
Abstract
The balance between T helper type 1 (TH1) cells and other TH cells is critical for antiviral and anti-tumour responses1,2,3, but how this balance is achieved remains poorly understood. Here we dissected the dynamic regulation of TH1 cell differentiation during in vitro polarization, and during in vivo differentiation after acute viral infection. We identified regulators modulating T helper cell differentiation using a unique TH1–TH2 cell dichotomous culture system and systematically validated their regulatory functions through multiple in vitro and in vivo CRISPR screens. We found that RAMP3, a component of the receptor for the neuropeptide CGRP (calcitonin gene-related peptide), has a cell-intrinsic role in TH1 cell fate determination. Extracellular CGRP signalling through the receptor RAMP3–CALCRL restricted the differentiation of TH2 cells, but promoted TH1 cell differentiation through the activation of downstream cAMP response element-binding protein (CREB) and activating transcription factor 3 (ATF3). ATF3 promoted TH1 cell differentiation by inducing the expression of Stat1, a key regulator of TH1 cell differentiation. After viral infection, an interaction between CGRP produced by neurons and RAMP3 expressed on T cells enhanced the anti-viral IFNγ-producing TH1 and CD8+ T cell response, and timely control of acute viral infection. Our research identifies a neuroimmune circuit in which neurons participate in T cell fate determination by producing the neuropeptide CGRP during acute viral infection, which acts on RAMP3-expressing T cells to induce an effective anti-viral TH1 cell response.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

Data availability
Sequencing data have been deposited at Gene Expression Omnibus under accession number GSE192728 and are publicly available. Source data are provided with this paper.
Code availability
There are no newly generated codes used in this paper.
References
Kamperschroer, C. & Quinn, D. G. The role of proinflammatory cytokines in wasting disease during lymphocytic choriomeningitis virus infection. J. Immunol. 169, 340–349 (2002).
Kaiko, G. E., Horvat, J. C., Beagley, K. W. & Hansbro, P. M. Immunological decision-making: how does the immune system decide to mount a helper T-cell response? Immunology 123, 326–338 (2008).
Rahimi, K. et al. Targeting the balance of T helper cell responses by curcumin in inflammatory and autoimmune states. Autoimmun. Rev. 18, 738–748 (2019).
Seder, R. A. & Paul, W. E. Acquisition of lymphokine-producing phenotype by CD4+ T cells. Annu. Rev. Immunol. 12, 635–673 (1994).
Swain, S. L., McKinstry, K. K. & Strutt, T. M. Expanding roles for CD4+ T cells in immunity to viruses. Nat. Rev. Immunol. 12, 136–148 (2012).
Varga, S. M. & Welsh, R. M. High frequency of virus-specific interleukin-2-producing CD4+ T cells and Th1 dominance during lymphocytic choriomeningitis virus infection. J. Virol. 74, 4429–4432 (2000).
Dardalhon, V., Korn, T., Kuchroo, V. K. & Anderson, A. C. Role of Th1 and Th17 cells in organ-specific autoimmunity. J. Autoimmun. 31, 252–256 (2008).
Skapenko, A., Leipe, J., Lipsky, P. E. & Schulze-Koops, H. The role of the T cell in autoimmune inflammation. Arthritis Res. Ther. 7, S4 (2005).
Chen, Z. & John Wherry, E. T cell responses in patients with COVID-19. Nat. Rev. Immunol. 20, 529–536 (2020).
Wauters, E. et al. Discriminating mild from critical COVID-19 by innate and adaptive immune single-cell profiling of bronchoalveolar lavages. Cell Res. 31, 272–290 (2021).
Gil-Etayo, F. J. et al. T-helper cell subset response is a determining factor in COVID-19 progression. Front. Cell Infect. Microbiol. 11, 624483 (2021).
Pavel, A. B. et al. Th2/Th1 cytokine imbalance is associated with higher COVID-19 risk mortality. Front. Genet. 12, 706902 (2021).
Wallrapp, A. et al. The neuropeptide NMU amplifies ILC2-driven allergic lung inflammation. Nature 549, 351–356 (2017).
Wallrapp, A. et al. Calcitonin gene-related peptide negatively regulates alarmin-driven type 2 innate lymphoid cell responses. Immunity 51, 709–723 (2019).
Kabata, H. & Artis, D. Neuro-immune crosstalk and allergic inflammation. J. Clin. Invest. 129, 1475–1482 (2019).
Chesné, J., Cardoso, V. & Veiga-Fernandes, H. Neuro-immune regulation of mucosal physiology. Mucosal Immunol. 12, 10–20 (2019).
Jacobson, A., Yang, D., Vella, M. & Chiu, I. M. The intestinal neuro-immune axis: crosstalk between neurons, immune cells, and microbes. Mucosal Immunol. 14, 555–565 (2021).
Enamorado, M. et al. Immunity to the microbiota promotes sensory neuron regeneration. Cell 186, 607–620 (2023).
Lighvani, A. A. et al. T-bet is rapidly induced by interferon-gamma in lymphoid and myeloid cells. Proc. Natl Acad. Sci. USA 98, 15137–15142 (2001).
Afkarian, M. et al. T-bet is a STAT1-induced regulator of IL-12R expression in naïve CD4+ T cells. Nat. Immunol. 3, 549–557 (2002).
Yosef, N. et al. Dynamic regulatory network controlling TH17 cell differentiation. Nature 496, 461–468 (2013).
Zhang, H. et al. An IL-27-driven transcriptional network identifies regulators of IL-10 expression across T helper cell subsets. Cell Rep. 33, 108433 (2020).
Takeda, K. et al. Essential role of Stat6 in IL-4 signalling. Nature 380, 627–630 (1996).
Kaplan, M. H., Schindler, U., Smiley, S. T. & Grusby, M. J. Stat6 is required for mediating responses to IL-4 and for the development of Th2 cells. Immunity 4, 313–319 (1996).
Khatun, A. et al. Single-cell lineage mapping of a diverse virus-specific naive CD4 T cell repertoire. J. Exp. Med. 218, e20200650 (2021).
Hale, J. S. et al. Distinct memory CD4+ T cells with commitment to T follicular helper- and T helper 1-cell lineages are generated after acute viral infection. Immunity 38, 805–817 (2013).
Noble, A., Staynov, D. Z. & Kemeny, D. M. Generation of rat Th2-like cells in vitro is interleukin-4-dependent and inhibited by interferon-gamma. Immunology 79, 562–567 (1993).
Abbas, A. K., Murphy, K. M. & Sher, A. Functional diversity of helper T lymphocytes. Nature 383, 787–793 (1996).
LaFleur, M. W. et al. A CRISPR-Cas9 delivery system for in vivo screening of genes in the immune system. Nat. Commun. 10, 1668 (2019).
Huang, B. et al. In vivo CRISPR screens reveal a HIF-1α-mTOR-network regulates T follicular helper versus Th1 cells. Nat. Commun. 13, 805 (2022).
Hay, D. L., Poyner, D. R. & Sexton, P. M. GPCR modulation by RAMPs. Pharmacol. Ther. 109, 173–197 (2006).
Xu, H. et al. Transcriptional atlas of intestinal immune cells reveals that neuropeptide α-CGRP modulates group 2 innate lymphoid cell responses. Immunity 51, 696–708 (2019).
Nagashima, H. et al. Neuropeptide CGRP limits group 2 innate lymphoid cell responses and constrains type 2 inflammation. Immunity 51, 682–695 (2019).
Zhang, S. J. et al. A signaling cascade of nuclear calcium-CREB-ATF3 activated by synaptic NMDA receptors defines a gene repression module that protects against extrasynaptic NMDA receptor-induced neuronal cell death and ischemic brain damage. J. Neurosci. 31, 4978–4990 (2011).
Kim, J. Y. et al. A critical role of STAT1 in streptozotocin-induced diabetic liver injury in mice: controlled by ATF3. Cell Signal. 21, 1758–1767 (2009).
Baral, P. et al. Nociceptor sensory neurons suppress neutrophil and γδ T cell responses in bacterial lung infections and lethal pneumonia. Nat. Med. 24, 417–426 (2018).
Garelja, M. L. et al. Pharmacological characterisation of mouse calcitonin and calcitonin receptor-like receptors reveals differences compared with human receptors. Br. J. Pharmacol. 179, 416–434 (2022).
Chu, H. M., Tan, Y., Kobierski, L. A., Balsam, L. B. & Comb, M. J. Activating transcription factor-3 stimulates 3′,5′-cyclic adenosine monophosphate-dependent gene expression. Mol. Endocrinol. 8, 59–68 (1994).
Chen, W. J., Lai, Y. J., Lee, J. L., Wu, S. T. & Hsu, Y. J. CREB/ATF3 signaling mediates indoxyl sulfate-induced vascular smooth muscle cell proliferation and neointimal formation in uremia. Atherosclerosis 315, 43–54 (2020).
Ma, W. et al. Localization and modulation of calcitonin gene-related peptide-receptor component protein-immunoreactive cells in the rat central and peripheral nervous systems. Neuroscience 120, 677–694 (2003).
Assas, B. M., Pennock, J. I. & Miyan, J. A. Calcitonin gene-related peptide is a key neurotransmitter in the neuro-immune axis. Front. Neurosci. 8, 23 (2014).
Chiu, I. M. et al. Bacteria activate sensory neurons that modulate pain and inflammation. Nature 501, 52–57 (2013).
Sui, P. et al. Pulmonary neuroendocrine cells amplify allergic asthma responses. Science 360, eaan8546 (2018).
Hanc, P. et al. Multimodal control of dendritic cell functions by nociceptors. Science 379, eabm5658 (2023).
Ding, W., Stohl, L. L., Wagner, J. A. & Granstein, R. D. Calcitonin gene-related peptide biases Langerhans cells toward Th2-type immunity. J. Immunol. 181, 6020–6026 (2008).
Edvinsson, L. CGRP antibodies as prophylaxis in migraine. Cell 175, 1719 (2018).
Assas, M. B. Anti-migraine agents from an immunological point of view. J. Transl. Med. 19, 23 (2021).
Mitsikostas, D. D. & Reuter, U. Calcitonin gene-related peptide monoclonal antibodies for migraine prevention: comparisons across randomized controlled studies. Curr. Opin. Neurol. 30, 272–280 (2017).
Wehbi, V. L. & Taskén, K. Molecular mechanisms for cAMP-mediated immunoregulation in T cells—role of anchored protein kinase A signaling units. Front. Immunol. 7, 222 (2016).
Boudard, F. & Bastide, M. Inhibition of mouse T-cell proliferation by CGRP and VIP: effects of these neuropeptides on IL-2 production and cAMP synthesis. J. Neurosci. Res. 29, 29–41 (1991).
Oh-hashi, Y. et al. Elevated sympathetic nervous activity in mice deficient in alphaCGRP. Circ. Res. 89, 983–990 (2001).
Tsujikawa, K. et al. Hypertension and dysregulated proinflammatory cytokine production in receptor activity-modifying protein 1-deficient mice. Proc. Natl Acad. Sci. USA 104, 16702–16707 (2007).
Dackor, R., Fritz-Six, K., Smithies, O. & Caron, K. Receptor activity-modifying proteins 2 and 3 have distinct physiological functions from embryogenesis to old age. J. Biol. Chem. 282, 18094–18099 (2007).
Picelli, S. et al. Full-length RNA-seq from single cells using Smart-seq2. Nat. Protoc. 9, 171–181 (2014).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 9, 357–359 (2012).
Trapnell, C. et al. Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 28, 511–515 (2010).
Love, M. I., Huber, W. & Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550 (2014).
Chen, E. Y. et al. Enrichr: interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 14, 128 (2013).
Kuleshov, M. V. et al. Enrichr: a comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 44, W90–W97 (2016).
Xie, Z. et al. Gene set knowledge discovery with Enrichr. Curr. Protoc. 1, e90 (2021).
Mootha, V. K. et al. PGC-1α-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 34, 267–273 (2003).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl Acad. Sci. USA 102, 15545–15550 (2005).
Zheng, G. X. et al. Massively parallel digital transcriptional profiling of single cells. Nat. Commun. 8, 14049 (2017).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587 (2021).
Waltman, L. & van Eck, N. J. A smart local moving algorithm for large-scale modularity-based community detection. Eur. Phys. J. B 86, 471 (2013).
Street, K. et al. Slingshot: cell lineage and pseudotime inference for single-cell transcriptomics. BMC Genom. 19, 477 (2018).
Corces, M. R. et al. An improved ATAC-seq protocol reduces background and enables interrogation of frozen tissues. Nat. Methods 14, 959–962 (2017).
Li, H. et al. The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079 (2009).
Zhang, Y. et al. Model-Based Analysis of ChIP-Seq (MACS). Genome Biol. 9, R137 (2008).
Robinson, J. T. et al. Integrative Genomics Viewer. Nat. Biotechnol. 29, 24–26 (2011).
Lun, A. T. & Smyth, G. K. csaw: a Bioconductor package for differential binding analysis of ChIP-seq data using sliding windows. Nucleic Acids Res. 44, e45 (2016).
Amemiya, H. M., Kundaje, A. & Boyle, A. P. The ENCODE blacklist: identification of problematic regions of the genome. Sci. Rep. 9, 9354 (2019).
Chen, Y., Lun, A. & Smyth, G. From reads to genes to pathways: differential expression analysis of RNA-Seq experiments using Rsubread and the edgeR quasi-likelihood pipeline. F1000Research 5, 1438 (2016).
Yu, G., Wang, L.-G. & He, Q.-Y. ChIPseeker: an R/Bioconductor package for ChIP peak annotation, comparison and visualization. Bioinformatics 31, 2382–2383 (2015).
Bioconductor Core Team & Bioconductor Package Maintainer. TxDb.Mmusculus.UCSC.mm10.knownGene: annotation package for TxDb object(s). R package version 3.4.7 (Bioconductor, 2019).
Carlson, M. org.Mm.eg.db: genome wide annotation for Mouse. R package version 3.8.2 (Bioconductor, 2019).
Kaya-Okur, H. S. et al. CUT&Tag for efficient epigenomic profiling of small samples and single cells. Nat. Commun. 10, 1930 (2019).
Olah, Z. et al. Ligand-induced dynamic membrane changes and cell deletion conferred by vanilloid receptor 1. J. Biol. Chem. 276, 11021–11030 (2001).
Acknowledgements
We acknowledge all of the members of the Kuchroo laboratory for inputs. The backbone construct for CRISPR screening was provided by the Genetic Perturbation Platform at the Broad Institute. IAb–GP66–77 and H-2Db–GP33–41 tetramers were provided by the NIH Tetramer Core Facility. V.K.K., A.H.S., A.R., J.G.D, and O.A. were supported by NIH grant U19AI133524. V.K.K. was supported by grant P01AI039671, P01AI056299, R01AI144166 and P01AI073748. V.K.K. and A.R. are supported by the Klarman Cell Observatory at the Broad Institute. I.M.C. was supported by NIH R01AI168005 and R01DK127257.
Author information
Authors and Affiliations
Contributions
Y.H., L.S., A.H.S., A.R. and V.K.K. conceived the study and wrote the manuscript with input from all of the authors. Y.H., L.S., A.H.S., A.R., I.M.C. and V.K.K. designed the experiments and interpreted the results. Y.H. and M.W.L. performed the screening experiments with the assistance from P.I.T.; K.G.-S. designed and generated the sgRNA library for screening. K.K., R.B., L.Y., Y.Z., R.T. and J.S. helped with in vitro experiments. A.W., H.S.C., L.D. and Y.-C.K. helped with neuronal and LCMV infection experiments. All sequencing data were analysed by Y.H., L.H. and C.L. with the assistance and guidance from A.H.S., O.A., G.S., A.R., Y.H. and J.G.D.
Corresponding authors
Ethics declarations
Competing interests
V.K.K. has an ownership interest in Tizona Therapeutics, Bicara Therapeutics, Larkspur Biosciences and Werewolf Therapeutics; financial interests in Biocon Biologics, Perkin Elmer, Elpiscience Biopharmaceutical, Equilium and Syngene. A.H.S. has patents/pending royalties on the PD-1 pathway from Roche and Novartis; is on advisory boards for Surface Oncology, SQZ Biotechnologies, Elpiscience, Selecta, Bicara Therapeutics, Monopteros, Fibrogen, Alixis, GlaxoSmithKline and Janssen; and has received research funding from Merck, Vertex, Moderna, Quark/Iome and AbbVie (unrelated to this project). A.R. is listed as a co-inventor on patent applications filed by the Broad Institute for inventions related to single-cell genomics; is an equity holder in Immunitas; and was a scientific advisory board member of Thermo Fisher Scientific, Syros Pharmaceuticals, Neogene Therapeutics and Asimov until 31 July 2020. From 1 August 2020, A.R. is an employee of Genentech and has equity in Roche. The other authors declare no competing interests.
Peer review
Peer review information
Nature thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Dynamic transcriptional regulation during Th1 differentiation.
(a) Expression dynamics of Th1 or Th2 signature genes. Cells were collected at the indicated time points during 3 days of in vitro culture for RNA extraction and qPCR. Two-tailed Student’s t test. Data are presented as mean +/− SD. n = 3, samples pooled by ≥ 3 mice. (b) Schematic of experimental design for temporal bulk RNA-seq of in vitro cultured T helper cells. (c) Expression dynamics of the signature genes of naïve, Th0, Th1 and Th2 cells detected in bulk RNA-seq and each showed distinct expression patterns and kinetics across the three conditions. (d) Three major dynamic phases at global transcriptome level during T helper cell in vitro differentiation. Spearman correlation matrix across collected time points is shown as a heatmap. (e) Relative expression of dynamically expressed genes during in vitro Th2 differentiation. (f) Top 1000 genes contributing to PC2 of early, intermediate, and late phase in Fig. 1b. (g) Heterogeneity among CD4+ T cells at day 8 of Armstrong infection. Cells are coloured by the relative expression of signature genes. (h) Frequency of each cell type in activated CD4+ T cells during LCMV Armstrong infection. Results are from one single wild type mouse. (i) Global lineage structure inferred by Slingshot using minimum spanning tree on the clusters of interest. (j) Z-normalized expression for top 50 marker genes for Naïve, Tcmp, Early active and Th1 cell clusters are shown in heatmap (FDR < 0.05, fold change > 1.2, rank by FDR increasingly; ribosomal protein small/large subunit (Rps-/Rpl-) genes or mitochondrial (Mt-) genes were excluded). Genes overlapped with in vitro dynamics gene clusters (Fig. 1a) were marked on the right. Enrichment p-value was calculated using Chi-squared test.
Extended Data Fig. 2 Determination of Th1 unique regulators under Th1/Th2 dichotomous culture condition.
(a) Modification of Th1/Th2 dichotomous culture condition by titrating cytokines (n = 3). (b) Th1/Th2 balance is not related to TCR clonality. Similar percentages of IFNγ+ and IL-13+ cells were detected in wild type and OT-II TCR transgenic cells under dichotomous condition. (n = 6, samples pooled by ≥ 3 mice per group). (c) UMAP embedding of the cells collected at 64 h from two technical replicates. Cells (dots) are coloured by clusters, technical replicates, cell cycle and relative expression of naïve, Th1 or Th2 signature genes. (d) Among the cells collected from 64 h, there is exclusive expression between Ifng and Il13 (top), between Th1 and Th2 program (bottom). (e) Differentially expressed genes between defined Th1 and Th2 cells at 64 h are shown with their expression profiles in each single cell as a heatmap. Two-tailed Student’s t test. Data are presented as mean +/− SD. n = X, samples pooled by ≥ 3 mice per group.
Extended Data Fig. 3 Validation of the function of Th1 regulator candidates using pooled genetic screens.
(a) The performance of in vitro gene perturbation strategy was evaluated by targeting positive control genes Tbx21 and Gata3 in Th1 and Th2 cells respectively. Naïve CD4+ T cells from Rosa26-Cas9 mice were activated for 24 h (anti-CD3 and anti-CD28), transduced with lentiviral-sgRNA and rested for 2 days with IL-2 for gene perturbation. The effect of each sgRNA was subsequently evaluated after 2 days in vitro Th1 (or Th2) polarization. Two-tailed Student’s t test. Data are presented as mean +/− SD. n = 4, samples pooled by ≥ 3 mice. (b) Technical replicates show high correlation of sgRNA distribution. From in vivo screening, total activated (CD44+) cell population was sorted as control sample and randomly split into two fractions for following steps to assess technical variations. (c) The distribution of sequencing depth for each guide in sgRNA library. (d) The distribution of sequencing depth for each guide in Naïve CD4+ T cells from in vivo screen. (e-g) The distribution of sequencing depth for each guide from in vitro screen of (e) Th1 condition, (f) Th2 condition, and (g) CD8 condition. (h) Sorted cell populations with similar cytokine expression show higher correlations of global sgRNA abundance. Global Pearson correlations were calculated between sorted cell populations from ex vivo screening based on the normalized sgRNA sequencing depth. (i) The recovery of sgRNAs from each screening setting. The value on the right of each row represents the number and percentage of sgRNAs recovered (>50X sequencing depth) in each screening. pDNA represents the plasmid of the cloned sgRNA library. (j) All detected sgRNAs were ranked with their unique functions in regulating IFNγ expression. Delta LFC value was calculated for each sgRNA in each screening setting by comparing LFC value in distinct sorted cell populations. sgRNAs ranking in the top with lower delta LFC values in Th1 screenings are IFNγ positive regulators, while sgRNAs ranking in the bottom with higher delta LFC values in Th1 screenings are IFNγ negative regulators. (k) All screen candidates were ranked based on their unique function in regulating IFNγ expression. Top-ranking IFNγ positive regulators (top left 30 genes) and negative regulators (top right 20 genes) are shown in the heatmap with their sgRNA enrichment score in distinct screening settings.
Extended Data Fig. 4 RAMP3 function as a neuropeptide receptor in T cells.
(a) Expression levels of RAMP family in ILC2 cells using data set from GSE136154 (n = 4). (b) The expression of RAMP family genes in CD4+ T cells after 4 h in vitro activation with anti-CD3 and anti-CD28. Cells were collected for RNA extraction to assess the expression of receptors (Ramp1, Ramp2, Ramp3) with RT-qPCR (n = 6). (c) The expression levels of RAMP family in CD8+ T cells after 4 h in vitro activation with anti-CD3 and anti-CD28. Cells were collected for RNA extraction to assess the expression of receptors (Ramp1, Ramp2, Ramp3) with RT-qPCR (n = 6). (d) Expression dynamics of Ramp3 during T cell in vitro differentiation under Th0, Th1 and Th2 conditions. (e) Expression pattern of GPCRs during T helper cell differentiation. (f) Pearson correlation between the expression of Ramp3 and each GPCR gene during T cell in vitro differentiation. (g) Calcrl gene expression shows similar expression dynamics as Ramp3 during Th1 in vitro differentiation. (h) Ramp3 sgRNAs are further depleted in IFNγhigh cells and IL-13low/neg cells from both in vitro screens.
Extended Data Fig. 5 Neuropeptide CGRP and ADM modulate T cell differentiation through Ramp3.
(a, b) CGRP and ADM promote Th1 / CD8+ T cell differentiation but antagonize Th2 differentiation. (a) Naïve CD8+ T cells were in vitro differentiated under Tc0 condition (anti-CD3 and anti-CD28) with / without CGRP Representative flow cytometry left panel, quantitation, right panel (n = 4). (b) ADM treatment show similar effects as CGRP in both CD4+ and CD8+ T cells (n = 4-5). (c) CGRP and ADM also inhibit IL-5 expression under Th2 condition after 4 days in vitro culture. Representative flow cytometry, left, quantitation, right (n = 8). (d) Naïve CD4+ T cells were cultured with Th1 / Th2 dichotomous condition for 3 days before analysing cytokine expression by flow cytometry, (representative flow cytometry, left panel, quantitation, right panel) (n = 5). (e) Cytokine secretion was analysed with LegendPlex after 3 days Th1 / Th2 dichotomous culture (n = 5). (f) CGRP induces the expression of Calca and Ramp family in CD4+ T cells. Isolated naïve CD4+ T cells were treated with the neuropeptide for 4 h and collected for qPCR analysis (n = 6). (g) Isoform specific expression analyses of Calca detected with qPCR in T cells (n = 4). (h) Calcitonin is detected in T cell lysate upon CGRP treatment for 72 h (n = 4). (i) The promotion of Th1 differentiation by CGRP is abrogated by BIBN4096BS (n = 8). (j). The promotion of Th1 differentiation by CGRP is abrogated in Calcrl knockout T cells (n = 5). (k) Gene expressions of Calcrl were detected by qPCR (n = 3). Two-tailed Student’s t test. Data are presented as mean +/− SD. n = X mice (h) / samples pooled by ≥ 3 mice (other panels) per group. Results shown are from one experiment, representative of at least three independent experiments.
Extended Data Fig. 6 Neuropeptide CGRP and ADM enhance Th1 differentiation through cAMP signalling.
(a-b) CGRP and ADM induce cAMP signalling in T cells. (a) Differentially expressed genes between CGRP and Vehicle group (after 4 h treatment) are presented in the heatmap. (b) Gene ontology enrichment analysis was performed with the upregulated genes in CGRP treatment group. (c) CGRP and ADM induce intracellular cAMP accumulation in CD4+ T cells. Naïve T cells were treated with CGRP or ADM for 30 min and intracellular cAMP was measured using ELISA (n = 3). (d-e) cAMP is involved in the CGRP-Ramp3 positive feedback loop. The expression of Ramp3 (d) and Calca (e) are significantly induced upon dbcAMP treatment for 4 h (n = 5). (f) Well differentiated T cells are less responsive to the CGRP / ADM treatment. Fold change of Calca, Ramp3 and Crem expression relative to control (n = 4). (g) dbcAMP treatment recapitulates CGRP and ADM effects in promoting IFNγ expression in CD8+ T cells. Cells were collected for intracellular cytokine staining after 3 days in vitro culture. Representative flow cytometry left panel, quantitation, right panel (n = 5). (h) Enhancement of Th1 differentiation by ADM is abrogated with CREB inhibitor 666-15 (50 μM). Cells were collected for intracellular cytokine staining after 3 days in vitro culture. Representative flow cytometry left panel, quantitation, right panel (n = 4-5). (i-j) Enhancement of Th1 differentiation by CGRP (i) and ADM (j) is blocked with adenylyl cyclase inhibitor SQ22536 (20 μM). Cells were collected for intracellular cytokine staining after 3 days in vitro culture (n = 7-8). Two-tailed Student’s t test. Data are presented as mean +/− SD (c-j). n = X samples pooled by ≥ 3 mice. Results shown are from one experiment, representative of at least three independent experiments.
Extended Data Fig. 7 CGRP-Ramp3-cAMP loop enhances Th1 differentiation through epigenomic remodelling and activating STAT1.
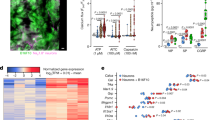
(a) Th1 pioneer program was induced by CGRP and ADM. Isolated naïve CD4+ T cells were treated with CGRP or ADM for 4 h and collected for qPCR analysis (n = 2-6). (b) Th1 pioneer program was induced by cAMP in a dose dependent manner. Isolated naïve CD4+ T cells were treated with dbcAMP for 4 h and collected for qPCR analysis (n = 4-5). (c) Categories of all cis-element open chromatin regions detected in ATAC-seq data of Th1 and Th2 cells. (d) Shared and unique Th1/Th2 differentially accessible chromatin regions were identified in the CGRP treated and control group. (e) Categories of differential cis-element open chromatin regions between Th1 and Th2 cells with or without CGRP treatment. (f) Overlapping of the genes proximal to differentially accessible regions and genes with differential expression between Th1 and Th2 cells. (g) CREB binding signals from CUT&Tag assay on in vitro differentiated Th1 cells. (h) Immunostaining of pSTAT1 in differentiated Th1 cells upon CGRP treatment for 8 h. Scale bar: 10 μm. Signal intensity of pSTAT1 is quantified (n = 36-159 sections from 2-3 mice). (i) The enhancement of Th1 differentiation by CGRP/ADMs is abrogated in Atf3 knockout T cells (n = 3). (j) Luciferase activity in 293 T cells transfected with luciferase reporters for the indicated cis-regulatory elements of Stat1 and plasmids encoding ATF3 (n = 3). Two-tailed Student’s t test. Data are presented as mean +/− SD (a, b and h-j). n = X mice (i) / samples (a, b and j) pooled by ≥ 3 mice per group. (k) Model graph of CGRP-RAMP3 axis in modulating T cell differentiation. Two-tailed Student’s t test. Data are presented as mean +/− SD. n = X samples pooled by ≥ 3 mice. Results shown are from one experiment, representative of at least three independent experiments.
Extended Data Fig. 8 CGRP enhances antigen-specific Th1 and Tc1 response during LCMV Armstrong infection.
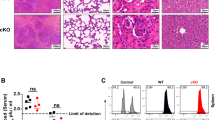
(a) CGRP in spleen is increased upon LCMV Armstrong infection. CGRP concentration in spleen homogenate was detected using ELISA (n = 3-4). (b) Th1 response was restrained in Calca−/− mice during LCMV Armstrong infection. Splenocytes were collected at day 8 post Armstrong infection, followed by restimulation with PMA / Ionomycin for intracellular cytokine expression analyses within total CD44+CD4+ T cells. Representative flow cytometry, left panel, quantitation, right (WT: n = 4; Calca−/−: n = 3). (c, d) IFNγ expression was restrained in CD8+ T cells of Calca−/− mice. Splenocytes were collected at Day8 post Armstrong infection, followed by restimulation with PMA / Ionomycin for another 4 h for intracellular cytokine staining within (c) total CD44+CD8+ T cells and (d) antigen specific CD8+ T cells (LCMV GP33-41 tetramer+). Representative flow cytometry, left panel, quantitation, right panel (WT: n = 4; Calca−/−: n = 3). (e) Gating strategy of distinct cell types in spleen upon LCMV Armstrong infection. (f) Flow cytometric quantification of immune cells in spleen from Calca−/− and WT mice at day 8 post LCMV Armstrong infection (WT: n = 4; Calca−/−: n = 3). (g) Calca−/− mice show higher weight of spleen (WT: n = 4; Calca−/−: n = 5). (h) CGRP is required for effective clearance of LCMV infection. Viral load was detected in serum from each mouse upon LCMV Armstrong infection. Centre line at the median, upper bound at 75th percentile, lower bound at 25th percentile, Whiskers extend to the minimum and maximum values (WT: n = 3; Calca−/−: n = 6). (i) Co-localization of beta-III Tubulin and CGRP in the spleen of WT and Calca−/− mice. Scale bar, 10 μm. (j) Immunostaining of CD3, CGRP and beta-III Tubulin in mouse spleen upon LCMV infection. Scale bar, 10 μm. Arrows indicates that T cells are close to CGRP+ neurons in the spleen. Percentage of CGRP+ among beta-III tubulin+ neurons that are close to CD3+ T cells (< 5 μm) are quantified (n = 19-28 sections from 2-3 mice). (k) Gene expressions of Calca and Trpv1 were detected in WT DRG (Vehicle: n = 3; RTX: n = 3). (l) Gene expressions of Trpv1 were detected in Calca−/− DRG (Vehicle: n = 3; RTX: n = 3). Mann-Whitney test (h); Two-tailed Student’s t test (other panels). Data are presented as mean +/− SD (b-d, f, g, j-l). n =X mice per group. Results shown are from one experiment, representative of at least three independent experiments.
Extended Data Fig. 9 CGRP-RAMP3 axis enhances antigen-specific Th1 and Tc1 response during LCMV Armstrong infection.
(a) At steady state, there is no difference on Th1 or Tc1 differentiation between Ramp3−/− and WT mice (WT: n = 5; Ramp3−/−: n = 3). (b) At steady state, Ramp3−/− mice have similar spleen size as WT mice (WT: n = 5; Ramp3−/−: n = 3). (c) Th1 response was restrained in Ramp3−/− mice upon LCMV Armstrong infection without affecting cell activation. At day 8 post Armstrong infection, splenocytes were restimulated with PMA / Ionomycin for intracellular cytokine expression analyses within CD4+ T cells. Representative flow cytometry, left, quantitation, right (WT: n = 3; Ramp3−/−: n = 4). (d) Tc1 response was restrained in CD8+ T cells of Ramp3−/− mice upon LCMV Armstrong infection without affecting cell activation. At Day8 post Armstrong infection, splenocytes were restimulated with PMA / Ionomycin for intracellular cytokine expression analyses within total CD44+CD8+ T cells. Representative flow cytometry, left, quantitation, right (WT: n = 3; Ramp3−/−: n = 4). (e) Antigen-specific Th1 response was restrained in Ramp3−/− mice upon LCMV Armstrong infection. At day 8 post Armstrong infection, splenocytes were restimulated with PMA / Ionomycin for intracellular cytokine expression analyses within antigen-specific CD4+ T cells (LCMV GP66-77 tetramer+). Representative flow cytometry, left panel, quantitation, right (WT: n = 3; Ramp3−/−: n = 4). (f) Tc1 response was restrained in CD8+ T cells of Ramp3−/− mice upon LCMV Armstrong infection. At Day8 post Armstrong infection, splenocytes were restimulated with PMA / Ionomycin for intracellular cytokine expression analyses within antigen-specific CD8+ T cells (LCMV GP33-41 tetramer+). Representative flow cytometry, left, quantitation, right (WT: n = 3; Ramp3−/−: n = 4). (g) Antigen-specific Th1 response was reduced in Ramp3−/− mice during LCMV Armstrong infection. Splenocytes were collected from spleen at Day 8 post Armstrong infection, followed by restimulation with LCMV GP61-80 for another 4 h for intracellular cytokine staining. Representative flow cytometry, left panel, quantitation, right (WT: n = 3; Ramp3−/−: n = 4). (h) Ramp3−/− mice show higher weight of spleen (WT: n = 4; Ramp3−/−: n = 5). (i) RAMP1 did not show function in regulating T cell response during LCMV Armstrong infection. At Day8 post Armstrong infection, splenocytes were restimulated with PMA / Ionomycin (left) or LCMV GP61-80 / LCMV GP33-41 (right) for intracellular cytokine expression analyses (WT: n = 5; Ramp1−/−: n = 4). Two-tailed Student’s t test. Data are presented as mean +/− SD. n = X mice per group. Results shown are from one experiment, representative of at least three independent experiments.
Supplementary information
Supplementary Fig. 1
Source images for western blotting and the flow cytometry sequential gating/sorting strategies.
Supplementary Tables
Supplementary Tables 1–9.
Source data
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Hou, Y., Sun, L., LaFleur, M.W. et al. Neuropeptide signalling orchestrates T cell differentiation. Nature 635, 444–452 (2024). https://doi.org/10.1038/s41586-024-08049-w
Received:
Accepted:
Published:
Issue date:
DOI: https://doi.org/10.1038/s41586-024-08049-w
This article is cited by
-
Targeting microglia-Th17 feed-forward loop to suppress autoimmune neuroinflammation
Journal of Neuroinflammation (2025)
-
Neurotransmitters: an emerging target for therapeutic resistance to tumor immune checkpoint inhibitors
Molecular Cancer (2025)
-
Pro-repair macrophages driven by CGRP rescue white matter integrity following intracerebral hemorrhage
Journal of Neuroinflammation (2025)
-
Integrating neuroscience and oncology: neuroimmune crosstalk in the initiation and progression of digestive system tumors
Molecular Cancer (2025)
-
Connection and communication between the nervous and immune systems
Nature Reviews Immunology (2025)
