
Abstract
Cholera toxin (CT) is the etiological agent of cholera. Here we report that multiple classes of fucosylated glycoconjugates function in CT binding and intoxication of intestinal epithelial cells. In Colo205 cells, knockout (KO) of B3GNT5, which encodes an enzyme required for synthesis of lacto and neolacto series glycosphingolipids (GSLs), reduces CT binding but sensitizes cells to intoxication. Overexpressing B3GNT5 to generate more fucosylated GSLs confers protection against intoxication, indicating that fucosylated GSLs act as decoy receptors for CT. KO of B3GALT5 causes increased production of fucosylated O-linked and N-linked glycoproteins and leads to increased CT binding and intoxication. KO of B3GNT5 in B3GALT5-KO cells eliminates production of fucosylated GSLs but increases intoxication, identifying fucosylated glycoproteins as functional receptors for CT. These findings provide insight into the molecular determinants regulating CT sensitivity of host cells.

This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others

Data availability
Data used to generate Fig. 1b,d are available from the Gene Expression Omnibus under accession number GSE242156. MS data used to generate Fig. 3a, Extended Data Fig. 4a–d, Fig. 4a, Extended Data Fig. 7a–c, Extended Data Fig. 8a–c and Extended Data Fig. 6a are available from GlycoPOST under accession number GPST000467. MS data used to generate Extended Data Fig. 6b,c are available from GlycoPOST under accession number GPST000465. Replicate immunoblot and lectin blot data are available from the Texas Data Repository (https://doi.org/10.18738/T8/N9NF8B). Source data are provided with this paper.
References
Grant, T. A., Balasubramanian, D. & Almagro-Moreno, S. JMM profile: Vibrio cholerae: an opportunist of human crises. J. Med. Microbiol. 70, 001423 (2021).
White, C., Bader, C. & Teter, K. The manipulation of cell signaling and host cell biology by cholera toxin. Cell Signal. 100, 110489 (2022).
Cuatrecasas, P. Gangliosides and membrane receptors for cholera toxin. Biochemistry 12, 3558–3566 (1973).
Cuatrecasas, P., Parikh, I. & Hollenberg, M. D. Affinity chromatography and structural analysis of Vibrio cholerae enterotoxin–ganglioside agarose and the biological effects of ganglioside-containing soluble polymers. Biochemistry 12, 4253–4264 (1973).
Heyningen, S. V. Cholera toxin: interaction of subunits with ganglioside GM1. Science 183, 656–657 (1974).
Holmgren, J., Lonnroth, I., Mansson, J. & Svennerholm, L. Interaction of cholera toxin and membrane GM1 ganglioside of small intestine. Proc. Natl Acad. Sci. USA 72, 2520–2524 (1975).
King, C. A. & Van Heyningen, W. E. Deactivation of cholera toxin by a sialidase-resistant monosialosylganglioside. J. Infect. Dis. 127, 639–647 (1973).
Holmgren, J., Lönnroth, I. & Svennerholm, L. Tissue receptor for cholera exotoxin: postulated structure from studies with GM1 ganglioside and related glycolipids. Infect. Immun. 8, 208–214 (1973).
Merritt, E. A. et al. Structural studies of receptor binding by cholera toxin mutants. Protein Sci. 6, 1516–1528 (1997).
Merritt, E. A. et al. Crystal structure of cholera toxin B-pentamer bound to receptor GM1 pentasaccharide. Protein Sci. 3, 166–175 (1994).
Turnbull, W. B., Precious, B. L. & Homans, S. W. Dissecting the cholera toxin–ganglioside GM1 interaction by isothermal titration calorimetry. J. Am. Chem. Soc. 126, 1047–1054 (2004).
Cervin, J. et al. GM1 ganglioside-independent intoxication by cholera toxin. PLoS Pathog. 14, e1006862 (2018).
Breimer, M. E., Hansson, G. C., Karlsson, K. A., Larson, G. & Leffler, H. Glycosphingolipid composition of epithelial cells isolated along the villus axis of small intestine of a single human individual. Glycobiology 22, 1721–1730 (2012).
Alisson-Silva, F. et al. Human evolutionary loss of epithelial Neu5Gc expression and species-specific susceptibility to cholera. PLoS Pathog. 14, e1007133 (2018).
Glass, R. I. et al. Predisposition for cholera of individuals with O blood group. Possible evolutionary significance. Am. J. Epidemiol. 121, 791–796 (1985).
Barua, D. & Paguio, A. S. ABO blood groups and cholera. Ann. Hum. Biol. 4, 489–492 (1977).
Chaudhuri, A. & De, S. Cholera and blood-groups. Lancet 2, 404 (1977).
Swerdlow, D. L. et al. Severe life-threatening cholera associated with blood group O in Peru: implications for the Latin American epidemic. J. Infect. Dis. 170, 468–472 (1994).
Harris, J. B. et al. Blood group, immunity, and risk of infection with Vibrio cholerae in an area of endemicity. Infect. Immun. 73, 7422–7427 (2005).
Harris, J. B. et al. Susceptibility to Vibrio cholerae infection in a cohort of household contacts of patients with cholera in Bangladesh. PLoS Negl. Trop. Dis. 2, e221 (2008).
Heggelund, J. E. et al. High-resolution crystal structures elucidate the molecular basis of cholera blood group dependence. PLoS Pathog. 12, e1005567 (2016).
Heggelund, J. E. et al. Both El Tor and classical cholera toxin bind blood group determinants. Biochem. Biophys. Res. Commun. 418, 731–735 (2012).
Holmner, A. et al. Novel binding site identified in a hybrid between cholera toxin and heat-labile enterotoxin: 1.9 Å crystal structure reveals the details. Structure 12, 1655–1667 (2004).
Bennun, F. R., Roth, G. A., Monferran, C. G. & Cumar, F. A. Binding of cholera toxin to pig intestinal mucosa glycosphingolipids: relationship with the ABO blood group system. Infect. Immun. 57, 969–974 (1989).
Wands, A. M. et al. Fucosylated molecules competitively interfere with cholera toxin binding to host cells. ACS Infect. Dis. 4, 758–770 (2018).
Prudden, A. R. et al. Synthesis of asymmetrical multiantennary human milk oligosaccharides. Proc. Natl Acad. Sci. USA 114, 6954–6959 (2017).
Heim, J. B., Hodnik, V., Heggelund, J. E., Anderluh, G. & Krengel, U. Crystal structures of cholera toxin in complex with fucosylated receptors point to importance of secondary binding site. Sci. Rep. 9, 12243 (2019).
Garber, J. M., Hennet, T. & Szymanski, C. M. Significance of fucose in intestinal health and disease. Mol. Microbiol. 115, 1086–1093 (2021).
Wands, A. M. et al. Fucosylation and protein glycosylation create functional receptors for cholera toxin. eLife 4, e09545 (2015).
Blanco, L. P. & DiRita, V. J. Bacterial-associated cholera toxin and GM1 binding are required for transcytosis of classical biotype Vibrio cholerae through an in vitro M cell model system. Cell Microbiol 8, 982–998 (2006).
Sethi, A. et al. Cell type and receptor identity regulate cholera toxin subunit B (CTB) internalization. Interface Focus 9, 20180076 (2019).
Sojitra, M. et al. Genetically encoded multivalent liquid glycan array displayed on M13 bacteriophage. Nat. Chem. Biol. 17, 806–816 (2021).
Yanagisawa, M., Ariga, T. & Yu, R. K. Cholera toxin B subunit binding does not correlate with GM1 expression: a study using mouse embryonic neural precursor cells. Glycobiology 16, 19g–22g (2006).
Bakker, H., Ashikov, A., Routier, F. H. & Gerardy-Schahn, R. in Handbook of Glycosyltransferases and Related Genes Ch. 124 (eds Taniguchi, N. et al.) 1403–1412 (Springer, 2014).
Foulquier, F. et al. TMEM165 deficiency causes a congenital disorder of glycosylation. Am. J. Hum. Genet 91, 15–26 (2012).
Togayachi, A. & Narimatsu, H. in Handbook of Glycosyltransferases and Related Genes Ch. 29 (eds Taniguchi, N. et al.) 311–320 (Springer, 2014).
Togayachi, A. & Narimatsu, H. in Handbook of Glycosyltransferases and Related Genes Ch. 9 (eds Taniguchi, N. et al.) 89–100 (Springer, 2014).
Lin, C. H. et al. Enhanced expression of β3-galactosyltransferase 5 activity is sufficient to induce in vivo synthesis of extended type 1 chains on lactosylceramides of selected human colonic carcinoma cell lines. Glycobiology 19, 418–427 (2009).
Lin, R. J. et al. B3GALT5 knockout alters gycosphingolipid profile and facilitates transition to human naive pluripotency. Proc. Natl Acad. Sci. USA 117, 27435–27444 (2020).
Isshiki, S. et al. Cloning, expression, and characterization of a novel UDP-galactose: β-N-acetylglucosamine β-galactosyltransferase (β3Gal-T5) responsible for synthesis of type 1 chain in colorectal and pancreatic epithelia and tumor cells derived therefrom. J. Biol. Chem. 274, 12499–12507 (1999).
Groth, T., Diehl, A. D., Gunawan, R. & Neelamegham, S. GlycoEnzOnto: a glycoenzyme pathway and molecular function ontology. Bioinformatics 38, 5413–5420 (2022).
Ancheta, L. R., Shramm, P. A., Bouajram, R., Higgins, D. & Lappi, D. A. Streptavidin–saporin: converting biotinylated materials into targeted toxins. Toxins 15, 181 (2023).
Stirpe, F. et al. Ribosome-inactivating proteins from the seeds of Saponaria officinalis L. (soapwort), of Agrostemma githago L. (corn cockle) and of Asparagus officinalis L. (asparagus), and from the latex of Hura crepitans L. (sandbox tree). Biochem. J. 216, 617–625 (1983).
Lee, L., Abe, A. & Shayman, J. A. Improved inhibitors of glucosylceramide synthase. J. Biol. Chem. 274, 14662–14669 (1999).
Singla, A. et al. Cholera intoxication of human enteroids reveals interplay between decoy and functional glycoconjugate ligands. Glycobiology 33, 801–816 (2023).
Moss, J., Fishman, P. H., Manganiello, V. C., Vaughan, M. & Brady, R. O. Functional incorporation of ganglioside into intact cells—induction of choleragen responsiveness. Proc. Natl Acad. Sci. USA 73, 1034–1037 (1976).
Gilbert, L. A. et al. Genome-scale CRISPR-mediated control of gene repression and activation. Cell 159, 647–661 (2014).
Tringali, C. et al. Modification of sialidase levels and sialoglycoconjugate pattern during erythroid and erytroleukemic cell differentiation. Glycoconj. J. 24, 67–79 (2007).
Arumugam, S. et al. Ceramide structure dictates glycosphingolipid nanodomain assembly and function. Nat. Commun. 12, 3675 (2021).
Chinnapen, D. J. et al. Lipid sorting by ceramide structure from plasma membrane to ER for the cholera toxin receptor ganglioside GM1. Dev. Cell 23, 573–586 (2012).
Kenworthy, A. K. et al. Cholera toxin as a probe for membrane biology. Toxins 13, 543 (2021).
Madunic, K. et al. Specific (sialyl-)Lewis core 2 O-glycans differentiate colorectal cancer from healthy colon epithelium. Theranostics 12, 4498–4512 (2022).
Robbe, C., Capon, C., Coddeville, B. & Michalski, J. C. Structural diversity and specific distribution of O-glycans in normal human mucins along the intestinal tract. Biochem. J. 384, 307–316 (2004).
Goto, Y. et al. Innate lymphoid cells regulate intestinal epithelial cell glycosylation. Science 345, 1254009 (2014).
Carroll, D. J. et al. Interleukin-22 regulates B3GNT7 expression to induce fucosylation of glycoproteins in intestinal epithelial cells. J. Biol. Chem. 298, 101463 (2022).
Pham, T. A. et al. Epithelial IL-22RA1-mediated fucosylation promotes intestinal colonization resistance to an opportunistic pathogen. Cell Host Microbe 16, 504–516 (2014).
Pickard, J. M. et al. Rapid fucosylation of intestinal epithelium sustains host–commensal symbiosis in sickness. Nature 514, 638–641 (2014).
Pickard, J. M. & Chervonsky, A. V. Intestinal fucose as a mediator of host–microbe symbiosis. J. Immunol. 194, 5588–5593 (2015).
Meng, D. et al. Bacterial symbionts induce a FUT2-dependent fucosylated niche on colonic epithelium via ERK and JNK signaling. Am. J. Physiol. Gastrointest. Liver Physiol. 293, G780–G787 (2007).
Muraoka, W. T. & Zhang, Q. Phenotypic and genotypic evidence for l-fucose utilization by Campylobacter jejuni. J. Bacteriol. 193, 1065–1075 (2011).
Stahl, M. et al. l-Fucose utilization provides Campylobacter jejuni with a competitive advantage. Proc. Natl Acad. Sci. USA 108, 7194–7199 (2011).
Staib, L. & Fuchs, T. M. Regulation of fucose and 1,2-propanediol utilization by Salmonella enterica serovar Typhimurium. Front. Microbiol 6, 1116 (2015).
Ruiz-Palacios, G. M., Cervantes, L. E., Ramos, P., Chavez-Munguia, B. & Newburg, D. S. Campylobacter jejuni binds intestinal H(O) antigen (Fucα1, 2Galβ1, 4GlcNAc), and fucosyloligosaccharides of human milk inhibit its binding and infection. J. Biol. Chem. 278, 14112–14120 (2003).
Chessa, D., Winter, M. G., Jakomin, M. & Bäumler, A. J. Salmonella enterica serotype Typhimurium Std fimbriae bind terminal α(1,2)fucose residues in the cecal mucosa. Mol. Microbiol. 71, 864–875 (2009).
Barra, J. L., Monferran, C. G., Balanzino, L. E. & Cumar, F. A. Escherichia coli heat-labile enterotoxin preferentially interacts with blood group A-active glycolipids from pig intestinal mucosa and A- and B-active glycolipids from human red cells compared to H-active glycolipids. Mol. Cell. Biochem. 115, 63–70 (1992).
Balanzino, L. E., Barra, J. L., Galván, E. M., Roth, G. A. & Monferran, C. G. Interaction of cholera toxin and Escherichia coli heat-labile enterotoxin with glycoconjugates from rabbit intestinal brush border membranes: relationship with ABH blood group determinants. Mol. Cell. Biochem. 194, 53–62 (1999).
Galván, E. M., Roth, G. A. & Monferran, C. G. Participation of ABH glycoconjugates in the secretory response to Escherichia coli heat-labile toxin in rabbit intestine. J. Infect. Dis. 180, 419–425 (1999).
Galván, E. M., Diema, C. D., Roth, G. A. & Monferran, C. G. Ability of blood group A-active glycosphingolipids to act as Escherichia coli heat-labile enterotoxin receptors in HT-29 cells. J. Infect. Dis. 189, 1556–1564 (2004).
Galván, E. M., Roth, G. A. & Monferran, C. G. Functional interaction of Escherichia coli heat-labile enterotoxin with blood group A-active glycoconjugates from differentiated HT29 cells. FEBS J. 273, 3444–3453 (2006).
Holmner, A., Askarieh, G., Okvist, M. & Krengel, U. Blood group antigen recognition by Escherichia coli heat-labile enterotoxin. J. Mol. Biol. 371, 754–764 (2007).
Marionneau, S. et al. Norwalk virus binds to histo-blood group antigens present on gastroduodenal epithelial cells of secretor individuals. Gastroenterology 122, 1967–1977 (2002).
Huang, P. et al. Noroviruses bind to human ABO, Lewis, and secretor histo-blood group antigens: identification of 4 distinct strain-specific patterns. J. Infect. Dis. 188, 19–31 (2003).
Lindesmith, L. et al. Human susceptibility and resistance to Norwalk virus infection. Nat. Med. 9, 548–553 (2003).
Harrington, P. R., Vinjé, J., Moe, C. L. & Baric, R. S. Norovirus capture with histo-blood group antigens reveals novel virus–ligand interactions. J. Virol. 78, 3035–3045 (2004).
Huang, P. et al. Norovirus and histo-blood group antigens: demonstration of a wide spectrum of strain specificities and classification of two major binding groups among multiple binding patterns. J. Virol. 79, 6714–6722 (2005).
Borén, T., Falk, P., Roth, K. A., Larson, G. & Normark, S. Attachment of Helicobacter pylori to human gastric epithelium mediated by blood group antigens. Science 262, 1892–1895 (1993).
Ilver, D. et al. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science 279, 373–377 (1998).
Ikehara, Y. et al. Polymorphisms of two fucosyltransferase genes (Lewis and Secretor genes) involving type I Lewis antigens are associated with the presence of anti-Helicobacter pylori IgG antibody. Cancer Epidemiol. Biomark. Prev. 10, 971–977 (2001).
Aspholm-Hurtig, M. et al. Functional adaptation of BabA, the H. pylori ABO blood group antigen binding adhesin. Science 305, 519–522 (2004).
Hage, N. et al. Structural basis of Lewisb antigen binding by the Helicobacter pylori adhesin BabA. Sci. Adv. 1, e1500315 (2015).
Chakraberty, R., Reiz, B. & Cairo, C. W. Profiling of glycosphingolipids with SCDase digestion and HPLC-FLD–MS. Anal. Biochem 631, 114361 (2021).
Doench, J. G. et al. Optimized sgRNA design to maximize activity and minimize off-target effects of CRISPR–Cas9. Nat. Biotechnol. 34, 184–191 (2016).
Li, W. et al. MAGeCK enables robust identification of essential genes from genome-scale CRISPR/Cas9 knockout screens. Genome Biol. 15, 554 (2014).
Liao, Y., Wang, J., Jaehnig, E. J., Shi, Z. & Zhang, B. WebGestalt 2019: gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 47, W199–W205 (2019).
Turner, S. D. qqman: an R package for visualizing GWAS results using Q–Q and Manhattan plots. J. Open Source Softw. 3, 731 (2018).
Varadi, M. et al. AlphaFold Protein Structure Database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 50, D439–D444 (2022).
Kadirvelraj, R. et al. Comparison of human poly-N-acetyl-lactosamine synthase structure with GT-A fold glycosyltransferases supports a modular assembly of catalytic subsites. J. Biol. Chem. 296, 100110 (2021).
Miller, J. J. et al. α-Galactosidase A-deficient rats accumulate glycosphingolipids and develop cardiorenal phenotypes of Fabry disease. FASEB J. 33, 418–429 (2019).
Barone, A., Benktander, J., Teneberg, S. & Breimer, M. E. Characterization of acid and non-acid glycosphingolipids of porcine heart valve cusps as potential immune targets in biological heart valve grafts. Xenotransplantation 21, 510–522 (2014).
Karlsson, H., Halim, A. & Teneberg, S. Differentiation of glycosphingolipid-derived glycan structural isomers by liquid chromatography/mass spectrometry. Glycobiology 20, 1103–1116 (2010).
Shajahan, A., Heiss, C., Ishihara, M. & Azadi, P. Glycomic and glycoproteomic analysis of glycoproteins—a tutorial. Anal. Bioanal. Chem. 409, 4483–4505 (2017).
Shajahan, A. et al. Comprehensive characterization of N- and O-glycosylation of SARS-CoV-2 human receptor angiotensin converting enzyme 2. Glycobiology 31, 410–424 (2021).
Shajahan, A., Supekar, N. T., Gleinich, A. S. & Azadi, P. Deducing the N- and O-glycosylation profile of the spike protein of novel coronavirus SARS-CoV-2. Glycobiology 30, 981–988 (2020).
Chai, W. et al. Negative-ion electrospray mass spectrometry of neutral underivatized oligosaccharides. Anal. Chem. 73, 651–657 (2001).
Acknowledgements
We thank A. Singla, D. Andrade Silva and M. Burns for comments on the manuscript. We thank E. Capota and D. Andrade Silva for technical support. We thank A. Wands, D. Carroll, H. Wu, H. Khan and M. Shiloh for advice and reagents. We thank the UT Southwestern Proteomics Core Facility and its director, A. Lemoff. We acknowledge support from the National Institutes of Health (R01GM090271 and R35GM145599 to J.J.K. and R24GM137782 to P.A.), the Swedish Cancer Foundation (22 2079 Pj to S.T.) and the Welch Foundation (I-1686 to J.J.K.). M.T.G. received support from the NIH (T32GM145467). We thank C. Cairo (University of Alberta) for the RhtrECI plasmid and R. Schnaar (Johns Hopkins) for the P4 inhibitor.
Author information
Authors and Affiliations
Contributions
A.C.G. and J.J.K. conceptualized the project and experimental approach, with input from A.B., R.S.R.K. and U.Y. A.C.G. conducted all cell culture experiments, flow cytometry experiments, functional assays and immunoblot experiments. A.B. prepared biotinylated CTB and CTB mutants with supervision from U.Y. S.A.A.-H. conducted MS analysis of intact glycolipids with supervision from P.A. M.T.G. performed MS analysis of procainamide-labeled glycans from glycolipids with supervision from P.A. D.Z. isolated neutral glycolipids and conducted MS analysis of their glycans with supervision from S.T. N.B.M. and S.A.A.-H. conducted N-linked and O-linked glycomic analyses with supervision from P.A. R.S.R.K. prepared EGCase and conducted preliminary analyses of glycolipids. X.Z. performed bioinformatics analyses with supervision from C.X. A.C.G. and J.J.K. wrote the manuscript with input from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Chemical Biology thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Characterization of B3GALT5-KO and B3GALT5-KO + OE cells.
(A, B) Representative histograms (left panel) from the flow cytometry analyses of cell surface binding of Lewis a antibody (A) or Lewis x antibody (B) to indicated cell lines. Bar graphs (right panel) show quantification from 3 independent trials. Error bars indicate mean ± SD. (C) Quantification of gMFI from flow cytometry analyses of cells treated with increasing concentrations of CTB. Data shown are from 3 independent trials and normalized to the maximum APC signal in WT cells. Error bars indicate mean ± SD. Indicated cell lines were incubated for 72 h with increasing concentrations of CTB-Saporin (D) or unconjugated saporin (E). Cell survival upon internalization of CTB-saporin was measured using the Cell Titer-Glo 2.0 assay. Data shown are luminescence values normalized to the signal from the untreated condition for each cell type. Each datapoint is a biological replicate consisting of 3 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates. (F) Cells pretreated with BFA or vehicle control for 0.5 h were incubated for 1.5 h with CT (1 nM) or buffer alone. Accumulation of cAMP was measured. Data shown are inverse of luminescence values normalized first to the total amount of cells plated for each cell line, then to the signal in CT-treated control cells. Each datapoint is a biological replicate consisting of 3 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates. (G) Control, B3GALT5-KO m1, m2 and KO + OE cells were treated with forskolin (10 µM) for 0.5 h and then analyzed as in panel F. Statistical analyses for panels A and B were performed by one-way ANOVA with Tukey correction and for panels C, F, and G by two-way ANOVA with Tukey correction.‘ns’ indicates not significant, **** indicates adjusted P-value < 0.0001. Exact P-values are as follows: 0.0004 and 0.0412 for control versus B3GALT5-KO m2 and B3GALT5-KO + OE; 0.0041 and 0.0024 for B3GALT5-KO m1 versus B3GALT5-KO m2 and B3GALT5-KO + OE, respectively (panel B); 0.0390 for control versus B3GALT5-KO m2 treated with 0.0375 μg/mL; 0.0002 and 0.0014 for control versus B3GALT5-KO m2 or m1 treated with 0.075 μg/mL, respectively (panel C).
Extended Data Fig. 2 Characterization of B3GNT5-KO and B3GNT5-KO + OE cells.
(A) Quantification of gMFI from flow cytometry analyses of cells treated with increasing concentrations of CTB. Data shown are from 3 independent trials and normalized to the maximum APC signal in WT cells. Error bars indicate mean ± SD. (B, C) Representative histograms (left panel) from the flow cytometry analyses of cell surface binding of Lewis x antibody (B) or Lewis a antibody (C) to control, B3GNT5-KO m1, m2 and KO + OE cells. Bar graphs (right panel) show quantification from 3 independent trials. Control, B3GNT5-KO m1 and KO + OE cells were incubated for 72 h with increasing concentrations of CTB-Saporin (D) or unconjugated saporin (E). Cell survival upon internalization of CTB-saporin was measured using the Cell Titer-Glo 2.0 assay. Data shown are luminescence values normalized to the signal from the untreated condition for each cell type. Each datapoint is a biological replicate consisting of 3 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates. (F) Cells pretreated with BFA or a vehicle control for 0.5 h were incubated for 1.5 h with CT (1 nM) or buffer alone. Accumulation of cAMP was measured. Data shown are inverse of luminescence values normalized first to the total amount of cells plated for each cell line, then to the signal in CT-treated control cells. Each datapoint is a biological replicate consisting of 3 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates. (G) Control, B3GNT5-KO m1, m2 and KO + OE cells were treated with forskolin (10 µM) for 0.5 h and then analyzed as in panel F. Statistical analyses for panels B and C were performed by one-way ANOVA with Tukey correction and for panels A, D, E, F, and G by two-way ANOVA with Tukey correction. ‘ns’ indicates not significant, **** indicates adjusted P-value < 0.0001. Exact P-values are as follows: 0.0219 for control versus B3GNT5-KO m1 treated with 1.25 μg/mL; 0.0010 and 0.0077 for control versus B3GNT5-KO m1 or m2 treated with 2.5 μg/mL, respectively (panel A); 0.0059 and 0.0036 for control versus B3GNT5-KO + OE treated with 2.5 or 12.5 μg/mL CTB-saporin (panel D).
Extended Data Fig. 3 Characterization of B3GALT5 + B3GNT5-dKO cells.
(A) Representative histograms (left panel) from the flow cytometry analyses of cell surface binding of Lewis x antibody to control, B3GALT5 -KO m1, B3GNT5-KO m1, and B3GALT5 + B3GNT5-dKO cells. Bar graph (right panel) shows quantification from 3 independent trials. (B, C) Control, B3GALT5-KO m1, and B3GALT5 + B3GNT5-dKO cells were incubated for 72 h with increasing concentrations of CTB-Saporin (B) or unconjugated saporin (C). Cell survival upon internalization of CTB-saporin measured using the Cell Titer-Glo 2.0 assay. Data shown are luminescence values normalized to the signal from the untreated condition for each cell type. Each datapoint is a biological replicate consisting of 3 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates. (D) Cells pretreated with BFA or a vehicle control for 0.5 h were incubated for 1.5 h with CT (1 nM) or buffer alone. Accumulation of cAMP was measured. Data shown are inverse of luminescence values normalized first to the total amount of cells plated for each cell line, then to the signal in CT-treated control cells. Each datapoint is a biological replicate (n = 2) consisting of 3 averaged technical replicates. (E) Control, B3GALT5-KO m1, m2 and KO + OE cells were treated with forskolin (10 µM) for 0.5 h and then analyzed as in panel D. Each datapoint is a biological replicate consisting of 3 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates. Statistical analyses for panel A were performed by one-way ANOVA with Tukey correction and for panels B, C, D, and E by two-way ANOVA with Tukey correction. **** indicates adjusted P-value < 0.0001. Exact P-values are as follows: 0.0003 and 0.0048 for B3GALT5 + B3GNT5-dKO versus control or B3GALT5-KO m1, respectively (panel A); 0.0002 and 0.0171 for control versus B3GALT5 + B3GNT5-dKO treated with 2.5 μg/mL CTB saporin or 12.5 μg/mL saporin (panels C and D, respectively).
Extended Data Fig. 4 Fucosylated lacto-series GSLs detected in control but not KO cell lines.
MS analysis of GSLs from control (A), B3GALT5-KO m1 (B), and B3GNT5-KO m1 (C) cells. (D) Example MS/MS spectrum of a fucosylated lacto-series glycolipid detected in control cells, confirming structure.
Extended Data Fig. 5 Validation of GSLs as decoy receptors for CT.
(A) Lectin blot with CTB-biotin of control and B3GALT5-KO m1 cell lysates, and pure GM1. Samples were treated for 16 h with endoglycoceramidase or a vehicle control. Data shown are a single representative trial of 3 independent biological replicates. (B-G) Control, B3GNT5-KO + OE, and B3GALT5-KO m1 cells were treated with P4 inhibitor of glycosphingolipid biosynthesis for 72 h then lysed for lectin blot analysis (B) or treated with BFA or a vehicle control for 0.5 h prior to incubation with CT (1 nM) for analysis of cAMP accumulation (C, D, and E). Alternately, cells were treated with forskolin (10 µM) for 0.5 h (F and G). Accumulation of cAMP was measured. Data shown are inverse of luminescence values normalized first to the total amount of cells plated for each cell line, then to the signal in CT-treated control cells. Each datapoint is a biological replicate (n = 2 in panel E, n = 3 in panels C, D, F, and G) consisting of 3 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates. Statistical analyses were performed by two-way ANOVA with Tukey correction. Exact P-values are 0.0002 for control versus inhibitor-treated B3GALT5-KO m1 as well as inhibitor-treated control versus untreated B3GALT5-KO m1; for inhibitor-treated control versus inhibitor-treated B3GALT5-KO m1, the exact P-value is 0.0052 (panel C).
Extended Data Fig. 6 Analysis of GSLs from B3GNT5-KO + OE cells.
(A) Glycolipids were isolated from B3GALT5-KO m1 or B3GNT5-KO + OE cells. Glycans were released with endoglycoceramidase, labeled with procainamide, and analyzed by mass spectrometry with HILIC-FL separation. (B, C) LC-ESI/MS analysis of oligosaccharides obtained by digestion of neutral GSLs from B3GNT5-KO + OE cells with endoglycoceramidase I. Diagnostic ions indicate carbohydrate linkage positions. (C) An MS spectrum displaying a series of C type fragment ions (C2α at m/z 528, C3α at m/z 690, C4α at m/z 1039, C5α at m/z 1201, C6 at m/z 1550, and C7 at m/z 1712), which identified an oligosaccharide with Hex-(Fuc-)HexNAc-Hex-(Fuc-)HexNAc-Hex-(Fuc-)HexNAc-Hex-Hex sequence. The ion at m/z 364 is obtained by double glycosidic cleavage of the 3-linked branch (C2/Z3β), and characteristic for an internal 4-linked GlcNAc substituted with a Fuc at 3-position that is a terminal Lex (refs. 90,94). Taken together this indicated an undecasaccharide with a terminal Lex determinant. (D, E) B3GALT5 + B3GNT5-dKO cells were incubated with GSLs extracted from B3GNT5-KO + OE cells, a commercial mixture of neutral GSLs, or purified GM1. Cells were then treated with BFA or a vehicle control for 0.5 h prior to incubation with CT (1 nM) for analysis of cAMP accumulation (D). Alternately, cells were treated with forskolin (10 µM) for 0.5 h (n = 2) (E). Accumulation of cAMP was measured. Data shown are inverse of luminescence values normalized first to the total amount of cells plated for each cell line, then to the signal in CT-treated control cells. Each datapoint is a biological replicate (n = 3 for panel D, n = 2 for panel E) consisting of 2 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates (panel D). Statistical analyses were performed by two-way ANOVA with Tukey correction. No significant differences were observed.
Extended Data Fig. 7 B3GALT5-KO cells exhibit increased fucosylation on N-linked glycoproteins.
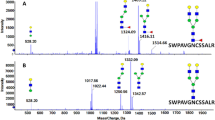
(A) N-linked glycoforms detected in control and B3GALT5-KO m1 cells by LC-MS/MS analysis. Quantification is based on triplicate analysis. (B) Example MS/MS of a complex, fucosylated N-linked glycan detected in B3GALT5-KO m1 cells confirming placement of fucose. (C) Bar graphs showing the relative enrichment of mono- vs di- vs tri-fucosylated N-linked glycans detected by LC-MS/MS analysis of control and B3GALT5-KO m1 cells. Statistical analysis was performed by two-tailed t-test with Holm-Šídák correction. Exact adjusted P-value = 0.013508 for mono-fucosylated N-linked glycans detected in control versus B3GALT5-KO m1. Exact adjusted P-value = 0.014105 for both di- and tri-fucosylated N-linked glycans detected in B3GALT5-KO m1 versus control.
Extended Data Fig. 8 B3GALT5-KO cells exhibit increased fucosylation on O-linked glycoproteins.
(A) O-linked glycoforms detected in control and B3GALT5-KO m1 cells by LC-MS/MS analysis. Quantification is based on triplicate analysis. (B) Example MS/MS of a fucosylated O-linked glycan detected in B3GALT5-KO m1 cells. (C) Example MS/MS of core 2 O-linked glycan detected in B3GALT5-KO m1 cells confirming structure. (D, E) B3GALT5-KO m1 cells were rescued with either WT or catalytically dead (mut) B3GALT5-OE. Rescued cells were treated with BFA or a vehicle control for 0.5 h prior to incubation with CT (1 nM) for analysis of cAMP accumulation (D). Alternately, cells were treated with forskolin (10 µM) for 0.5 h (E). Accumulation of cAMP was measured. Data shown are inverse of luminescence values normalized first to the total amount of cells plated for each cell line, then to the signal in CT-treated control cells. Each datapoint is a biological replicate consisting of 2 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates. Statistical analyses were performed by two-way ANOVA with Tukey correction. No significant differences were observed.
Extended Data Fig. 9 Validation of SLC35C1-KO cell lines.
(A) GM1 was treated with EGCase or vehicle, then detected by lectin blot using WT CTB-biotin, W88K CTB-biotin, and H18L CTB-biotin. Data presented are from 3 distinct membranes processed simultaneously (separated by vertical lines) and are a single representative trial from 3 biological replicates. (B, C) Representative histograms (left panel) from flow cytometry analyses of anti-Lex antibody (B) and AAL (C) binding to surfaces of control, B3GALT5-KO m1, SLC35C1-KO, and B3GALT5 + SLC35C1-dKO cells. Quantification of gMFIs (right panel) from 3 biological replicates are normalized to the maximum signal in control cells. Lysates from control, B3GALT5-KO m1, SLC35C1-KO and B3GALT5 + SLC35C1-dKO cells were analyzed by immunoblot probing with anti-Lex antibody or lectin blot probing with AAL (D). Data shown are a single representative trial from 3 biological replicates. (E) Control, B3GALT5-KO m1, SLC35C1-KO and B3GALT5 + SLC35C1-dKO cells were incubated for 72 h with unconjugated saporin (12.5 μg/mL). Survival data shown are luminescence values normalized to the signal from the untreated condition for each cell type. Each datapoint indicates the mean of 2 biological replicates, each consisting of 3 averaged technical replicates. (F) Cells pretreated with BFA or a vehicle control for 0.5 h were incubated for 1.5 h with CT (1 nM) or buffer alone. Accumulation of cAMP was measured. Data shown are inverse of luminescence values normalized first to the total amount of cells plated for each cell line, then to the signal in CT-treated control cells. Each datapoint is a biological replicate (n = 2) consisting of 3 averaged technical replicates. (G) Cells were treated with forskolin (10 µM) for 0.5 h and then analyzed as in panel F. Each datapoint is a biological replicate consisting of 3 averaged technical replicates. Error bars indicate mean ± SD of 3 biological replicates. Statistical analyses were performed by one-way (panels B and C) or two-way (panels F and G) ANOVA with Tukey correction. **** indicates adjusted P-value < 0.0001. Exact P-value for control versus B3GALT5-KO m1 treated with AAL is 0.0018.
Extended Data Fig. 10 CTB-interacting glycoproteins.
(A) Top 50 proteins identified from CTB-biotin pulldown from B3GALT5 + B3GNT5-dKO versus B3GALT5 + SLC35C1-dKO cell lysates. Fold-change is the abundance in the B3GALT5 + B3GNT5-dKO pulldown as compared to the B3GALT5 + SLC35C1-dKO pulldown. Unadjusted P-values are derived from two-tailed student’s t-test. (B) Gene ontology (GO) analysis using Fisher’s exact test with Bonferroni correction to identify pathways enriched in CTB-biotin pulldown from B3GALT5 + B3GNT5-dKO cell lysates as compared to B3GALT5 + SLC35C1-dKO cell lysates. Top 25 pathways are shown. (C) Two hits (FLOT1 and MUC1, boldface font in panel A) were selected for confirmation. Total lysates and material from CTB-biotin pulldowns were analyzed by immunoblot using antibodies against FLOT1 and MUC1.
Supplementary information
Supplementary Information
Supplementary Fig. 1.
Supplementary Table 1
Reagents list.
Supplementary Table 2
Oligonucleotide list.
Supplementary Table 3
CRISPR KO sequence validation.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 9
Statistical source data.
Source Data Extended Data Fig. 9
Uncropped western blot.
Source Data Extended Data Fig. 10
Uncropped western blot.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Ghorashi, A.C., Boucher, A., Archer-Hartmann, S.A. et al. Fucosylation of glycoproteins and glycolipids: opposing roles in cholera intoxication. Nat Chem Biol 21, 555–566 (2025). https://doi.org/10.1038/s41589-024-01748-5
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41589-024-01748-5
