
Abstract
Fasting is associated with a range of health benefits1,2,3,4,5,6. How fasting signals elicit changes in the proteome to establish metabolic programmes remains poorly understood. Here we show that hepatocytes selectively remodel the translatome while global translation is paradoxically downregulated during fasting7,8. We discover that phosphorylation of eukaryotic translation initiation factor 4E (P-eIF4E) is induced during fasting. We show that P-eIF4E is responsible for controlling the translation of genes involved in lipid catabolism and the production of ketone bodies. Inhibiting P-eIF4E impairs ketogenesis in response to fasting and a ketogenic diet. P-eIF4E regulates those messenger RNAs through a specific translation regulatory element within their 5′ untranslated regions (5′ UTRs). Our findings reveal a new signalling property of fatty acids, which are elevated during fasting. We found that fatty acids bind and induce AMP-activated protein kinase (AMPK) kinase activity that in turn enhances the phosphorylation of MAP kinase-interacting protein kinase (MNK), the kinase that phosphorylates eIF4E. The AMPK–MNK–eIF4E axis controls ketogenesis, revealing a new lipid-mediated kinase signalling pathway that links ketogenesis to translation control. Certain types of cancer use ketone bodies as an energy source9,10 that may rely on P-eIF4E. Our findings reveal that on a ketogenic diet, treatment with eFT508 (also known as tomivosertib; a P-eIF4E inhibitor) restrains pancreatic tumour growth. Thus, our findings unveil a new fatty acid-induced signalling pathway that activates selective translation, which underlies ketogenesis and provides a tailored diet intervention therapy for cancer.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others
Data availability
All data necessary for the conclusions of the study are provided with the article. Sequencing data generated for the PolyRibo-seq analysis of fed and fasted livers from WT and eIF4ES209A mice, chow- and ketogenic-diet-fed mice livers and T3M4 xenograft tumours from mice with different treatments are available at the Gene Expression Omnibus under SuperSeries accession numbers GSE232342, GSE267336 and GSE267398. The mass spectrometry metabolomics data have been deposited to the Metabolomics Workbench under Study ID ST003199. Source data are provided with this paper.
References
de Cabo, R. & Mattson, M. P. Effects of intermittent fasting on health, aging, and disease. N. Engl. J. Med. 381, 2541–2551 (2019).
Sutton, E. F. et al. Early time-restricted feeding improves insulin sensitivity, blood pressure, and oxidative stress even without weight loss in men with prediabetes. Cell Metab. 27, 1212–1221 e1213 (2018).
Tiwari, S., Sapkota, N. & Han, Z. Effect of fasting on cancer: a narrative review of scientific evidence. Cancer Sci. 113, 3291–3302 (2022).
Harris, L. et al. Intermittent fasting interventions for treatment of overweight and obesity in adults: a systematic review and meta-analysis. JBI Database System. Rev. Implement. Rep. 16, 507–547 (2018).
Tan-Shalaby, J. Ketogenic diets and cancer: emerging evidence. Fed. Pract. 34, 37S–42S (2017).
Dowis, K. & Banga, S. The potential health benefits of the ketogenic diet: a narrative review. Nutrients 13, 1654 (2021).
Yuan, C. L. et al. Preserved protein synthesis in the heart in response to acute fasting and chronic food restriction despite reductions in liver and skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 295, E216–E222 (2008).
Afolabi, P. R. et al. The effect of total starvation and very low energy diet in lean men on kinetics of whole body protein and five hepatic secretory proteins. Am. J. Physiol. Endocrinol. Metab. 293, E1580–E1589 (2007).
Zahra, A. et al. Consuming a ketogenic diet while receiving radiation and chemotherapy for locally advanced lung cancer and pancreatic cancer: the University of Iowa experience of two phase 1 clinical trials. Radiat. Res. 187, 743–754 (2017).
Hopkins, B. D. et al. Suppression of insulin feedback enhances the efficacy of PI3K inhibitors. Nature 560, 499–503 (2018).
Puchalska, P. & Crawford, P. A. Metabolic and signaling roles of ketone bodies in health and disease. Annu. Rev. Nutr. 41, 49–77 (2021).
Sengupta, S., Peterson, T. R., Laplante, M., Oh, S. & Sabatini, D. M. mTORC1 controls fasting-induced ketogenesis and its modulation by ageing. Nature 468, 1100–1104 (2010).
Dowling, R. J. et al. mTORC1-mediated cell proliferation, but not cell growth, controlled by the 4E-BPs. Science 328, 1172–1176 (2010).
Waskiewicz, A. J., Flynn, A., Proud, C. G. & Cooper, J. A. Mitogen-activated protein kinases activate the serine/threonine kinases Mnk1 and Mnk2. EMBO J. 16, 1909–1920 (1997).
Furic, L. et al. eIF4E phosphorylation promotes tumorigenesis and is associated with prostate cancer progression. Proc. Natl Acad. Sci. USA 107, 14134–14139 (2010).
Wendel, H. G. et al. Dissecting eIF4E action in tumorigenesis. Genes Dev. 21, 3232–3237 (2007).
Scheper, G. C. & Proud, C. G. Does phosphorylation of the cap-binding protein eIF4E play a role in translation initiation? Eur. J. Biochem. 269, 5350–5359 (2002).
Zhulyn, O. et al. Evolutionarily divergent mTOR remodels translatome for tissue regeneration. Nature 620, 163–171 (2023).
Morita, M. et al. mTOR coordinates protein synthesis, mitochondrial activity and proliferation. Cell Cycle 14, 473–480 (2015).
Guillon, J. et al. tRNA biogenesis and specific aminoacyl-tRNA synthetases regulate senescence stability under the control of mTOR. PLoS Genet. 17, e1009953 (2021).
Casals, N. et al. Regulation of the expression of the mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase gene. Its role in the control of ketogenesis. Biochem. J. 283, 261–264 (1992).
Kersten, S. et al. Peroxisome proliferator-activated receptor alpha mediates the adaptive response to fasting. J. Clin. Invest. 103, 1489–1498 (1999).
Montagner, A. et al. Liver PPARalpha is crucial for whole-body fatty acid homeostasis and is protective against NAFLD. Gut 65, 1202–1214 (2016).
Xu, Y. et al. Translation control of the immune checkpoint in cancer and its therapeutic targeting. Nat. Med. 25, 301–311 (2019).
Reich, S. H. et al. Structure-based design of pyridone-aminal eFT508 targeting dysregulated translation by selective mitogen-activated protein kinase interacting kinases 1 and 2 (MNK1/2) inhibition. J. Med. Chem. 61, 3516–3540 (2018).
Venable, A. H. et al. Fasting-induced HMGCS2 expression in the kidney does not contribute to circulating ketones. Am. J. Physiol. Renal Physiol. 322, F460–F467 (2022).
Truitt, M. L. et al. Differential requirements for eIF4E dose in normal development and cancer. Cell 162, 59–71 (2015).
Hsieh, A. C. et al. The translational landscape of mTOR signalling steers cancer initiation and metastasis. Nature 485, 55–61 (2012).
Wu, J. C., Merlino, G. & Fausto, N. Establishment and characterization of differentiated, nontransformed hepatocyte cell lines derived from mice transgenic for transforming growth factor alpha. Proc. Natl Acad. Sci. USA 91, 674–678 (1994).
Grygiel-Gorniak, B. Peroxisome proliferator-activated receptors and their ligands: nutritional and clinical implications—a review. Nutr. J. 13, 17 (2014).
DiMarco, N. M., Beitz, D. C. & Whitehurst, G. B. Effect of fasting on free fatty acid, glycerol and cholesterol concentrations in blood plasma and lipoprotein lipase activity in adipose tissue of cattle. J. Anim. Sci. 52, 75–82 (1981).
Abe, I. et al. Lipolysis-derived linoleic acid drives beige fat progenitor cell proliferation. Dev. Cell 57, 2623–2637 e2628 (2022).
Fukunaga, R. & Hunter, T. MNK1, a new MAP kinase-activated protein kinase, isolated by a novel expression screening method for identifying protein kinase substrates. EMBO J. 16, 1921–1933 (1997).
Li, Y. et al. Protein phosphatase 2A negatively regulates eukaryotic initiation factor 4E phosphorylation and eIF4F assembly through direct dephosphorylation of Mnk and eIF4E. Neoplasia 12, 848–855 (2010).
Bujak, A. L. et al. AMPK activation of muscle autophagy prevents fasting-induced hypoglycemia and myopathy during aging. Cell Metab. 21, 883–890 (2015).
Minokoshi, Y. et al. AMP-kinase regulates food intake by responding to hormonal and nutrient signals in the hypothalamus. Nature 428, 569–574 (2004).
Shveygert, M., Kaiser, C., Bradrick, S. S. & Gromeier, M. Regulation of eukaryotic initiation factor 4E (eIF4E) phosphorylation by mitogen-activated protein kinase occurs through modulation of Mnk1-eIF4G interaction. Mol. Cell. Biol. 30, 5160–5167 (2010).
Parra, J. L., Buxade, M. & Proud, C. G. Features of the catalytic domains and C termini of the MAPK signal-integrating kinases Mnk1 and Mnk2 determine their differing activities and regulatory properties. J. Biol. Chem. 280, 37623–37633 (2005).
Goto, S., Yao, Z. & Proud, C. G. The C-terminal domain of Mnk1a plays a dual role in tightly regulating its activity. Biochem. J 423, 279–290 (2009).
Xiao, B. et al. Structural basis for AMP binding to mammalian AMP-activated protein kinase. Nature 449, 496–500 (2007).
Oakhill, J. S. et al. AMPK is a direct adenylate charge-regulated protein kinase. Science 332, 1433–1435 (2011).
Pinkosky, S. L. et al. Long-chain fatty acyl-CoA esters regulate metabolism via allosteric control of AMPK beta1 isoforms. Nat. Metab. 2, 873–881 (2020).
Li, X. et al. Erratum: structural basis of AMPK regulation by adenine nucleotides and glycogen. Cell Res. 25, 398 (2015).
Chen, L. et al. Conserved regulatory elements in AMPK. Nature 498, E8–E10 (2013).
Xin, F. J., Wang, J., Zhao, R. Q., Wang, Z. X. & Wu, J. W. Coordinated regulation of AMPK activity by multiple elements in the alpha-subunit. Cell Res. 23, 1237–1240 (2013).
Steinberg, G. R. & Hardie, D. G. New insights into activation and function of the AMPK. Nat. Rev. Mol. Cell Biol. 24, 255–272 (2023).
Gouirand, V. et al. Ketogenic HMG-CoA lyase and its product beta-hydroxybutyrate promote pancreatic cancer progression. EMBO J. 41, e110466 (2022).
Gwinn, D. M. et al. AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol. Cell 30, 214–226 (2008).
Hoppe, S. et al. AMP-activated protein kinase adapts rRNA synthesis to cellular energy supply. Proc. Natl Acad. Sci. USA 106, 17781–17786 (2009).
Basu, S. S. & Blair, I. A. SILEC: a protocol for generating and using isotopically labeled coenzyme A mass spectrometry standards. Nat. Protoc. 7, 1–12 (2011).
Liu, X. et al. High-resolution metabolomics with acyl-CoA profiling reveals widespread remodeling in response to diet. Mol. Cell Proteomics 14, 1489–1500 (2015).
MacLean, B. et al. Skyline: an open source document editor for creating and analyzing targeted proteomics experiments. Bioinformatics 26, 966–968 (2010).
Kawaguchi, A., Yoshimura, T. & Okuda, S. A new method for the preparation of acyl-CoA thioesters. J. Biochem. 89, 337–339 (1981).
Jo, S., Kim, T., Iyer, V. G. & Im, W. CHARMM-GUI: a web-based graphical user interface for CHARMM. J. Comput. Chem. 29, 1859–1865 (2008).
Lundborg, M. & Lindahl, E. Automatic GROMACS topology generation and comparisons of force fields for solvation free energy calculations. J. Phys. Chem. B 119, 810–823 (2015).
Gu, X. et al. Deconvoluting AMP-activated protein kinase (AMPK) adenine nucleotide binding and sensing. J. Biol. Chem. 292, 12653–12666 (2017).
Yan, Y. et al. Structure of an AMPK complex in an inactive, ATP-bound state. Science 373, 413–419 (2021).
Feenstra, K. A., Hess, B. & Berendsen, H. J. C. Improving efficiency of large time-scale molecular dynamics simulations of hydrogen-rich systems. J. Comput. Chem. 20, 786–798 (1999).
Gao, Y. et al. CHARMM-GUI supports hydrogen mass repartitioning and different protonation states of phosphates in lipopolysaccharides. J. Chem. Inf. Model. 61, 831–839 (2021).
Acknowledgements
We thank members of the Ruggero laboratory for discussion, L. Song (UCSF) for providing the T3M4 cell line, and L. Qiu and D. Wang (UCSF) for performing hydrodynamic tail vein injection. We thank eFFECTOR Therapeutics for providing eFT508. We thank the Preclinical Therapeutics Core (PTC), and The Center for Advanced Technology (CAT) at UCSF for their assistance in our study. Mass Spectrometry was provided by the Mass Spectrometry Resource at UCSF (A. L. Burlingame, Director) supported by the Dr. Miriam and Sheldon G. Adelson Medical Research Foundation (AMRF) and the UCSF Program for Breakthrough Biomedical Research (PBBR). Metabolomics and lipidomics was performed and analysed by Metabolomics Platform from University of Chicago Medicine Comprehensive Cancer Center (RRID: SCR_022932) supported by H. Shah. and R. AminiTabrizi. Mouse primary hepatocytes were provided by the Liver Cell Isolation, Analysis and Immunology Core from UCSF Liver Center, supported by S. Dhillon. H.Y. is funded by American Heart Association (grant nos. P0540503, P0564550). S.K. is funded by NIH (grant no. DK125281) and the Howard Hughes Medical Institute. M.G. is funded by NIH grant no. R01GM137109. D.R. is funded by NIH (grant no. R35CA242986) and the American Cancer Society (American Cancer Society Research Professor Award). Illustrations in Figs. 1a,c,g,h, 2e,g,k, 3a,b,d,f,k, 4a,d,j and 5a,c,e,g–k and Extended Data Figs. 5d,e,k,m and 8a,h were created using BioRender (BioRender.com).
Author information
Authors and Affiliations
Contributions
H.Y. and D.R. designed experimental outline and wrote the paper. D.R. supervised the project. H.Y., V.A.Z. and H.T. performed experiments. J.L., S.O., J.A.O.-P., M.G., S.K., A.L.B., H.S., Q.E. and I.S. assisted with experiments, analysis and provided research expertise.
Corresponding author
Ethics declarations
Competing interests
D.R. is a shareholder of eFFECTOR Therapeutics, Inc., and a member of its scientific advisory board. The other authors declare no competing interests.
Peer review
Peer review information
Nature thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
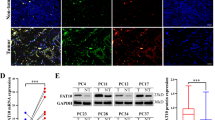
Extended Data Fig. 1 Ketogenesis is controlled at the translation level during fasting through P-eIF4E.
a. Quantification for the indicated proteins and phosphorylation states of immunoblot of liver lysates from mice were given chow ad libitum or fasted for 24 (n = 3 mice). b. Blood glucose levels (n = 5 mice), c. Serum insulin levels (n = 8 mice), d. Body weight of WT and eIF4ES209A mice in fed and 24 h fasted conditions (n = 5 mice). e. Serum fatty acids (FA) levels (n = 4 or 5 mice) and f. Serum glycerol levels (n = 5 mice) in 24 h fasted WT and eIF4ES209A mice. g. Relative levels of indicated metabolites in 24 h fasted WT or eIF4ES209A livers (n = 3 mice). h. Body weight of WT treated with eFT508 or vehicle after 24 h fasting (n = 4 mice). i. Serum fatty acids (FA) levels (n = 4 mice), and j. Glycerol levels in 24 h fasted WT mice treated with vehicle or eFT508 (n = 4 or 5 mice). All values represent the mean ± SEM; Two-way ANOVA for a, d, g, h, and two-sided Student’s t-test for b, c, e, g, f, i, j.
Extended Data Fig. 2 P-eIF4E is required for fasting-induced translation through the PRTE motif on 5′UTR.
a. Representative polysome profiles of 24 h fasted WT and eIF4ES209A liver lysates separated on a sucrose gradient; Inset highlights the free/RNP/80 S/light polysome (2–7) and heavy polysomal fractions (8–13) used for translational profiling by polysomal RNAseq. b. Top pathways enriched in the 445 genes that are significantly translationally upregulated upon fasting in WT conditions but fail to increase in eIF4ES209A livers based on the wiki pathway database. c. Percentages of the house keeping gene, Beta-2 microglobulin (B2M) mRNA distributed in sucrose gradient fractions against its total mRNA, of the 24 h fasted WT and eIF4ES209A livers (n = 3 mice). d. Representative images of immunohistochemistry (IHC) of Hmgcs2 on sections from 24 h fasted WT or eIF4ES209A livers. e. Representative images, and f. Quantification of IHC of PPARα on liver sections from 24 h fasted WT, PPARα knockout (PPARα−/−), or eIF4ES209A mice (n = 3 mice). g. Relative mRNA levels of the downstream targets of PPARα, Cpt2 and Hsd17b10 in fasted WT and eIF4ES209A liver normalized to B2M (n = 3 mice). h. Representative image of luciferase expression of WT and eIF4ES209A mice injected with PPARα 5′UTR linked luciferase in 24 h fasted condition. i. Relative expression of Fluc with no 5′UTR normalized to the expression of the internal control Rluc at full medium (FM) or 24 h induction of fasting mimicking medium (Fast: serum free with FA, n = 3 biological independent samples). j. Relative expression of Fluc with Hmgcs2 5′UTR, k. eIF5A 5′UTR, l. HNRNPC 5′UTR, m. RPL26 5′UTR, and n. ABCD3 5′UTR or C to G transversion mutated PRTE motif in corresponding 5′UTR at full medium (FM) or 24 h induction of fasting mimicking medium (n = 3 biological independent samples). All values represent the mean ± SEM; Two-way ANOVA for g, One-way ANOVA for e, and two-sided Student’s t-test for f, i, j, k, l, m, n.
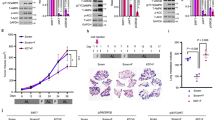
Extended Data Fig. 3 Fatty acids activate the MNK-P-eIF4E axis by enhancing AMPK activity.
a. Immunoblot and quantification of indicated proteins in primary hepatocytes isolated from mice liver treated with BSA, 200 μM palmitic acid (PA), oleic acid (OA), or linoleic acid (LA) for 4 h after overnight serum starvation, and immunoblot quantification of P-eIF4E/eIF4E in the AML12 cells and primary hepatocytes in response to BSA or different fatty acids (n = 3 biological independent samples). b. Immunoblot of indicated proteins in AML12 cells or primary hepatocytes treated with 100μM fatty acids mix (FA) for 0, 1, 2, 3 h after overnight serum starvation (n = 3 biological independent samples). c. Percentages of indicated mRNA distributed in sucrose gradient fractions of AML12 cells treated with BSA or 150 μM linoleic acid for 4 h in overnight serum-free medium (n = 3 biological independent samples). d. Percentages of indicated mRNA distributed in sucrose gradient fractions of livers from WT or fat specific adipose triglyceride lipase knockout mice (ATGL−/−) after 24 h fasting (n = 3 mice). e. Relative expression of Fluc with PPARα 5′UTR or empty 5′UTR after BSA, 150 μM linoleic acid or linoleic acid plus preincubation of 1 μM eFT508 in serum-free conditions (n = 3 biological independent samples). f. Immunoblot of indicated proteins in AML12 cells treated with different inhibitors before and during 4 h of linoleic acid stimuli in serum-free conditions. g Immunoblot and quantification of indicated proteins in AML12 cells pretreated with vehicle or AMPK inhibitor (Bay-3827) in serum-free conditions upon 4 h fatty acids stimuli (n = 3 biological independent samples). h. Immunoblot of indicated proteins in primary hepatocytes treated with vehicle or AMPK inhibitor (Compound C) in serum-free conditions upon fatty acids (150μM mix of PA, OA and LA) stimuli (n = 3 biological independent samples). i. Percentages of PPARα, Hmgcs2 and B2M mRNAs distributed in sucrose gradient fractions against their total mRNA levels of AML12 cells preincubated with vehicle or 10 μM AMPK inhibitor Compound C in serum-free conditions upon 4 h linoleic acid stimuli (n = 3 biological independent samples). j. Immunoblot of indicated proteins in fed and 24 h fasted livers (n = 3 mice). k. Blood BHB levels of 24 h fasted eIF4ES209A mice treated with vehicle or AMPK inhibitor Compound C before fasting and 12 h after fasting (n = 4 mice). All values represent the mean ± SEM; Two-way ANOVA for a, c, d, e, i, One-way ANOVA for g, and two-sided Student’s t-test for k.
Extended Data Fig. 4 AMPK phosphorylates MNK.
a. Relative AMPK activity on MNK1 as substrate in the presence of vehicle or 10 μM AMPK inhibitor compound C (n = 3 biological independent samples). b. MSMS spectra belonging to tryptic peptides spanning amino acids A36 to K49 (panel A), D165 to R210 (B), and N392 to R410 (C) of human MNK1, obtained by HCD fragmentation of precursor ions 841.3626+2, 1157.8012+4, and 1067.4977+2 respectively. An excess mass of 79.9663 compared to the unmodified sequence, corresponding to the phosphate group, is observed in the precursor and in sequence ions than contain serine 39, serine 168 and serine 394. Experimental masses of the most representative sequence ion peaks are labelled in the spectra, also indicating the fragment type (Roepstorff-Fohlmann-Biemann ions nomenclature). The position of the fragmentation events generating these ions are indicated in the sequences over the spectra. A table indicating the theoretical masses of the sequence ions according to the proposed modified sequences is shown in the right side, with red font indicating observed fragments. SPhospho, phosphorylated serina; CCM, carboxymethylated cysteine. c. Relative AMPK activity on the same amount of purified wild type MNK1, S39A, S168A, S394A and triple mutation of S39A, S168A and S394A MNK1, and AMPK’s known substrate SAMStide; all readings were normalized by subtracting the values obtained with the AMPK inhibitor (n = 3 biological independent samples). d. Silver staining of purified human eIF4E-GST and human wild type or mutated MNK1-GST. e. Stacked 1H NMR (300 MHz, DMSO-d6) spectra of linoleic acid (green), CoASH (red), and linoleoyl-CoA (blue). A characteristic shift of the triplet at 2.2 ppm in linoleic acid to 2.8 ppm is seen in linoleoyl-CoA, corresponding to the protons alpha to the thioester carbonyl (indicated by arrows on the chemical structure and in the spectrum). All values represent the mean ± SEM; One-way ANOVA for c, and two-sided Student’s t-test for a.
Extended Data Fig. 5 Linoleic acid activates AMPK through directly binding to AMPKγ.
a. Relative AMPKα1β1γ1 activity on its substrate SAMStide in the presence of vehicle (Veh), BSA, 100 μM linoleic acid (LA), 10 μM linoleoyl-CoA (LA-CoA) or 100 μM AMP (n = 5 biological independent samples) b. Relative AMPKα1β1γ1 activity on its substrate peptide SAMStide in the presence of vehicle (Veh), or the indicated linoleic acid and AMP combinations (n = 3 biological independent samples). c. Silver staining of blue native gel with recombinant AMPK complex with bodipy or C12-bodipy. d. Immunoblot of indicated proteins in whole cell lysates from fed and 24 h fasted livers, and linoleic acid conjugated or butyric acid conjugated beads pull-downed proteins; the total proteins of each sample on membrane were shown using ponceau S staining. e. Immunoblot of residual AMPKγ in the purified AMPKα1β1γ1 incubated linoleic acid conjugated beads or butyric acid beads after competing with different concentrations of free linoleic acid or butyric acid. f. Relative AMPKα1β2γ1 activity on its substrate SAMStide, in the presence of vehicle (Veh), BSA, 100 μM linoleic acid (LA), 10 μM linoleoyl-CoA (LA-CoA), 10 μM palmitoyl-CoA (PA-CoA) or 100 μM AMP (n = 3 biological independent samples). g. Relative AMPKα1β2γ1 activity on its substrate SAMStide, with different concentrations of linoleic acid (n = 5 biological independent samples). h. The isolated AMPKγ1-subunit remains stable. Backbone RMSD values of the γ-subunit are report without (blue line) and with (orange line) AMP bound at CBS sites −1, −3, and −4. The relatively low and stable RMSDs over these simulations confirm that simulations of γ can be run in the absence of the rest of the AMPK complex. i. A heat map of linoleic acid contacts with AMPKγ1 reveals primary sites of interaction. Structure of the γ subunit (PDB 4RER) with residues colored by contact time with linoleic acid aggregated across all lipid binding simulations, highlighting identified lipid binding Sites 1 and 2. The color spectrum runs from white to red, with white being no/minimal fatty acid contact and red being high contact. Site 1 is a hydrophobic pocket buried among the main and side chains of L173, K177, L285, I289, V293, L315, I318, and Y29; Site 2 is buried within F91, I94, L95, A250, Y255, K58, F62, V65, and L109. A third site, Site 3, at the bottom of the figure also sampled frequent contact with linoleic acid, but was not tested as a candidate for mutation given the site’s proximity to the binding interface with the β-subunit, reduced contact times compared to Sites 1 and 2, and the diffuse nature of the interaction surface. j. Proposed mutations to Sites 1 (left column) and 2 (right column) for disrupting linoleic acid binding. The images highlight the wild type residues (top row) and proposed mutations (bottom row). Mutations were selected based on fatty acid binding simulations, aiming to crowd out the hydrophobic pockets that linoleic acid was seen to enter in order to prevent binding. k. Scheme of expression of different AMPKγ1 mutants in Hek293T cells, and purification of AMPKα1β1γ1 complex with different AMPKγ mutants using anti-flag beads; AMPKα1 and AMPKγ1 levels in the pulled-down samples were shown using immunoblot; relative AMPK activity of AMPK complex with different AMPKγ1 mutants when used SAMStide as substrate and reaction with AMPK inhibitor compound C was used as a negative control (n = 3 biological independent samples). l. Immunoblot of different AMPKγ1 mutants in the input and bound to the beads. m. Silver staining of purified human wild type or mutated AMPKα1β1γ1 complex. All values represent the mean ± SEM; One-way ANOVA for a, b, f, linear regression analysis was used for g, and two-sided Student’s t test for g, i, j, k. Schematics in d, e, k, m is created using BioRender (BioRender.com).
Extended Data Fig. 6 Ketogenic diet activates P-eIF4E dependent translational regulation of ketogenic mRNAs.
a. Quantification of indicated protein in the immunoblot of liver lysates from mice fed with regular chow or ketogenic diet (n = 3 mice). b. Quantification of indicated protein in the immunoblot of liver lysates from mice on ketogenic diet treated with vehicle or AMPK inhibitor (Comp. C) (n = 3 mice). c. Body weight of WT and eIF4ES209A mice fed with regular chow and ketogenic diet (n = 5 or 7 mice). d. Serum glycerol (n = 7 mice), e. Serum fatty acids (FA) (n = 7 mice) in WT and eIF4ES209A mice fed with ketogenic diet. f. Percentages of indicated mRNA distributed in sucrose gradient fractions of ketogenic diet fed WT and eIF4ES209A livers (n = 3 mice). g. Immunoblot of indicated proteins, h. Representative images of immunohistochemistry (IHC) of Hmgcs2, and i. Triglyceride levels (n = 3 or 4 mice) in the liver from eIF4ES209A mice injected with vehicle or Hmgcs2 cDNA with Myc tag. j. Immunoblot of indicated proteins and k. Representative images of IHC of Myc in the liver from eIF4ES209A mice injected with vehicle or PPARα cDNA with Myc tag. l. qPCR analysis of PPARα and Hmgcs2 mRNA levels in the liver from WT mice, and eIF4ES209A mice with hydrodynamic injection of vehicle or PPARα cDNA (n = 3 mice). All values represent the mean ± SEM; Two-way ANOVA for a, b, c, l, and two-sided Student’s t test for d, e, i.
Extended Data Fig. 7 Combination of eFT508 and ketogenic diet systemically inhibits pancreatic tumour growth.
a. Tumour size, b. Tumour weight, c. Blood BHB levels, and d. Body weight of xenograft mouse models with the human pancreatic cancer cell line AsPC1, treated with chow, eFT508, ketogenic diet or eFT508 and ketogenic diet after the tumours reach 150 mm3 (n = 4 mice). e. Tumour weight (n = 9 or 10 biological independent samples) f. Mouse body weight (n = 5 mice), g. Food intake (n = 5 days), h. Blood glucose levels (n = 5 mice), i. Blood glycerol levels (n = 5 mice), j. Serum fatty acids levels (n = 5 mice), and k. Serum insulin levels (n = 4 or 5 mice) of xenograft mouse models with the human pancreatic cancer cell line T3M4 with different treatment after the tumours reach 150 mm3. l. Relative cell survival of T3M4 cells treated without or with 1 mM BHB in fasting-mimicking medium (no glucose serum-free medium with 100 μM linoleic acid) for 48 h (n = 3 biological independent samples. m. Mouse body weight (n = 5 mice) and n. Represent tumour images of xenograft mouse models with the human pancreatic cancer cell line T3M4, treated with ketogenic diet or eFT508 and ketogenic diet or eFT508 and ketogenic diet with 1% BHB in the drinking water after the tumours reach 150 mm3. All values represent the mean ± SEM; Two-way ANOVA for a, b, c, d, f, h, l, m, One-way ANOVA for e, and two-sided Student’s t test for g, i, j, k.
Extended Data Fig. 8 Combination of eFT508 and ketogenic diet intrinsically affects pancreatic tumour growth.
a. AsPC1 cells were treated with 100 μM linoleic acid or BSA for 4 h or 24 h in low glucose serum-free (SF) medium and cell lysates were analyzed by immunoblotting for the indicated proteins and phosphorylation states (n = 3 biological independent samples). b. Percentages of indicated mRNA distributed in sucrose gradient fractions of tumours from xenograft mouse models with the human pancreatic cancer cell line T3M4, treated with ketogenic diet or eFT508 and ketogenic diet (n = 3 biological independent samples). c. Immunoblot of GFP and relative cell numbers of control T3M4 and PPARα-GFP cDNA overexpressed T3M4 in full medium. d. Tumour weight and representative images of end point tumours from xenograft mouse models with the control T3M4 or T3M4 cells overexpressing PPARα cDNA treated with ketogenic diet (Keto) and vehicle (Veh) or a combination of eFT508 and ketogenic diet (n = 9 or 10 biological independent samples). e. Immunoblot of indicated proteins from the xenograft tumours with the control T3M4 or PPARα cDNA overexpressed T3M4 cells treated vehicle (Veh) eFT508 upon a ketogenic diet (n = 3 mice). f. Blood BHB levels of xenograft mouse models with the control T3M4 or PPARα cDNA overexpressed T3M4 cells treated with vehicle (Veh) or eFT508 upon a ketogenic diet (n = 5 mice). g. Relative cell survival levels of T3M4 or AsPC1 pancreatic cancer cells treated with eFT508, PPARα inhibitor, eFT508 and PPARα inhibitor, and β-hydroxybutyrate in the presence of eFT508 in fasting-mimicking medium (FFM: no glucose serum-free medium with 100 μM linoleic acid) for 2 days (n = 3 biological independent samples). h. Scheme of T3M4 stably expressing luciferase reporter with wild type PPARα 5′UTR (WT) or PRTE mutated PPARα 5′UTR (Mut). i. The quantification of luciferase signal and representative images of the tumours from the xenograft mouse model in response to regular chow or ketogenic diet (n = 7 or 8 mice). j. The quantification of luciferase signal and representative images of the tumours from the xenograft mouse model in response to vehicle or eFT508 upon ketogenic diet (n = 6 mice). All values represent the mean ± SEM; Two-way ANOVA for b, d, f, and One-way ANOVA for g, and two-sided Student’s t test for i, j; Schematics in a, h is created using BioRender (BioRender.com).
Supplementary information
Supplementary Data 1
Raw western blot data in Supplementary Figs. 1–6. This PDF contains all fully annotated raw western blots analysed in this paper. All western blots contain molecular weight annotations.
Supplementary Data 2
Translational remodelling revealed by PolyRibo-seq during the fed–fasted transition in WT livers. This Excel file contains the log2 fold changes (logFC) of translation efficiency (TE) and adjusted P value of significantly changed transcripts between fed and 24 h fasted livers.
Supplementary Data 3
Metabolomics and short-chain acyl-CoA profiles revealed by mass spectrometry of 24 h fasted WT livers and eIF4ES209A livers. This Excel file contains a datasheet of (log2) fold change of metabolites in 24 h fasted eIF4ES209A livers compared to WT livers, as well as adjusted P value; and another datasheet of the peak area of short-chain Acyl-CoAs in fasted WT livers and eIF4ES209A livers and control samples.
Supplementary Data 4
Translatome changes revealed by PolyRibo-seq during the fed–fasted transition in WT livers, and 24 h fasted eIF4ES209A livers. This Excel file contains the log2 fold changes (logFC) of translation efficiency (TE) and adjusted P value of the genes that are significantly upregulated on fasting in WT livers compared to fed, but significantly downregulated in fasted eIF4ES209A livers compared to WT livers.
Supplementary Data 5
Translational remodelling revealed by PolyRibo-seq of livers from ketogenic diet and chow fed WT mice. This Excel file contains log2 of translation efficiency (TE) at different conditions of the genes that are significantly upregulated on fasting in WT livers compared to fed, but significantly downregulated in fasted eIF4ES209A livers compared to WT livers, and P values of changed transcripts between livers from mice fed with ketogenic diet and chow for 24 h.
Supplementary Data 6
Translational remodelling revealed by PolyRibo-seq of livers from ketogenic diet and chow fed WT mice. This Excel file contains log2 fold changes (logFC) of translation efficiency (TE) and the adjusted P value of significantly changed transcripts between mice fed with ketogenic diet and chow for 24 h.
Supplementary Data 7
Short-chain acyl-CoA profiles revealed by mass spectrometry of livers from mice fed with ketogenic diet and chow for 24 hours. This Excel file contains the peak area of short-chain Acyl-CoAs in livers from mice fed with ketogenic diet and chow for 24 h, and control samples.
Supplementary Data 8
Translatome changes revealed by PolyRibo-seq in T3M4 xenograft tumours under different treatments. This Excel file contains a datasheet (TE between keto and chow) with the log2 fold change of translation efficiency (TE) and adjusted P value of significantly changed transcripts between T3M4 tumours from xenograft mice fed with ketogenic diet and chow; and a second datasheet (TE between eFT508 to Veh) with the log2 fold change of translation efficiency (TE) and adjusted P value of significantly changed transcripts between T3M4 tumours from xenograft mice treated with eFT508 or vehicle on ketogenic diet.
Supplementary Data 9
Table of molecular dynamics simulations summary.
Supplementary Data 10
Table of PRTE motif sequence in the 5′ UTR of each mRNA and the corresponding C to G mutated sequence.
Source data
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Yang, H., Zingaro, V.A., Lincoff, J. et al. Remodelling of the translatome controls diet and its impact on tumorigenesis. Nature 633, 189–197 (2024). https://doi.org/10.1038/s41586-024-07781-7
Received:
Accepted:
Published:
Issue date:
DOI: https://doi.org/10.1038/s41586-024-07781-7
This article is cited by
-
Serine metabolism in tumor progression and immunotherapy
Discover Oncology (2025)
-
Therapeutic potential of β-hydroxybutyrate in the management of pancreatic neoplasms: exploring novel diagnostic and treatment strategies
Lipids in Health and Disease (2024)
-
Creating a dietary vulnerability
Nature Reviews Cancer (2024)
-
Clinical research framework proposal for ketogenic metabolic therapy in glioblastoma
BMC Medicine (2024)
