
Abstract
Nuts are the natural source of healthy lipids, proteins, and omega-3. They are susceptible to fungal and mycotoxins contamination because of their high nutritional value. Twenty-five species comprising 12 genera were isolated from 80 samples of dried fruits and nuts using the dilution plate method. Peanut recorded the highest level of contamination followed by coconut; almond and raisin were the lowest. Aspergillus was the most prevalent genus and A. niger, was the most dominant species. The morphological identification of the selected A. niger isolates as they were detected in high frequency of occurrence was confirmed by using 18SrRNA sequence. Ochratoxin biosynthesis gene Aopks was detected in the tested isolates. Lipase production by the selected A. niger isolates was determined with enzyme activity index (EAI) ranging from 2.02 to 3.28. A. niger-26 was the highest lipase producer with enzyme activity of 0.6 ± 0.1 U/ml by the trimetric method. Lip2 gene was also detected in the tested isolates. Finally, the antibacterial and antibiofilm efficiency of crude lipase against some human pathogens was monitored. Results exhibited great antibacterial efficacy with minimum bactericidal concentration (MBC) of 20 to 40 µl/100 µl against Escherichia coli, Pseudomonas aeruginosa, Proteus mirabilis, and Methicillin-resistant Staphylococcus aureus (MRSA). Interestingly, significant anti-biofilm efficacy with inhibition percentages of 95.3, 74.9, 77.1 and 93.6% was observed against the tested pathogens, respectively.
Similar content being viewed by others


Introduction
Dried fruits and nuts are enriched source of healthy fatty acids, protein, potassium, dietary fibers and bioactive compounds1. They protect the mankind from the risks of obesity, cardiovascular illnesses, type 2 diabete and hypertension2, 3. Alternaria, Aspergillus, Candida, Fusarium, Mucor, Rhizopus, Penicillium, Trichoderma, and Cladosporium are the most common genera causing nuts spoilage, and their ingestion may cause mycoses especially in immunocompromised patients4,5.
Many species of fungi produce mycotoxins, secondary metabolites of small molecular sizes (MW < 300) that are toxic to humans and animals cause mycotoxicoses when ingested, leading to cancer and liver diseases6, 7. A total of 145 secondary metabolites were discovered in Aspergillus section Nigri; among them are ochratoxin A (OTA), which are the most toxic to humans and animals5. OTA is the causative agent of Balkan endemic nephropathy, urothelial tumors, and testicular cancer in humans8,9,10.
Lipase, an enzyme belonging to the serine hydrolase class, catalyzes the hydrolysis of fats and oils to glycerol and fatty acids without requiring cofactors11. Fungi are considered the best producers of lipase among all microorganisms, especially, Aspergillus niger which was generally recognized as safe (GRAS) by Food and Drug Administration (FDA) in the United States12. Lipase has a wide range of industrial applications, such as in the food industry, as detergent additives, pharmaceutical industry, and biofuel production; therefore, the universal demand for the lipase enzyme is increasing13.
Enzymes including isomerases, lyases, oxidoreductases, transferases, esterases and hydrolases have been reported to induce antibacterial efficacy14. Lipase is a hydrolytic enzyme, has antimicrobial and antifouling properties15. However, its mode of action and its effects in most of the cases have not been clarified fully14.
Biofilm is a complex medium involving live and dead bacterial cells, exopolysaccharides, proteins and carbohydrates on a material surface with a serious problem in biomedical applications16. Several steps were involved in biofilms development beginning with surface adherence, microcolony formation, maturation and finally detachment stages17. Biofilms protect pathogenic bacteria from human immune system, antibiotics and severe environmental conditions18. Several bio-active and chemically synthesized compounds have been performed to suppress biofilm formation by pathogenic bacteria19,20,21,22. Revitalize aminoglycosides also have been used to inhibit biofilm and pathogenic bacterial infections23. Attenuating motility properties can be considered as highly potential for controlling biofilm formation since attachment was one of the main steps in biofilm formation24,25. Using enzymes is also a good policy for biofilm elimination because enzymes are rabidly eco-friendly and degradable26,27.
This study was established for the isolation and identification of mycobiota associated with four kinds of nuts, determination the ochratoxigenic potential of some A. niger isolates, their lipolytic activity, and finally studying the ability of crude lipase from A. niger to inhibit the growth and biofilm formation of some human pathogens.
Results
Mycobiota contaminating nuts
Twenty-five fungal species comprising 12 genera were obtained from the 80 tested samples of nuts by using dilution plate method. Aspergillus was the most prevalent genus as it was isolated from 100% of the samples. Penicillium was the second genus in frequency as it was isolated from 62.5% of total samples. From the above genera A. niger, A. flavus, P. chrysogenum and P. oxalicum were the most frequent species (Table 1). Rhizopus stolonifer was isolated from 60% of peanut, 45% of almond and 35% of raisin but not detected in coconut. The remaining genera and species were isolated in rare frequency accounting collectively 1.59 × 103 CFU/g as illustrated in Table 1.
Multiple alignment of different A. niger isolates
The 5.8S gene in rDNA sequences were subjected to multiple alignments using the BioEdit program (http://www.mbio.ncsu.edu/BioEdit/page2.html). Among the five isolates of A. niger, 5.8S gene nucleotide sequences showed 98% similarity all strains. When the sequences were aligned with the database sequences, they showed 95% similarity with A. niger strain AHBR5, except the A. niger-27 similarity sequence, which shared 98% similarity with A. niger strain AHBR5 (Fig. 1). Phylogenetic tree was drawn with MEGA 7.1 program show that more similarity among Aspergillus niger and low similarity with A. niger AHBR5 (Fig. 2).

Comparison alignment of nucleotide sequences of ITS region in five A. niger isolates with different relation nucleotide sequences of some strains at NCBI database using BioEdit program.

Phylogenetic tree and relationship among five strains of A. niger compared with some A. niger strains at NCBI.
Ochratoxigenic potential of A. niger
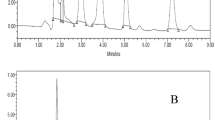
Our results indicated that all the tested A. niger isolates had ability to produce ochratoxins by using flourometric method with variable levels (2.6–3.2 ppb) with the highest reading recorded by A. niger-27 recovered from coconuts as shown in Table 2.
Detection of ochratoxins biosynthesis genes
Polymerase chain reaction (PCR) was applied using two sets of primer for gene involved in ochratoxin biosynthetic pathway. Bands of the fragments of Aopks gene can be visualized in all tested A. niger isolates at 549 bp (Fig. 3).

PCR amplification of Aopks genes (549 bp) for A. niger isolates. Whereas, (1) A. niger-26 and (2) A. niger-27, (3) A. niger-28, (4) A. niger-29 and (5) A. niger-30. Sample lanes from different gels have been juxtaposed together in this figure.
Preliminary screening of A. niger isolates for lipase production
The tested A. niger isolates had ability to produce lipase enzyme in solid medium containing tween 80 with enzyme activity index (EAI) ranging from 2.02 to 3.28 as summarized in Table (3). White precipitate diameter was between 11.5 ± 0.5 and 21.8 ± 7.42 mm. A. niger-29 showed the highest diameter 21.8 ± 7.42 mm and the lowest was observed in A. niger-27 with 11.5 ± 0.5 mm.
Assay of lipase enzyme
Lipase activity was determined in liquid medium by using trimetric method showed that A. niger-26 obtained from almond recorded the highest lipase activity (0.6 ± 0.1 U/ml-min) followed by A. niger-30 (0.3 ± 0.1 U/ml-min), A. niger-28 and A. niger-29 with the same reading (0.233 ± 0.11547 U/ml-min) and A. niger-27 was the least (0.2 ± 0.1 U/ml-min) (Table 3).
Detection of A. niger lip2 gene
PCR was performed for Lip2 gene detection in the tested A. niger isolates using two sets of primers. Lip2 gene was detected at 1276 bp in all the A. niger isolates (Fig. 4).

PCR amplification of Lip2 genes (1276 bp) for A. niger isolates. whereas, (A) A. niger-29, (B) A. niger-26, (C ) A. niger-28, (D) A. niger-27 and (E) A. niger-30.
Studying the virulence properties of target human pathogens in presence of crude lipase obtained from A. niger
In the current study, we extended the utility of using crude lipase from A. niger to explore its potential as antibacterial against some human pathogens. This was performed using INT reduction assay. Results exhibited an excellent effect of crude lipase against both Gram negative and Gram positive tested strains. Where MIC ranged from 10 to 20 µl/100 µl and MBC from 20 to 40 for Escherichia coli, Proteus mirabilis, Pseudomonas aeruginosa, and Methicillin-resistant Staphylococcus aureus (MRSA) Table 4.
Antibiofilm activity lipase enzyme
In our study crude lipase from A. niger MW029470 was examined as antibiofilm agent against four human pathogens by spectrophotometric methods. The ability of the four tested human pathogens Escherichia coli, Pseudomonas aeruginosa, Proteus mirabilis and Methicillin-resistant Staphylococcus aureus (MRSA) to form biofilm were confirmed before treatment with lipase as shown in Fig. 5A–D{C}, respectively. The results exhibited significant inhibition for biofilm formation in the four tested pathogens. The highest significant percentages of inhibition were 95.3, 74.9, 77.1 and 93.6 for Escherichia coli, Pseudomonas aeruginosa, Proteus mirabilis, and Methicillin-resistant Staphylococcus aureus (MRSA) Fig. 5A–D{50}, respectively.

Antibiofilm activity of crude lipase produced by A. niger isolated from nuts against some human pathogenic bacteria. (A) Escherichia coli; (B) Pseudomonas aeruginosa; (C) Proteus mirabilis; (D) methicillin-resistant Staphylococcus aureus (MRSA). C: control (amount of biofilm of the tested strains). 20, 30, 40, 50, 60, and 70 µl: added volumes of crude lipase for determination the optimum volume in inhibiting biofilm. Shown are the averages from at least three independent measurements. The error bars indicate the standard deviations. Asterisk: means values are highly significant compared with control.
Scanning electron microscopy (SEM)
Results of SEM were revealed in Figs. 6 and 7. For antibacterial efficacy of crude lipase for the tested bacteria, the micrographs showing that some cells shorten and getting smaller such as Escherichia coli (Fig. 6a,b). Other cells were curved and divided like Proteus mirabilis (Fig. 6c,d). Cells of Pseudomonas aeruginosa (Fig. 6e,f) distortion occur in cell shape to spherical instead of bacillus. Finally, cells of Methicillin-resistant Staphylococcus aureus (MRSA) that begin to swell up with irregular spherical shape Fig. 6g,h.

Scanning electron microscopy micrographs of treated bacteria with crude lipase. (a,c,e,g) Untreated Escherichia coli, Proteus mirabilis, Pseudomonas aeruginosa and methicillin-resistant Staphylococcus aureus (MRSA), respectively (control). (b,d,f,h) Treated Escherichia coli, Proteus mirabilis, Pseudomonas aeruginosa and methicillin-resistant Staphylococcus aureus (MRSA) with crude lipase, respectively (treatments).

Scanning electron microscopy micrographs of biofilm structure. (a,c,e,g) Biofilm formation by untreated Escherichia coli, Proteus mirabilis, Pseudomonas aeruginosa and methicillin-resistant Staphylococcus aureus (MRSA), respectively (control). (b,d,f,h) Biofilm formation by treated Escherichia coli, Proteus mirabilis, Pseudomonas aeruginosa and methicillin-resistant Staphylococcus aureus (MRSA) with crude lipase, respectively (treatments).
SEM micrographs for biofilm structure revealed that, in control, there are typically heterogeneous distributions of biofilm with higher number of adhered cells also cells arranged in the form of aggregates or simply as individualized cells without slimy material in their vicinity (Fig. 7a,c,e,g). In contrast, to treatment with crude lipase where micrographs showing a uniform layer of cells with negligible clumping (Fig. 7b,d,f,h).
Discussion
Nuts and dried fruits are healthful foods that protect human body from many chronic diseases. Their high nutritional value makes them a suitable medium for fungal contamination. In the current study, peanut were the highest contaminated samples this may be due to the high moisture content of peanut samples in harmony with Ismail28, who reported that peanut samples were highly deteriorated with fungi than coconut. Aspergillus was the most prevalent genus followed by Penicillium. This were previously confirmed by Khosravi et al.29, who showed that Aspergillus followed by Penicillium were the most frequent genera deteriorated 60 samples of nuts. From the above genera A. niger, A. flavus, P. chrysogenum and P. oxalicum were the most frequent species (Table 1) and these results were previously obtained by Ismail28,30,31,32,33. In contrast, Aspergillus section Flavi was the highest recorded in peanuts seeds followed by Aspergillus section Nigri and Aspergillus section Circumdati was the least34. Past study by Tournas et al5 found the same results that A. niger followed by Penicillium were the most common mold in nuts and dried fruits. Rhizopus stolonifer was isolated from peanuts, almonds and raisins in high and moderate frequency of occurrence (Table 1). In a study by Abdulla35, reported that Aspergillus, Rhizopus and Penicillium genera were more frequently detected than other genera of fungi in nuts.
The molecular identification of the tested A. niger confirmed the morphological identity and more similarity among A. niger isolates was observed and low similarity with A. niger AHBR5 except A. niger-27 (Figs. 1, 2) and the obtained results in agreement with Perrone et al36,37. All the tested A. niger isolates were ochratoxin producers with variable readings by using fluorometric method (Table 2). In past investigation by Al-Sheikh38 confirmed that 57% and 60% of A. niger and A. carbonarius, respectively deteriorated peanut were ochratoxin producers. Magonli et al39, demonstrated that 32% of Aspergillus section Nigri obtained from peanut seeds in Argentinean had ability to produce ochratoxin A. Alhussaini30, found that 33.3% of Aspergillus section Nigri biserriate and one isolate of uniserriate isolated from nuts were ochratoxin A producers. The tested A. niger isolates recovered from baby foods recorded positive results for ochratoxins production40. Our obtained results were in-disagreement with past study by Palumbo & O’Keeffe41, reported that all the tested 171 isolates of Aspergillus section Nigri isolated from almonds showed negative results for ochratoxin A production. In this study, ochratoxin biosynthesis gene Aopks was detected in all the tested isolates at 549 bp (Fig. 3). The obtained results in harmony with Massi et al.42, who reported that pks gene was detected in all Aspergillus niger positive ochratoxgenic strains isolated from Brazilian foods amongst, nuts and dried fruits. Aopks genes were detected at 549 bp in A. niger isolates that had ability to produce ochratoxins43, 44. All tested A. niger and A. tubingensis isolated from beef showed positive results for the presence of pks genes45. The selected A. niger isolates were lipase producers qualitatively on Tween 80 solid medium and quantitatively by using trimetric titration method with the highest activity recorded by A. niger-26 isolated from almond (Table 3). A. niger is well-recognized to be the best producer of lipase enzyme and is favored in many industrial processes46, 47. A. niger, Fusarium oxysporum and Nectria haematococca isolated from beef luncheon were the highest lipase producers48. Rai et al.49, isolated lipase producer A. niger from some oil contaminated soil samples. Earlier studies also, confirmed that A. niger was the highest lipase producing strain50,51,52,53,54. Putri et al.12, optimized the production of lipase by A. niger by using agro-waste and revealed that 1% olive oil was the highest inducer, yielding dry lipase extract with highest activity unit (176 U/ml enzyme). Lip2 gene was visualized at 1276 bp in all the tested A. niger isolates (Fig. 4). Yang et al.55, reported Lip2 gene a novel lipase gene cloned from A. niger. Lipase exhibits antibacterial activity against Escherichia coli, Proteus mirabilis and Pseudomonas aeruginosa with MBC of 20 µl/100 µl (Table 4). This may be due to that lipase acting on the lipopolysaccharide of Gram negative cell wall as well as the esters of exopolysaccharide present in the biofilm. Furthermore, lipolytic enzyme acts on a lipid substrate Such as phospholipids and other hydrophobic molecules, to hydrolyze or esterify a bond16. Lipases are esterases capable of hydrolyzing any ester bond. They act on the lipoprotein, lipopolysaccharide and phospholipids which surrounds the peptidoglycan layer leading to the hydrolysis of the lipid bilayer. The lipopolysaccharide complex is an endotoxin present on the outer membrane of the cell wall and this toxicity leads to a wide spectrum of nonspecific pathophysiological reactions including fever, changes in white blood cell counts, disseminated intravascular coagulation, hypotension, shock and death. When lipase works on lipid A, the chances of infection are reduced56. In most of the Gram positive bacteria, lipoteichoic acids are present and the lipid tail present here plays a major role in the bacterial attachment. There is a possibility for the lipase to act on this lipid tail thereby preventing its adherence to a surface16. Our results confirmed antibacterial activity of lipase on Gram positive bacteria (MRSA) with MBC of 40 µl/100 µl (Table 4). Bacterial biofilms pose a great threat to human life not only because they involved in a lot of chronic infectious human diseases but also, they highly resistant to different antimicrobial agents. This generates a strong demand for finding suitable anti biofilm agents57. Bacterial biofilms are common populations of bacterial cells surrounded by a self-produced matrix of extracellular polymeric substances (EPS) 58. EPS surrounding mixture include various exopolysaccharides, lipids, secreted proteins some of which can form amyloid fibers and extracellular DNA59. Most of the antimicrobial agents fail to penetrate the biofilm owing to the presence of EPS which acts as a barrier protecting the bacterial cells within the biofilm. So, the remedy will be the use of compounds that able to degrade the biofilm EPS. Enzymes have been recognized to be effective for the degradation of the biofilms EPS60,61. Plants contain various anti-biofilm compounds, as they have to prevent bacterial growth on their surfaces62,63. Since lipase, an esterase, is a hydrolyzing enzyme, it is having the ability to act on the EPS produced by the organisms64, by degrading protein components and the high molecular weight lipid of the biofilm65. In the current investigation crude lipase, was examined as antibiofilm agent against four human pathogens by spectrophotometric methods. The highest significant percentages of inhibition were 95.3, 74.9, 77.1 and 93.6 for Escherichia coli, Pseudomonas aeruginosa, Proteus mirabilis and Methicillin-resistant Staphylococcus aureus (MRSA), respectively. Although, all added volumes of lipase significantly inhibited biofilm formation, the suitable volume that gives highest inhibition percentage was 50 µl (Fig. 5). Scanning electron microscopy (SEM) has been used widely for qualitative observation of biofilm before and after treatments, biofilm disturbance due to its high resolution and is usually applied in biological assays of biofilm removal effectiveness also antimicrobial treatments66,67,68. Results of SEM confirmed antbacterial and antibiofilm properties of crude lipase against the tested human pathogens.
In conclusion, Peanuts were the highest contaminated samples among the tested types. A. niger was the most isolated species from nuts. All the selected A. niger isolates were lipase producers with highest enzyme activity was recorded by A. niger MW029470 and showed positive results for the presence of Lip 2 gene. Crude lipase from A. niger MW029470 showed highly inhibition of the tested pathogens growth with MBC of 20 to 40 µl/100 µl and significantly inhibited biofilm formation of 4 biofilm former human bacterial pathogens. The significant percentages of inhibition were 95.3, 74.9, 77.1, and 93.6 for Escherichia coli, Pseudomonas aeruginosa, Proteus mirabilis and Methicillin-resistant Staphylococcus aureus (MRSA), respectively.
Materials and methods
Collection of nuts samples
Eighty samples of peanut, almond, coconut and raisin (20 samples of each type) were purchased from different supermarkets at Qena Governorate, Egypt. All samples were kept in a refrigerator until mycological analysis.
Isolation of fungi
The modified method described by Tournas et al.5 was employed for isolation of mycobiota contaminating nuts. A known weight of each sample was blended with 90 ml of 0.1% peptone in blender jar under aseptic conditions for minute. Serial dilutions were made to obtain the suitable one. One ml of the suitable dilution was poured in sterilized petri plate followed by 20 ml of rose Bengal chloramphenicol agar (RBCA) medium containing g/l (peptone; 5, glucose; 10, kH2PO4; 1, MgSO4. 7H2O; 0.5, rose Bengal; 0.05, chloramphenicol; 0.1, and agar 15.5). Triplicates of each sample were prepared. Plates were incubated for a week at 28 °C. The developed fungal colonies were counted, examined and identified (based on macro- and microscopic features)69.
Sequence analysis of 5.8S-ITS region
ITS1 and ITS2 regions together with 5.8S gene in rDNA from A. niger strains were amplified as designed by Hermosa et al.70. The purified bands were determined using the sequencer Gene analyzer 3121 in Scientific Research Center, Biotechnology and Genetic Engineering Unit, Taif University, KSA. The realized sequence was aligned using Molecular Evolutionary Genetics Analysis (MEGA) version 5.10. Then, a consensus sequence was generated from each alignment made. The sequencing data were compared against the Gene Bank database (http://www.ncbi.nlm.nih.gov/BLAST/), where a nucleotide blast program was chosen to identify the homology between the PCR fragments and the sequences on the Gene Bank database.
Accessions numbers
Sequences were deposited in GenBank under accession numbers MW029470-MW029474.
Ochratoxins production by A. niger isolates
Five isolates of Aspergillus niger with the highest number of colonies and high frequency of occurrence were tested for their ability to produce ochratoxins by cultivation in conical flasks containing 50 ml of yeast extract sucrose (YES) liquid medium with composition sucrose, 40 g, yeast extract 20 g, and distilled water, 1000 m1. Incubation the flasks at 28 °C for fifteen days71. Filtration through a fluted filter paper (Whatman 2 V, Whatmanplc, Middlesex,UK). Total ochratoxins were determined according to the method mentioned by El-Dawy et al.44 in 10 ml fungal filtrate by adding 90 ml (methanol: water) (80:20 v/v) and the filtrate was diluted (1:4) with distilled water and re-filtered through a glass-fiber filter paper. Ten milliliters of the glass-fiber filtrate were placed on Ochra test WB SR Column (VICAM, Watertown, MA, USA) and allowed to elute at 1–2 drops/s. The columns were washed twice with 10 ml of distilled water, and ochratoxins were eluted from the column by adding 1 ml of methanol HPLC and delivered in clean cuvette. 1.5 ml ochratoxin eluting agent was added and the total ochratoxins concentration were measured after calibration VICAMSeries-4 fluorometer set at 360 nm excitation and 450 nm emissions72.
Molecular detection of ochratoxin-producing genes
DNA extraction and purification were performed using DNA Promega Kit DNeasy Blood & Tissue (Valencia, CA, USA). Two published primers were used for the specific detection of ochratoxin biosynthesis genes. The sequence of primers was as following: Aopks-F '5-CAGACCATCGACACTGCATGC-'3, Aopks-R '5- CTGGCGTTCCAGTACCATGAG-'373. The 630 bp fragments were amplified, PCR was performed in a reaction volume of 25 μl according to Hussein et al.74 The reactions were done in a C1000.
Thermo Cycler BioRad, Germany with initial denaturizing at 94 °C for 5 min, followed by 30 cycles of 1 min. at 94 °C, 1 min. at 58 °C and extension at 72 °C for 1 min43, then final step as extension at 72 °C for 10 min. PCR products were checked on a 1.3% agarose gel and stained with ethidium bromide.
Screening A. niger isolates for lipase production
Tween 80 agar plate was used for screening the tested isolates for lipase production containing (g/l peptone, 15; NaCl, 5; CaCl2, 1; tween 80, 10 and agar, 15) and pH of the media was adjusted to 7. 250 µl of fungal spore suspension (8 × 107 spores/ ml) was inoculated to 8 mm cavity on the media and incubated at 28 °C for 4 days. Appearance of white precipitate around the fungal colony indicates the ability to produce lipase enzyme49.
Quantitative estimation of lipase
Trimetric method was applied for assay lipase75 with some modification. Two disks (8 mm) of tested isolates were inoculated to minimal medium containing (g/l 1 yeast extract, 1 KCl, 1 MgSO4.7H2O), pH 6 and supplemented with (1% v/v tween 80) and incubated at 30 °C on shaker incubator at 150 rpm for 3 days. 1.5 ml suspension from liquid medium was centrifuged at 3000 rpm for 10 min. Assay mixture containing 500 µl 0.1 M phosphate buffer (pH 6.8), 500 µl tween 80 and 250 µl crude enzyme was incubated at 37 °C for 20 min. Three ml of acetone: ethanol (1:1) was added to stop the reaction. The liberated fatty acids were titrated with 1 N NaOH solution with bromothymol blue indicator. In control 250 µl distilled water was added instead of crude enzyme. Lipase activity (U/ml-min) was calculated from the following equation:
T is the titration volume, C is the control, N is the normality of NaOH, df is the volume of assay/volume of enzyme, t is the incubation time, v is the sample.
Detection of lip2 gene in Aspergillus niger isolates
Aspergillus niger lip2 gene was detected by using 2 documented primers. The sequences of primers were as following: P1 (5’-CTCAAGAGTATCCTGCACTG-3’) and P2 (5’-CTGAACCTTCCTTGGGATAG-3’)55. Twenty-five μl as volume was used for PCR reaction by mixing 12.5 µl of EmeraldAmp Max PCR Master Mix (Takara, Japan), 1 µl of each primer, 6 µl of DNA template and 4.5 µl of water was added to make the volume up to 25 μl. Applied biosystem 2720 thermal cycler was used for performing the reaction with initial denaturizing at 94 °C for 5 min, followed by 35 cycles of 30 s. at 94 °C, 50 s. as annealing temperature at 59 °C and 1 s. at 72 °C. Ten minutes at 72 °C was used as the final extension. The PCR products were checked on 1.5% agarose gel in 1 × TBE buffer. A gelpilot 100 bp plus DNA Ladder was used to determine the fragment sizes. The gel was photographed by a gel documentation system. Data was analyzed through computer software.
Studying the virulence properties of target human pathogens in presence of crude lipase obtained from A. niger
Determination of minimum inhibitory concentration (MIC)
MIC was evaluated by p-iodonitrotetrazolium violet choloride (INT) formazon assay (0.2 mg/ml, SIGMA-ALDRICH). Overnight cultures of Escherichia coli, Pseudomonas aeruginosa, Proteus mirabilis, and Methicillin-resistant Staphylococcus aureus (MRSA) were adjusted to OD595 of 0.01 into tryptic soy broth (TSB). 100 µl of each freshly prepared bacterial culture were placed into 96-well plates plus different volumes of crude lipase (10–100 µl, 8 replicates were made for each volume in complete raw). After 24 h incubation at 37 °C, to confirm bacterial growth suppression and deficiency of metabolic activity, 40 µl INT was added to the microplate wells and re-incubated at 37 °C for 30 min. The MIC in the INT assay was defined as the lowest concentration that suppressed bacterial growth and prevented color change62,76,77,78.
Determination of minimum bactericidal concentration (MBC)
The bactericidal efficacy was defined as a 99.9% decrease in CFU (3 logs) in the initial inoculum during 24 h of incubation. The MBC was determined by inoculating sterilized tryptic soy agar (TSA) fresh plates with 50 µl from each well of overnight MIC plates. Viable colonies were counted after 24 h at 37 °C. The limit of detection for this assay was 10 cfu ml−162,79.
Static biofilm assay
The tested bacterial strains {Escherichia coli, Pseudomonas aeruginosa, Proteus mirabilis and Methicillin-resistant Staphylococcus aureus (MRSA)} were obtained kindly from international Luxor hospital. The ability of the verified pathogens for biofilm formation was determined using 96-well polystyrene plates80. Bacterial strains were subcultured on tryptic soy agar for 24 h at 37 °C, suspended in tryptic soy broth and adjusted to an OD595 of 0.02. 130 µl of each adjusted isolate culture were put in the microtitre plate (U bottom, Sterilin) at 37 °C for 24 h. After incubation the wells were washed six times with distilled water, Furthermore, the wells were stained with 0.1% crystal violet for 10 min. the wells were again washed with distilled water (4 times) to remove excess stain81. Finally, the wells were destained by 210 µl of ethanol 96% and the OD595 was read using infiniteF50 Robotic (Ostrich) microplate plate to quantify the amount of biofilm.
Antibiofilm efficacy of crude lipase
The effect of crude lipase enzyme with the highest activity from A. niger MW029470 free of ochratoxin after 3 days of incubation as antibiofilm against four human pathogenic biofilm former bacteria was done by spectrophotometric methods. Different volumes (20, 30, 40, 50, 60, and 70 µl) were added to 130 µl of the tested pathogens at OD595 of 0.02 after 24 h incubation at 37 °C for allowing biofilm formation. The plates then incubated for further 24 h and then stained with crystal violet as described previously27.
Scanning electron microscopy (SEM) analysis of antibacterial and antibiofilm efficacy of crude lipase
Preparation of samples for antibacterial was performed as described by Wang et al.82. While for biofilm was done as described by Kong et al.83 and Chin et al.84 with little modifications. Biofilms were allowed to form on the slides at 37 °C for 24 h alone (control) and after treatment with crude lipase following which, the samples were fixed in 4% (v/v) glutaraldehyde in 0.05 M phosphate buffer (pH 7.0) at 4 °C for 12 h. Subsequently, the samples were washed three times in phosphate buffer, dehydrated through a graded ethanol series, dried in a critical-point drying apparatus with liquid carbon dioxide; slides coated with gold and viewed using (JEOL JSM-5500LV, Japan).
Statistical analysis
The variability degree of results was expressed in form of means ± standard deviation (mean ± SD) based on triplicates determinations (n = 3 for replicate plates). The data were statistically analyzed by one-way ANOVA analysis and compared using the least significant difference (LSD) test at 0.05 (*) levels. It was done to compare between control and treatments.
References
Carughi, A. et al. Pairing nuts and dried fruit for cardiometabolic health. Nutr. J. 5, 15–23. https://doi.org/10.1186/s12937-016-0142-4 (2015).
Li, M., Fan, Y., Zhang, X., Hou, W. & Tang, Z. Fruit and vegetable intake and risk of type 2 diabetes mellitus: Meta-analysis of prospective cohort studies. BMJ Open 4, e005497. https://doi.org/10.1136/bmjopen-2014-005497 (2014).
Hemandez-Alonso, P., Salas-Salvado, J., Baldrich-Mora, M., Juanola-Falgarona, M. & Bullo, M. Beneficial effect of pistachio consumption on glucose metabolism, insulin resistance, inflammation, and related metabolic risk markers: A randomized clinical trial. Diabetes Care 37, 3098–3105. https://doi.org/10.2337/dc14-1431 (2014) (Epub 2014 Aug 14).
Weidenborner, M. Pine nuts: The mycobiota and potential mycotoxins. Can. J. Microbiol. 47, 460–463 (2001).
Tournas, V. H., Niazi, N. S. & Kohn, J. S. Fungal presence in selected tree nuts and dried fruits. Microbiol. Insights. 8, 1–6. https://doi.org/10.4137/MBI.S24308 (2015).
Agriopoulou, S., Koliadima, A., Karaiskakis, G. & Kapolos, J. Kinetic study of aflatoxins degradation in the presence of ozone. Food Control 61, 221–226. https://doi.org/10.1016/j.foodcont.2015.09.013 (2016).
Benedict, K., Chiller, T. M. & Mody, R. K. Invasive fungal infections acquired from contaminated food or nutritional supplements. Foodborne Path. Dis. 13(7), 343–349. https://doi.org/10.1089/fpd.2015.2108 (2016).
Pfohl-Leszkowicz, A. Ochratoxin A and aristolochic acid involvement in nephropathies and associated urothelial tract tumours. Arh. Hig. Rada. Toksikol. 60(4), 465–483. https://doi.org/10.2478/10004-1254-60-2009-2000 (2009).
Wafa, E. W., Yahya, R. S., Sobh, M. A., Eraky, I., El Baz, H., El Gayar, H. A. M., Betbeder, A. M. & Creppy, E. E. Human ochratoxicosis and nephropathy in Egypt: A preliminary study. Hum. Exp. Toxicol. 17, 124–129 (1998).
Schwartz, G. G. Hypothesis: Does ochratoxin A cause testicular cancer?. Cancer Causes Control. 13, 91–100. https://doi.org/10.1023/A:1013973715289 (2002).
Singh, A. K. & Mukhopadhyay, M. Overview of fungal lipase: A review. Appl. Biochem. Biotechnol. 166(2), 486–520. https://doi.org/10.1007/s12010-011-9444-3 (2012).
Putri, D. N., Khootama, A., Perdani, M. S., Utami, T. S. & Hermansyah, H. Optamization of Aspergillus niger lipase production by solid state fermentation of agro-industrial waste. Energy Rep. 6, 331–335 https://doi.org/10.1016/j.egyr.2019.08.064 (2020).
Hasan, F., Shah, A. A. & Hameed, A. Industrial applications of microbial lipases. Enzyme Microb. Technol. 39(2), 235–251 (2006).
Kristensen, J.B., Meyer, R.L., Laursen, B.S., Shipovskov, S., Besenbacher, F. & et al. Antifouling enzymes and the biochemistry of marine settlement. Biotechnol. Adv. 26, 471–481 (2008).
Carvajal, J.C., McDaniel, C.S. & Wales, M.E. Enzymatic antimicrobial and antifouling coating and polymeric materials. In US Patent/0238811 A1 (2009).
Prabhawathi, V., Boobalan, T., Sivakumar, P. M. & Doble, M. Antibiofilm properties of interfacially active lipase immobilized porous polycaprolactam prepared by LB technique. PLoS ONE 9(5), e96152. https://doi.org/10.1371/journal.pone.0096152 (2014).
Achchi, N. I. B., Khan, F. & Kim, Y. M. Inhibition of virulence factors and biofilm formation of Acinetobacterbaumannii by naturally-derived and synthetic drugs. Curr. Drug Targets. https://doi.org/10.2174/1389450121666201023122355 (2020).
Khan, F., Tabassum, N., Pham, D. T. N., Oloketuyi, S. F. & Kim, Y. M. Molecules involved in motility regulation in Escherichiacoli cells: A review. Biofouling 36(8), 889–908 (2020).
Khan, F., Khan, M. M. & Kim, Y. M. Recent progress and future perspectives of antibiofilm drugs immobilized on nanomaterials. Curr. Pharm. Biotechnol. 19(8), 631–643. https://doi.org/10.2174/1389201019666180828090052 (2018).
Khan, F., Oloketuyi, S. F. & Kim, Y. M. Diversity of bacteria and bacterial products as antibiofilm and antiquorum sensing drugs against pathogenic bacteria. Curr. Drug Targets 20(11), 1156–1179. https://doi.org/10.2174/1389450120666190423161249 (2019).
Khan, F. et al. Chitosan and their derivatives: Antibiofilm drugs against pathogenic bacteria. Colloids Surf. B Biointerfaces 185, 110627. https://doi.org/10.1016/j.colsurfb.2019.110627 (2020).
Mulat, M., Pandita, A. & Khan, F. Medicinal plant compounds for combating the multi-drug resistant pathogenic bacteria: A review. Curr. Pharm. Biotechnol. 20(3), 183–196. https://doi.org/10.2174/1872210513666190308133429 (2019).
Khan, F., Lee, J. W., Javaid, A., Park, S. K. & Kim, Y. M. Inhibition of biofilm and virulence properties of Pseudomonas aeruginosa by sub-inhibitory concentrations of aminoglycosides. Microb. Pathog. 146, 104249. https://doi.org/10.1016/j.micpath.2020.104249 (2020) (Epub 2020 May 11).
Khan, F., Pham, D. T. N., Oloketuyi, S. F. & Kim, Y. M. Regulation and controlling the motility properties of Pseudomonasaeruginosa. Appl. Microbial. Biotechnol. 104(1), 33–49. https://doi.org/10.1007/s00253-019-10201-w (2020).
Khan, F., Tabassum, N., Anand, R. & Kim, Y. M. Motility of Vibrio spp.: Regulation and controlling strategies. Appl. Microbiol. Biotechnol. 1–22 https://doi.org/10.1007/s00253-020-10794-7 (2020).
Xavier, J. B., Picioreanu, C., Rani, S. A., Van Loosdrecht, M. C. M. & Stewart, P. S. Biofilm control strategies based on enzymatic disruption of the extracellular polymeric substance matrix_a modeling study. Microbiology 51, 3817–3832 (2005).
Elamary, R., & Salem, W.M. Optimizing and purifying extracellular amylase from soil bacteria to inhibit clinical biofilm-forming bacteria. PeerJ 8, e10288 https://doi.org/10.7717/peerj.10288 (2020).
Ismail, M. A. Deterioration and spoilage of peanuts and desiccated coconuts from two sub-Saharan tropical east african countries due to the associated mycobiota and their degradative enzymes. Mycopathologia 150(2), 67–84. https://doi.org/10.1023/a:1010863507652 (2001).
Khosravi, A. R., Shokri, H. & Ziglari, T. Evaluation of fungal flora in some important nut products (pistachio, peanut, hazelnut and almond) in Tehran, Iran. Pak. J. Nutr. 6(5), 460–462 (2007).
Alhussaini, M. S. Mycobiota and mycotoxins of nuts and some dried fruits from Saudi Arabia. J. Am. Sci. 8 (12), 525–534, http://www.jofamericanscience.org (2012).
Kazemi, A. et al. Mold contamination of untreated and roasted with salt nuts (walnuts, peanuts and pistachios) sold at markets of Tabriz, Iran. Jundishapur. J. Microbiol. 7(1), e8751. https://doi.org/10.5812/jjm.8751 (2014).
Abbas, M., Naz, S. A., Shafigue, M., Jabeen, N. & Abbas, S. Fungal contamination in dried fruits and nuts: A possible source of mycosis and mycotoxicosis. Pak. J. Bot. 51(4), 1523–1529, https://doi.org/10.30848/PJB2019-4(31) (2019).
Zohri, A. A. & Abdel-Gawad, K. M. Survey of mycoflora and mycotoxins of some dried fruits in Egypt. J. Basic Microbiol. 33(4), 279–288. https://doi.org/10.1002/jobm.3620330413 (1993).
Sultan, Y. & Magan, N. Mycotoxigenic fungi in peanuts from different geographic regions of Egypt. Mycotoxin Res. 26, 133–140 (2010).
Abdulla, N. Q. F. Evaluation of fungal flora and mycotoxin in some important nut products in Erbil local markets. Res. J. Environ. Earth Sci. 5(6), 330–336 (2013).
Perrone, G., Susca, A., Epifani, F. & Mule, G. AFLP characterization of Southern Europe population of Aspergillus section Nigri from grapes. Int. J. Food Microbiol. 111(Suppl 1), S22–S27. https://doi.org/10.1016/j.ijfoodmicro.2006.03.009 (2006).
Botton, A. et al. A cDNA-AFLP approach to study ochratoxin A production in Aspergilluscarbonarius. Int. J. Food Microbiol. 127, 105–115. https://doi.org/10.1016/j.ijfoodmicro.2008.06.037 (2008).
Al-Sheikh, H. M. LAMP-PCR detection of ochratoxigenic Aspergillus species collected from peanut kernel. Genet. Mol. Res. (GMR) 14(1), 634–644, https://doi.org/10.4238/2015.January.30.5 (2015).
Magonli, C., Astoreca, A., Posone, M.L., Fernandez-Juri, M.G., Barberis, C. & Dalcero, A.M. Ochratoxin A and Aspergillus section Nigri in peanut seeds at different months of storage in Cordoba, Argentina. Int. J. Food Microbiol. 1, 119(3), 213–8, https://doi.org/10.1016/j.ijfoodmicro.2007.07.056 (2007).
Yassein, A. S., El-Said, A. H. M. & El-Dawy, E. G. A. Biocontrol of toxigenic Aspergillus strains isolated from baby foods by essential oils. Flavour Fragr. J. 35, 182–189. https://doi.org/10.1002/ffj.3551 (2020).
Palumbo, J. D. & O’Keeffe, T. L. Distribution and mycotoxigenic potential of Aspergillus section Nigri species in naturally contaminated almonds. J. Food Prot. 76(4), 702–706. https://doi.org/10.4315/0362-028X.JFP-12-431 (2013).
Massi, F. P., Sartori, D., Ferranti, L. deS., Imanaka, B. T., Taniwak, M. H. Vieira, M. L. C. & Fungaro, M. H. P. Prospecting for the incidence of genes involved in ochratoxin and fumonisin biosynthesis in Brazilian strains of Aspergillus niger and Aspergillus welwitschiae. Int. J. Food Microbiol. 16(221), 19–28, https://doi.org/10.1016/j.ijfoodmicro.2016.01.010 (2016).
El‐Hamaky, A. M., Atef, A. H., El Yazeed, H. A. & Refai, M. K. Prevalence and detection of toxigenic A. flavus, A. niger and A. ochraceus by traditional and molecular biology methods in feeds. Int. J. Curr. Res. 8, 25621–25633 (2016).
El-Dawy, E. G. A., Yassein, A. S. & El-Said, A. H. M. Detection of mycobiota, aflatoxigenic and ochratoxigenic genes, and cytotoxic ability in spices. Food Sci. Nutr. 7(8), 2595–2604. https://doi.org/10.1002/fsn3.1113 (2019).
Hussein, M. A. & Gherbawy, Y. Genotypic identification of ochratoxigenic Aspergilli that contaminated beef luncheon and their protease activity. Rend. Lincei-Sci. Fis. 30(4), 767–773. https://doi.org/10.1007/s12210-019-00845-1 (2019).
Macris, B., Kourentzi, E. & Hatzinkolaou, D. G. Studies on localization and regulation of lipase production by Aspergillusniger. Process Biochem. 31, 807–812 (1996).
Mala, J. G., Edwinoliver, N. G., Kamini, N. R. & Puvanakrishnan, R. Mixed substrate solid state fermentation for production and extraction of lipase from Aspergillusniger MTCC 2594. J. Gen. Appl. Microbiol. 53(4), 247–253. https://doi.org/10.2323/jgam.53.247 (2007).
Saleem, A. Effect of some food preservatives on lipolytic activity of beef luncheon fungi. Mycobiology. 36, 167–172 (2008).
Rai, B., Shreshtha, A., Sharma, S. & Joshi, J. Screening, optimization and process scale up for pilot scale production of lipase by Aspergillus niger. Biomed. Biotechnol. 2(3), 54–59, 10.12691bb-2-33 (2014). https: //pubs.scieup.com/bb-2-3-3.
Falony, G., Armas, J. C., Mendoza, J. C. D. & Hernandez, J. L. M. Production of extracellular lipase from Aspergillusniger by solid state fermentation. Food Technol. Biotechnol. 44(2), 235–240 (2006).
Mahadik, N. D., Puntambekar, U. S., Bastawde, K. B., Khire, J. M. & Gokhale, D. V. Production of acidic lipase by Aspergillusniger in solid state fermentation. Proc. Biochem. 38(5), 715–721 (2002).
Olama, Z. A. & El-Sabaeny, A. H. Lipase production by Aspergillusniger under various growth conditions using a solid state fermentation. Microbiologia. (Madrid) 9, 134–141 (1993).
Kamini, N. R., Mala, J. G. S. & Puvanakrishnan, P. Lipase production from Aspergillusniger by solid state fermentation using gingelly oil cake. Process. Biochem. 35, 501–511 (1999).
Mukhtar, H., Hanif, M., Ur-Rehman, A., Nawaz, A. & Ul-Haq, I. Studies on lipase production by Aspergillus niger through solid state fermentation. Pak. J. Bot. 47(SI), 351–354 (2015).
Yang, J., Sun, J. & Yan, Y. lip2, a novel lipase gene cloned from Aspergillusniger exhibits enzymatic characteristics distinct from its previously identified family member. Biotechnol. Lett. 32, 951–956. https://doi.org/10.1007/s10529-010-0238-4 (2010).
Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2, 114–122 (2003).
Pruteanu, M., Hernández Lobato, J. I., Stach, T. & Hengge, R. Common plant flavonoids prevent the assembly of amyloid curli fibres and can interfere with bacterial biofilm formation. Environ. Microbiol. https://doi.org/10.1111/1462-2920.15216 (2020).
Flemming, H. C. & Wuertz, S. Bacteria and archaea on Earth and their abundance in biofilms. Nat. Rev. Microbiol. 17, 247–260 (2019).
Flemming, H. C. & Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 8, 623–633 (2010).
Kalpana, B. J., Aarthy, S. & Pandian, S. K. Antibiofilm activity of α-amylase from Bacillussubtilis S8–18 against biofilm forming human bacterial pathogens. Appl. Biochem. Biotechnol. 167, 1778–1794. https://doi.org/10.1007/s12010-011-9526-2 (2012).
Lequette, Y., Boelsb, G., Clarissea, M. & Faille, C. Using enzymes to remove biofilms of bacterial isolates sampled in the food-industry. Biofouling 26, 421–431. https://doi.org/10.1080/08927011003699535 (2010).
Elamary, R. B., Albarakaty, F. M. & Salem, W. M. Efficacy of Acacia nilotica aqueous extract in treating biofilm-forming and multidrug resistant uropathogens isolated from patients with UTI syndrome. Sci. Rep. 10(1), 1–14. https://doi.org/10.1038/s41598-020-67732-w (2020).
Silva, N. L., Zimmer, K. R., Macedo, A. J. & Trentin, S. D. Plant natural products targeting bacterial virulence factors. Chem. Rev. 116(16), 9162–9236. https://doi.org/10.1021/acs.chemrev.6b00184 (2016).
Rosenberg, M., Gutnick, D. & Rosenberg, E. Adherence of bacteria to hydrocarbons: A simple method for measuring cell-surface hydrophobicity. FEMS Microbiol. Lett. 9, 29–33 (1980).
Reifsteck, F., Wee, S. & Wilkinson, B. J. Hydrophobicity-hydrophilicity of staphylococci. J. Med. Microbiol. 24, 65–73 (1987).
Whittaker, C., Ridgway, H. & Olson, B. H. Evaluation of cleaning strategies for removal of biofilms from reverse-osmosis membranes. Appl. Environ. Microbiol. 48, 395–403 (1984).
Vickery, K., Pajkos, A. & Cossart, Y. Removal of biofilm from endoscopes: Evaluation of detergent efficiency. Am. J. Infect. Control 32, 170–176. https://doi.org/10.1016/j.ajic.2003.10.009 (2004).
Vyas, N., Sammons, R. L., Addison, O., Dehghani, H. & Walmsley, A. D. A quantitative method to measure biofilm removal efficiency from complex biomaterial surfaces using SEM and image analysis. Sci. Rep. 6, 32694. https://doi.org/10.1038/srep32694 (2016).
Domsch, K. H., Gams, W. & Anderson, T. H. Compendium of Soil Fungi, Taxonomically Revised by W. Gams. 672 (IHW, 2007).
Hermosa, M. R. et al. Genetic diversity shown in Trichoderma biocontrol isolates. Mycol. Res. 108, 897–906. https://doi.org/10.1017/s0953756204000358 (2004).
Gabal, M. A., Hegazy, S. M. & Nagwa, Y. H. Aflatoxin production by Aspergillusflavus field isolates. Vet. Hum. Toxicol. 39, 519–521 (1994).
Lewis, L. et al. Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in eastern and central Kenya. Environ. Health Perspect. 113(12), 1763–1767. https://doi.org/10.1289/ehp.7998 (2005).
Reddy, K. V., Naveen, K. & Reddy, I. B. Incidence and molecular detection of ochratoxigenic fungi from Indian cereal grains. Int. J. Pharm. Biol. Sci. 4(3), 31–40 (2013).
Hussein, M. A., El-Said, A. H. M. & Yassein, A. S. Mycobiota associated with strawberry fruits, their mycotoxin potential and pectinase activity. Mycology 11(2), 158–166, https://doi.org/10.1080/21501203.2020.1759719 (2020).
Elegado, F., Legaspi, C. L., Paet, J. M., Querlibin, F., Tolentino, J. E., Vilela, J., Jr, A. P., Maloles, J. & Zarate, J. Screening, identification and optimization of extracellular lipase production of yeast (Cryptococcus flavescens) isolated from a tree canopy fern in the mount Makiling forest reserve, Philippines. In AIP Conference Proceedings Vol. 2155 (1), 020029, https://doi.org/https://doi.org/10.1063/1.5125533 (AIP Publishing LLC, 2019).
Eloff, J. N. A sensitive and quick microplate method to determine the minimal inhibitory concentration of plant extracts for bacteria. Planta. Med. 46, 711–713 (1998).
Lall, N., Henley-Smith, C.J., De Canha, M.N., Oosthuizen, C.B. & Berrington, D. Viability reagent, presto blue, in comparison with other available reagents, utilized in cytotoxicity and antimicrobial assays. Int. J. Microbiol. (2013).
Salem, W. M., El-Hamed, D. S., Sayed, W. & Elamary, R. Alterations in virulence and antibiotic resistant genes of multidrug-resistant Salmonellaserovars isolated from poultry: The bactericidal efficacy of Alliumsativum. Microb. Pathog. 108, 91–100. https://doi.org/10.1016/j.micpath.2017.05.008 (2017).
Sirelkhatim, A. et al. Review on zinc oxide nanoparticles: Antibacterial activity and toxicity mechanism. Nano-Micro. Lett. 7(3), 219–242 (2015).
Khan, F. et al. Streptomycin mediated biofilm inhibition and suppression of virulence properties in Pseudomonasaeruginosa PAO1. Appl. Microbiol. Biotechnol. 104(2), 799–816. https://doi.org/10.1007/s00253-019-10190-w (2020) ((epub 2019 Dec 9)).
Merritt, D.J., Turner, S.R., Commander, L.E. & Dixon, K.W. (eds). Proceedings of the Fifth Australian Workshop on Native Seed Biology Brisbane, Australia (2005).
Wang, J., Ma, M., Yang, J., Chen, L., Yu, P., Wang, J. & Zeng, Z. In vitro antibacterial activity and mechanism of monocaprylin against Escherichia coli and Staphylococcus aureus. J. Food Prot. 81(12), 1988–1996 (2018).
Kong, C. et al. Suppression of Staphylococcusaureus biofilm formation and virulence by a benzimidazole derivative, UM-C162. Sci. Rep. 8(1), 1–16 (2018).
Chin, C. Y., Hara, Y., Ghazali, A. K., Yap, S. J., Kong, C., Wong, Y. C., & Nathan, S. Global transcriptional analysis of Burkholderia pseudomallei high and low biofilm producers reveals insights into biofilm production and virulence. BMC Genomics 16(1), 471(2015).
Acknowledgements
Special thanks to South Valley University, Qena, Egypt. The authors also, extended their appreciation to Taif University, Saudi Arabia for funding this work through the program of Taif University Researchers Supporting Project number (TURSP-2020/59), Taif University, Taif, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
A.Y. conceived, designed the manuscript, performed the practical work and wrote some parts of the article. R.E. shared in the manuscript design, performed practical work, wrote some parts of the article, analyzed the data. M.H. assisted in the molecular identification part and gave some feedbacks about the research. All the authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yassein, A.S., Hassan, M.M. & Elamary, R.B. Prevalence of lipase producer Aspergillus niger in nuts and anti-biofilm efficacy of its crude lipase against some human pathogenic bacteria. Sci Rep 11, 7981 (2021). https://doi.org/10.1038/s41598-021-87079-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-87079-0
This article is cited by
-
Biogenesis, characterization, and applications of Spirulina selenium nanoparticles
Microbial Cell Factories (2025)
-
Enhanced production of extracellular L-asparaginase in batch culture via nitrous acid-induced mutagenesis of Aspergillus oryzae
Microbial Cell Factories (2025)
-
Optimizing lipase production by Bacillus subtilis on cheese whey and evaluating its antimicrobial, antibiofilm, anti virulence and biosafety properties
Scientific Reports (2025)
-
Antimicrobial, antibiofilm and antioxidant activities of bioactive secondary metabolites of marine Scarus ghobban gut-associated Aspergillus niger: In-vitro and in-silico studies
Scientific Reports (2025)
-
Extracellular matrix-degrading enzymes as a biofilm control strategy for food-related microorganisms
Food Science and Biotechnology (2023)
