, 1834–1837 (2013).
Kim, M. J. et al. Population genetic characterization of the endangered dung beetle Copris tripartitus (Coleoptera: Scarabaeidae) using novel microsatellite markers. J. Asia. Pac. Entomol. 25, 101899 (2022).
Um, S. et al. Naphthoquinone–oxindole alkaloids, coprisidins A and B, from a gut-associated bacterium in the dung beetle, Copris tripartitus. Org. Lett. 18, 5792–5795 (2016).
Park, S.-H. et al. Tripartilactam, a cyclobutane-bearing tricyclic lactam from a Streptomyces sp. in a dung beetle’s brood ball. Org. Lett. 14, 1258–1261 (2012).
Blodgett, J. A. V. et al. Common biosynthetic origins for polycyclic tetramate macrolactams from phylogenetically diverse bacteria. Proc. Natl Acad. Sci. 107, 11692–11697 (2010).
Prasad, S. B. & Verma, A. K. Cantharidin-mediated ultrastructural and biochemical changes in mitochondria lead to apoptosis and necrosis in murine Dalton’s lymphoma. Microsc. Microanal. 19, 1377–1394 (2013).
Zhao, Y. et al. SoNar, a highly responsive NAD+/NADH sensor, allows high-throughput metabolic screening of anti-tumor agents. Cell Metab. 21, 777–789 (2015).
Dettner, K. et al. Occurrence of terpene anhydride Palasonin and Palasoninimide in blister beetle Hycleus lunata (Coleoptera: Meloidae). Biochem. Syst. Ecol. 31, 203–205 (2003).
Lang, E. & Lang, F. Triggers, inhibitors, mechanisms, and significance of eryptosis: the suicidal erythrocyte death. Biomed Res. Int. 2015, (2015).
Rivero-Pino, F., Guadix, A. & Guadix, E. M. Identification of novel dipeptidyl peptidase IV and α-glucosidase inhibitory peptides from Tenebrio molitor. Food Funct. 12, 873–880 (2021).
Zielińska, E., Baraniak, B. & Karaś, M. Identification of antioxidant and anti-inflammatory peptides obtained by simulated gastrointestinal digestion of three edible insects species (Gryllodes sigillatus, Tenebrio molitor, Schistocerca gragaria). Int. J. Food Sci. Technol. 53, 2542–2551 (2018).
Zielińska, E., Karaś, M., Baraniak, B. & Jakubczyk, A. Evaluation of ACE, α-glucosidase, and lipase inhibitory activities of peptides obtained by in vitro digestion of selected species of edible insects. Eur. Food Res. Technol. 246, 1361–1369 (2020).
Tang, J.-J. et al. Dopamine derivatives from the insect Polyrhachis dives as inhibitors of ROCK1/2 and stimulators of neural stem cell proliferation. Tetrahedron 70, 8852–8857 (2014).
Shi, Y.-N. et al. Bioactive compounds from the insect Aspongopus chinensis. Bioorg. Med. Chem. Lett. 24, 5164–5169 (2014).
de Carvalho, F. M. et al. Brazilian red propolis: Extracts production, physicochemical characterization, and cytotoxicity profile for antitumor activity. Biomolecules 10, 726 (2020).
Arbiser, J. L. et al. Solenopsin, the alkaloidal component of the fire ant (Solenopsis invicta), is a naturally occurring inhibitor of phosphatidylinositol-3-kinase signaling and angiogenesis. Blood 109, 560–565 (2007).
Zhang, Y. et al. Molecular mechanisms of novel peptides from silkworm pupae that inhibit α-glucosidase. Peptides 76, 45–50 (2016).
Kong, W.-H. et al. Antiobesity effects and improvement of insulin sensitivity by 1-deoxynojirimycin in animal models. J. Agric. Food Chem. 56, 2613–2619 (2008).
Liu, S. et al. Antioxidant activity and phenolic compounds of Holotrichia parallela Motschulsky extracts. Food Chem. 134, 1885–1891 (2012).
Pettit, G. R., Meng, Y., Herald, D. L., Knight, J. C. & Day, J. F. Antineoplastic Agents. 553. The Texas Grasshopper Brachystola m agna. J. Nat. Prod. 68, 1256–1258 (2005).
Hall, F., Reddivari, L. & Liceaga, A. M. Identification and characterization of edible cricket peptides on hypertensive and glycemic in vitro inhibition and their anti-inflammatory activity on RAW 264.7 macrophage cells. Nutrients 12, 3588 (2020).
Acknowledgements
We would like to thank JKUAT, UEA and the RUFORUM for their support.
Author information
Authors and Affiliations
Contributions
J.I., S.N., K.K. and J.K. contributed to the research design, wrote and revised the manuscript; J.I. processed data, conceptualization and formal analysis; J.I., S.N., K.K. and J.N. data curation and investigation; J.I. drafted the manuscript; all authors reviewed the manuscript. All authors contributed to this work and approved the final text of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ishara, J., Ekaette, I., Matendo, R. et al. Potential health benefits of insect bioactive metabolites and consumer attitudes towards edible insects. npj Sci Food 9, 195 (2025). https://doi.org/10.1038/s41538-025-00549-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41538-025-00549-x
