
Abstract
Combination antiretroviral therapy controls human immunodeficiency virus-1 (HIV) but cannot eradicate latent proviruses in immune cells, which reactivate upon treatment interruption. Anti-latency therapies like “shock-and-kill” are being developed but are yet to succeed due to the complexity of latency mechanisms. This review discusses recent advances in understanding HIV latency via mathematical modeling, covering key regulatory factors and models to predict latency reversal, highlighting gaps to guide future therapeutic approaches.
Similar content being viewed by others

Introduction
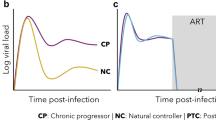
HIV latency remains a major barrier to achieving a HIV cure. When HIV enters a susceptible cell, its RNA is reverse-transcribed and integrates semi-randomly1 into the host genome, forming a provirus [Fig. 1a]. The provirus can follow one of two developmental fates: active-replication or latency. Actively replicating proviruses sustain a relatively high gene-expression activity, driving HIV proliferation and acquired immunodeficiency syndrome (AIDS) progression. In particular, after ~40h2 of activity, the accumulation of viral products within the intracellular space drives host-cell lysis, allowing the hundreds of assembled virions3 to diffuse and infect nearby susceptible cells [Fig. 1b, c]. Antiretroviral therapy (ART) acts disrupting the critical steps of the HIV replication program, interrupting viral proliferation, and allowing the progressive depletion of productively-infected cells. However, even after decades of fully-effective treatment, ART interruption causes almost certain viremia rebound within 2-8 weeks4,5. Evidence suggest that this rebound is due to long-lived proviruses activating from latency6,7,8 [Fig. 1d]. Latent proviruses are unbale to sustain significant viral activity9 due to molecular blocks hindering their gene-expression program. However, their dormancy is reversible and they switch to active replication as soon as the blocks are lifted. Moreover, as long as the provirus is silent, the hosting cell is virtually indistinguishable from its healthy counterpart, allowing the provirus to survive without replicating. Therefore, given their ability to potentially ignite HIV proliferation after years of dormancy, latent proviruses represent the main obstacle for curing HIV10,11.

a HIV cell infection, proliferation, and latency reversal. Once HIV enters a susceptible cell (i.e., active CD4 + T-lymphocyte), is reverse-transcribed, and integrates within the host genome, forming a provirus. b Transitioning into active-replication. Upon sufficiently-high transactivator of transcription (Tat) production, a provirus enters active replication, forcing the host-cell to fabricate and release virions. c host-cell loss and HIV proliferation. The accumulation of viral products leads to host-cell apoptosis within ~40 h of active replication. d Transitioning into latency. Conversely, upon low Tat concentrations, the provirus remains silent. As opposed to what was initially believed, active replication and latency can be detected both in active and resting lymphocytes, represented by green and purple cells, respectively. e Latency-reversal. Upon increased Tat levels a dormant provirus activates from latency. The shock-and-kill therapy employes latency reversing agents to enhance Tat production and facilitate latency-reversal. [Created in BioRender. Palma, P. (2025) https://BioRender.com/0fw3cwk].
Curative strategies now aim at reducing the reservoir of latent HIV proviruses. The most studied method is called shock-and-kill12. This method implies using molecules known as latency reversing agents (LRAs) to disrupt the mechanisms that keep the provirus silent (shock phase). Then, once activated, the provirus would be cleared by cytolytic effects and immune response (kill phase) [Fig. 1e, f]. Developing effective LRAs is critical. It requires identifying the molecular factors regulating proviral latency and their mechanisms of action [Tables 1–3].
Initial observations suggested that proviral activation is a secondary effect of the host-cell immune-activation13,14. In other words, the state of a provirus depends on the state of its host15. However, efforts to reverse latency solely by altering the host-cell state has proven this principle to be inaccurate16. Proviruses do not always enter latency when the host-cell becomes quiescent, and some latent proviruses fail to activate when the immune system is active16,17. Mathematical modeling applied to single-cell experimental data has revealed that HIV self-regulates its transcription through an intrinsic mechanism based on the viral protein transactivator of transcription (Tat)17,18,19,20. These models demonstrated that HIV latency is not simply an “OFF” state but rather a dynamically maintained basal regime, characterized by low Tat levels. When Tat accumulates beyond a critical threshold, it triggers a positive feedback-loop that bypass host regulation17,19,21, amplifies viral transcription18,20,22, and shift the provirus into a robust viral regime [Fig. 1b, d]. Subsequent findings suggested that proviral fate is also regulated by post-transcriptional steps, including splicing, nuclear export, and translation23. Host factors such as RNA-binding proteins (e.g., MATR3, PTB, and PSF) and specific microRNAs act to sequester, degrade, or otherwise inhibit the efficient export and translation of HIV RNAs, thereby enforcing a block to virus reactivation24. According to these findings, the development of anti-latency strategies requires understanding of what host factors are regulating basal transcriptional and post-transcriptional activity, and how the Tat circuit reacts to basal activity20,25,26,27,28,29. Experimental assays that reveal the molecular microenvironment (MME) of integrated proviruses, together with HIV gene-expression models were developed to aid the design of personalized LRAs, predicting the efficacy of a given treatment through in-silico experiments30,31,32,33,34.
Recent literature reviews on HIV latency and anti-latency therapies primarily focus on molecular mechanisms, latency-reversing agents, and host factors driving viral persistence35,36,37,38. In contrast, our review adopts a systems biology perspective, examining HIV gene-expression as a circuit influenced by feedback loops, stochastic events, and regulatory motifs. In the first section of this review, we present the models developed to understand the basal HIV gene-expression dynamics27,28,29, and the main factors regulating its steps. In the second section we focus on the Tat circuit models, developed to elucidate how Tat reacts to the basal expression activity and identify the threshold to active-replication17,18,20,25,26. Building on the knowledge reviewed in these first two section we conclude showing the models developed to simulate perturbations to the HIV gene-expression circuit and predict the efficacy of anti-latency therapies, to proceed with their development39,40,41,42,43,44. By bridging virology with quantitative modeling, our interdisciplinary approach offers fresh insights into HIV latency mechanisms and provides a framework for optimizing therapeutic interventions, paving the way for future research at the nexus of systems biology and HIV treatment.
Basal HIV gene-expression control
Analyzes of basal HIV gene-expression reveal that viral activity is generally low on average and subject to significant intrinsic variability27 – often described as transcriptional noise. Knowing that latency exit occurs stochastically, understanding the mechanisms of transcriptional noise and its main regulatory factors could provide strategies to reverse latency. Early modeling efforts attributed this intrinsic noise to stochastic fluctuations inherent in the mRNA synthesis and degradation processes. This hypothesis led to the development of the constitutive model of HIV gene-expression30 [Fig. 2a]. In this model the HIV promoter—located in the 5’ long-terminal-repeat (LTR) – actively sustains HIV transcription (mRNA), leading to the synthesis of viral proteins (P).

a Bursting model. In this model the HIV promoter is allowed to switch between an active and inactive state, represented by LTRON and LTROFF, respectively. The transition is driven by the rates kon and koff, which represent the frequency of the molecular interactions dynamically shaping the LTR configuration that push the LTR towards a favorable or inhibited state, respectively. The LTR becomes active when the epigenetic editors (EE) allow for an open chromatin, and when the positive transcription factors (TFs), transcription machinery components (i.e., general transcription factors), and positive elongation factors (EF) are successfully recruited by the LTR. On the other hand, the LTR inactivates when the EE promote the formation of thick chromatin, when negative TF and/or EF are recruited and their positive counterparts are dismissed. When the LTR is transcriptionally-active, viral RNA is transcribed at rate kbasal, representing successful recruitment of RNA polymerase II (RNAPII), and viral proteins are synthesized by it at rate kP. Simultaneously, viral transcripts and proteins are lost by degradation at rate dm, and dP, respectively. b Multi-bursting model. In this model the molecular interactions regulating the two transcriptional steps initiation and elongation are explicitly tracked. The LTR switches to its active state LTRON at rate kon, allowing transcription initiation, while it becomes transcriptionally inactive, LTROFF at rate koff. The LTR activates upon removal of epigenetic blocks, and recruitment of positive TF and GTFs. Upon RNAPII recruitment transcription may be paused at rate kpause, alternatively, it is successfully performed at rate kbasal. The pausing takes place when elongation blocks are in place, such as recruitment of negative EF and/or nucleosome-1 positioning. In that case the LTR assumes a paused configuration LTRpause. When transcription is paused, RNAPII may abort transcription at rate kabort, or is released and produces elongated transcripts at rate krelease. This dynamic results in the production of RNAPII convoys. The convoys were modeled considering the number of RNAPII involved (Npol), their time-spacing (tspace), the elongation rate vel and the time before completing an iteration, tproc, (convoys not depicted). [Created in BioRender. Palma, P. (2025) https://BioRender.com/6uc4cie].
Although the constitutive model could not be validated biologically27,45, later studies proposed an alternative explanation. They suggested that the HIV promoter stochastically alternates between active and inactive states, promoting or inhibit transcription, respectively. This mechanism forms the basis of the two-state, or random telegraph, model of HIV expression27,45,46,47 [Fig. 2a]. In this model, stochastic transitions between LTR states generate randomly timed transcriptional bursts.
These postulated LTR “ON” and “OFF” states could be explained biologically. In fact, fluctuating molecular interactions at the provirus integration site (IS) dynamically alter the LTR configuration, “locking” and “unlocking” transcriptional steps28,29. According to this view, identifying the factors interacting with the LTR and shaping its configuration would lead to regulatory factors of HIV transcription, and potential transcription-control strategies. These factors can be divided based on the transcriptional step they regulate (i.e., initiation, elongation). Transcription initiation requires an LTR surrounded by open chromatin, and successful recruitment of positive transcription factors (TFs) and general transcription factors (GTFs)48. This configuration allows RNA polymerase II (RNAPII) to dock and start processing the viral genome. Subsequently, transcription elongation requires RNAPII maturation and nucleosome-1 displacement19,49,50.
Subsequent studies using single-molecule RNA microscopy showed that the HIV promoter fluctuates over two independently-regulated inactive states, characterized by two distinct time scales28. The first time scale represents the time-delay between subsequent emergence of LTR configurations favorable for transcription initiation. Whereas the second time scale is shorter and represents the interval between elongation-free LTR configuration, characterizing the length of transcriptional bursts. It was also observed that transcriptional bursts are characterized by convoys of RNAPII that reinitiate transcription upon successful elongation. Moreover, the same studies demonstrated that RNAPII pausing is not obligatory but stochastic29. These observations led to the development of the multi-scale HIV bursting model28,29 [Fig. 2b].
Despite the validity of the discussed HIV circuit models, recent studies showed that viral expression is also regulated during transcription completion, and post-transcriptional steps, including splicing and nuclear export23. These observations led to the development of more comprehensive models to understand how these steps affect the HIV expression dynamics and what are the involved molecular factors51,52.
From a broader perspective, the LTR state and subsequent post-transcriptional steps, depend on the epigenetic profile and the composition of the molecular cluster fluctuating at the provirus IS. These factors are dynamically regulated by the host and characterize the provirus MME45,53,54 [Fig. 3a]. The host configures the MMEs along its genome to control the gene-expression activity of the genomic entities in-location. Therefore, MMEs at diverse genomic coordinates would be subjected to diverse regulatory inputs over time [Fig. 3b–d]. This explains why different IS shows different expression distributions and why non-specific stimuli, such as the host-cell immune activation, do not homogeneously affect all proviruses. Following this scheme, the range of expression levels that a provirus can potentially experience depends on the genomic entities populating the IS27,46,55,56. For example, the MMEs of transcriptionally-repressed locations, such as centromeric heterochromatin, would never shift to favorable configurations, underlying a null expression potential. In contrast, MMEs at genic or intergenic regions are eventually regulated by the host to induce or inhibit local activity, underlying a variable activation potential.

a Molecular microenvironment (MME) at the provirus integration site (IS). MMEs represent the epigenetic profile and the cluster of diffusing molecules at a given genomic location. The epigenetic profile is determined by the level of DNA methylation and nucleosomes positioning. The molecular cluster is composed of molecular groups, each regulating specific transcriptional and post-transcriptional steps. These main groups are the positive and negative transcription factors (TF+/ TF−) in yellow, positive, and negative elongation factors (EF+/ EF−) in purple, transcription machinery component in blue, and epigenetic editors (EE) in green. The cluster composition affects what reaction would likely rearrange the LTR configuration, and drive splicing and nuclear export. On the other hand, the epigenetic profile determines the chromatin thickness and affect the ability of surrounding molecules to interact with the LTR. b Heuristic representation of MME favorability and allowed MME configurations range. An MME characterized by open chromatin and a favorable molecular cluster composition would favor gene-expression activity. Different ISs would experience the formation of different range of MMEs, depending on the genomic entities populating the location. For example, IS1 represents a provirus integrated within a repressed region, characterized by the static unfavorable MME1. This provirus would eventually experience only a negligible expression activity. Conversely IS2 and IS3 are within expressed regions and their MME can assume different configurations, allowing, in turn, a variable range of gene-expression distributions. c Heuristic representation of basal HIV expression distribution vs IS genomic coordinates and basal HIV expression potential. The MME characterizing a provirus IS also determines the provirus’ basal activity. This activity is modulated both in terms of average and variability (noise). ISs within diverse genomic regions would be potentially exposed to different MMEs depending on the nature of the surrounding genomic entities. According to this view, the basal gene-expression potential – the area underlined by the possible MMEs configurations – is a single-values metric describing the viral expression energy associated with a single genomic location. d Holistic representation of basal HIV expression distributions vs MME. The MME at the provirus IS modulates the basal expression activity. [Created in BioRender. Palma, P. (2025) https://BioRender.com/4la57ft].
Proviral fate
Host-driven HIV expression alone is not sufficient to explain the gene-expression levels, which clearly distinguish active-replication from latency18. The transition between these two regimens requires the interplay of viral factors. In-vitro experiments using the LTR-GFP-IRES-Tat (LGIT) HIV model vector showed that the transition from latency to active-replication is strongly regulated by the viral protein Tat. Upon a threshold level of Tat expression, clones experience a phenotypic bifurcation towards either one of the two developmental fates20. Whereas below and above the threshold, clones stably maintain latency and active-replication, respectively20. Biochemical studies showed that Tat promotes HIV gene-expression bypassing the host during transcriptional and post-transcriptional steps. Tat augments its own production forming a positive feedback-loop, which amplifies viral expression up to ~100 folds57.
Stochastic models were developed to capture the Tat circuit architecture and predict proviral activation20,25,26,34,58. The first models [Fig. 4a] tracked the Tat-driven reactions aimed at lifting elongation blocks20,34, recruiting the positive transcription elongation factor (pTEFb) and nucleosome remodeling complexes (i.e., SWI/SNF)59. Emphasis was given to pTEFb recruitment by deacetylated Tat, and binding to the TAR element, forming and unfolding the Tatd LTR complex. Then Tat is reversibly acetylated, by the host histone acetyltransferase p30059,60, activating, and forming Tata LTR. According to this model, Tat and pTEFb acetylation/deacetylation toggling would explain the stochastic time to activation60. Acetylated Tat promote transcription, then it is released in its deacetylated form and eventually recycled61. Coupling this Tat circuit with an always-active LTR, viral protein synthesis, and reactions of mRNA import/export, was sufficient to well predict phenotypic bifurcation in-vitro62.

a HIV gene-expression model with Tat feedback-loop. In this model the LTR toggles between an active state LTRON and inactive state LTROFF, with rates kon and koff, respectively. An active LTR transcribes viral RNA at rate \({k}_{{basal}}\), which is exported in the cytoplasm at rate \({k}_{{Exp}-{mRNA}}\). Subsequently, viral proteins are synthesized from the mRNA at rate \({k}_{P}\), and the protein \({Tat}\) is synthesized at rate \({k}_{{Tat}}\). Simultaneously, viral transcripts, Tat and the other proteins are lost by degradation at rate \({d}_{m,}\), \({d}_{{Tat}}\), and \({d}_{P}\), respectively. Subsequently, Tat reversibly binds to the LTR at rates \({k}_{{bind}}\) and \({k}_{{unbind}}\) forming the \({Ta}{t}_{d}{LTR}\) complex. Then \({Tat}\) can be acetylated by the host histone acetyltransferase p300 at rate \({k}_{{acetyl}}\), and deacetylated by SirT1 at rate \({k}_{{deacetyl}}\). Finally, acetylated \({Tat}\) triggers transactivated HIV mRNA transcription at rate \({k}_{{transact}}\), resulting the release of Tat in its deacetylated form the LTR, where it can be eventually recycled. b Heuristic model illustrating the relationship between TTA and MME favorability at the proviral IS. (Top Panel) A schematic representation shows how the TTA decreases as the favorability of the MME at the proviral IS increases (black lines). Due to the intrinsic stochasticity of HIV gene expression, the TTA for each MME level is not a single value but rather a distribution—illustrated by the bell-shaped curves along the x-axis. The gray lines delineate the expected TTA boundaries for each MME, while the shaded regions indicate the range of MME favorability associated with two distinct ISs. (Bottom Panel) The lower portion of the figure displays the dynamics of Tat expression for proviruses subjected to different MME conditions, demonstrating how variations in MME favorability impact Tat-driven feedback and viral reactivation. [Created in BioRender. Palma, P. (2025) https://BioRender.com/s0hgdfm].
This model set the foundation to simulate and predict proviral fate in-vitro. Moreover, its analysis suggested that enhancing Tat acetylation facilitate the transitioning to active-replication34. However, the circuit’s intrinsic instability, which allows the two phenotypes to emerge, also causes high sensitivity to small thermal fluctuation. This sensitivity is reflected in a very low probability of observing longer times to activation (TTA), and therefore, long-term latency25. Real-time expression kinetics at the single-cell level showed that basal activity drives Tat pulses, suggesting the presence of some counteracting motif to Tat-amplification25. In the absence of any explicit repressive mechanisms, such as bi-stability and self-cooperativity, it was proposed an enzymatic resistor based on SirT125. This resistor would act rapidly deacetylating Tat and competing with the p300-driven acetylation, decreasing the circuit sensitivity60,63. According to this model stronger resistors accompanied by weaker feedback-loop determine shorter Tat pulses following basal inputs25,26. Subsequent flow cytometry and single-cell imaging analyses on Jurkat cells highlighted the presence of a transient single-molecule threshold, requiring excess of inactive Tat to achieve at least one active molecule58. This result was modeled, introducing an additional Tat inactive state. Therefore, at its core, the Tat circuit is a non-latching positive feedback loop that generates transient pulses of expression, with the strength of the response varying with the regulatory inputs17,18.
According to this view, a provirus transit to active-replication—activates—when its basal activity is sufficiently high to trigger the self-maintenance of high Tat levels20. As discussed, the basal expression levels is stochastic and the distribution is regulated by the host through the provirus MME27,46,55,56. It follows that the MME at the provirus IS impacts the random TTA55— defined as the time required by a provirus to overcome the Tat activation threshold. A provirus exposed to a favorable MME would experience an early transition to active-replication. Conversely, proviruses with a relatively unfavorable MME would eventually activate later in time. This delay would be caused by an increased LTR toggling between active and inactive states due to increasingly significant molecular fluctuations, which lower the average transcriptional activity and increase the noise. Moreover, since the TTA is intrinsically stochastic, identical proviruses exposed to the same MME would transit to active replication at different times20,64.
Subsequent studies showed that Tat has other functions: it opens the chromatin at the LTR and aids GTFs recruitment, facilitating transcription initiation. Moreover, Tat promotes mRNA export and engages the cellular translation machinery, enhancing post-transcriptional steps. Direct evidence of Tat’s differential role in pediatric HIV infection during viral activation remains sparse, necessitating further investigation to uncover age-specific variation in Tat function and their implications for optimizing HIV management in children.
HIV latency and latency reversal
The MME at the proviral IS orchestrates proviral activity. Long-term latency arises when the MME strongly represses viral expression, even though the IS potentially allows more permissive MME configurations. According to this view, latency reversal occurs when the MME is reconfigured into a permissive state. These reconfigurations can happen spontaneously, driven by host-cell reaction network dynamics, or be induced artificially through LRAs.
Initial studies on latency reversal employed transcription activators and chromatin remodelers to facilitate transcription initiation. Activators, such as TNFα and PKC agonists (i.e., prostratin and bryostatin) act as TFs enhancers and were observed to amplify mean HIV expression levels in-vitro48, though with limited efficacy ex-vivo and in-vivo16. The combined use of chromatin remodelers, such as histone deacetylase inhibitors (HDACi) (i.e., vorinostat, panobinostat, and romidepsin), methylation inhibitors, and bromodomain inhibitors has demonstrated a synergic effect, significantly enhancing treatments’ efficacy44,65,66. Moreover, when the intrinsic stochastic nature of gene-expression was discovered67, expression variability was included as a diagnostic metric33. This inclusion led to the discovery of noise modulators33 and the development of a theory explaining LRAs synergies27,45. The theory states that while transcription activators mainly act lifting transcription initiation blocks, increasing the average expression (burst frequency kon↑), noise modulators act relaxing the elongation blocks, increasing the expression noise (burst size kbasal/koff↑). Therefore, each class independently modulates one transcriptional step and their combined usage lead to synergistic effect, amplifying HIV expression level [Fig. 5a–c]. This theory was validated experimentally, leading to the selection of novel LRAs33.

a LTR state, blocks to transcriptional steps and LRAs. The schematic illustrates the key transcriptional blocks at the HIV LTR. Basal transcriptional activity is inhibited by blocks at the initiation step, while elongation blocks cause RNA polymerase pausing. In this context, LRAs that act as transcription activators are designed to overcome the initiation blocks, and the ones that act as noise modulators are intended to relieve elongation blocks. b HIV gene-expression modulation. HIV gene expression is characterized by bursting dynamics. Here, the burst frequency, \({k}_{{on}}\), is primarily governed by the removal or persistence of initiation blocks, whereas the burst size, \({k}_{{basal}}/{k}_{{off}}\), reflects the extent to which transcription is paused at the elongation checkpoint. c LRAs synergy. According to the synergy theory, when transcription activators are applied alone, they shift the system state along a diagonal representing constant burst size (path 1). In contrast, noise modulators alone increase the burst size (path 2). When combined, these LRAs act synergistically, producing an amplified increase in the average transcriptional output (path 3). d HIV gene-expression model explicitly considering LRAs treatment. This schematic illustrates a model of HIV transcription incorporating the modulatory effects of LRAs, specifically PKC agonists and HDACIs. In this framework, the transcription factor NFκB is released into the cytoplasm at a basal rate \({k}_{{NFkB}}\), a process enhanced by PKC agonists, \({\varphi }_{{PKCa}}\). NFκB translocate to the nucleus, \({k}_{{Imp}-{NFkB}}\) and binds the HIV LTR at rate \({k}_{{on}}\), forming the LTR-NFκB complex that initiates basal transcription, \({k}_{{basal}}\). HDACIs further amplify this transcriptional activity, \({\varphi }_{{HDACI}}\), by altering chromatin structure. The transcribed HIV mRNA is exported to the cytoplasm, \({k}_{{Exp}-{mRNA}}\) and translated into viral proteins (\({k}_{P}\)), including Tat, \({k}_{{Tat}}\). Deacetylated Tat is imported into the nucleus, \({k}_{{Imp}-{Tat}}\), binds the active LTR, \({k}_{{bind}}\), and undergoes acetylation, \({k}_{{acetyl}}\). The acetylated Tat-LTR complex significantly enhances transcription (\({k}_{{transact}}\)), establishing a positive feedback loop that drives robust viral gene expression. [Created in BioRender. Palma, P. (2025) https://BioRender.com/ygeoibo].
Building on detailed insights into basal and Tat-activated HIV expression dynamics, researchers have developed mathematical models that simulate the effects of latency LRAs to predict treatment efficacy30,31,32,42. These models typically extend the random telegraph framework, introducing the effect of LRAs on specific steps of the expression program. A recent model includes dose-dependent parameters that represent the nuclear levels of TFs – accounting for both their baseline release and the boost provided by PKC agonist stimulation [Fig. 5d]. In this framework, an active LTR drives transcription at a basal rate in the absence of Tat. Once Tat is acetylated, transcription is significantly enhanced. Both in basal and viral regime, HDACis stimulation further increase the transcriptional output. By coupling the stochastic promoter activity with pharmacokinetic equations, this integrative model forecast the overall reactivation probability of latent cells under different LRA dosing regimens and the kinetic patterns of viral reactivation42. Ultimately, such models serve as quantitative tools for optimizing “shock-and-kill” strategies in HIV cure research – enabling personalized predictions of LRA efficacy based on the specific transcriptional blocks a given provirus experiences.
Personalized medicine is inherently challenged by both inter-individual and intra-individual variability. This important level of heterogeneity complicates the direct applicability of traditional models to predict individual responses. To address this challenge, advanced modeling strategies must be employed68. One promising approach is stochastic modeling, which captures population-level variability and can simulate a wide range of patient responses. In this context, patient-specific parameterization—where models are fine-tuned for individual patients or groups with similar characteristics—proves particularly effective69. Training such models on rich datasets (e.g., virological, and immunological information, including high-throughput single-cell transcriptomics and proteomics) enables them to reflect the unique molecular profile of each patient. Furthermore, the use of Bayesian methodologies allows for real-time updates70. As new patient data become available, predictions can be refined continuously, thereby supporting the ongoing optimization of therapeutic strategies.
Discussion and conclusions
Despite extensive research, significant gaps remain in understanding the mechanisms that regulate HIV gene-expression, particularly concerning the transition from latency to active-replication, known as viral reactivation. These gaps present substantial obstacles to the development of effective LRAs and the accurate prediction of viral rebound—defined as the resurgence of detectable virus levels—following ART interruption. One critical challenge is elucidating how extracellular stimuli impact the MMEs along host-cell DNA and modulate HIV gene-expression. Host factors, such as TFs, transcriptional machinery components (i.e., GTFs, RNAPII), and epigenetic editors, play pivotal roles in regulating the activity of the LTR, shaping its molecular configuration. However, comprehensive models that integrate how variations in MME at the provirus IS influence HIV gene-expression dynamics, such as transcriptional burst size and frequency, are still lacking. Mathematical and computational modeling plays a crucial role in bridging these knowledge gaps, providing tools to predict and simulate the behavior of latent reservoirs under different conditions. Modeling helps clarify the interactions between host factors and viral elements that govern HIV latency and reactivation. By capturing how MME variations affect transcriptional noise and activation thresholds, models allow researchers to identify key regulatory mechanisms and predict how latent proviruses transition to active-replication. This predictive capability is essential for developing personalized LRAs tailored to target high-risk proviruses based on their IS and reactivation potential.
The location of IS within the host genome, along with the nature and frequency of extracellular stimuli, profoundly affects whether a provirus goes dormant or becomes actively-replicating. Developing a comprehensive reactivation likelihood map, informed by robust modeling, that links specific MMEs to the probability of latent proviruses to synthesize Tat and transition to active-replication in response to stimuli, such as immune activation, would be invaluable. Such a map, generated through modeling, would enable clinicians to identify high-risk proviruses more prone to reactivation and tailor LRA therapies more effectively.
The MME at the provirus IS is quantitatively determined using a combination of high-throughput sequencing and advanced computational analyses. IS sequencing precisely maps the locations where HIV integrates into the host genome across different cells and tissues, providing a spatial framework. This mapping is then complemented by epigenomic profiling techniques such as ChIP-seq, which detects histone modifications indicative of active or repressive chromatin states, and ATAC-seq, which measures chromatin accessibility. Additionally, RNA-seq data help correlate these epigenetic signatures with transcriptional activity, offering insights into the activity surround the IS. Computational models integrate these multidimensional datasets to generate quantitative scores representing the MME. These scores encompass parameters like DNA methylation levels, nucleosome positioning, histone mark distributions, and the presence of regulatory elements such as enhancers or promoters. While many models have been initially validated using data from resting CD4 + T cells, recent advances allow them to be adapted to other HIV-harboring cell types, including macrophages and tissue-resident cells, by incorporating cell-specific71. Thus, with comprehensive data integration, these models are broadly applicable and not confined to specific cell types, enabling tailored analysis of the MME in diverse biological compartments.
Predicting viral rebound following ART interruption remains a significant challenge in HIV cure research. While viral reactivation marks the initial process by which latent proviruses become transcriptionally active, viral rebound represents the clinical manifestation of renewed virus replication and detectable levels in plasma. Current predictive models, largely based on population-level dynamics, often fail to capture the heterogeneity of proviruses within an individual’s latent reservoir. Proviruses integrated at different MME may exhibit distinct latency-reversal rates, complicating accurate predictions of viral rebound. Mathematical modeling can incorporate data on IS-specific reactivation potential into viral dynamics models, significantly enhancing predictive accuracy. These improved models can guide treatment decisions and better manage patient expectations
A deeper understanding of the mechanisms by which host factors regulate HIV transcription will enable the development of predictive models that map reactivation potential across different genomic regions. Such models will facilitate more accurate predictions of time to viral rebound and support the creation of personalized LRAs targeting high-risk proviruses. Enhanced modeling approaches also enable the development of in-vitro systems that accurately simulate latency and reactivation dynamics, providing a platform to evaluate the efficacy of novel LRAs and optimize therapeutic strategies.
In pediatric populations, the dynamics of viral reactivation differ significantly from adults due to factors such as immune system maturation. Children, especially infants, have developing immune systems characterized by higher thymic activity and a greater proportion of naïve T cells compared to adults. This influences both the establishment and reactivation of the HIV reservoir. Research indicates that early initiation of ART in children can reduce reservoir size and lower reactivation risk. However, adherence to ART remains a critical challenge; adolescents often face adherence difficulties, leading to higher rates of virological failure and increased risk of viral rebound compared to adults. Mathematical models that incorporate age-specific immune dynamics and adherence patterns are necessary to predict reactivation risk more accurately in children and to design effective interventions. The EPIICAL consortium’s study on perinatally infected infants who initiated ART early highlighted that faster CD4 T-cell reconstitution correlates with viral load resurgences, emphasizing the complex interplay between immune recovery and viral reactivation in pediatric populations72,73,74. Modeling these interactions offers critical insights into how immune recovery influences reactivation risk, guiding personalized treatment strategies and improving outcomes.
In conclusion, a thorough understanding of how host factors regulate HIV transcription, aided by advanced modeling, is crucial for creating predictive reactivation maps across various genomic regions. These models improve predictions of time-to-reactivation and guide the design of personalized LRAs to target high-risk proviruses. By integrating mathematical modeling with experimental data, researchers can develop accurate in vitro models of latency dynamics and LRA effectiveness, moving closer to the goal of HIV eradication.
Data availability
No datasets were generated or analysed during the current study.
References
Schröder, A.R.W. et al. HIV-1 integration in the human genome favors active genes and local hotspots. Cell 110, 521–529 (2002).
Perelson, A. S., Neumann, A. U., Markowitz, M., Leonard, J. M. & Ho, D. D. HIV-1 dynamics in vivo: virion clearance rate, infected cell life-span, and viral generation time. Science 271, 1582–1586 (1996).
Seth, N. et al. Expansion and contraction of HIV-specific CD4 T cells with short bursts of viremia, but physical loss of the majority of these cells with sustained viral replication. J. Immunol. 175, 6948–6958 (2005).
Colby, D. J. et al. Rapid HIV RNA rebound after antiretroviral treatment interruption in persons durably suppressed in Fiebig I acute HIV infection. Nat. Med. 24, 923–926 (2018).
Pannus, P. et al. Rapid viral rebound after analytical treatment interruption in patients with very small HIV reservoir and minimal on-going viral transcription. J. Int. AIDS Soc. 23, 25453 (2020).
Finzi, D. et al. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 278, 1295–1300 (1997).
Chun, T. W. et al. Presence of an inducible HIV-1 latent reservoir during highly active antiretroviral therapy. Proc. Natl. Acad. Sci. USA. 94, 13193–13197 (1997).
Pierson, T., McArthur, J. & Siliciano, R. F. Reservoirs for HIV-1: mechanisms for viral persistence in the presence of antiviral immune responses and antiretroviral therapy. Annu. Rev. Immunol. 18, 665–708 (2000).
Finzi, D. et al. Latent infection of CD4 + T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat. Med. 5, 512–517 (1999).
Richman, D. D. et al. The challenge of finding a cure for HIV infection. Science 323, 1304–1307 (2009).
Weinberger, A. D. & Weinberger, L. S. Stochastic fate selection in HIV-infected patients. Cell 155, 497–499 (2013).
Deeks, S. G. HIV: Shock and kill. Nature 487, 439–440 (2012).
Pearson, R. et al. Epigenetic silencing of human immunodeficiency virus (HIV) transcription by formation of restrictive chromatin structures at the viral long terminal repeat drives the progressive entry of HIV into latency. J. Virol. 82, 12291–12303 (2008).
Tyagi, M. et al. Establishment of HIV latency in primary CD4+ cells is due to epigenetic transcriptional silencing and P-TEFb restriction. J. Virol. 84, 6425–6437 (2010).
Coffin, J. M. & Swanstrom, R. HIV pathogenesis: dynamics and genetics of viral populations and infected cells. Cold Spring Harb. Perspect. Med. 3, 012526–012526 (2013).
Ho, Y. C. et al. Replication-competent noninduced proviruses in the latent reservoir increase barrier to HIV-1 cure. Cell 155, 540–551 (2013).
Razooky, B. S. et al. A hardwired HIV latency program. Cell 160, 990–1001 (2015).
Razooky B. S. et al. Nonlatching positive feedback enables robust bimodality by decoupling expression noise from the mean. PLoS Biol. 15, e2000841 (2017).
Morton, E. L. et al. Transcriptional circuit fragility influences HIV proviral fate. Cell Rep. 27, 154 (2019).
Weinberger, L. S. et al. Stochastic gene expression in a lentiviral positive-feedback loop: HIV-1 Tat fluctuations drive phenotypic diversity. Cell 122, 169–182 (2005).
Karn, J. The molecular biology of HIV latency: breaking and restoring the Tat-dependent transcriptional circuit. Curr. Opin. Hiv. Aids 6, 4–11 (2011).
Singh, A. & Weinberger, L. S. Stochastic gene expression as a molecular switch for viral latency. Curr. Opin. Microbiol. 12, 460–466 (2009).
Yukl, S. A. et al. HIV latency in isolated patient CD4 + T cells may be due to blocks in HIV transcriptional elongation, completion, and splicing. Sci. Transl. Med. 10, eaap9927 (2018).
Kobayashi-Ishihara, M. & Tsunetsugu-Yokota, Y. Post-transcriptional HIV-1 latency: A promising target for therapy?. Viruses 16, 666 (2024).
Weinberger, L. S. & Shenk, T. An HIV feedback resistor: auto-regulatory circuit deactivator and noise buffer. PLoS Biol. 5, e9 (2006).
Weinberger, L. S. et al. Transient-mediated fate determination in a transcriptional circuit of HIV. Nat. Genet. 40, 466–470 (2008).
Singh, A. et al. Transcriptional bursting from the HIV-1 promoter is a significant source of stochastic noise in HIV-1 gene expression. Biophys. J. 98, L32–L34 (2010).
Tantale, K. et al. A single-molecule view of transcription reveals convoys of RNA polymerases and multi-scale bursting. Nat. Commun. 7, 12248–12248 (2016).
Tantale, K. et al. Stochastic pausing at latent HIV-1 promoters generates transcriptional bursting. Nat. Commun. 12, 4503 (2021).
Chavali, A. K. et al. Distinct promoter activation mechanisms modulate noise-driven HIV gene expression. Sci. Rep. 5, 17661–17661 (2015).
Wong, V. C. et al. NF-κB-chromatin interactions drive diverse phenotypes by modulating transcriptional noise. Cell Rep. 22, 585–599 (2018).
Bullock, M. E. et al. A transcriptional cycling model recapitulates chromatin-dependent features of noisy inducible transcription. PLoS Comput. Biol. 18, e1010152 (2022).
Dar, R. D. et al. Screening for noise in gene expression identifies drug synergies. Science 344, 1392–1396 (2014).
Cao, Y. et al. Probabilistic control of HIV latency and transactivation by the Tat gene circuit. Proc. Natl. Acad. Sci. USA. 115, 12453–12458 (2018).
Vemparala, B., Chowdhury, S., Guedj, J. & Dixit, N. M. Modelling HIV-1 control and remission. npj Syst. Biol. Appl. 10, 1–11 (2024).
Kulpa, D. A., Paiardini, M. & Silvestri, G. Immune-mediated strategies to solving the HIV reservoir problem. Nat. Rev. Immunol. 1–12. https://doi.org/10.1038/s41577-025-01136-7 (2025)
Jones, J. E., Gunderson, C. E., Wigdahl, B. & Nonnemacher, M. R. Impact of chromatin on HIV-1 latency: a multi-dimensional perspective. Epigenet. Chromatin 18, 9 (2025).
D’Orso, I. The HIV-1 transcriptional program: from initiation to elongation control. J. Mol. Biol. 437, 168690 (2025).
Deng, Q., Guo, T., Qiu, Z. & Chen, Y. Towards a new combination therapy with vectored immunoprophylaxis for HIV: modeling “shock and kill” strategy. Math. Biosci. 355, 108954–108954 (2023).
Deng, Q., Guo, T., Qiu, Z. & Chen, Y. Modeling the cytotoxicity of Romidepsin reveals the ineffectiveness of this drug in the “shock and kill” strategy. Chaos Solitons Amp Fractals 173, 113702–113702 (2023).
Guo, X., et al. The Nonequilibrium mechanism of noise-enhanced drug synergy in HIV latency reactivation. iScience 25, 104358 https://doi.org/10.1016/j.isci.2022.104358 (2022).
Gupta, V., Dixit, N. M. Trade-off between synergy and efficacy in combinations of HIV-1 latency-reversing agents. PLOS Comput. Biol. 14, e1006004 (2018).
Ke, R. et al. Determinants of the efficacy of HIV latency-reversing agents and implications for drug and treatment design. JCI Insight 3, e123052 (2018).
Laird, G. M. et al. Ex vivo analysis identifies effective HIV-1 latency–reversing drug combinations. J. Clin. Investig. 125, 1901–1912 (2015).
Dae, R. D. et al. Transcriptional burst frequency and burst size are equally modulated across the human genome. Proc. Natl. Acad. Sci. USA. 109, 17454–17459 (2012).
Dey, S. S. et al. Orthogonal control of expression mean and variance by epigenetic features at different genomic loci. Mol. Syst. Biol. 11, 806–806 (2015).
Skupsky, R. et al. HIV promoter integration site primarily modulates transcriptional burst size rather than frequency. PLOS Comput. Biol. 6, 22364–22376 (2010).
Ruelas, D. S. & Greene, W. C. An integrated overview of HIV-1 latency. Cell 155, 519–529 (2013).
Mizutani, T. et al. Loss of the Brm-type SWI/SNF chromatin remodeling complex is a strong barrier to the Tat-independent transcriptional elongation of human immunodeficiency virus type 1 transcripts. J. Virol. 83, 11569–11580 (2009).
Vos, S. M. et al. Structure of paused transcription complex Pol II–DSIF–NELF. Nature 560, 601–606 (2018).
Likhoshvai, V. A. et al. Mathematical model of the Tat-Rev regulation of HIV-1 replication in an activated cell predicts the existence of oscillatory dynamics in the synthesis of viral components. BMC Genom.15, 1–18 (2014).
Bensussen, A. et al. Dynamics of the gene regulatory network of HIV-1 and the role of viral non-coding RNAs on latency reversion. Front. Genet. 9, 1364 (2018).
Raser, J. M. & O’Shea, E. K. Control of Stochasticity in Eukaryotic Gene Expression. Science 304, 1811–1814 (2004).
Özbudak, E. M. et al. Regulation of noise in the expression of a single gene. Nat. Genet. 31, 69–73 (2002).
Jordan, A. et al. The site of HIV-1 integration in the human genome determines basal transcriptional activity and response to Tat transactivation. EMBO J. 20, 1726–1738 (2001).
Miller-Jensen, K. et al. Varying virulence: epigenetic control of expression noise and disease processes. Trends Biotechnol. 29, 517–525 (2011).
Karn, J. et al. Tackling tat. J. Mol. Biol. 293, 235–254 (1999).
Aull, K. H. et al. Transient Thresholding: A Mechanism Enabling Noncooperative Transcriptional Circuitry to Form a Switch. Biophys. J. 112, 2428–2438 (2017).
Mahmoudi, T. et al. The SWI/SNF chromatin-remodeling complex is a cofactor for Tat transactivation of the HIV promoter. J. Biol. Chem. 281, 19960–19968 (2006).
Ott, M. et al. Tat acetylation: a regulatory switch between early and late phases in HIV transcription elongation. Novartis Found. Symp. 259, 182–196 (2004).
Pagans, S. et al. SIRT1 regulates HIV transcription via Tat deacetylation. PLoS Biol. 3, e41(2005).
Wu, Y. HIV-1 gene expression: lessons from provirus and non-integrated DNA. Retrovirology 1, 13 (2004).
Michael, S. et al. Stochastic amplification and signaling in enzymatic futile cycles through noise-induced bistability with oscillations. Proc. Natl. Acad. Sci. USA. 102, 2310–2315 (2005).
Miller-Jensen, K. et al. Genetic selection for context-dependent stochastic phenotypes: Sp1 and TATA mutations increase phenotypic noise in HIV-1 gene expression. PLoS Comput. Biol. 9, e1003135 (2013).
Reuse, S. et al. Synergistic activation of HIV-1 expression by deacetylase inhibitors and prostratin: implications for treatment of latent infection. PLoS ONE 4, e1005063 (2009).
Cnops, L. et al. An In-Depth Comparison of latency-reversing agent combinations in various in vitro and ex vivo HIV-1 latency models identified bryostatin-1 + JQ1 and ingenol-B + JQ1 to potently reactivate viral gene expression. PLOS Pathog. 11, e1005063 (2015).
Kærn, M.et al. Stochasticity in gene expression: from theories to phenotypes. Nat. Rev. Genet. 6, 451–464 (2005).
Kapelner, A. et al. Evaluating the effectiveness of personalized medicine with software. front. Big Data 4, 572532 (2021).
Luedtke, A. R. & van der Laan, M. J. Super-learning of an optimal dynamic treatment rule. Int. J. Biostat. 12, 305–332 (2016).
Zhao, B., Zhong, Y., Kang, J. & Zhao, L. Bayesian learning of Covid-19 vaccine safety while incorporating adverse events ontology. Ann. Appl. Stat. 17, 2887–2902 (2023).
DeMarino, C. et al. Differences in transcriptional dynamics between T-cells and macrophages as determined by a three-state mathematical model. Sci. Rep. 10, 2227–2227 (2020).
Morris, S. E. et al. Quantifying the dynamics of hiv decline in perinatally infected neonates on antiretroviral therapy. JAIDS J. Acquir. Immune Defic. Syndr. 85, 209 (2020).
Morris, S. E. et al. Healthy dynamics of CD4 T cells may drive HIV resurgence in perinatally-infected infants on antiretroviral therapy. PLOS Pathog. 18, e1010751 (2022).
Schröter, J. & de Boer, R. J. What explains the poor contraction of the viral load during paediatric HIV infection?. J. Theor. Biol. 570, 111521 (2023).
Singh, A. Stochastic analysis of genetic feedback circuit controlling HIV cell-fate decision. in Proc. IEEE 51st IEEE Conference on Decision and Control (CDC)4918–4923. https://doi.org/10.1109/cdc.2012.6425836 (IEEE, 2012).
Jordan, A., Defechereux, P. & Verdin, E. The site of HIV-1 integration in the human genome determines basal transcriptional activity and response to Tat transactivation. EMBO J. 20, 1726–1738 (2001).
Jordan, A., Bisgrove, D. & Verdin, E. HIV reproducibly establishes a latent infection after acute infection of T cells in vitro. EMBO J. 22, 1868–1877 (2003).
Acknowledgements
This work was funded by NIH PAVE, a Martin Delaney Collaboratory (Grant Number: UM1AI164566). This work was also supported by the Italian Ministry of Health with Current Research funds.
Author information
Authors and Affiliations
Contributions
G.R.: drafted and finilized all manuscript materials, including the main text, figures, and tables. E.E., J.M.C., N.C., and P.P.: reviewed the manuscript, provided feedback, and approved the final version.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rasi, G., Emili, E., Conway, J.M. et al. Mathematical modeling and mechanisms of HIV latency for personalized anti latency therapies. npj Syst Biol Appl 11, 64 (2025). https://doi.org/10.1038/s41540-025-00538-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41540-025-00538-6
