
Abstract
The function of most long noncoding RNAs (lncRNAs) is unknown. However, recent studies reveal important roles of lncRNAs in regulating cancer-related pathways. Human antisense lncRNA-NKX2-1-AS1 partially overlaps the NKX2-1/TTF1 gene within chromosomal region 14q13.3. Amplification of this region and/or differential expression of genes therein are associated with cancer progression. Herein we show higher levels of NKX2-AS1 and NKX2-1 in lung adenocarcinomas relative to non-tumor controls but no correlation between NKX2-1-AS1 and NKX2-1 levels across specimens, or with amplification of the 14q13.3 region, suggesting that NKX2-1-AS1 and NKX2-1 are independently regulated. Loss-and-gain of function experiments showed that NKX2-1-AS1 does not regulate NKX2-1 expression, or nearby genes, but controls genes in trans. Genes up-regulated by NKX2-1-AS1-knockdown belong to cell adhesion and PD-L1/PD-1 checkpoint pathways. NKX2-1-AS1 negatively regulates endogenous CD274/PD-L1, a known target of NKX2-1, and the transcriptional activity of -1kb-CD274 promoter-reporter construct. Furthermore, NKX2-1-AS1 interferes with NKX2-1 protein binding to the CD274-promoter, likely by NKX2-1 protein-NKX2-1-AS1 interactions. Finally, NKX2-1-AS1 negatively regulates cell migration and wound healing, but not proliferation or apoptosis. These findings support potential roles of NKX2-1-AS1 in limiting motility and immune system evasion of lung carcinoma cells, highlighting a novel mechanism that may influence tumorigenic capabilities of lung epithelial cells.
Similar content being viewed by others

Introduction
Genomic regions that are frequently amplified in human lung cancer often contain genes that control tumorigenesis. However, the contribution of each gene within these amplified genomic regions to the tumorigenesis process is often not fully defined1. For example, the 14q13.3 cytoband is a genomic region amplified in ~15% of lung cancers2,3,4 and mutations or deletions in this chromosomal region are frequently found in lung and thyroid cancer patients5,6. Within this region, the transcription factors NKX2-1 (thyroid transcription factor-1; also known as TTF1), NKX2-8 and PAX9 are candidate genes that cooperate to control lung cancer cell growth, although other genes in this region are also likely to facilitate the tumorigenesis process4,7. Recently, by whole transcriptome sequencing, a few long non-coding RNAs have been identified that map to this region, including lnc-NKX2-8 and lnc-SLC25A218,9, NANCI10 and NKX2-1-AS111; however, their contribution to lung tumorigenesis is not fully understood.
lncRNAs are transcripts of more than 200 nucleotides in length, but in contrast to protein-coding genes, they lack the structural features needed to be translated into proteins12. The functions of an increasing number of individual lncRNAs have been determined, and the dysregulation of lncRNAs has been linked to a number of diseases. However, for the majority of the annotated lncRNAs, the functional relevance is unknown13. The lack of knowledge about conserved sequences or structures among mammalian lncRNAs has made it difficult to infer the potential functions of these genes as has been done for many protein-coding genes. Efforts are in progress, though, to identify these domains in fungi14 and ultimately in vertebrates15. Therefore, studies about the functions of individual lncRNAs are warranted to determine their role as part of regulatory networks16. This is especially relevant for novel lncRNAs associated with human diseases. Recent studies have revealed the role of an increasing number of lncRNAs in regulating key cancer pathways at a transcriptional, post-transcriptional and epigenetic level17,18,19,20. Moreover, lncRNAs have been shown to be differentially expressed across various stages of tumor differentiation19,21. Evidence indicates that the lncRNA NKX2-1-AS1 is specifically detected in carcinoma cell lines of lung origin22 and, like the adjacent protein-coding gene NKX2-1, is highly expressed in primary lung adenocarcinomas compared to squamous carcinomas11,21,23 and in some small cell carcinomas24, but its contribution to lung tumorigenesis is not well understood.
To evaluate the potential regulatory and biological functions of the NKX2-1-AS1 transcript, we performed expression analyses, loss and gain of function experiments, and functional tests. We have uncovered in this study a role for the NKX2-1-AS1 transcript in regulating genes that control cell adhesion and the migration of lung tumor cell lines. We also identified a role for NKX2-1-AS1 in the regulation of CD274, the gene encoding the Programmed Death-Ligand 1 (PD-L1). These effects are independent of the transcription or translation of the adjacent gene encoding the transcription factor NKX2-1. These findings highlight a new mechanism by which NKX2-1-AS1 might regulate tumorigenic properties of lung cells.
Results
NKX2-1-AS1 and NKX2-1 are co-expressed at variable levels in lung carcinoma cells, lung cell lines and normal tissues
Previous RNA-sequencing analyses of non-small cell lung carcinomas (NSCLCs) have shown that NKX2-1-AS1 is highly expressed in primary lung adenocarcinomas compared to squamous carcinomas11,21,23 and in some small cell carcinomas24, a pattern of expression that is similar to that of the adjacent protein-coding gene NKX2-1. To validate whether the NKX2-1-AS1 transcript annotated in public databases (ENST00000521292.2, hg38 chr14:36,519,278–36,523,016, Fig. 1A) is detected in lung tumors, we measured the expression of NKX2-1-AS1 and NKX2-1 using qPCR in a select number of lung adenocarcinomas (AC, n = 8) and squamous cell carcinomas (SCC, n = 8) and their matched non-tumor controls, described in Table 1. This analysis showed that, compared to non-tumor matching controls, NKX2-1-AS1 and NKX2-1 expression was increased in adenocarcinoma, whereas levels of both NKX2-1-AS1 and NKX2-1 were decreased in squamous cell carcinoma [AC vs SCC: NKX2-1-AS1 p = 0.030; NKX2-1 p = 0.029] (Fig. 1B), in concordance with previous RNA-sequencing findings that NKX2-1-AS1 expression is higher in lung adenocarcinomas than in squamous cell carcinomas11,21. However, we noticed that the relative expression of NKX2-1 and NKX2-1-AS1 differed considerably across tumors. Furthermore, there was no significant correlation between the expression of these two genes across the specimens evaluated in this study (Fig. 1C), or across the tumors in a publicly available NSCLC expression GEO dataset (GDS3627)25 (Supplementary Fig. S1). Variable NKX2-1-AS1 expression was observed in the presence of relatively constant NKX2-1 expression, and vice versa. These results suggest that in certain conditions co-transcription of these genes is either biased towards one direction and/or that transcripts for these genes are degraded at different rates. Alternatively, these findings might suggest that NKX2-1-AS1 and NKX2-1 are independently regulated.

NKX2-1-AS1 expression patterns in human non-small cell lung carcinoma (NSCLC). (A) Schematic representation of the relative chromosomal location of NKX2-1-AS1 lncRNA and adjacent NKX2-1 protein coding-gene in human chromosomal region 14q13.3. Arrows indicate direction of transcription. Boxes indicate exons, dotted lines indicate introns and colored boxes indicate coding regions. (B) Levels of expression of NKX2-1-AS1 and NKX2-1 in NSCLCs (SCC = squamous cell carcinoma; and AC = adenocarcinoma) relative to their corresponding non-tumor control determined by qPCR (n = 8; *p < 0.03). (C) Correlation plot of the expression of NKX2-1-AS1 and NKX2-1 determined by qPCR in the above tumors and their corresponding non-tumor specimens analyzed in this study. (D) Amplification status of the NKX2-1-AS1 locus determined by qPCR of genomic DNA and expressed as copy number of the NKX2-1-AS1 gene per genome plotted relative to NKX2-1-AS1 expression level in each sample (n = 12).
NKX2-1-AS1 is located in the 14q13.3 chromosomal region, which is amplified in ∼15% of lung cancers2,4. To evaluate whether differences in the expression of NKX2-1-AS1 in tumor samples was due to amplification of this locus, we analyzed the copy number of NKX2-1-AS1 by genomic qPCR. We normalized the values to LINE1, a repetitive sequence with relatively conserved copy number in human normal and neoplastic cells26,27. Normal human iPSC DNA was used to generate the calibration curves. We found that all but two samples tested contained one copy of NKX2-1-AS1 per genome (Fig. 1D). Moreover, the two samples with a higher copy number (5 or 11 copies) have lower NKX2-1-AS1 expression than many samples with only one copy. Despite the variable expression levels of NKX2-1-AS1 in the specimens studied, including some with very high levels of NKX2-1-AS1, a link between expression level and copy number was not observed.
To determine whether NKX2-1-AS1 is also expressed in cell lines known to express NKX2-1, we measured the expression of NKX2-1-AS1 and NKX2-1 by RT-PCR (Fig. 2A) in human lung carcinoma cell lines. Some lines, such as H441 and H661, express both NKX2-1 and NKX2-1-AS1, but others, including H1299, BEAS-2B, A549 and Calu-6, do not express detectable levels of either transcript. We also quantified the relative level of expression of NKX2-1-AS1 and NKX2-1 in H441 and H661 cells relative to normal lung and thyroid (Fig. 2B). H441 cells express the highest levels of both genes among the cell lines tested; therefore, most studies were performed in this cell line, with confirmatory studies performed in H661 cells. Both genes are expressed at higher levels in the thyroid than in the lung (Fig. 2B).

NKX2-1-AS1 follows tissue-specific patterns of expression similar to NKX2-1 in human cells. (A) Expression of NKX2-1-AS1 and NKX2-1 determined by RT-PCR in lung cell lines. The NKX2-1-AS1 PCR fragments were sequenced to confirm the identity of the sequence. (B) Relative expression patterns of NKX2-1-AS1 and NKX2-1 in tissues and cell lines, including normal human adult lung and thyroid and H441 and H661 cell lines, as determined by qPCR (n = 3; *p < 0.05; **p < 0.01). (C) 5′-RACE analysis of NKX2-1-AS1 in human thyroid identified multiple transcription initiation sites within the 500 bp 5′ of the transcription initiation site reported in Ensembl. Thyroid total RNA was used as it has higher levels of NKX2-1-AS1 expression than the lung. The sensitivity of the method did not allow us to analyze lung RNA. Thirty clones, generated in three independent experiments, were sequenced. The black bar indicates the region recently identified in a genome wide analysis of accurate lncRNA transcription initiation sites13. (D) Time course analysis of NKX2-1-AS1 and NKX2-1 transcript stability in H441 cells by qPCR after inhibition of transcription by actinomycin D treatment (n = 3; *p < 0.05).
These data indicate that the overall pattern of expression of NKX2-1-AS1 mature transcript is similar to that of NKX2-1 in normal and tumor tissues and cell lines, but that the relative expression levels of these genes vary among specimens, and that increased levels of expression of NKX2-1-AS1 in certain lung tumors is not explained by locus amplification.
NKX2-1-AS1 transcripts can be initiated at multiple transcription start sites
Most lncRNA sequences are derived from RNA-sequencing assemblies, and in general, their 5′ ends are not accurate13. By 5′-RACE analyses of the NKX2-1-AS1 transcripts in normal human thyroid, which has higher level of expression of NKX2-1-AS1 compared to lung, we identified multiple transcription initiation sites for the NKX2-1-AS1 transcript in a 500 bp region surrounding the TSS that has been reported in Ensembl (ENST00000521292.2, hg38 chr14:36,519,278) (Fig. 2C). These results are supported by a recent in silico study of the potential TSS of lncRNAs13, which found that most of the TSS for NKX2-1-AS1 are located around 100 bp upstream of the TSS reported in Ensembl. These findings suggest the presence of longer NKX2-1-AS1 transcripts that have not been identified in previous RNA-sequencing analyses.
NKX2-1-AS1 transcript is more stable than NKX2-1 transcript
Since divergent transcription is a common phenomenon that can result in shorter transcripts that are rapidly degraded, or longer transcripts that might perform unique biological functions28,29, we measured the relative stability of the non-coding NKX2-1-AS1 transcript compared to that of the protein-coding NKX2-1 transcript. For these analyses, we inhibited transcription using actinomycin D and collected cells for RNA analysis by qPCR at different time points from 0 to 8 h. These analyses showed that despite the relatively low expression of NKX2-1-AS1, the transcript is more stable; 50% of the initial transcripts could be detected at more than 4 hours, whereas NKX2-1 levels were reduced by 50% within approximately 2 hours (Fig. 2D).
NKX2-1-AS1 does not regulate expression of NKX2-1 or other genes in the 14q13.3 chromosomal region
LncRNAs can regulate genes within nearby or distant genomic regions. To evaluate whether the NKX2-1-AS1 transcript regulates the expression of adjacent genes within the 14q13.3 genomic region (Fig. 3A), we performed loss of function experiments using a mix of three siRNAs targeting NKX2-1-AS1 and a non-silencing control in H441 and H661 cells. Using qPCR, we showed that reduction of NKX2-1-AS1 expression by ~70% at 48 h or by ~90% at 72 h in H441 cells using the mix of three siRNAs (Fig. 3B) did not affect expression of NKX2-1 at the mRNA (Fig. 3C) or protein (Fig. 3D,E) level. Individual siRNAs were also tested and no significant down-regulation of NKX2-1 was observed in H441 cells (Supplementary Fig. S2). Similarly, no reduction in NKX2-1 mRNA levels was observed in H661 cells after knockdown of NKX2-1-AS1 (Supplementary Fig. S3).

NKX2-1-AS1 does not regulate expression of genes in the 14q13.3 chromosomal region. (A) Scheme of human chr14 within the 14q13.3 cytoband region indicating selected genes neighboring NKX2-1-AS1. (B) qPCR analyses of NKX2-1-AS1 in H441 cells treated with a pool of three siRNAs targeting NKX2-1-AS1 exon 2 show significant down-regulation of NKX2-1-AS1 at 48 h and 72 h post transfection (n = 6; **p = 0.01 and ***p = 0.005 respectively). (C) qPCR analyses of the neighboring protein coding gene NKX2-1 in NKX2-1-AS1 knocked-down cells show no changes in NKX2-1 RNA levels (n = 6). (D) Representative western blots of NKX2-1 protein in non-silencing control [c] and NKX2-1-AS1 siRNA [si] treated H441 cells normalized to β-actin. (E) Densitometry of the western blot signals normalized to β-actin indicates that NKX2-1 protein levels also remained unchanged; n = 3. (F) Expression levels of NKX2-1-AS1 in the knockdown cells at 48 h and of other genes in the 14q13.3 chromosomal region as determined by microarray analysis; n = 6. (G) qPCR analysis in the NKX2-1-AS1 knockdown cells at 24, 48 and 72 h of MBIP; (H) NKX2-8; and (I) PAX9. n = 6, *p = 0.002.
We also measured gene expression following NKX2-1-AS1 knockdown in H441 cells using microarrays, which showed that the expression of most genes in the 14q13.3 cytoband (Fig. 3A), including MBIP, SFTA3 (NANCI), NKX2-8, and SLC25A21, was unchanged at 48 h, with the exception of a small but significant effect on PAX9 (p = 0.02, FDR q = 0.17) (Fig. 3F). We validated these findings by qPCR at 24, 48 and 72 h after knockdown of NKX2-1-AS1 (Fig. 3G–I). Down-regulation of PAX9 was observed, but only at 72 h; expression of the other genes in the cytoband was not changed. Overall, these data support a limited role of the NKX2-1-AS1 transcript in regulating genes in cis.
NKX2-1-AS1 does not control alternative splicing of NKX2-1 RNA or translation of NKX2-1 protein
Analyses of the role of divergent lncRNAs with 5′ regions that overlap a protein-coding mRNA have demonstrated that some of them can control alternative splicing and/or protein translation30. As shown in Fig. 3D,E, no changes in NKX2-1 protein levels were observed in cells treated with NKX2-1-AS1 siRNAs for 48 h, despite a 70% reduction in NKX2-1-AS1 levels. Furthermore, no changes in the ratio of the two major NKX2-1 variants (variant 1 and variant 2) were detected by RT-qPCR (data not shown). These data suggest that the partial overlapping of the 5′ ends of NKX2-1-AS1 and NKX2-1 does not control the selection of different transcription initiation sites, splicing of NKX2-1 RNA, or translation of NKX2-1 protein, as has been shown for other divergent overlapping mRNA-lncRNA pairs30.
Overexpression of NKX2-1 does not change the expression levels of NKX2-1-AS1
To evaluate whether NKX2-1 had any effect on NKX2-1-AS1 expression, we performed NKX2-1 gain of function experiments in H441 cells. We found that overexpression of NKX2-1 by more than 300-fold did not result in alterations in the expression of endogenous NKX2-1-AS1, but significantly increased expression of the positive control target gene SFTPC by 2.7-fold. These data suggest that NKX2-1 and NKX2-1-AS1 do not have a direct reciprocal transcriptional regulatory relationship (Supplementary Fig. S4).
NKX2-1-AS1 negatively regulates PD-1/PD-L1 signaling and adherens junction pathways
The microarray experiment showed considerable changes in gene expression after 48 h of NKX2-1-AS1 knockdown in H441 cells (140 genes at FDR q < 0.01. The top twenty genes that were down-regulated or up-regulated by NKX2-1-AS1 knockdown are listed in Fig. 4A,B. Results for selected genes within these lists were validated by RT-qPCR (Fig. 4C). GSEA was then used to identify biological terms, pathways and processes that were coordinately up- or down-regulated after NKX2-1-AS1 knockdown (Supplementary Materials). Two of the gene sets with the most significant coordinate up-regulation following NKX2-1-AS1 silencing are the Reactome PD-1 signaling and KEGG adherens junction pathways (p < 0.001, FDR q < 0.25). Validation of selected genes in the adherens junction pathway (Supplementary Material) are shown in Fig. 4D.

NKX2-1-AS1 knockdown alters gene expression patterns in H441 cells. (A) List of the top 20 genes down- and (B) up-regulated by NKX2-1-AS1 knockdown determined by microarray analysis, 48 h after treatment (n = 6). (C) qPCR validation of down-regulated and up-regulated genes in NKX2-1-AS1 knockdown cells at 48 h after treatment (n = 3, *p < 0.05, **p < 0.01). (D) Real time PCR validation of adherens junction related genes up-regulated by NKX2-1-AS1 knockdown (n = 3; *p < 0.05; **p < 0.01). (E) Representative western blots of PD-L1 and PTPN1 protein in non-silencing control [c] and NKX2-1-AS1 siRNAs [si] treated H441 cells normalized to β-actin. (F) Densitometry of the western blot signals normalized to β-actin indicates that PD-L1 (n = 9; **p = 0.002) and PTPN1 (n = 3; *p = 0.05) protein levels are increased by NKX2-1-AS1 knockdown.
The PD-1 signaling pathway includes the gene CD274, which encodes PD-L1. The up-regulation of CD274 mRNA and PD-L1 protein following NKX2-1-AS1 knockdown was validated in H441 cells by qPCR (Fig. 4C) and western blot (Fig. 4E,F) respectively and in H661 cells by RT-qPCR (Supplementary Fig. S5A). An inverse trend between the regulation of NKX2-1-AS1 and PD-L1 in tumors relative to non-tumor controls is observed in the specimens tested in this work as well as in a publicly available GEO dataset (GDS3627) (Supplementary Fig. S6). The up-regulation of genes involved in adherens junction formation, including PTPN1 (protein tyrosine phosphatase, non-receptor type 1) that promotes migration and invasion in breast cancer cells31 and in epidermal dendritic cells32, FARP2 (FERM, ARH/RhoGEF and pleckstrin domain protein 2), SNAI2 (snail family transcriptional repressor 2), and MET (MET proto-oncogene, receptor tyrosine kinase), was validated by qPCR in H441 cells (Fig. 4D), and the up-regulation of MET was also validated by qPCR in H661 cells (Supplementary Fig. S5B), suggesting a potential role for NKX2-1-AS1 in the regulation of cell-cell interaction and motility processes. Furthermore, CLDN1, a gene encoding a tight junction component, was also up-regulated by NKX2-1-AS1 knockdown in both cell lines (Fig. 4C and Supplementary Fig. S5A).
NKX2-1-AS1 impairs binding of NKX2-1 protein to the CD274 gene promoter
To identify potential mechanisms of negative regulation of CD274 by NKX2-1-AS1, we used ChIP-qPCR to evaluate the effect of NKX2-1-AS1-knockdown on the binding of NKX2-1, which is known to directly activate the transcription of CD27433. Reducing the expression of NKX2-1-AS1 by siRNA resulted in increased binding of NKX2-1 protein (Fig. 5A) (n = 5, p = 0.005) to the CD274 promoter region. However, the binding of NKX2-1 protein to the proximal promoters of PTPN1 and CLDN1, which were also negatively regulated by NKX2-1-AS1, was unchanged (Fig. 5B,C).

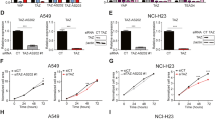
NKX2-1-AS1 overexpression reduces CD274 expression levels in A549 cells in part by impairing NKX2-1 protein binding to the CD274 promoter. ChIP-qPCR analysis of NKX2-1 protein binding to (A) CD274 promoter (n = 4; p = 0.005), (B) CLDN1 promoter (n = 5), and (C) PTPN1 promoter (n = 5) in H441 cells transfected with NKX2-1-AS1 siRNAs or non-silencing control. (D) NKX2-1 co-transfection with the −1kbCD274-Luc vector results in higher luciferase activity (3-fold in the absence of NKX2-1-AS1, 0ug). NKX2-1-AS1 overexpression reduces the activity of the −1kbCD274 promoter in a dose-dependent manner both in the absence (E) (n = 3-4; ANOVA p = 0.003) or presence (F) (n = 3-4; ANOVA p = 0.0001) of NKX2-1 overexpression. NKX2-1-AS1 overexpression reduces the expression of the endogenous CD274 gene in a dose-dependent manner both in the absence (G) (n = 3-4; ANOVA p = 0.001) or presence (H) (n = 3-4; ANOVA p = 0.05) of NKX2-1 overexpression. (I) RIP-qPCR analysis of NKX2-1-AS1 pull down by NKX2-1 antibody compared to IgG control (n = 6; p = 0.05).
NKX2-1-AS1 inhibits transcription from the CD274 proximal promoter and reduces NKX2-1-mediated activation of CD274 transcription
To confirm that NKX2-1 increased CD274 expression by direct activation of the CD274 proximal promoter, we performed co-transfection experiments in A549 cells with plasmids containing NKX2-1 or NKX2-1-AS1 overexpression constructs and a luciferase reporter driven by 1.1 kb of the CD274 promoter. NKX2-1 overexpression increased the activity of the CD274 reporter by 3-fold (Fig. 5D, 0 ug). Conversely, NKX2-1-AS1 overexpression reduced the activity of the CD274 reporter in a dose-dependent manner, both in the absence (ANOVA p = 0.003) and presence (ANOVA p = 0.0001) of NKX2-1 overexpression (Fig. 5E,F, respectively). Furthermore, NKX2-1-AS1 overexpression reduced the expression of endogenous CD274 in a dose-dependent manner, both in the absence (ANOVA p = 0.001) and presence (ANOVA p = 0.05) of NKX2-1 overexpression (Fig. 5G,H, respectively). NKX2-1-AS1 overexpression also reduced the expression of endogenous CLDN1 (ANOVA p = 0.0002) in a dose-dependent manner, but did not affect the expression of PTPN1, RBPJ or FARP2 (Supplementary Fig. S7).
NKX2-1-AS1 interacts with NKX2-1 protein
One of the mechanisms by which lncRNAs inhibit gene expression is by acting as decoys and preventing binding of transcription factors to DNA34. To evaluate whether NKX2-1-AS1 controls the binding of NKX2-1 to the CD274 promoter, we used RNA immunoprecipitation followed by qPCR (RIP-qPCR) to investigate the potential interaction of the NKX2-1-AS1 lncRNA and NKX2-1 protein. In H441 cells, endogenous NKX2-1-AS1 co-immunoprecipitated with NKX2-1 protein (p = 0.05 versus IgG control) (Fig. 5I), suggesting that NKX2-1-AS1-NKX2-1 interactions may inhibit NKX2-1 binding to CD274 promoter.
NKX2-1-AS1 does not change the histone methylation landscape of the CD274 promoter
Another mechanism by which lncRNA mediates the inhibition of gene expression is through the recruitment of chromatin-modifying proteins to alter epigenetic marks on gene regulatory regions34. To evaluate this possibility, we used ChIP in H441 cells to measure H3K4me3 (active chromatin) and H3K27me3 (silenced chromatin) marks and binding of the histone 3 lysine 27 methyltransferase EZH2 to the promoters of CD274, CLDN1, and PTNP1 (which were up-regulated by the knockdown of NKX2-1-AS1) and the NKX2-1 promoter. We found that only H3K4me3 was enriched above IgG background levels at these promoters, and no significant changes in the enrichment of any of these marks was observed in NKX2-1-AS1-knocked down cells (Supplementary Fig. S8). These data suggest that NKX2-1-AS1 inhibition of CD274 is not linked to epigenetic gene regulation by histone methylation mechanisms.
NKX2-1-AS1 inhibits cell migration but has no significant effect on proliferation, invasion or apoptosis
Because of the changes observed in genes involved in cell-cell interaction, we evaluated whether changes in NKX2-1-AS1 expression levels have any effect on the motility of lung tumor cells. Using the mix of three siRNAs described above, we reduced NKX2-1-AS1 expression by 70-90%. Cell monolayer scratch (wound healing) assays performed on these cells showed that decreasing NKX2-1-AS1 expression produced a statistically significant (p < 0.002) increase in wound healing activity of these cells, underscoring a role in cell motility (Fig. 6A,B). Furthermore, migration and invasion analysis using polypropylene or extracellular matrix covered trans-wells showed that reduction of NKX2-1-AS1 significantly increased migration of H441 cells (Fig. 6C) but had no effect on cell invasion (Fig. 6D). Conversely, overexpression of NKX2-1-AS1 cDNA by more than 100-fold in H441 cells by transient transfection did not change the level of expression of NKX2-1 mRNA (Supplementary Fig. S4) but resulted in a moderate but significant reduction in wound healing activity (p < 0.00001) (Fig. 6E,F), supporting a role for NKX2-1-AS1 transcript in the inhibition of lung tumor cell migration by mechanisms independent of regulation of NKX2-1 expression.

NKX2-1-AS1 inhibits cell motility in H441 cells. (A) Representative wound healing analysis of H441 cells treated with NKX2-1-AS1 siRNAs or non-silencing siRNA control. Cells were treated with the siRNAs for 24 h before the scratch was performed (0 h). Three images per scratch were taken at 0, 24, 48 and 72 h in 3 independent experiments. (B) Average wound area closed determined in the above images (*p < 0.002). (C) Migration of NKX2-1-AS1 knockdown H441 cells was compared to non-silencing control in transwell experiments (n = 3; *p < 0.05). (D) Invasion of H441 cells treated with NKX2-1-AS1 siRNA compared to non-silencing control in collagen-covered transwells (n = 3). (E) Representative wound healing analysis of H441 cells transfected with CMV-NKX2-1-AS1 or with empty vector control. After treatment for 24 h the scratch was performed (0 h). Three images per scratch were taken at 0, 24, and 48 h in three independent experiments. (F) Average wound area closed (**p < 0.00001).
To further evaluate potential biological functions of the NKX2-1-AS1 transcript, we tested whether NKX2-1-AS1 controls other classic tumorigenic properties of lung epithelial cells, including cell growth, cell cycle progression and apoptosis. Reduction of NKX2-1-AS1 by 70-90% using the mix of three siRNAs as described above resulted in minimal, and not statistically significant, cell growth reduction at 48 or 72 h (Fig. 7A). Cell cycle analysis of these samples by flow cytometry did not show changes in the number of cells in each stage of the cell cycle (Fig. 7B) or an increase in apoptosis as measured by annexin-V binding (Fig. 7C).

NKX2-1-AS1 knockdown does not affect proliferation or apoptosis of H441 cells. (A) Cell growth was determined by counting cells at 24, 48 and 72 h after treatment with a pool of 3 siRNAs targeting NKX2-1-AS1 (n = 3). (B) Analysis of cell cycle stage by measuring DNA cell content by flow cytometry in NKX2-1-AS1 knockdown cells compared to non-silencing control at 48 h after treatment (n = 3). No significant change in cell number in each cell cycle stage was observed in NKX2-1-AS1 knockdown cells compared to non-silencing control. (C) No significant change in apoptosis was observed in NKX2-1-AS1 knockdown cells compared to non-silencing control as measured by annexin-V binding.
Discussion
Because the 14q13.3 chromosomal region is believed to contribute to the pathobiology of lung tumorigenesis, we sought to elucidate the potential contribution of newly identified genes residing within this region to lung tumorigenesis. Recent findings point to a potential role for NKX2-1-AS1, a long non-coding RNA (lncRNA) mapped to this region, as a regulator of lung tumor cell properties: its selective expression in cell lines of lung and thyroid origin22; its increased expression in lung adenocarcinomas11,21,23; and its close proximity to the transcription factor NKX2-1 (thyroid-transcription factor 1, TTF1, NKX2.1), which is already known to contribute to lung cancer progression35. In this study, we report our novel findings on the potential regulatory and biological functions of the NKX2-1-AS1 transcript in lung cancer cells.
Despite the fact that the 14q13.3 chromosomal region, where NKX2-1-AS1 and NKX2-1 are located, is amplified in ~15% of adenocarcinomas2,4 and is the highest focally amplified region in lung adenocarcinomas1, we found no link between NKX2-1-AS1 expression levels and the copy number of its locus. Although we cannot rule out a role for amplification in the expression of these genes in other lung tumor samples, our data suggest that an active mechanism of gene regulation is in place that controls NKX2-1-AS1 expression in cells where the 14q13.3 region is not amplified. The relatively higher stability of the NKX2-1-AS1 transcript observed in our studies might be one such factor contributing to the increased levels of this gene in lung adenocarcinomas.
Due to their relative position, the NKX2-1 and NKX2-1-AS1 genes are classified as bidirectional and divergent (transcribed from opposite strands in opposite directions)29. Moreover, the 5′ regions of the NKX2-1 and the NKX2-1-AS1 transcripts overlap, sharing > 300 bp of complementary sequence. The lncRNAs that are divergent to protein coding genes are thought to arise as a consequence of the transcription process in which the polymerase II works bidirectionally36. These antisense, noncoding transcripts are in many instances short RNA forms that are rapidly degraded. However, longer lncRNAs (>1-2 kb), which are frequently detected as spliced forms, do not degrade as readily and might have independent functions37,38. Although dissecting these two processes is difficult, demonstrating the relevance of the lncRNA transcript (e.g., NKX2-1-AS1) to gene regulation and cellular behavior could help recognize multiple functions that are attributed to a particular genomic region (e.g., 14q13.3 in lung tumorigenesis).
In this study, we found evidence for a role for the NKX2-1-AS1 transcript in the regulation of lung gene expression. We were able to detect NKX2-1-AS1 transcript by various methods, including qPCR and 5′-RACE, and showed that this transcript is more stable than the NKX2-1 transcript. Furthermore, NKX2-1-AS1 transcripts are initiated at multiple start sites within a 500 bp window. We found that loss of NKX2-1-AS1 expression does not affect expression of genes in cis (including NKX2-1) with the exception of PAX9, a gene ∼140 kb 3′ to NKX2-1-AS1, which is moderately regulated by reduction of NKX2-1-AS1 levels. Instead, the NKX2-1-AS1 transcript affects the expression of genes in trans; in particular, genes involved in cell adhesion and in PD-L1/PD-1 signaling pathways in different cell lines were upregulated following NKX2-1-AS1 knockdown.
Of particular interest was the finding that NKX2-1-AS1 inhibits the expression of CD274 (PD-L1) mRNA and protein. PD-L1 is expressed in tumors of many different origins, including NSCLC, and binds to the programmed death-1 (PD-1) receptor expressed on T-cells, B-cells, dendritic cells and natural killer T-cells39. High PD-L1 expression enables cancers to evade the host immune system40, thereby indirectly making them more aggressive. In fact, NSCLC patients with high PD-L1 expression exhibit poor overall survival41 and anti-PD-L1 and/or anti-PD-1 are now being used to treat many forms of cancer42,43, although only approximately 20% of patients respond to this type of treatment.
It has been previously shown that NKX2-1 positively regulates CD274 expression33. Now we show that NKX2-1-AS1 negatively regulates CD274 transcription by interfering with the recruitment of NKX2-1 protein to the CD274 promoter. As we have shown that NKX2-1-AS1 is immunoprecipitated with NKX2-1 protein, it is possible that NKX2-1-AS1 acts as a decoy, preventing the binding of NKX2-1 protein to the CD274 promoter. Further studies will be focused on analyzing these specific interactions.
We also observed that reducing the expression of NKX2-1-AS1 increases migration of tumor cells with minimal effect on cell proliferation. This role for NKX2-1-AS1 appears to be mediated through its suppression of the transcription of several genes involved in cell adhesion, including PTPN1, which promotes migration and invasion in breast cancer cells31 and in epidermal dendritic cells32 and dephosphorylates β-catenin, adding to the adhesive properties of the cells by activation of cadherin44, and CLDN1, which encodes a tight junction component necessary for cell migration in skin wound healing and in human lung carcinoma cells45,46. This suggests that levels of NKX2-1-AS1 can modulate cell motility, a tumorigenic property that controls the dissemination and spreading of primary tumors.
Interestingly, however, the knockdown of NKX2-1-AS1 did not result in reduced cell growth, cell cycle arrest, or an increase in apoptosis. These results stand in contrast to those of a recent study by Balbin et al.11, which suggested a role for NKX2-1-AS1 in cell growth, since the down-regulation of NKX2-1-AS1 by siRNAs that were individually transfected in H441 cells produced changes in cell number up to 100 h in culture. In initial experiments, we had tested the effect of reducing NKX2-1-AS1 by individual siRNAs, and also observed a significant reduction in cell growth at 72 h post transfection (data not shown). However, in order to reduce non-specific effects of the siRNAs, we combined three different siRNAs, each of which was able to reduce NKX2-1-AS1 expression>50%, and the combination of which resulted in 90% reduction in NKX2-1-AS1 at 72 h post transfection. Because the effect of this combination on cell number was not significant in our study, it suggests that an experiment using only one siRNA might produce off-target effects.
In this study, we have characterized molecular and functional features of the NKX2-1-AS1 mature transcript. However, we cannot establish whether the process of NKX2-1-AS1 transcription itself controls the transcription of NKX2-1 or vice versa, or whether this event has regulatory or biological relevance. Answering these questions will require a different approach. These studies will be the focus of future studies. For example, the transcription of the lncRNA HAND2-AS1 (upperhand, Uph), which is divergent to the heart developmental transcription factor HAND2, has been shown to be necessary for HAND2 transcription by establishing a permissive chromatin environment; in that work, however, the function of the Uph mature transcript itself was not evaluated47.
Further understanding of the mechanisms regulating PD-L1 expression could lead to new approaches to treat aggressive malignancies48. Our data suggest that high levels of NKX2-1-AS1 are important for maintaining low levels of PD-L1 expression, presumably to limit the capacity of tumor cells to evade the immune system defenses.
In summary, this study is the first to show the role of the NKX2-1-AS1 transcript in lung gene regulation in the context of human lung cancer. Increased expression of NKX2-1-AS1 in lung carcinomas might be a protective mechanism by which cancer cells limit cell migration, metastatic spread and immune system evasion. These findings, therefore, could lead to the discovery of new mechanisms to target in future therapeutic interventions.
Methods
Cell lines
NKX2-1-AS1 is specifically detected in carcinoma cells of lung origin22. We selected for our studies human lung carcinoma cell lines that are known to express both NKX2-1 and NKX2-1-AS1: NCI-H441 (derived from a papillary adenocarcinoma of the lung) and NCI-H661 (derived from lung epithelial cells from large cell lung cancer metastasized to lymph nodes). We used as negative controls cells that express low or undetectable levels of NKX2-1 nor NKX2-1-AS1: NCI-H1299 (derived from NSCLC cells metastasized to lymph nodes) and A549 cells (derived from a lung epithelial carcinoma). In addition, we tested the expression in Calu-6 (derived from an anaplastic carcinoma of lung origin) and BEAS-2B (a virally transformed lung/bronchus epithelial cell line) [American Type Culture Collection (ATCC)]. Cells were grown in the conditions recommended by ATCC. The 14q13.3 chromosomal region is known to be amplified in NCI-H661 and not amplified in NCI-H4414.
RNA isolation and qPCR
RNA purification, reverse transcription and quantitative real-time PCR (qPCR) were performed as described previously using the OneStep Real-Time PCR System (Applied Bio-systems)49. Data was normalized to β-actin. qPCR assays are listed in Supplementary Table S1. The identity of NKX2-1 and NKX2-1-AS1 qPCR products was confirmed by DNA sequencing.
Human lung samples
Total RNA and the corresponding genomic DNA from NSCLC and adjacent control tissue were purchased from OriGene Technologies, Inc. Sample diagnosis was verified at OriGene by an independent board-certified pathologist. Patient demographic and clinical information for each sample were provided by OriGene. Normal human lung and thyroid total RNAs were purchased from Clontech/Takara Bio USA, ThermoFisher and Zymogen.
5′ RACE analysis
Rapid Amplification of cDNA Ends (RACE) was used to identify the 5′ ends of the NKX2-1-AS1 transcript in primary human samples. Human thyroid RNA was used as this tissue has higher levels of NKX2-1-AS1 expression than the lung and was sufficient for the detection limits of this assay. Total RNA (1ug) was used for first strand cDNA synthesis using Smarter® RACE 5′ (Takara/Clontech Inc.) following the manufacturer’s instructions with modifications. In the rapid amplification step with the NKX2-1-AS1 specific primer GSP1 (Supplementary Table S1) we increased the number of cycles to 30 to improve amplification of transcripts with low expression. To reduce the amplification of unwanted products during the sequential nested PCR with the specific NKX2-1-AS1 nested primer NGSP1 (Supplementary Table S1), the annealing temperature was increased to 70 °C and the number of cycles to 30. PCR products were separated by gel electrophoresis and the fragments purified using a gel extraction kit (Qiagen). The fragments were cloned into the vector provided in the RACE kit. DNA from isolated clones was purified using the QIAprep kit (Qiagen) and sequenced using NGSP1 primer, M13F, and M13R primers from the vector.
NKX2-1 RNA isoform analysis
By qPCR we analyzed the level of expression of the major variants of NKX2-1 mRNA. One TaqMan assay spanning exons 1 and 2 of NKX2-1 variant 1 and exons 2 and 3 of variant 2 was used to determine the level of the main NKX2-1 mRNA variants. A second TaqMan Assay spanning exons 1 and 2 of variant 2 was used to detect only NKX2-1 variant 2 (Supplementary Table S1).
RNA half-life analysis
To measure NKX2-1-AS1 half-life, NCI-H441 cells were seeded into 6-well plates at a density of 2 × 105 cells/well and cultured for 24 hours to reach 60% confluency. To inhibit transcriptional activity, 5 mg/ml of actinomycin D (Sigma) were added to the medium. Cells were harvested at 15 min, 30 min, 45 min, 1 hr, 2 hr, 3 hr, 4 hr, 5 hr, 6 hr, 8 hr, and 24hrs. Total RNA was purified using RNeasy kit (Qiagen) and treated with DNA-Free kit (Qiagen). Isolated total RNA (1ug) was reverse transcribed (RT) and analyzed by qPCR using the NKX2-1-AS1 and NKX2-1 TaqMan assays (Supplementary Table S1).
RNAi gene knockdown
We performed siRNA transfections to down-regulate NKX2-1-AS1 levels. We tested four different custom made Silencer® Select siRNAs designed to target the second exon of NKX2-1-AS1 and the region of the first exon not overlapping with the NKX2-1 sequence NKX2-1-AS1 siRNA A, B, C and D (Supplementary Table S1). We selected the three siRNAs (A, B and D) that individually caused >50% down regulation of NKX2-1-AS1 at 48 h post transfection. The mix of these three siRNAs resulted in a reduction of >90% of NKX2-1-AS1 levels at 72 h in culture. We seeded 150,000 cells/well in 6-well plates. Using Lipofectamine RNAiMAX (Thermo Fisher Scientific), 10uM of each siRNA or a non-silencing negative control RNA were added in Opti-MEM to 70% confluent cells. Cells were harvested at 24, 48, and 72 h post transfection and processed for total RNA or total protein isolation43, using either TRIzol or spin-column RNA extraction, followed by treatment with DNA-Free kit.
NKX2-1-AS1 cloning and overexpression
The full-length NKX2-1-AS1 (1,775 bp) was synthesized and cloned in the Nhe-BamH1 sites of the pCDNA3.1+ vector (GenScript). Overexpression of NKX2-1-AS1 or empty vector control was performed in H441 or A549 cells using Lipofectamine LTX with Plus™ reagent (Life Technologies). RNA was isolated after 48 h and expression of NKX2-1-AS1 and NKX2-1 and other genes determined by RT-qPCR.
Promoter-Luciferase activity studies
A 1.1 kb fragment upstream of the transcription start site of the human CD274 gene was synthesized and cloned in the pGL3-basic plasmid (Promega) (CD274-Luc) (GenScript). A549 cells were transfected with various constructs using Lipofectamine 3000 (ThermoFisher); the total amount of DNA used in each transfection was maintained constant by completion with pCDNA3.1 + plasmid. −1kbCD274-Luc (4ug) or 0-Luc control were transfected in the presence of various concentrations of pCDNA3.1-NKX2-1-AS1 vector (0, 0.5, 1, 2.5, 3, 5, 7 ug) alone or in combination with the pCDNA3.1-NKX2-1 vector (2ug). pCMV-Green Renilla luciferase vector construct (ThermoFisher) was co-transfected to measure transfection efficiency. Dual-Luciferase Reporter assay system (Promega) was used to measure luciferase activity in a Biotex Synergy 2 instrument following the manufacturer’s protocol. Total protein concentration and Renilla luciferase activity were used for normalization. RT-qPCR analyses were performed on the same samples to assess level of overexpression of NKX2-1 and NKX2-1-AS1 and endogenous CD274 using assays described in Supplementary Table S1.
Western blots
Protein extracts were run on Nu-Page 4-12% Bis-Tris polyacrylamide gels (Invitrogen) and transferred to nitrocellulose membranes in an iBlot2 (Thermo Fisher Scientific). Blots were treated with Odyssey blocking buffer (LI-COR) at room temperature for 1 h and incubated with rabbit monoclonal TTF1 (NKX2-1) antibody (Abcam ab133737; 1:4000) and mouse monoclonal β-actin antibody (Sigma A54441; 1:4000) overnight at 4 °C overnight, and detected with anti-rabbit HRP conjugated secondary antibody (company catalog; 1:5000), and detected with anti-rabbit IgG DyLight 800 from (Cell Signaling Technology 5151S; 1:8000 in 1XTBST) and anti-mouse IgG DyLight 680 (Invitrogen 35518; 1:8000 in 1XTBST) and quantitated using a LI-COR imaging system. PD-L1 and PTPN1 (PTP1B) proteins were similarly analyzed using mouse PD-L1 antibody (Abcam ab210931; 1:4000) and rabbit PTP1B antibody (Cell Signaling Technology 5311S; 1:2000), respectively.
Growth, proliferation and apoptosis assays
To determine the effect of NKX2-1-AS1 levels on cell growth, H441 cells were transfected with the mix of three siRNAs or control as described above, cultured, trypsinized and counted at 0 h, 24 h, 48 h, and 72 h after seeding. Proliferation was determined with Click-iT® EdU Flow Cytometry Assay Kit (Invitrogen) coupled to Alexa Fluor®488 dye and analyzed by standard flow cytometry methods at the Boston University Flow Cytometry Core to determine the percentage of cells in each cell cycle phase. Apoptosis was determined using ApoDETECT™ (Invitrogen) that uses Annexin V-FITC and propidium iodide to detect cell surface exposure of phosphatidylserine in viable cells by flow cytometry following the manufacturer’s protocol.
Wound healing, migration and invasion assays
We tested the effect of reduced NKX2-1-AS1 levels on the wound healing behavior of H441 cell monolayers upon induced injury. Healing of a scratch in confluent H441 cells transfected with non-silencing control or NKX2-1-AS1 siRNAs was analyzed using the analytical function in Adobe Photoshop CS4 Suite. Migration and invasion were determined using CytoSelect™ 24-well Cell Migration and Invasion Assay (8 µm, Fluorometric Format) (Cell Biolabs, Inc.). Cell migration analysis was performed 48 h and 72 h after NKX2-1-AS1 siRNAs transfection using polycarbonate membrane inserts. After 24 h migrating cells were dissociated from the membrane and detected in a fluorescence reader using the CyQuant® GR Dye (Invitrogen). For invasion assays we used basement membrane-coated inserts following the same protocol.
Microarray analysis
Affymetrix Human Gene 2.0 ST arrays were used to profile RNA (extracted using either TRIzol or spin columns) from H441 cells treated with the pool of three siRNAs targeting the lncRNA NKX2-1-AS1 or non-silencing control (n = 3 per group). Arrays were normalized using the Robust Multiarray Average (RMA) with the affy R package (version 1.36.1) and an Entrez Gene-specific probeset mapping (version 16.0.0) from the Molecular and Behavioral Neuroscience Institute at the University of Michigan. Differential expression was assessed using the moderated (empirical Bayesian) t test implemented in the limma R package (version 3.14.4); specifically, simple linear models were created using lmFit, adjusting for RNA extraction protocol, followed by empirical Bayesian adjustment with eBayes. Correction for multiple hypothesis testing was accomplished using the Benjamini-Hochberg false discovery rate (FDR), performed across the genes that were expressed above the median value of at least one of the 12 microarrays. All analyses were performed using the R environment for statistical computing (version 2.15.1). Gene Set Enrichment Analysis (GSEA) (version 2.0.13) was used to perform pre-ranked analyses (default parameters with random seed 1234), using a list of all Entrez Gene identifiers ranked by the t statistic computed between NKX2-1-AS1 knockdown and control, and the Entrez Gene versions of the Biocarta, KEGG, Reactome, Gene Ontology (GO), cytoband, and transcription factor and microRNA motif gene sets obtained from the Molecular Signatures Database (MSigDB), version 4.0. Detailed analyses are included in the Supplementary Materials.
NKX2-1-AS1 locus amplification analyses
Genomic DNA corresponding to the tumors described in Table 1 obtained from OriGene was used to determine the copy number of the NKX2-1-AS1 locus using a custom designed SYBR Green Assay (Supplementary Table S1). To normalize we used a SYBR Green assay designed to amplify LINE1 loci (Supplementary Table S1), a repetitive sequence with conserved copy number in most human normal and neoplastic cells26,27. Normal human iPSC DNA was used to generate the calibration curve.
Chromatin Immunoprecipitation Assays (ChIP)
Analyses were performed with the MAGnify Chromatin Immunoprecipitation kit (ThermoFisher) according to the manufacturer’s protocol with minor modifications. Cells were trypsinized, washed with 1 x PBS and fixed with 1% formaldehyde in 1x PBS at room temperature with rotation for 10 min. Chromatin was sonicated to 300–500 bp fragments using Pico Bioruptor (Diagenode) for 12 cycles 30 sec on, 30 sec off. Lysate was centrifuged at 20,000 × g for 5 min at 4 °C to remove insoluble material. DNA content was measured in a NanoDrop and equal amount of soluble chromatin was used for ChIP assays. 5ug of anti-TTF1 antibody (NKX2-1 protein)(Abcam, ab133737) and nonspecific rabbit IgG were added to sheared chromatin and incubated with rotation at 4 °C overnight. ChIPs were analyzed by qPCR using 2X Fast SYBR master mix (ThermoFisher) and promoter-specific assays (Supplementary Table S1).
RNA Immunoprecipitation Assays (RIP)
Analyses were performed with the Nuclear RNA-Binding Protein Immunoprecipitation Kit (Millipore) according to manufacturer’s protocol with minor modifications. Cells were grown in 150 mm culture dish and allowed to reach 90% confluency. Cells were crosslinked using 1% formaldehyde in media, and washed with 1x PBS supplemented with protease and RNAse inhibitors. Nuclear extracts were isolated and sonicated to 100–300 bases fragments using Pico Bioruptor (Diagenode) for 10 cycles (30 sec on and 30 sec off). Lysates were centrifuged at 20,000 × g for 10 min at 4 °C to remove insoluble material. RIP incubations were performed overnight at 4 °C with rotation using 5ug or 10 ug of anti-TTF1 (NKX2-1) antibody (Abcam, ab133737) or nonspecific rabbit IgG. One aliquot was saved for input. RNA purification was performed using RNeasy kit (Qiagen). RT was performed using SensiScript kit (Qiagen) and qPCR was performed using 2X Fast Taqman master mix (ThermoFisher) and assays to detect NKX2-1-AS1 lncRNA and NKX2-1 mRNA as control (Supplementary Table S1).
Statistical Analysis
Data were obtained from at least three independent experiments (n ≥ 3) and presented as mean ± SEM unless other stated in the Figure legend. The significance of differences was calculated using parametric t-test for two-group unpaired comparisons, two-tails; non-parametric t-tests were used to analyze the RIP and ChIP assays. p < 0.05 was considered statistically significant. For tumor samples, we used the Wilcoxon Mann Whitney test to analyze expression values of NKX2-1 and NKX2-1-AS1 in adenocarcinoma (n = 8) relative to their corresponding non-tumor control versus squamous cell tumors (n = 8) relative to their corresponding non-tumor controls. All statistical analyses were performed in GraphPad Prism 7.0c.
Data Availability
The datasets generated and analyzed during this study are available in the Gene Expression Omnibus (GEO) repository, Series GSE104691. This work was partially supported by R01 HL127426 (MIR) and CTSA grant UL1-TR000157.
References
Campbell, J. D. et al. Distinct patterns of somatic genome alterations in lung adenocarcinomas and squamous cell carcinomas. Nat. Genet. 48, 607–616 (2016).
Weir, B. A. et al. Characterizing the cancer genome in lung adenocarcinoma. Nature 450, 893–898 (2007).
Perner, S. et al. TTF1 expression in non-small cell lung carcinoma: association with TTF1 gene amplification and improved survival. J. Pathol. 217, 65–72 (2009).
Kwei, K. A. et al. Genomic profiling identifies TITF1 as a lineage-specific oncogene amplified in lung cancer. Oncogene 27, 3635–3640 (2008).
Harris, T. et al. Both gene amplification and allelic loss occur at 14q13.3 in lung cancer. Clin. Cancer Res. 17, 690–699 (2011).
Hwang, D. H. et al. KRAS and NKX2-1 Mutations in Invasive Mucinous Adenocarcinoma of the Lung. J. Thorac. Oncol. 11, 496–503 (2016).
Hsu, D. S. et al. Characterizing the developmental pathways TTF-1, NKX2-8, and PAX9 in lung cancer. Proc. Natl. Acad. Sci. USA 106, 5312–5317 (2009).
Volders, P. J. et al. LNCipedia: a database for annotated human lncRNA transcript sequences and structures. Nucleic Acids Res. 41, D246–51 (2013).
Volders, P. J. et al. An update on LNCipedia: a database for annotated human lncRNA sequences. Nucleic Acids Res. 43, 4363–4364 (2015).
Herriges, M. J. et al. The NANCI-Nkx2.1 gene duplex buffers Nkx2.1 expression to maintain lung development and homeostasis. Genes Dev. 31, 889–903 (2017).
Balbin, O. A. et al. The landscape of antisense gene expression in human cancers. Genome Res. 25, 1068–1079 (2015).
Quinn, J. J. & Chang, H. Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 17, 47–62 (2016).
Hon, C. C. et al. An atlas of human long non-coding RNAs with accurate 5′ ends. Nature 543, 199–204 (2017).
Li, S. & Breaker, R. R. Identification of 15 candidate structured noncoding RNA motifs in fungi by comparative genomics. BMC Genomics 18, 785-017–4171-y (2017).
Amaral, P. P. et al. Genomic positional conservation identifies topological anchor point RNAs linked to developmental loci. Genome Biol. 19, 32-018–1405-5 (2018).
Ward, M., McEwan, C., Mills, J. D. & Janitz, M. Conservation and tissue-specific transcription patterns of long noncoding RNAs. J. Hum. Transcr. 1, 2–9 (2015).
Sang, H., Liu, H., Xiong, P. & Zhu, M. Long non-coding RNA functions in lung cancer. Tumour Biol. 36, 4027–4037 (2015).
Peng, W. X., Koirala, P. & Mo, Y. Y. LncRNA-mediated regulation of cell signaling in cancer. Oncogene (2017).
Huarte, M. et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response. Cell 142, 409–419 (2010).
White, N. M. et al. Transcriptome sequencing reveals altered long intergenic non-coding RNAs in lung cancer. Genome Biol. 15, 429-014–0429-8 (2014).
Zhao, W., Luo, J. & Jiao, S. Comprehensive characterization of cancer subtype associated long non-coding RNAs and their clinical implications. Sci. Rep. 4, 6591 (2014).
Krupp, M. et al. CellLineNavigator: a workbench for cancer cell line analysis. Nucleic Acids Res. 41, D942–8 (2013).
Gruber, M. P. et al. Human lung project: evaluating variance of gene expression in the human lung. Am. J. Respir. Cell Mol. Biol. 35, 65–71 (2006).
Sato, T. et al. PRC2 overexpression and PRC2-target gene repression relating to poorer prognosis in small cell lung cancer. Sci. Rep. 3, 1911 (2013).
Kuner, R. et al. Global gene expression analysis reveals specific patterns of cell junctions in non-small cell lung cancer subtypes. Lung Cancer 63, 32–38 (2009).
Zhao, X. et al. An integrated view of copy number and allelic alterations in the cancer genome using single nucleotide polymorphism arrays. Cancer Res. 64, 3060–3071 (2004).
Kubo, T. et al. MET gene amplification or EGFR mutation activate MET in lung cancers untreated with EGFR tyrosine kinase inhibitors. Int. J. Cancer 124, 1778–1784 (2009).
Yoon, J. H. et al. LincRNA-p21 suppresses target mRNA translation. Mol. Cell 47, 648–655 (2012).
Sigova, A. A. et al. Divergent transcription of long noncoding RNA/mRNA gene pairs in embryonic stem cells. Proc. Natl. Acad. Sci. USA 110, 2876–2881 (2013).
Carrieri, C. et al. Long non-coding antisense RNA controls Uchl1 translation through an embedded SINEB2 repeat. Nature 491, 454–457 (2012).
Liu, X. et al. PTP1B promotes aggressiveness of breast cancer cells by regulating PTEN but not EMT. Tumour Biol. 37, 13479–13487 (2016).
Martin-Granados, C. et al. A key role for PTP1B in dendritic cell maturation, migration, and T cell activation. J. Mol. Cell. Biol. 7, 517–528 (2015).
Maeda, Y. et al. Kras(G12D) and NKX2-1 haploinsufficiency induce mucinous adenocarcinoma of the lung. J. Clin. Invest. 122, 4388–4400 (2012).
Sun, W., Yang, Y., Xu, C. & Guo, J. Regulatory mechanisms of long noncoding RNAs on gene expression in cancers. Cancer. Genet. 216-217, 105–110 (2017).
Winslow, M. M. et al. Suppression of lung adenocarcinoma progression by NKX2-1. Nature 473, 101–104 (2011).
Engreitz, J. M. et al. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 539, 452–455 (2016).
Clark, M. B. et al. Genome-wide analysis of long noncoding RNA stability. Genome Res. 22, 885–898 (2012).
Yoon, J. H., Kim, J. & Gorospe, M. Long noncoding RNA turnover. Biochimie 117, 15–21 (2015).
Konishi, J. et al. B7-H1 expression on non-small cell lung cancer cells and its relationship with tumor-infiltrating lymphocytes and their PD-1 expression. Clin. Cancer Res. 10, 5094–5100 (2004).
Zitvogel, L. & Kroemer, G. Targeting PD-1/PD-L1 interactions for cancer immunotherapy. Oncoimmunology 1, 1223–1225 (2012).
Wang, A. et al. The prognostic value of PD-L1 expression for non-small cell lung cancer patients: a meta-analysis. Eur. J. Surg. Oncol. 41, 450–456 (2015).
Topalian, S. L. et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N. Engl. J. Med. 366, 2443–2454 (2012).
Brahmer, J. R. et al. Safety and activity of anti-PD-L1 antibody in patients with advanced cancer. N. Engl. J. Med. 366, 2455–2465 (2012).
Xu, G., Arregui, C., Lilien, J. & Balsamo, J. PTP1B modulates the association of beta-catenin with N-cadherin through binding to an adjacent and partially overlapping target site. J. Biol. Chem. 277, 49989–49997 (2002).
Volksdorf, T. et al. Tight Junction Proteins Claudin-1 and Occludin Are Important for Cutaneous Wound Healing. Am. J. Pathol. 187, 1301–1312 (2017).
Shiozaki, A. et al. Claudin 1 mediates TNFalpha-induced gene expression and cell migration in human lung carcinoma cells. PLoS One 7, e38049 (2012).
Anderson, K. M. et al. Transcription of the non-coding RNA upperhand controls Hand2 expression and heart development. Nature 539, 433–436 (2016).
Chen, J., Jiang, C. C., Jin, L. & Zhang, X. D. Regulation of PD-L1: a novel role of pro-survival signalling in cancer. Ann. Oncol. 27, 409–416 (2016).
Tagne, J. B. et al. Genome-wide analyses of NKX2-1 binding to transcriptional target genes uncover novel regulatory patterns conserved in lung development and tumors. PLoS One 7, e29907 (2012).
Acknowledgements
We thank Drs Jeannie Lee, Darrell Kotton and Jay Mizgerd for insightful comments about this work and Drs Ross Summer and Freddy Romero for critical reading of the manuscript. We also thank Sana Krimberg and Dr. Zhaotang Wu for technical support.
Author information
Authors and Affiliations
Contributions
H.K. contributed to hypothesis development, experimental design, human sample analysis, data interpretation and manuscript writing, G.M., L.M. and J.B.T. performed experimental work, A.C.G. analyzed microarray data, Y.C. contributed to hypothesis development, and M.I.R. contributed to hypothesis development, experimental design, data interpretation, and manuscript writing.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kathuria, H., Millien, G., McNally, L. et al. NKX2-1-AS1 negatively regulates CD274/PD-L1, cell-cell interaction genes, and limits human lung carcinoma cell migration. Sci Rep 8, 14418 (2018). https://doi.org/10.1038/s41598-018-32793-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-018-32793-5
Keywords
This article is cited by
-
Integrative transcriptomic and structural modeling reveal CASP1, TLR3, PYCARD, and CD274 as immune-modulatory drivers in breast cancer
Naunyn-Schmiedeberg's Archives of Pharmacology (2026)
-
Long non-coding RNAs in cancer: multifaceted roles and potential targets for immunotherapy
Molecular and Cellular Biochemistry (2024)
-
Identification of peroxiredoxin II and its related molecules as potential biomarkers of dermal mesenchymal stem cell homing using network analysis
Applied Biological Chemistry (2022)
-
Human embryonic mesenchymal lung-conditioned medium promotes differentiation to myofibroblast and loss of stemness phenotype in lung adenocarcinoma cell lines
Journal of Experimental & Clinical Cancer Research (2022)
-
The regulatory role of long non- coding RNAs as a novel controller of immune response against cancer cells
Molecular Biology Reports (2022)
