
Abstract
Molecular hydrogen is the electron donor for the ancient exergonic reductive acetyl-coenzyme A pathway (acetyl-CoA pathway), which is used by hydrogenotrophic methanogenic archaea. How the presence of iron-sulfides influenced the acetyl-CoA pathway under primordial early Earth geochemistry is still poorly understood. Here we show that the iron-sulfides mackinawite (FeS) and greigite (Fe3S4), which formed in chemical garden experiments simulating geochemical conditions of the early Archaean eon (4.0–3.6 billion years ago), produce abiotic H2 in sufficient quantities to support hydrogenotrophic growth of the hyperthermophilic methanogen Methanocaldococcus jannaschii. Abiotic H2 from iron-sulfide formation promoted CO2 fixation and methanogenesis and induced overexpression of genes encoding the acetyl-CoA pathway. We demonstrate that H2 from iron-sulfide precipitation under simulated early Earth hydrothermal geochemistry fuels a H2-dependent primordial metabolism.
Similar content being viewed by others

Main
Formation of iron-sulfide minerals is an ancient and globally widespread process known to proceed via mackinawite (FeS)1 and the intermediate greigite (Fe3S4) (refs. 2,3) while producing H2 (refs. 4,5,6,7). The ancient occurrence of hydrothermal iron-sulfide rich deposits in the geological record extend into the early Archaean Eon (Eoarchaean 4.0–3.6 billion years ago) and exhibit fossil features interpreted as some of the oldest signatures for life on Earth8,9. Chemical garden experiments have shown the mackinawite–greigite transition under hydrothermal conditions analogous to Eoarchaean geochemical environments7,10. Such iron-sulfide chemical gardens are mixtures of inorganic chemicals which often form precipitates resembling organic structures and have been applied in emergence of life studies to better understand connections between aqueous geochemistry and mineralogy in simulated hydrothermal springs10,11. However, links between abiotic H2 production in iron-sulfide chemical gardens simulating Eoarchaean hydrothermal systems and early life are scarce.
Molecular H2 can be produced abiotically in numerous geological environments, most notably by serpentinization involving mafic (Mg and Fe-rich) or ultramafic rocks12,13. Since their discovery in 197714, seafloor hydrothermal springs venting abiotic H2 were viewed as potential energy sources and setting for the emergence of life15,16,17,18,19. Reactive iron-sulfide minerals are ubiquitous in hydrothermal vent black-smoker chimneys, which are surrounded by metalliferous sediments underlying the hydrothermal plumes of precipitated black iron-sulfide particles15,20,21. Modern black smokers can exhibit natural variations in fluid chemistry and temperature within a single vent system22, providing temperature gradients and sources of chemical energy15,16,21 to support a high diversity of microorganisms, including methanogenic archaea20. The availability of H2 plays a key role as the main source of electrons for biological systems and thereby microbial growth at hydrothermal systems23,24.
Models of early metabolism in the Eoarchaean predict that abiotic H2 was a potentially important electron donor and CO2 served as a key electron acceptor for the first cells25,26,27. Anaerobic organisms that use the H2-dependent reductive acetyl-coenzyme A (acetyl-CoA) pathway for CO2 fixation, such as methanogens and acetogens, are modern representatives that have preserved vestiges of the first metabolisms28,29. Geochemical evidence has shown that methanogenic microbes have been present since the early Archaean at least 3.5 billion years ago29, and it has been proposed that methanogens could have emerged in an iron-sulfide hydrothermal black-smoker environment16. Furthermore, physiological predictions of the last universal common ancestor (LUCA) based on genome data indicated that LUCA was possibly (hyper)thermophilic30, anaerobic and H2-dependent31, and used the acetyl-CoA pathway to fix CO2 (refs. 27,31,32,33,34). The acetyl-CoA pathway is the most ancient among carbon-fixation pathways35, as it is short, non-cyclic, exergonic33,35,36 and the only carbon-fixation pathway that can be reconstituted in the laboratory to take place without enzymes25,37.
The acetyl-CoA pathway is furthermore replete with enzymes that depend on simple Fe(Ni)S cofactors27,31,33,36,38. For example, the CO2 reducing-and-fixing enzyme formylmethanofuran dehydrogenase in methanogens contains an anomalously high 46 [4Fe–4S] clusters39. Other enzymes such as ferredoxin and CO dehydrogenase, which are common to methanogens, have iron-sulfur clusters with similar structure to iron-sulfide minerals, such as mackinawite and greigite18,36, which form in hydrothermal environments. These observations have led to the hypothesis that iron-sulfide clusters in these modern enzymes might be relics of an ancient metabolism, which possibly originated in an ancient iron-sulfide rich setting31,40.
These data and observations suggest an ancient origin of the acetyl-CoA pathway as a potential primordial metabolism25,32,33,34,35,41. Because of the possibility that hyperthermophilic methanogens harbour one of the most ancient metabolisms30, we investigated acetyl-CoA pathway-dependent growth of the methanogenic hyperthermophile Methanocaldococcus jannaschii during iron-sulfide mineral formation.
Ferruginous (anoxic and iron-rich42) Archaean oceans contained dissolved ferrous iron concentrations of 0.1–10 mM (refs. 43,44). Sulfide may have been relatively rare as shown by its apparent dearth in the Hadean and Eoarchaean, but the origin of sulfide could have been derived from volcanic sulfur dioxide and polymerized sulfides45,46. In the absence of sulfidic fluids, green rust was probably a major constituent of Hadean and Archaean chemical gardens under ferruginous conditions47,48. Sulfate was extremely rare in the Archaean oceans49, and the salinity has been predicted to be similar50 if not greater51 than now found in ocean water. We attempted to reflect these ancient conditions of the Eoarchaean oceans in our chemical gardens (Supplementary Methods).
As our model organism, we chose the methanogenic hyperthermophile M. jannaschii (DSM strain 2661), which was originally isolated from hydrothermal iron-sulfide sediment at the base of a black smoker on the East Pacific Rise52. M. jannaschii serves as a model organism for methanogenesis53,54,55; it uses H2 and CO2 as its sole carbon and energy source and has an optimal growth temperature ~85 °C (refs. 52). Thus, tracing both H2 and CO2 and the product CH4 can reveal the active metabolism of this organism in controlled experiments, which is advantageous, because only a small amount of substrates and products need to be measured to study the energy metabolism.
Results and discussion
Iron-sulfide chimney formation, mineralogy and habitability
To precipitate iron-sulfide, we created sulfidic precipitation mounds (experiment 1, Supplementary Table 1) within a ferruginous aqueous environment germane to hydrochemical conditions predicted for the Hadean Ocean at the time of the origin of life7,10,18. Acidic sulfidic solution (0.5 M Na2S, pH 3) was injected into an amorphous ferruginous solution (0.5 M Fe(II)Cl2, pH 6) (chimney protocol in Supplementary Methods). Within 10 min, iron-sulfide minerals crystallized forming a black-chimney structure reaching an average height of 2.0 cm (s.d. = 0.1). In this initial phase of the experiment, the chimney showed the most rapid growth (growth rate = 0.2 cm min−1). After 1 hour, the average height of the chimney was 2.5 cm (s.d. = 0.1, growth rate = 0.01 cm min−1). No growth of the chimney was observed after 1 hour. All experiments were performed under an anoxic N2 atmosphere in an anoxic chamber. Additional information about imaging, processing and quantifying the chimney growth is displayed in the Supplementary Methods.
The chimneys were stable at room temperature. Upon heating of the injection fluid, however, bubbles formed in the sulfidic fluid and caused the chimney structures to collapse into sediment, which collected at the bottom of the flask. Therefore, all hydrothermal experiments at 80 °C were conducted using sedimented iron-sulfide material (for details of preparation, see experiments 2, 3, 4 and 5 in Supplementary Table 1), which we describe as ‘sedimentary iron-sulfide chemical gardens’. Future experiments could test if higher pressure (similar to the deep sea) would increase the boiling temperature of the injected sulfidic fluid and reduce formation of gas bubbles.
Raman spectroscopy and scanning electron microscopy (SEM) revealed that the hydrothermal (80 °C) sedimentary iron-sulfide chemical gardens (experiment 5a, Supplementary Table 1) comprised mostly mackinawite (FeS) and greigite (Fe3S4), with traces of NaCl (Fig. 1c–f). Under anoxic conditions, metastable mackinawite, an iron-monosulfide precursor, transforms into greigite when heated to 70–75 °C through partial oxidation of mackinawite with water2,7.

a, Iron-sulfide chimney precipitated over the course of 10 min at 25 °C (experiment 1, Supplementary Table 1). The three images (from left to right) were taken 1, 5 and 10 min after the injection started. After 10 min, the chimney reached a height of 2.0 cm (s.d. = 0.1). b, M. jannaschii cells accumulate on iron-sulfide particles (experiment 4a). Overlay of a bright field image of the iron-sulfide particles and a fluorescence image of the cells. This was observed across three experimental replicates (Methods). c–f, Mineralogical analysis (experiment 5a). c,d, Raman spectroscopy identifies that mackinawite (FeS, c) and greigite (Fe3S4, d) are the primary minerals formed in the sedimentary iron-sulfide chemical gardens at 80 °C. This was observed across three experimental replicates (Supplementary Table 1). Important Raman peaks are labelled for mackinawite and greigite7,57,58,59. The peak at 1,300 cm−1 is due to laser beam oxidation. e, Raman images of a sedimentary iron-sulfide chemical garden. Most crystals have an octahedral shape typical for greigite. f, EDX of iron-sulfide particles with peaks indicating relative abundances of O, Na, S, Cl and Fe. keV, kiloelectron volt.
Mackinawite is a tetragonal iron-monosulfide1. Greigite is the cubic thio-spinel of iron, exhibiting mostly octahedral and sometimes cubic crystals2,3,56. Next to the Raman peaks for mackinawite at 212, 275–276, 385–386 and 583–584 cm−1 (Fig. 1c)57,58 a broad peak at 1,300 cm−1 was evident of slight laser beam oxidation of the sample, which converted mackinawite into haematite (Fe2O3). The greigite Raman spectra (Fig. 1d) show peaks at 191, 250, 299–301 and 350–354 cm−1 (refs. 7,59). The Raman images display an octahedral crystal morphology fitting for greigite3 (Fig. 1e). The energy dispersive X-ray (EDX) spectrum highlights the presence of iron (30.78 at.%) and sulfur (41.33 at.%) in the sample (Fig. 1f). Magnetic properties of the iron-sulfide minerals from the chemical garden (experiment 5a, Supplementary Table 1) was confirmed with a 96-ring magnet plate (ALPAQUA) (Extended Data Fig. 1), indicating the presence of greigite, which is a ferrimagnetic mineral2,56,60,61. These results are consistent with earlier chemical garden experiments7, which also produced semipermeable colloidal membranes of iron-monosulfide minerals mimicking natural hydrothermal chimneys10.
Iron-sulfur clusters play a critical role in various metalloenzymes that are present in both archaea and bacteria7, and therefore mackinawite and greigite are considered key minerals for the emergence of life7,40,59,62 because they structurally resemble active centres of proteins and enzymes36,59,63,64. The presence of both minerals in hydrothermal environments hints at the possibility that iron-rich cofactors in proteins and enzymes geologically originate from mineral precursors63. It is therefore notable that mackinawite and greigite are the major precipitated minerals in our chemical gardens simulating sulfidic Eoarchaean hydrothermal springs (Fig. 1d). We note that the pyrite structure is different because the ferrous iron is ligated to six sulfur pairs, which makes the mineral less reactive by comparison65.
To test the habitability of the sedimentary iron-sulfide chemical gardens, we added stationary-phase culture of the hyperthermophilic methanogen M. jannaschii52 to the chemical gardens (experiment 4a, Supplementary Table 1). Fluorescence microscopy revealed M. jannaschii cells physically associated with iron-sulfide particles (Fig. 1b), consistent with minerals found in black smokers that are colonized by thermophilic archaea66,67. The pH of the sedimentary chemical gardens was 5.5, which is close to the optimal pH for growth of M. jannaschii68. Next, we investigated mechanisms linking the mineralogy of the iron-sulfide chemical gardens to microbial habitability.
Abiotic H2 production and methanogenesis in chemical gardens
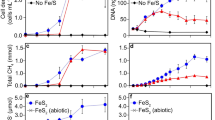
We detected hydrogen gas (H2) emanating from the sedimentary iron-sulfide chemical gardens (experiment 2, Supplementary Table 1) in amounts proportional to the concentrations of Fe(II) and Na2S (Fig. 2a). Abiotic H2 was only detectable in flasks where no headspace was present (Supplementary Methods), which could be due to increasing pressure building without a larger headspace volume. The concentration of abiotic H2 produced in the chemical gardens ranged from 124 ± 1 to 781 ± 138 µM, spanning Fe(II) and Na2S concentrations of 10–500 mM, respectively (P = 0.0206) (Fig. 2a). This is in the range of H2 concentrations in the fluids of black smokers, from 0.05 to 1 mM (ref. 12). Abiotic H2 production is probably explained by the presence of mackinawite (FeS) and greigite (Fe3S4) above 70 °C (Fig. 1c,d), namely that H2 is released during the hydration of iron-monosulfide6,7,69 according to:

a, Production of abiotic H2 in the chemical gardens increases with increasing Fe(II) concentrations at 80 °C (experiment 2, Supplementary Table 1). Error bars indicate standard error of the means, individual points represent biological replicates (n = 3). b, The concentration of abiotic H2 produced in the sedimentary iron-sulfide chemical garden (500 mM FeCl2 and Na2S, 80 °C) correlates positively with the amount of 13CH4 produced by M. jannaschii. c, Gas consumption and production by M. jannaschii in the sedimentary iron-sulfide chemical gardens at 80 °C supplemented with 13C-bicarbonate to trace CO2 fixation and methanogenesis (experiment 3). Error bars represent standard deviations across three biological replicates. Note that the 13C-labelling of CO2 decreases over time, as the 13C-labelling of methane increases, indicating a transfer of 13C from CO2 to CH4 by M. jannaschii. The abiotically produced H2 increases over the first 4 h and then decreases afterwards, due to consumption by M. jannaschii.
A similar reaction of mackinawite and H2S was found to be used in an anaerobic metabolism by H2-producing bacteria in a syntrophic partnership with hydrogenotrophic methanogens70. Addition of stationary-phase M. jannaschii culture to the chemical garden reduced abiotic H2 significantly compared to controls (P = 0.0001) without added methanogens (Extended Data Fig. 2), indicating that the hydrogenotrophic M. jannaschii cells consume H2 for growth. Others54 defined the lower limit for growth at 17–23 µM of H2, which is less H2 than the iron-sulfide chemical gardens produced (experiment 2, Fig. 2a). No H2 was detectable in controls where FeCl2 and Na2S were not added. We investigated the effects of abiotic H2 from the sedimentary chemical gardens on the physiology of M. jannaschii (experiment 3, Fig. 2c) by tracking the production of H2 and 13CH4 at 80 °C (experiment 3, Supplementary Table 1) using 13CO2 carbon substrate. After inoculation, an increase in 13CH4 was observed concomitant with a decrease in 13CO2, as expected for M. jannaschii methanogenesis (Fig. 2c). Over the course of 10 h, 13C-labelled CO2 decreased constantly, while abiotic H2 increased in concentration until 4 h, after which the rate of H2 consumption by M. jannaschii began to outpace H2 production. We observed a direct correlation (R2 = 0.97, P < 0.005) between abiotic H2 produced by the chemical garden and 13C-labelled CH4 produced by M. jannaschii (Fig. 2b) indicating that, consistent with earlier studies54,55, hydrogenotrophic methanogenesis by M. jannaschii in the iron-sulfide chemical gardens was limited by H2.
M. jannaschii growth in iron-sulfide chemical gardens
We compared the growth of M. jannaschii at 80 °C in the iron-sulfide chemical garden (experiment 5a) to growth in MMC medium (experiment 5c) as a positive control and sterile water (experiment 5b) as a negative control. Similar to experiment 4, the sterile water negative control was used to account for the dilution factor introduced by inoculating the stationary-phase culture into the chemical garden, making the presence of iron-sulfide the only difference between experiments 5a and 5b (Supplementary Table 1).
M. jannaschii reached exponential growth (growth rate µ = 0.14 h−1) in the iron-sulfide chemical gardens after 24 h reaching a peak cell concentration of 2.3 × 107 cells ml−1 (Fig. 3a) (experiment 4a, Supplementary Table 1). However, the growth rate in the chemical gardens (experiment 4a) was 30% lower (P = 0.0424) than the positive controls (µ = 0.20 h−1), where M. jannaschii was grown in their standard MMC growth medium (experiment 4c) and reached a peak cell concentration of 3.0 × 107 cells ml−1, lower than previous reports52 possibly because we did not shake cultures during incubation. The lower growth rate in the iron-sulfide chemical gardens (Fig. 3a) could be explained by the lack of additional salts, trace metals (for example, Ni, Mo, Co, W and Zn) and nutrients compared to MMC medium, where nitrogen in the form of ammonia was present at tenfold higher concentrations68. Traces of nitrogen and other essential metals were inevitably transferred with the inoculum to the chemical gardens and were probably available—albeit at ultra-low concentrations—for growth (Fig. 3a).

a, Comparison of M. jannaschii growth in three experimental settings: sedimentary iron-sulfide chemical gardens (experiment 4a, Supplementary Table 1), MMC growth medium (experiment 4c) and sterile water (experiment 4b). All experiments were performed at 80 °C, error bars indicate standard error of the means across three biological replicates. b, Principal-component (PC) analysis shows the significant difference of M. jannaschii gene expression in non-identical treatments (experiment 5) (analysis of similarity P = 0.001). Grey circles highlight transcriptomes from 25 and 80 °C experiments, respectively. Coloured symbols correspond to the same three experimental conditions that are displayed in a (yellow diamonds, MMC media control; orange triangles, iron-sulfide chemical garden; blue squares, sterile water negative control).
No growth was observed in the negative (water) control (experiment 4b) (Fig. 3a). Because the addition of Fe(II)Cl2 and Na2S are the only parameters that differed compared to the control (additional nutrients, vitamins or trace metals above those present in the inoculum were not added to the chemical gardens), exponential growth in the chemical gardens can be attributed to H2 from the reaction of FeCl2 and Na2S (Fig. 2c and Extended Data Fig. 2). Nitrogen limitation cannot explain the large difference in growth rate between the water control and the chemical garden, because both experiments received the same amount of inoculum. This indicates that the growth in the chemical gardens was mainly limited by H2 compared to nitrogen, indicating that the abiotic H2 from the chemical garden was sufficient enough to rescue M. jannaschii from H2 limitation.
Gene expression in iron-sulfide chemical gardens
RNA was extracted from the 80 °C sterile water control (experiment 5b), 10 mM iron-sulfide chemical garden (experiment 5a) and MMC medium (experiment 5c), which was determined to be 2.52, 2.90 and 10.00 ng µl−1, respectively (Supplementary Methods). This RNA was used for transcriptomic analysis of M. jannaschii, which showed that gene expression was significantly different between the three 80 °C treatments (10 mM iron-sulfide, MMC medium and sterile water, experiments 5a–c, Supplementary Table 1), indicating that each condition was associated with different physiological states (Fig. 3b). The sedimentary iron-sulfide chemical garden transcriptomes uncovered biological (analysis of similarity P = 0.001) and technical variation in between replicates exceeding that in controls (Fig. 3b), but nonetheless show a reproducible and unique gene expression profile induced by iron-sulfide chemical gardens. Key genes that were overexpressed by M. jannaschii in the chemical gardens reflect the H2-dependent nature of M. jannaschii metabolism.
In contrast to the MMC medium, 120 genes were downregulated in M. jannaschii grown in sedimentary iron-sulfide chemical gardens, whereas 57 genes were downregulated in M. jannaschii cells grown in chemical gardens compared to the water controls (experiment 5, Fig. 4 and Supplementary Table 1). However, numerous genes encoding key enzymes of the H2-dependent and CO2-fixing acetyl-CoA pathway for methanogenesis were overexpressed in cells grown in chemical gardens when compared to the controls (Fig. 4 and Supplementary Tables 2 and 3). A total of 34 and 46 genes were overexpressed in cells grown in chemical gardens as compared to MMC medium (Fig. 4a and Supplementary Table 2) and water (Fig. 4b and Supplementary Table 3), respectively. The conditions of the chemical garden promoted the expression of the acetyl-CoA pathway in M. jannaschii compared to other cellular processes. There were 96 genes upregulated and 241 genes downregulated in M. jannaschii cells grown in MMC medium at the optimal 80 °C compared to MMC media at 25 °C (Extended Data Fig. 3).

Gene expression is compared to two sets of controls (experiments 5a–c, Supplementary Table 1). a, Significant overexpression of genes (n = 34, P < 0.05) in M. jannaschii transcriptomes from 80 °C iron-sulfide chemical gardens (red, experiment 5a) relative to the MMC medium (beige, experiment 5c) (P = 0.05, one-sided t-test). b, Significant overexpression of genes (n = 46, P < 0.05) in M. jannaschii transcriptomes from 80 °C iron-sulfide chemical gardens (red, experiment 5a) relative to sterile water negative controls (blue, experiment 5b) (P = 0.05, one-sided t-test). The vertical line separates expressed genes that either increased or decreased in rank in the chemical gardens, the dotted horizontal line represents the P value cutoff for determining statistical significance. Overexpressed genes in the chemical gardens that are part of the methanogenic acetyl-CoA pathway are labelled. The full spellings of the gene acronyms of the acetyl-CoA pathway are provided in Supplementary Tables 1 and 2. Note that the genes of the methanogenic acetyl-CoA pathway were only overexpressed in the iron-sulfide chemical gardens, and were overexpressed relative to the MMC medium and sterile water negative controls.
The overexpressed genes of the methanogenic acetyl-CoA pathway (Fig. 4 and Supplementary Tables 2 and 3) consisted of CO dehydrogenase/CO-methylating acetyl-CoA synthase complex subunit beta (CODH/ACS), methyl-coenzyme M (methyl-COM) reductase subunits alpha/beta/gamma (mcrA, mcrB and mcrG), methyl-COM reductase operon proteins C/D (mcrC and mcrD), tetrahydromethanopterin S-methyltransferase subunits A–H (mtrA–H), formylmethanofuran dehydrogenase (fmdA, fmdB and fmdC), formylmethanofuran-tetrahydromethanopterin N-formyltransferase (ftr), acetyl-CoA decarbonylase/synthase complex subunits alpha/beta/gamma (ACDS) and coenzyme B–coenzyme M (CoB–CoM) heterodisulfide reductase subunits B/C (hdrB and hdrC). With the exception of only four genes (mcrC, mcrD, ftr and hdrB,C) all other acetyl-CoA pathway genes were overexpressed in the sedimentary iron-sulfide chemical gardens compared to both sterile water and MMC medium controls (Fig. 4). This shows that the chemical gardens were generally associated with a stimulated activity of the acetyl-CoA pathway in M. jannaschii, even compared to the growth conditions of the MMC medium, which was optimized for growth of hyperthermophilic methanogens66 (Fig. 3a).
Methanogenesis requires several carbon carriers for the reduction of CO2 to CH4, which are the coenzymes methanofuran, tetrahydromethanopterin and CoM18,71. CoB is not a carbon carrier but is also required in the terminal step of methanogenesis71. Next to methane production through CO2 fixation, the acetyl-CoA pathway also produces the metabolic intermediate acetyl-CoA, which is for example a precursor to lipids and pyruvate72. The key multi-enzyme complex involved in the acetyl-CoA formation is CODH/ACS36,41 where CODH catalyses the CO2 reduction to CO and ACS catalyses the C–C bond formation from CO and a methyl group to form acetyl-CoA73. This process allows methanogens to use CO as a carbon source. The multi-enzyme complex ACDS, which was also overexpressed in our sedimentary iron-sulfide chemical gardens (Fig. 4), catalyses the reversible cleavage of acetyl-CoA in methanogens74. Electrons for the methanogenic acetyl-CoA pathway primarily come from H2 (ref. 71). Considering that the genes responsible for the enzymatic conversions for most of these steps were overexpressed in the iron-sulfide chemical gardens, it appears that this chemolithoautotrophic pathway in M. jannaschii was probably driven by the abiotic H2 released during the formation of mackinawite and greigite in the chemical gardens.
Interestingly, under the non-growing physiological state in sterile H2O we observed an overexpression of some genes that encode enzymes of the acetyl-CoA pathway when compared to growth in MMC medium only (Extended Data Fig. 4 and Supplementary Table 4). This might indicate that non-growing archaeal cells experiencing H2 limitation and/or nutrient limitation can maintain a physiological readiness to quickly react to suddenly appearing favourable growth conditions in a hydrothermal environment66,75.
The key enzyme complex CODH/ACS (Fig. 4), which is necessary to make cell carbon18, contains metal sulfur clusters affine with structure iron-sulfide minerals including greigite36,64. Although CODH/ACS was overexpressed when cultivated in sterile H2O controls compared to MMC medium (Extended Data Fig. 4), the overexpression was six times higher in the chemical gardens (Fig. 4) by comparison. Specifically, CODH/ACS transcript levels had a fold-increase of 11.6 (one-sided t-test, P = 0.0014) in the iron-sulfide chemical garden relative to the sterile H2O, compared to 1.9-fold increase (one-sided t-test, P = 0.0007) in sterile H2O compared to MMC medium. This shows that while CODH/ACS is slightly overexpressed in a non-growing state, once the cells are placed into the iron-sulfide chemical garden, where they are alleviated from H2 limitation, they increase the expression of CODH/ACS even more as well as other genes of the acetyl-CoA pathway. This further supports the conclusion that the cells remain physiologically ready for growth even if the cells are temporarily in a non-growing state.
Overexpression of fmd, ftr and mtrA–H in the iron-sulfide chemical gardens is notable considering that they are part of the first CO2-reduction steps that take place in the acetyl-CoA pathway71. This is consistent with the stable isotope labelling experiment showing fixation of 13CO2 by M. jannaschii in the iron-sulfide chemical gardens (Fig. 2c). Interestingly, mtrA–H, fmd and ftr were only overexpressed in cells cultivated in chemical gardens (Fig. 4), but not in the sterile water controls compared to MMC medium (Extended Data Fig. 4 and Supplementary Table 4), indicating an enhancement of the CO2-reduction step by iron-sulfide conditions. This suggests that the iron-sulfide chemical garden promoted carbon fixation by M. jannaschii, which is consistent with the exponential growth observed in the iron-sulfide chemical gardens (Fig. 3a). No additional bicarbonate was added to the iron-sulfide chemical gardens and therefore some of the overexpression of fmd, ftr or mtrA–H relative to MMC medium could be a response to lower CO2 levels and carbon limitation. However, this cannot explain the overexpression of genes in these cells compared to the sterile H2O controls (because they also did not receive additional bicarbonate). Thus, it seems likely that the iron-sulfide chemical garden is a major factor influencing the overexpression of these genes.
Mcr catalyses the reduction of methyl-CoM with CoB to methane in the last step of the acetyl-CoA pathway, which is coupled to the formation of the heterodisulfide (CoM–S–S–CoB)32,76,77. Genes encoding all subunits of the mcr protein were overexpressed in the iron-sulfide chemical gardens relative to MMC medium and sterile H2O controls (Fig. 4 and Supplementary Tables 2 and 3). In contrast, only the beta subunit was overexpressed in the sterile H2O control compared to MMC medium and its expression level was comparably lower (Supplementary Table 4). We also detected overexpression of two mcr operon proteins D and G in the iron-sulfide chemical garden relative to MMC medium.
The heterodisulfide reductase gene (hdr) was only overexpressed in an active growing state compared to a non-active growing state (iron-sulfide chemical garden relative to sterile H2O, Fig. 4b). It was not overexpressed when comparing actively growing states to one another, specifically comparing the iron-sulfide chemical gardens to the MMC medium (Fig. 4a). This shows that hdr was important during active growth and not during dormancy. Hdr is an important iron-sulfur protein in the acetyl-CoA pathway that catalyses the reversible reduction of the heterodisulfide (CoM–S–S–CoB) back to CoM–SH and CoB–SH with H2 leading to the formation of methane76,78. The overexpression of these acetyl-CoA pathway genes mcr and hdr is furthermore consistent with the stable isotope labelling of 13CH4 which was produced by M. jannaschii in the iron-sulfide chemical gardens (Fig. 2c). M. jannaschii overexpressed a disproportionally higher percentage of genes assigned to archaeal clusters of orthologous genes (arCOGs)79 involved in energy production and conversion and translation in iron-sulfide chemical gardens compared to the controls (Extended Data Fig. 5). All these results indicate that cellular control of the acetyl-CoA pathway responsible for energy conservation and CO2 fixation was overexpressed in the chemical gardens (Fig. 4a). These experimental results support metabolic and evolutionary models based on phylogenomic reconstructions of LUCA, indicating that the acetyl-CoA pathway was present in LUCA, which might have lived in a hydrothermal environment31,34.
Furthermore, it has been determined that reduced iron can promote electron transfer from H2 to ferredoxin, which might have been important for cellular energy metabolism via the acetyl-CoA pathway before the emergence of enzymes and cofactors80. It therefore seems possible that reduced iron-sulfides could also facilitate electron transfer between H2 and ferredoxin in M. jannaschii, which would help to explain how M. jannaschii can achieve exponential growth in the iron-sulfide chemical garden.
We compared the effect of temperature to iron-sulfide environments as a factor that might influence the overexpression of genes involved in the acetyl-CoA pathway. Gene expression of M. jannaschii at 80 and 25 °C (experiments 5c,d and Supplementary Table 1) in MMC medium was different (Fig. 3b). Temperature is known to have a strong effect on the gene expression of thermophilic archaea75. In our experiments, a total of 96 genes were overexpressed at 80 °C compared to 25 °C (experiment 5, Supplementary Table 1), with two of the overexpressed genes indirectly involved in regulation of the methanogenic acetyl-CoA pathway (Extended Data Fig. 3). These include methyl-COM reductase system component A2 (atwA), which is an ATP-binding protein81, and an NADPH-dependent F420 reductase (npdG). Despite M. jannaschii being a hyperthermophile, the number of overexpressed acetyl-CoA pathway-associated genes at high temperature (n = 2) was lower compared to the iron-sulfide experiments (n = 11). This indicates that the iron-sulfide chemical garden environment had a larger positive influence on the activity of the H2-dependent acetyl-CoA pathway metabolism relative to temperature.
While the genes encoding the acetyl-CoA pathway in the sedimentary iron-sulfide chemical garden were overexpressed relative to MMC medium, the growth rate of M. jannaschii was lower in the chemical gardens by comparison (Fig. 3a). The slower growth rate could be explained by the presence of additional bicarbonate (the carbon source), micronutrients, vitamins and trace metals supplied in the MMC medium, and excluded in the iron-sulfide chemical gardens (which only received FeCl2 and Na2S).
To conclude, the physiology of many methanogenic archaea is defined by H2-dependent syntrophy, where H2 is produced by a partner organism82. This relationship, whereby both partner organisms live together in a metabolic partnership, is considered to have an ancient origin83. Our results suggest, that on the early Eoarchaean Earth abiotic iron-sulfide geochemistry could have replaced H2-producing syntrophic partner organisms and provide an abiotic H2 source for H2-dependent methanogenic archaea. This could have been an ancient precursor to modern syntrophic partnerships, whereby H2 is sourced from biological fermentations.
Outlook
We provide experimental testing of a key aspect of hydrothermal and iron-sulfide-rich environments for the emergence of life, namely that abiotic H2 produced in a sedimentary iron-sulfide chemical garden could have provided sufficient energy to fuel the survival and growth of Archaea in an Eoarchaean hydrothermal environment. Our results show that abiotic H2 produced by iron-sulfide mineral redox reactions is sufficient to promote exponential chemolithoautotrophic growth of a hyperthermophilic methanogen under ferruginous conditions. This physiological response was explained by the overexpression of acetyl-CoA pathway-encoding genes by M. jannaschii in the sedimentary iron-sulfide chemical gardens and an exergonic CO2-fixation pathway that has been described as a ‘free lunch that you are paid to eat’84. Our findings provide experimental support for theories36,85,86 predicting that the extreme and energy-limited conditions of iron-sulfide-rich environments of the Eoarchaean would have promoted an acetyl-CoA pathway-based metabolism. Our study points to FeS–Fe3S4 chemical gardens as potential hatcheries of life, primordial environments that could theoretically support a continuous evolution of the first metabolizing cells, through the progenote, to a methanogen.
Methods
The hyperthermophilic methanogen M. jannaschii52 (DSM strain 2661, German Collection of Microorganisms and Cell Cultures GmbH) was cultivated in an MMC growth medium68, which was prepared at the Institute of Microbiology and German Archaea Centre at the University of Regensburg (Supplementary Methods). The strain was recultivated from the Bacteria Bank Regensburg and adopted to the medium by at least two serial transfers in MMC. The stationary-phase cell cultures were used to inoculate the experiments that are part of this study.
In total, five different sets of experiments were executed (Supplementary Table 1). Throughout our paper we refer to these experiments as: experiment 1, chimney formation (Fig. 1a); experiment 2, abiotic H2 formation (Fig. 2a and Extended Data Fig. 2); experiment 3, stable isotope labelling (Fig. 2b,c); experiment 4, M. jannaschii colonization and growth curve (Figs. 1b and 3a); and experiment 5, transcriptomics (Figs. 3b and 4 and Extended Data Figs. 3–5) and mineralogical analysis (Fig. 1c–f and Extended Data Fig. 1).
A detailed chimney formation protocol for experiment 1 is in Supplementary Methods. In experiment 2 and 3 (Supplementary Table 1), gas measurements of 13CO2, 13CH4 and H2 were performed using a GC-MS QP2020 NX connected to a headspace autosampler (Shimadzu) (see Supplementary Methods for protocol).
In experiment 4 (Supplementary Table 1) we compared the growth of M. jannaschii in the sedimentary iron-sulfide chemical garden at 80 °C (experiment 4a) to its growth in MMC medium (experiment 4c) as a positive control. As an additional negative control, we also measured the growth of M. jannaschii in sterile water (experiment 4b). The sterile water negative control accounted for the dilution factor introduced in the chemical garden, making the presence of iron-sulfide the only difference between both experiments 4a and 4b. Using the chemical garden from experiment 4a we tested the colonization of M. jannaschii on the iron-sulfide particles (Fig. 1b). Cell counts and particle attachment were visualized using an inverted fluorescence microscope (Leica Thunder Imager DMi) based on autofluorescence of the coenzyme F420 present in M. jannaschii87.
In experiment 5 (Supplementary Table 1) we compared the transcriptomic response of M. jannaschii at 80 °C in the sedimentary iron-sulfide chemical garden (experiment 5a) to the gene expression in MMC medium (experiment 5c) as a positive control and sterile water (experiment 5b) as a negative control. Furthermore, we performed transcriptomes on stationary-phase cultures stored at 25 °C (experiment 5d) as an additional low temperature comparison. Similar to experiment 4 the sterile water negative control was used to account for the dilution factor introduced by inoculating the stationary-phase culture into the chemical garden, making the presence of iron-sulfide the only difference between experiments 5a and 5b.
In experiment 5, RNA was extracted using the Direct-zol RNA Microprep kit (ZYMO Research)75, with several changes to the protocol to improve RNA extraction from the chemical gardens. Phosphate was added to reduce adsorbtion of RNA to the iron-sulfide minerals and chloroform was added to improve RNA recovery (Supplementary Methods). Transcriptomes were prepared using the Revelo RNA-Seq kit (Tecan) and raw reads were mapped against the annotated genome of M. jannaschii88 using BLASTx with DIAMOND89 to measure gene expression levels.
The mineralogy of sedimentary iron-sulfide chemical gardens (experiment 5a, Supplementary Table 1) was analysed using Raman spectroscopy according to previously reported methods47 and SEM with an EDX detector (protocol in Supplementary Methods). Full details on transcriptome preparations, bioinformatic analysis, cell counts, gas analysis, Raman spectroscopy, EDX analysis and MMC medium preparations are provided in Supplementary Methods90,91.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
Transcriptome data have been deposited in the NCBI short read archive under Bioproject ID PRJNA1157004.
Code availability
All code used in this study is available via GitHub at https://github.com/williamorsi/bioinformatics_code_orsi.
References
Lennie, A. R., Redfern, S. A., Schofield, P. F. & Vaughan, D. J. Synthesis and Rietveld crystal structure refinement of mackinawite, tetragonal FeS. Mineral. Mag. 59, 677–683 (1995).
Hunger, S. & Benning, L. G. Greigite: a true intermediate on the polysulfide pathway to pyrite. Geochem. Trans. 8, 1 (2007).
Lennie, A. R. et al. Transformation of mackinawite to greigite: an in situ X-ray powder diffraction and transmission electron microscope study. Am. Mineral. 82, 302–309 (1997).
Drobner, E., Huber, H., Wächtershäuser, G., Rose, D. & Stetter, K. O. Pyrite formation linked with hydrogen evolution under anaerobic conditions. Nature 346, 742–744 (1990).
Heinen, W. & Lauwers, A. M. Organic sulfur compounds resulting from the interaction of iron sulfide, hydrogen sulfide and carbon dioxide in an anaerobic aqueous environment. Orig. Life Evol. Biosph. 26, 131–150 (1996).
Russell, M. J. & Hall, A. The emergence of life from iron monosulphide bubbles at a submarine hydrothermal redox and pH front. J. Geol. Soc. 154, 377–402 (1997).
White, L. M., Bhartia, R., Stucky, G. D., Kanik, I. & Russell, M. J. Mackinawite and greigite in ancient alkaline hydrothermal chimneys: identifying potential key catalysts for emergent life. Earth Planet. Sci. Lett. 430, 105–114 (2015).
Dodd, M. S. et al. Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 543, 60–64 (2017).
Papineau, D. et al. Metabolically diverse primordial microbial communities in Earth’s oldest seafloor-hydrothermal jasper. Sci. Adv. 8, eabm2296 (2022).
Russell, M. J., Hall, A. J. & Turner, D. In vitro growth of iron sulphide chimneys: possible culture chambers for origin‐of‐life experiments. Terra Nova 1, 238–241 (1989).
Barge, L. M. et al. From chemical gardens to chemobrionics. Chem. Rev. 115, 8652–8703 (2015).
Klein, F., Tarnas, J. D. & Bach, W. Abiotic sources of molecular hydrogen on Earth. Elements 16, 19–24 (2020).
Truche, L., McCollom, T. M. & Martinez, I. Hydrogen and abiotic hydrocarbons: molecules that change the world. Elements 16, 13–18 (2020).
Corliss, J. B. et al. Submarine thermal springs on the Galapagos Rift. Science 203, 1073–1083 (1979).
Baross, J. A. & Hoffman, S. E. Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life. Orig. Life Evol. Biosph. 15, 327–345 (1985).
Corliss, J., Baross, J. & Hoffman, S. An hypothesis concerning the relationship between submarine hot springs and the origin of life on Earth. Oceanol. Acta 80, 59–69 (1981).
Goldford, J. E., Hartman, H., Marsland, R. III & Segrè, D. Environmental boundary conditions for the origin of life converge to an organo-sulfur metabolism. Nat. Ecol. Evol. 3, 1715–1724 (2019).
Martin, W., Baross, J., Kelley, D. & Russell, M. J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 6, 805–814 (2008).
Yamamoto, M., Nakamura, R. & Takai, K. Deep‐sea hydrothermal fields as natural power plants. ChemElectroChem 5, 2162–2166 (2018).
Karson, J. A., Kelley, D. S., Fornari, D. J., Perfit, M. R. & Shank, T. M. Discovering the Deep: A Photographic Atlas of the Seafloor and Ocean Crust (Cambridge Univ. Press, 2015).
Li, Y., Kitadai, N. & Nakamura, R. Chemical diversity of metal sulfide minerals and its implications for the origin of life. Life 8, 46 (2018).
Dick, G. J. The microbiomes of deep-sea hydrothermal vents: distributed globally, shaped locally. Nat. Rev. Microbiol. 17, 271–283 (2019).
Petersen, J. M. et al. Hydrogen is an energy source for hydrothermal vent symbioses. Nature 476, 176–180 (2011).
Reveillaud, J. et al. Subseafloor microbial communities in hydrogen‐rich vent fluids from hydrothermal systems along the Mid‐Cayman Rise. Environ. Microbiol. 18, 1970–1987 (2016).
Preiner, M. et al. A hydrogen-dependent geochemical analogue of primordial carbon and energy metabolism. Nat. Ecol. Evol. 4, 534–542 (2020).
Russell, M. J., Hall, A. J. & Martin, W. Serpentinization as a source of energy at the origin of life. Geobiology 8, 355–371 (2010).
Lane, N., Allen, J. F. & Martin, W. How did LUCA make a living? Chemiosmosis in the origin of life. BioEssays 32, 271–280 (2010).
Woese, C. R. Bacterial evolution. Microbiol. Rev. 51, 221–271 (1987).
Ueno, Y., Yamada, K., Yoshida, N., Maruyama, S. & Isozaki, Y. Evidence from fluid inclusions for microbial methanogenesis in the early Archaean era. Nature 440, 516–519 (2006).
Ciccarelli, F. D. et al. Toward automatic reconstruction of a highly resolved tree of life. Science 311, 1283–1287 (2006).
Weiss, M. C. et al. The physiology and habitat of the last universal common ancestor. Nat. Microbiol. 1, 16116 (2016).
Borrel, G., Adam, P. S. & Gribaldo, S. Methanogenesis and the Wood–Ljungdahl pathway: an ancient, versatile, and fragile association. Genome Biol. Evol. 8, 1706–1711 (2016).
Varma, S. J., Muchowska, K. B., Chatelain, P. & Moran, J. Native iron reduces CO2 to intermediates and end-products of the acetyl-CoA pathway. Nat. Ecol. Evol. 2, 1019–1024 (2018).
Moody, E. R. et al. The nature of the last universal common ancestor and its impact on the early Earth system. Nat. Ecol. Evol. 8, 1654–1666 (2024).
Fuchs, G. Alternative pathways of carbon dioxide fixation: insights into the early evolution of life? Annu. Rev. Microbiol. 65, 631–658 (2011).
Russell, M. J. & Martin, W. The rocky roots of the acetyl-CoA pathway. Trends Biochem. Sci. 29, 358–363 (2004).
Martin, W. F. Older than genes: the acetyl CoA pathway and origins. Front. Microbiol. 11, 817 (2020).
Sojo, V., Herschy, B., Whicher, A., Camprubi, E. & Lane, N. The origin of life in alkaline hydrothermal vents. Astrobiology 16, 181–197 (2016).
Wagner, T., Ermler, U. & Shima, S. The methanogenic CO2 reducing-and-fixing enzyme is bifunctional and contains 46 [4Fe-4S] clusters. Science 354, 114–117 (2016).
Eck, R. V. & Dayhoff, M. O. Evolution of the structure of ferredoxin based on living relics of primitive amino acid sequences. Science 152, 363–366 (1966).
Ragsdale, S. W. & Pierce, E. Acetogenesis and the Wood–Ljungdahl pathway of CO2 fixation. Biochim. Biophys Acta 1784, 1873–1898 (2008).
Poulton, S. W. & Canfield, D. E. Ferruginous conditions: a dominant feature of the ocean through Earth’s history. Elements 7, 107–112 (2011).
Song, H. et al. The onset of widespread marine red beds and the evolution of ferruginous oceans. Nat. Commun. 8, 399 (2017).
Krissansen-Totton, J., Arney, G. N. & Catling, D. C. Constraining the climate and ocean pH of the early Earth with a geological carbon cycle model. Proc. Natl Acad. Sci. USA 115, 4105–4110 (2018).
Hutchison, W., Finch, A. A. & Boyce, A. J. The sulfur isotope evolution of magmatic-hydrothermal fluids: insights into ore-forming processes. Geochim. Cosmochim. Acta 288, 176–198 (2020).
Kawasumi, S. & Chiba, H. Redox state of seafloor hydrothermal fluids and its effect on sulfide mineralization. Chem. Geol. 451, 25–37 (2017).
Helmbrecht, V., Weingart, M., Klein, F., Braun, D. & Orsi, W. D. White and green rust chimneys accumulate RNA in a ferruginous chemical garden. Geobiology 21, 758–769 (2023).
Russell, M. J. Green rust: the simple organizing ‘seed’ of all life? Life 8, 35 (2018).
Crowe, S. A. et al. Sulfate was a trace constituent of Archean seawater. Science 346, 735–739 (2014).
Marty, B., Avice, G., Bekaert, D. V. & Broadley, M. W. Salinity of the Archaean oceans from analysis of fluid inclusions in quartz. C. R. Geosci. 350, 154–163 (2018).
Knauth, L. P. in Geobiology: Objectives, Concepts, Perspectives (ed. Noffke, N.) 53–69 (Elsevier, 2005).
Jones, W., Leigh, J., Mayer, F., Woese, C. & Wolfe, R. Methanococcus jannaschii sp. nov., an extremely thermophilic methanogen from a submarine hydrothermal vent. Arch. Microbiol. 136, 254–261 (1983).
Bult, C. J. et al. Complete genome sequence of the methanogenic archaeon, Methanococcus jannaschii. Science 273, 1058–1073 (1996).
Ver Eecke, H. C. et al. Hydrogen-limited growth of hyperthermophilic methanogens at deep-sea hydrothermal vents. Proc. Natl Acad. Sci. USA 109, 13674–13679 (2012).
Topçuoğlu, B. D., Meydan, C., Nguyen, T. B., Lang, S. Q. & Holden, J. F. Growth kinetics, carbon isotope fractionation, and gene expression in the hyperthermophile Methanocaldococcus jannaschii during hydrogen-limited growth and interspecies hydrogen transfer. Appl. Environ. Microbiol. 85, e00180–19 (2019).
Skinner, B. J., Erd, R. C. & Grimaldi, F. S. Greigite, the thio-spinel of iron; a new mineral. Am. Mineral. 49, 543–555 (1964).
Lafuente, B. et al. The power of databases: the RRUFF project. Highlights Mineral. Crystallogr. 1, 25 (2015).
Genchev, G. & Erbe, A. Raman spectroscopy of mackinawite FeS in anodic iron sulfide corrosion products. J. Electrochem. Soc. 163, C333–C338 (2016).
Mielke, R. E. et al. Iron-sulfide-bearing chimneys as potential catalytic energy traps at life’s emergence. Astrobiology 11, 933–950 (2011).
Roberts, A. P., Chang, L., Rowan, C. J., Horng, C. S. & Florindo, F. Magnetic properties of sedimentary greigite (Fe3S4): an update. Rev. Geophys. https://doi.org/10.1029/2010rg000336 (2011).
Dekkers, M. J., Passier, H. F. & Schoonen, M. A. Magnetic properties of hydrothermally synthesized greigite (Fe3S4)—II. High-and low-temperature characteristics. Geophys. J. Int. 141, 809–819 (2000).
Russell, M. J. & Ponce, A. Six ‘must-have’ minerals for life’s emergence: olivine, pyrrhotite, bridgmanite, serpentine, fougerite and mackinawite. Life 10, 291 (2020).
McGlynn, S. E., Mulder, D. W., Shepard, E. M., Broderick, J. B. & Peters, J. W. Hydrogenase cluster biosynthesis: organometallic chemistry nature’s way. Dalton Trans. https://doi.org/10.1039/B821432H (2009).
Nitschke, W., McGlynn, S. E., Milner-White, E. J. & Russell, M. J. On the antiquity of metalloenzymes and their substrates in bioenergetics. Biochim. Biophys. Acta 1827, 871–881 (1827).
Finklea, S. L., Cathey, L. & Amma, E. Investigation of the bonding mechanism in pyrite using the Mössbauer effect and X-ray crystallography. Acta Crystallogr. A 32, 529–537 (1976).
Wirth, R., Luckner, M. & Wanner, G. Validation of a hypothesis: colonization of black smokers by hyperthermophilic microorganisms. Front. Microbiol. 9, 524 (2018).
Klingl, A. et al. Analysis of the surface proteins of Acidithiobacillus ferrooxidans strain SP5/1 and the new, pyrite-oxidizing Acidithiobacillus isolate HV2/2, and their possible involvement in pyrite oxidation. Arch. Microbiol. 193, 867–882 (2011).
Gambelli, L. et al. An archaellum filament composed of two alternating subunits. Nat. Commun. 13, 710 (2022).
Rickard, D. & Luther, G. W. Chemistry of iron sulfides. Chem. Rev. 107, 514–562 (2007).
Thiel, J., Byrne, J. M., Kappler, A., Schink, B. & Pester, M. Pyrite formation from FeS and H2S is mediated through microbial redox activity. Proc. Natl Acad. Sci. USA 116, 6897–6902 (2019).
Brock, T. D., Madigan, M. T., Martinko, J. M. & Parker, J. Brock Biology of Microorganisms (Prentice-Hall, 2003).
Muchowska, K. B., Varma, S. J. & Moran, J. Nonenzymatic metabolic reactions and life’s origins. Chem. Rev. 120, 7708–7744 (2020).
Can, M., Armstrong, F. A. & Ragsdale, S. W. Structure, function, and mechanism of the nickel metalloenzymes, CO dehydrogenase, and acetyl-CoA synthase. Chem. Rev. 114, 4149–4174 (2014).
Gu, W., Gencic, S., Cramer, S. P. & Grahame, D. A. The A-cluster in subunit β of the acetyl-CoA decarbonylase/synthase complex from Methanosarcina thermophila: Ni and Fe K-edge XANES and EXAFS analyses. J. Am. Chem. Soc. 125, 15343–15351 (2003).
Grünberger, F. et al. Uncovering the temporal dynamics and regulatory networks of thermal stress response in a hyperthermophile using transcriptomics and proteomics. mBio 14, e02174–23 (2023).
Hedderich, R., Hamann, N. & Bennati, M. Heterodisulfide reductase from methanogenic archaea: a new catalytic role for an iron-sulfur cluster. Biol. Chem. 386, 961–970 (2005).
Ermler, U., Grabarse, W., Shima, S., Goubeaud, M. & Thauer, R. K. Crystal structure of methyl-coenzyme M reductase: the key enzyme of biological methane formation. Science 278, 1457–1462 (1997).
Wagner, T., Koch, J., Ermler, U. & Shima, S. Methanogenic heterodisulfide reductase (HdrABC-MvhAGD) uses two noncubane [4Fe-4S] clusters for reduction. Science 357, 699–703 (2017).
Makarova, K. S., Wolf, Y. I. & Koonin, E. V. Archaeal clusters of orthologous genes (arCOGs): an update and application for analysis of shared features between Thermococcales, Methanococcales, and Methanobacteriales. Life 5, 818–840 (2015).
Brabender, M. et al. Ferredoxin reduction by hydrogen with iron functions as an evolutionary precursor of flavin-based electron bifurcation. Proc. Natl Acad. Sci. USA 121, e2318969121 (2024).
Prakash, D., Wu, Y., Suh, S.-J. & Duin, E. C. Elucidating the process of activation of methyl-coenzyme M reductase. J. Bacteriol. 196, 2491–2498 (2014).
Schink, B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Mol. Biol. Rev. 61, 262–280 (1997).
Martin, W. & Müller, M. The hydrogen hypothesis for the first eukaryote. Nature 392, 37–41 (1998).
Shock, E. L., McCollom, T. & Schulte, M. D. in Thermophiles (eds Wiegel, J. & Michael, A. W. W.) 79–96 (CRC, 1998).
Martin, W. & Russell, M. J. On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Philos. Trans. R. Soc. Lond. B 358, 59–85 (2003).
Wächtershäuser, G. in The Molecular Origins of Life (ed. Brack, A.) 206–218 (Cambridge Univ. Press, 1998).
Muralidharan, V., Rinker, K., Hirsh, I., Bouwer, E. & Kelly, R. Hydrogen transfer between methanogens and fermentative heterotrophs in hyperthermophilic cocultures. Biotechnol. Bioeng. 56, 268–278 (1997).
Graham, D. E., Kyrpides, N., Anderson, I. J., Overbeek, R. & Whitman, W. B. Genome of Methanocaldococcus (Methanococcus) jannaschii. Methods Enzymol. 330, 40–123 (2001).
Buchfink, B., Xie, C. & Huson, D. H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12, 59–60 (2015).
Widdel, F. Theory and measurement of bacterial growth. Di Dalam Grundpraktikum Mikrobiol 4, 1–11 (2007).
Orsi, W. D. et al. Carbon assimilating fungi from surface ocean to subseafloor revealed by coupled phylogenetic and stable isotope analysis. ISME J. 16, 1245–1261 (2022).
Acknowledgements
This work was supported by the Deutsche Forschungsgemeinschaft project OR 417/8-1 granted to W.D.O. The work was also supported in part by the Transregio Collaborative Research Center (CRC 235, ‘Emergence of Life’) Project-ID 364653263—TRR 235 granted to W.D.O. We thank W. Martin for comments on the original version of the paper.
Funding
Open access funding provided by Ludwig-Maximilians-Universität München.
Author information
Authors and Affiliations
Contributions
V.H., R.R., D.G. and W.D.O. designed the experiments. V.H. performed the experiments. W.D.O. and V.H. analysed data. All authors commented on the paper and contributed to the writing process.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Ecology & Evolution thanks Julie Huber, Helene Ver Eecke and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Iron-sulfide particles are magnetic.
Sedimentary iron-sulfide chemical garden precipitates align with magnetic rings on a ring magnet plate, showing the presence of the magnetic iron-sulfide mineral greigite (Fe3S4).
Extended Data Fig. 2 Abiotic H2 produced in the 500 mM sedimentary iron-sulfide chemical gardens at 80 °C with and without M. jannaschii.
Error bars show standard error of the means across three biological replicates, P value is the result of a one-sided T-test.
Extended Data Fig. 3 Volcano plot analysis of gene expression of M. jannaschii transcriptomes in MMC Medium at 25 and 80 °C (Experiments 5c, d, Supplemental Table 1).
Of all significant genes (P = 0.05, one-sided t-test), 96 are overexpressed at 80 °C (dark blue, Experiment 5c) compared to 25 °C. 241 genes were down regulated at 80 °C compared to 25 °C (light blue, Experiment 5 d). The vertical line separates expressed genes that either increased or decreased in rank in the MMC medium, the dotted horizontal line represents the P-value cutoff for determining statistical significance (one-sided T-test). Of all overexpressed genes, two are part of the methanogenic acetyl CoA pathway: methyl coenzyme M reductase system component A2 (atwA) and NADPH-dependent F420 reductase (npdG).
Extended Data Fig. 4 Volcano plot analysis of gene expression of M. jannaschii transcriptomes in sterile water relative to MMC medium (Experiments 5b, c, Supplemental Table 1).
Of all significant genes (P = 0.05, one-sided t-test), 59 are overexpressed in the sterile water controls (blue, Experiment 5b) compared to MMC medium (beige, Experiment 5c). Significantly overexpressed genes are listed in Supplemental Table 4. The full list of overexpressed genes is provided in Supplemental Table 4.
Extended Data Fig. 5 Functional annotation of overexpressed genes. (Experiments 5a-c, Supplemental Table 1).
For gene analysis, the arCOGs (Archaeal Clusters of Orthologous Genes) database was used79. (a) Out of 34 overexpressed genes in the sedimentary iron-sulfide chemical garden relative to MMC medium (Fig. 4a), the majority of genes has a function in metabolism (blue) or information, storage and processing (red), especially in coenzyme transport (H) and translation (J). (b) Out of 46 overexpressed genes in the sedimentary iron-sulfide chemical garden relative to sterile H2O, most genes have a function in metabolism (blue), especially in energy production and conversion (C). Genes regulating cellular processes and signaling (green) make a proportion of 17 % of the overexpressed genes. (c) In the sterile H2O controls relative to MMC medium, 59 genes were overexpressed, with 44 % having a metabolic function (blue).
Supplementary information
Supplementary Information
Supplementary Tables 1–4 and Methods.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Helmbrecht, V., Reichelt, R., Grohmann, D. et al. Simulated early Earth geochemistry fuels a hydrogen-dependent primordial metabolism. Nat Ecol Evol 9, 769–778 (2025). https://doi.org/10.1038/s41559-025-02676-w
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41559-025-02676-w
This article is cited by
-
Photoholes within cyanobacterial mats can account for the origin of anammox bacteria and ancient nitrogen loss
Nature Ecology & Evolution (2026)
