
Abstract
Xanthoparmelia conspersa is rich in specific secondary metabolites but an unexplored lichen species. This work determined the chemical composition and biological activities (anti-microbial, anti-protozoal, and cytotoxic) of its methanolic and hexane extracts. Additionally, we evaluated the potential of these extracts in modulating cancer signaling pathways in HeLa cells. The phytochemical analysis revealed that usnic acid was the predominant constituent in the hexane extract, while stictic acid was in the methanolic one. Among tested cell lines (VERO, FaDu, SCC-25, HeLa), cytotoxic selectivity was detected for X. conspersa hexane extract against the FaDu (SI 7.36) and HeLa (SI 2.19) cells. A noticeably better anti-microbial potential was found for hexane extract, however, the overall anti-microbial activity was relatively weak (28, 21, and 20% inhibition of Candida glabrata, Cryptococcus neoformans, and Escherichia coli, respectively). On the contrary, the anti-parasitic action of hexane extract was significant, with an IC50 value of 2.64 µg/mL against Leishmania donovani – amastigote and 7.18 µg/mL against Trypanosoma brucei. The detailed evaluation of the cancer-related signaling pathways in HeLa cells, done by two distinct methodologies (luciferase reporter tests), revealed that especially the hexane extract and usnic acid exhibited selective inhibition of Stat3, Smad, NF-κB, cMYC, and Notch pathways.
Similar content being viewed by others
Introduction
Nature is a rich source of diverse chemical compounds, offering an incredible array of scaffold structures and complex molecular architectures. These natural products have played a pivotal role in the advancement of pharmacotherapy, particularly in the treatment of cancer and infectious diseases1. Botanical extracts and their isolated active compounds have been shown to effectively target specific pathways and enzymes in various cell types and disease models2,3,4. Despite significant progress, the need to explore natural sources for active compounds remains unmet. Additionally, the emerging concept of polypharmacology, which involves targeting multiple physiological pathways with complex mixtures, is gaining increasing popularity5. Lichen species have long been recognized for their high potential as sources of natural bioactive compounds. With the adventage of modern technologies, we can now uncover and explore the hidden pharmacological properties of these complex compounds, and explore their significant health benefits.
Lichens (Fungi lichenisati) are self-supporting, stable, mutualistic symbionts composed of fungi (mainly Ascomycota) and algae (Chlorophyceae) or cyanobacteria6. They produce characteristic secondary metabolites (aliphatic or aromatic) unique to lichen-forming fungi. These chemical compounds accumulate in the lichen cortex, for example, atranorin or usnic acid, or in the medullary layer, such as protocetraric acid, on the outer surfaces of the hyphae6. The genus Xanthoparmelia belongs to the Family Parmeliaceae, order Lecanorales, and contains about 800 species, of which more than 400 have been described based on morphological and chemical characteristics7,8. Up to now, over 1050 secondary metabolites isolated from lichens have been shown to exert biological and therapeutic activities9,10. For the genus Xanthoparmelia, over 92 compounds of diverse chemical structures (phenolics, anthraquinones, depsides, depsidones, monocyclic compounds, and aliphatic acids) were described11. Some of these molecules have been reported to possess antibiotic12, antioxidant13,14, anti-proliferative15, anti-pyretic, anti-tumor16, anti-bacterial, photoprotective17, and even anti-viral (e.g., HIV)18,19 properties. They were also proved important factors in metal homeostasis and pollution tolerance of lichen thalli20.
Secondary metabolites isolated from X. conspersa include usnic acid, stictic, and norstictic acids21. Usnic acid showed relatively potent anti-microbial22,23 and anti-tumor effects24,25. It caused a decrease in the proliferation of human breast cancer and human lung cancer cells without DNA damage20 and exhibited anti-viral activity against the Junin virus26. Additionally, this dibenzofuran was found to have strong UV absorption abilities and function as a filter for excessive UV-B irradiation27. A depsidone, stictic acid, showed anti-microbial activity on Klebsiella pneumoniae28 and against two fungi species, Candida albicans and C. glabrata29,30.
Currently, the search for drugs targeting cancer is considered of tremendous importance. HeLa cells are one of the most frequently used cell lines in scientific research31,32. A complex network of signaling pathways in cancer cells and their key transcription factors regulate cytological processes, including differentiation, cell death, proliferation, oncogenic transformation, and apoptosis33,34. Of a large number of cancer-associated genes and driver gene mutations identified, the vast majority belong to 13 different signal-transduction pathways including such transcription factors as Stat3, Smad, AP-1, NF-kB, E2F, MYC, Ets, Notch, FoxO, Wnt, Hdghog, miR-21, and k-Ras35,36. These are signaling nodes for multiple oncogenic pathways, which transduce intracellular and extracellular signals to the nucleus and control the expression of genes responsible for multiple physiological processes such as cell growth, proliferation, differentiation, positioning, migration, metabolism, and apoptosis37,38.
Given the aforementioned details and our current limited knowledge of X. conspersa, the objective of this study was to determine the chemical composition and explore the biological activities (anti-microbial, anti-leishmanial, anti-trypanosomal, and cytotoxic) of the methanolic and hexane extracts of this species. In addition, the anti-cancer potential of these extracts in inhibiting cancer signaling pathways in HeLa, a cervical cancer cell line, was evaluated.
Results and discussion
Phytochemical analysis of Xanthoparmelia conspersa extracts
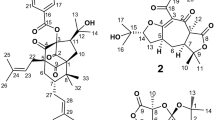
The secondary metabolites detected in the studied lichen species can be classified into four chemical classes: phenols, depsides, depsidones, and dibenzofurans (Table 1). Phenols were represented by lecanoric acid (1), atranol (2), methyl everninate (3), methyl 4-formyl-3,5-dihydroxybenzoate (4), 3-butyryl-β-resorcylic acid (5), proatranorin II (6) and dimethoxybenzoic acid derivatives (7, 8). Haemathamnolic acid isomers (9, 10), hypothamnolic acid (11), and thamnolic acid (12) constituted depsides. Depsidones included stictic (15), cryptostictic (14), norstictic (17), hypoconstictic (18), and constictic (19) acids, as well as two unidentified benzofuro-benzodioxepin-carboxylic structures (13, 16). Several molecules based on the dibenzofuran skeleton were found in the last category. These covered differently substituted: dibenzofuran-carboxylic acid, dibenzofuran-dicarboxylate, dibenzofurandione-based structures (20, 21, 24, 26, 28, 31, 33, 35, 36) and also usnic (29), isousnic (30), decarbousnic (22), placodiolic (25, 27) and pseudoplacodiolic (23) acids with their derivatives (32, 24). Compounds placed in the category of others were organic acids: gluconic acid (37), methylisocitric acid (38), and citric acid (39).
As can be seen on the DAD chromatogram (Fig. 1A), the predominant constituent in the hexane extract was usnic acid. It was accompanied by small amounts of placodiolic acid, other dibenzofurans and a few phenols (Table 1). Usnic acid was also detected in significant amounts in the methanolic extract. However, based on UV and mass spectra, the maximal signal on the methanolic extract chromatogram was assigned as stictic acid (Fig. 1B). From the qualitative point of view, the methanolic extract was abundant in numerous compounds belonging to all four chemical classes (phenols, depsides, depsidones, and dibenzofurans) (Table 1). However, besides stictic and usnic acids, other constituents had minor contributions to the methanolic extract profile. Our study’s results agree with the literature data, which indicated that usnic acid is the major constituent of the upper cortex of X. conspersa thallus. In contrast, stictic acid is predominant in the medulla39. Additionally, constictic, cryptostictic and norstictic acids are all minor constituents39, as we detected.
The fragmentation pathway of usnic acid (33)(C18H16O7, m/z 343.0811) could be carried out in three directions. The first possibility was the formation of the ion at m/z 299 by the loss of CO2. The second was the elimination of methyl substituent, resulting in the formation of radical at m/z 328. The last variant revealed the loss of the C4H4O2 fragment, which produced the daughter ion at m/z 259, which, by subsequent elimination of CO, yielded an ion at m/z 231 (Fig. 2). Similar fragmentation patterns were observed for placodiolic acid and other dibenzofurans, which presented a loss of CH3OH, CO, CH3, CO2, or H2O40,41,42 (Table 1). The pseudomolecular ion of stictic acid (17) (C19H13O9, m/z 385) yielded a daughter ion at m/z 341, obtained by the loss of CO2 from the parent ion. The subsequent loss of CO2 and CH3 produced radicals at m/z 282, which was further diminished by another methyl group, resulting in an ion at m/z 267. Likewise, cryptostictic, norstictic, hypoconstictic, and constictic acids showed the fragmentation with a loss of two CO2 moieties [M-H-44-44]−40,43.
As described above, LC-MS analysis allowed us to characterize secondary metabolites present in the investigated extracts of X. conspersa. The qualitative determination revealed numerous compounds of different chemical structures, the UV spectra proved that usnic and stictic acid dominate quantitatively and most probably significantly contribute to the observed biological activities of the extracts.

The DAD (λ = 320 nm) chromatograms of the studied X. conspersa extracts. (A) – hexane extract, (B) – methanolic extract. UV spectra are shown for the main constituents of the extracts.

The fragmentation pathway of usnic acid.
Anti-microbial activity
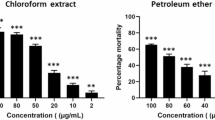
The results of anti-bacterial and anti-fungal activity of X. conspersa methanolic and hexane extracts, as well as the positive controls (anti-bacterial fluoroquinolone ciprofloxacin; antifungal amphotericin B) are shown in Table 2. Crude extracts were initially tested in a primary screen at 50 µg/mL in duplicate, and percent inhibitions (% inh.) were calculated relative to negative and positive controls. Extracts showing < 50% inhibition did not proceed to the secondary assay. The observed antimicrobial potential depended on the extract type and the microorganism species. Based on the results, it was found that X. conspersa hexane extract had better anti-bacterial activity compared to methanolic extract. Microbiological assays of the hexane extract showed activity against the fungal pathogens Candida glabrata and Cryptococcus neoformans, described by 28 and 21% inhibition, respectively (Table 2). C. glabrata is an important agent responsible for the surface (oral cavity, esophagus, vaginal or urinary duct) or systemic mycoses. The main problem with the infections caused by C. glabrata is their innate resistance to drugs that are very effective against other Candida species. Thus, finding agents effective against this species is of particular importance since, with the increasing use of immunosuppressants, the number of cases of candidiasis is increasing44. Amphotericin B, used as a positive control and used to treat C. glabrata infections, is nephrotoxic for humans and can trigger multiple undesired side effects. Also, C. neoformans is an important pathogen which causes cryptococcosis. It is a subacute or chronic mycosis that attacks the central nervous system, lungs (organ mycoses, deep mycoses) or the skin and subcutaneous tissue (superficial mycoses)45. The antibacterial activity of the hexane extract of X. conspersa was mostly against Escherichia coli (20% inhibition). E. coli is a facultatively anaerobic Gram-negative rod belonging to the Enterobacteriaceae family46. Most E. coli strains are components of the flora of the human large intestine and rarely cause disease in healthy individuals. Nonetheless, many pathogenic strains can cause inflammation of the urinary tract, sepsis, meningitis, intestine diseases, and diarrhea. This species is difficult to treat as it is resistant to many known antimicrobial agents47.
Although the activity described above was relatively weak and the examined lichen extracts showed lower ability to inhibit the growth of the other fungi (Candida albicans, C. krusei, Aspergillus fumigatus), and other bacteria (Staphylococcus aureus, MRS, Pseudomonas aeruginosa, M. intracellular) pathogenic to humans (Table 2) it should be underlined that screened extracts concentration was relatively low. The literature sources report a wide range of concentrations in anti-microbial screening. Therefore, the strict value selected in this study (50 µg/mL) may be considered as a limiting factor. Nevertheless, some studies have shown that the depsidone - stictic acid isolated from X. conspersa thalli, had antimicrobial activity against Klebsiella pneumoniae28 and against two fungi species Candida albicans and C. glabrata30.
Anti-parasitic activity
Hexane extract of X. conspersa showed > 90% inhibition of growth against L. donovani – amastigotes, against intracellular L. donovani amastigotes in THP1 human acute monocytic leukemia cell line (L. donovani – amastigotes + THP1), and against trypomastigote forms of Trypanosoma brucei at 20 µg/mL (Table 3). The results are similar to the anti-leishmanial drugs used as control. Still, in the case of intracellular amastigotes with differentiated THP1 cells infected in vitro with L. donovani, hexane extract of X. conspersa showed higher inhibition (99%) than amphotericin B as control (81%). Because methanolic extract caused very little inhibition, only hexane extract was subjected to a secondary assay for dose-response analysis. The IC50 value was 1.99 µg/mL against human acute monocyclic leukemia cell line (THP1), 2.64 µg/mL against L. donovani - amastigote and 6.17 µg/mL against intracellular Leishmania donovani amastigotes in THP1 (Table 3). Assays with macrophage-amastigotes models are close to the pathophysiological conditions of leishmaniasis and appropriate for in vitro screening to discover new drugs48. Notably, the observed IC50 value of the extract and reference compound pentamidine against L. donovani – amastigotes and L. donovani – amastigotes + THP1 were very similar (Table 3), indicating the extract’s high potency.
The results also confirmed the high biological activity of the hexane extract of X. conspersa against Trypanosoma brucei (100% inhibition at the concentration of 20 µg/mL in primary assay). In the secondary screening, the IC50 value against T. brucei for active extract was 7.18 µg/mL and for the activity of the reference compound alpha-difluoromenthylornithine (DFMO) was similar (5.35 µg/mL) (Table 3).
Both investigated parasitic organisms still pose a threat to human health. L. donovani is flagellate (Leishmania) from the genus Trypanosomatidae and the obligate intracellular protozoan49. The parasite causes visceral leishmaniasis (Dum-Dum fever, black fever, kaza-azra)50. 12 million people suffer from leishmaniasis yearly, of which 1/4 cases are visceral leishmaniasis51. The reservoirs of infection are wild and domestic animals (e.g. dogs), although in the case of L. donovani, the vector typically transmits the pathogen directly from person to person. T. brucei is an extracellular protozoan parasite that causes African trypanosomiasis, transmitted by tsetse flies from the genus Glossina (G. morsitans, G. pallidipes, G. swynnertoni). It parasitizes the mammalian hemolymphatic and CNSs and causes persistent infections in humans (sleeping sickness) and domestic animals (nagana) that are fatal52,53.
The main compound of the studied X. conspersa hexane extract, usnic acid, showed an antiparasitic effect against intracellular amastigote forms, with disorganization of parasite kinetoplast and mitochondria, but with nosignificant ultrastructural damage to the host cells54. Moreover, usnic acid was effective as well against L. major, L. infantum, and L. tropica promastigotes at IC 50 values of 10.76 μg/ml, 13.34 μg/ml, and 21.06 μg/ml, respectively. It not only inhibited proliferation but also caused apoptosis through upregulation of p53, Bax, Casp-3, and Casp-9 geneexpression and downregulation of the level of Bcl-2 gene expression55. Additionally, usnic acid was already proven to dose-dependently (5–30 μg/ml) inhibit the growth of other T. cruzi species epimastigotes in vitro. Thetreatment resulted in damage to mitochondria with a marked increase in kinetoplast volume, enlargement of the flagellar pocket, and vacuolation of cytoplasm and the mitochondrial matrix.
Cytotoxicity and anti-proliferative selectivity
The cytotoxicity of X. conspersa was evaluated on non-cancerous VERO cells and cancer cell lines originating from Head and Neck Cancers (HNC) – FaDu and SCC-25, and cervical adenocarcinoma – HeLa, as was presented in Table 4; Fig. 3. X. conspersa extracts showed moderate to weak cytotoxicity towards VERO cells with CC50 (50% cytotoxic concentration) between 113.4 and 205.25 µg/mL (Table 4). Anti-proliferative selectivity was observed for X. conspersa hexane extract against the FaDu (SI 7.36) and HeLa (SI 2.19). Notably, the CC50 on FaDu cells was 15.41 µg/mL, which indicates high cytotoxic activity56,57. The X. conspersa methanolic extract also showed selective toxicity towards the FaDu (SI 2.95) and HeLa (SI 5.91). Usnic and stictic acids were included in the studies as the main components of hexane and methanolic extracts, respectively. These acids demonstrated greater activity against all studied cell lines compared to the extracts. This suggests that these compounds significantly contribute to the overall activity of the extracts. As shown in Fig. 1, usnic acid is the predominant compound in the hexane extract, while stictic acid is more abundant in the methanolic extract. Additionally, the significant presence of both acids in the methanolic extract suggests the possibility of synergistic activity. Atraric acid, representing a different phenolic structure, was used for comparative evaluation. Atraric acid showed significant (p < 0.02) selectivity toward HeLa, while the stictic acid against both cell lines belonging to HNC and usnic acid only on FaDu cells. In general, the SCC-25 cells were resistant to the tested samples, except the static acid, which showed potent cytotoxicity (CC50 9.01 µg/mL) and potential antiproliferative selectivity (SI 3.88) against these cells.
CC50 – 50% cytotoxic concentration (mean ± SD; µg/mL); SI – Selectivity Index (CC50VERO/ CC50Cancer); Statistical significance of higher cytotoxicity towards cancer-originating cell lines was evaluated regarding results obtained for non-cancerous VERO cells (2-way ANOVA; Dunnett’s multiple comparisons test); a highly significant (p < 0.01) anticancer cytotoxicity was labeled where applicable.
Table 4. Cytotoxicity of X. conspersa extracts and selected bioactive compounds.
Herein, the cytotoxicity profile of X. conspersa extracts towards non-cancer and selected cancer-originating cells was reported for the first time. Kocovic et al.58 reported the cytotoxicity of Xanthoparmelia stenophylla acetonic and hexane extracts on non-cancerous cells derived from human lung (MRC-5) and cancer cells – HeLa and HCT 116 (colon carcinoma). The acetonic extract showed moderate toxicity toward MRC-5 with CC50 of 51.63 µg/mL, but the exact CC50 of the hexane extract was not evaluated (CC50 > 100 µg/mL)58. The toxicity of usnic acid against MRC-5 (CC50 53.14 µg/mL) reported by58 was comparable to the one we observed on the VERO cells (CC50 49.14 µg/mL). Interestingly, we have observed higher cytotoxicity of usnic acid on HeLa cells (CC50 34.48 µg/mL) than was previously reported (CC50 > 100 µg/mL)58. However, the cytotoxicity of X. conspersa hexane extract (CC50 51.84 µg/mL) on HeLa was similar to the one reported for X. stenophylla hexane extract (CC50 51.37 µg/mL)58. It was also shown that Xanthoparmelia mexicana did not show significant cytotoxic activity against MCF-7 (breast adenocarcinoma) cells59, and Xanthoparmelia somloensis extract (MeOH/H2O, 50/50, %v/v) induced anticancer effect against MCF-7 and MDA-MB-231 (breast adenocarcinoma) with CC50 of 58.8 and 45.8 µg/mL, respectively. In contrast, no significant activity was found against cells originating from lung cancers - A549 and H129960.
Shrestha et al.61 studied the anticancer activities of extracts from several species of North American lichen against Raji cells (lymphoblast-like cells originating from Burkitt’s Lymphoma). From the 17 studied lichen species, 14 extracts showed cytotoxicity against Raji cells, and the highest toxicity was found for Xanthoparmelia chlorochroa and Tuckermannopsis ciliaris with CC50 of 28.3 and 28.4 µg/mL, respectively. Extracts from X. chlorochroa and T. ciliaris extracts did not affect the viability of normal lymphocytes. Further, it was found that both extracts not only decreased the proliferation of Raji cells but also caused a cell cycle arrest at the G0/G1 phase, induced apoptosis, and upregulated the p53 gene expression. The upregulation of TK1 expression was observed only for T. ciliaris61. The composition of X. chlorochroa extract shared some similarities with extracts from X. conspersa analyzed herein, both containing usnic, constictic, and norstictic acids as major compounds.

Dose-response influence of tested samples on cell lines.
The influence on the cancer-related signaling pathways in HeLa cells
As described above, the tested extracts and individual compounds were selectively toxic towards FaDu (SI 2.95) and/or HeLa cells, yet their detailed mechanism of action was not known. Therefore, in this work, we used a well-established HeLa cancerous cell line model to evaluate the influence of X. conspersa extracts on the signal transduction pathways important in cancer progression. To evaluate the modulating effect of these test agents, we performed screening tests using a battery of 13 Firefly luciferase (FFLuc) reporter gene vectors in the first step. The preliminary results showed that the hexane extract of X. conspersa was more potent than the methanolic one in inhibiting these activated pathways (Fig. 4). The hexane extracts potency of inhibition was as follows: MYC, Wnt, Notch and next Stat3, Smad, Hdghog, Ets, Ap-1, NF-κB, and k-Ras. pTK was used as the vector control. We observed dose-dependent activity in the concentration range of 15, 25, and 40 µg/mL (Fig. 5) and higher than the resveratrol analog, a reference compound used. In all tested concentrations, usnic acid also revealed higher activity than resveratrol analog and atraric acid. It inhibited the activation of 11 signaling pathways (except FoxO and miR-21), Wnt, MYC, and Hdghog exhibited strong inhibition at lower concentration (25 µM), while Notch, Ets, NF-κB, Stat3, Smad, Ap-1, E2F NF-κB and k-Ras at higher concentration (50µM). The inhibition was similar to the hexane extract, confirming that usnic acid significantly contributes to the biological activity of the extract. Interestingly, hexane extract, usnic acid, and resveratrol analog did not inhibit FFLuc expression driven by the minimal thymidine kinase promoter (pTK - control), indicating it was not significant for general cytotoxicity, changes of general gene expression, or luciferase enzyme inhibition. Similarly, the activation of the apoptotic mediators FoxO and miR-21 was not observed with the usnic acid in all tested concentrations (Table S1), as well as with hexane and methanolic extracts in the concentration of 40 µg/mL, and with hexane extract in concentrations of 25 µg/mL and 15 µg/mL.

The inhibition of activation of Ap-1, E2F, Ets, FoxO, Hdghog, k-Ras, miR-21, MYC, NF-κB, Notch, pTK, Smad, Stat3, Wnt signalling by hexane extract of X. conspersa and reference/control compounds: RA– resveratrol analog; XCM – the methanol extract of X. conspersa; XCH – the hexane extract of. X. conspersa; IL-6, TGF-β, PMA, Wnt 3a – Inducer (promoter) of cancer processes.

Dose-response activity of X. conspersa extracts (A), usnic and atraric acids (B) against cancer-related signaling pathways in HeLa cells.
Additionally, k-Ras as a transcription factor in FFLuc gene assay was weakly activated by hexane extract of X. conspersa in the concentration of 40 µg/mL and 25 µg/mL and by usinic acid in concentrations of 50µM and 25µM (Table S2). The phenolic analog compound, atraric acid exhibited lower potency against the tested signaling pathways than the active antitumor compound, resveratrol analog. This may suggest that other transduction molecules mediate its antiproliferative activity.
Since the results of the FFLuc screening assays revealed a significant influence of X. conspersa extracts and usnic acid on the cancer-related signaling pathway in HeLa cells, we continued the research by detailed determination of the selected pathways in the second step.
Further analysis conducted using Gaussia Luciferase Reporter System (GLRS) (Fig. 6), a secreted luciferase that was normalized by the secretion of SEAP (secreted alkaline phosphatase), showed that usnic acid and hexane extract of X. conspersa have a dose-response in all the reporters analyzed, whereas the highest inhibitory potential in small doses was for NFkβ, cMyc, NFAT, SMAD, STAT, K-Ras and Notch signaling pathways. Atraric acid showed inhibitory activity but only in high doses (> 40 µg/ml). The methanol extract of. X. conspersa in low doses activates TWIST1 and NFAT.
By using 2 reporter systems, we show that usnic and, to a lesser degree atraric, acids have inhibitory effects on various signaling pathways in a dose-dependent manner. Some of these pathways were inhibited at low doses, in particular by usnic acid. In addition, the inhibition profile of usnic acid closely resembled that of the hexane extract, further indicating that usnic acid is a major contributor to the extract’s biological activity.
Several of these pathways are well-known promoters of tumorigenesis, tumor aggressiveness, migration, or proliferation of multiple cancer cells35,36,38. Here, we show that extracts from X .conspersa, most notably usnic acid, interfere with these pathways and reduce proliferation and viability at relatively low doses, while having a reduced effect on non-cancer cells, as presented using VERO cells.
In our studies, among the tested signaling pathways, Myc, Wnt, and Hdghog showed strong inhibition by usnic acid when compared to the other pathways. We are the first to report this activity in HeLa cells. MYC (induced nuclear protein antigen) was activated by phorbol 12-myristate-13-acetate, a protein kinase activator (PMA) (Table S1, Fig. 4). MYC, as well as c-, L-, and N-myc belong to the proto-oncogene family and are central regulators of cell growth, which abnormal expression may be responsible for carcinogenesis62. MYC encoded by the MINA gene directly regulates target genes responsible for the proliferation and growth of cancer cells and shows elevated expression in up to 70% of all human malignancies63. Kumar et al.64 have reported that usnic acid suppresses the downstream target genes of c-jun/AP-1 and c-myc at mRNA levels.

The expanded dose-response activity of X. conspersa extracts usnic and atraric acids against cancer-related signaling pathways in HeLa cells (GLuc reporter system). Abbreviations used in the figure stand for A – atranic acid, U – usnic acid, XCH – hexene extract of X. conspersa, XCM – methanol extract of. X. conspersa. The results were shown as fold change calculated to the non-treated control and normalized against SEAP.
The Wnt signaling pathway was activated by the protein Wnt-3a, a protein encoded by the WNT3A gene in humans65. The WNT3A gene is clustered in human chromosome 1q42, where during carcinogenesis, recombination results in chromosomal translocation, gene amplification, and deletion66. The WNT family of genes consists of structurally related genes encoding the secretion of proteins in signaling pathways essential during development, cell differentiation, morphogenesis, and repair, but often dysregulated during tumor formation65. Some of the lichen secondary metabolites like squamatic acid, salazinic acid, and lecanoric acid were reported to inhibit the Wnt pathway in GBM cell line models67.
The next most inhibited pathway in our studies was Hdghog. The Hedgehog (Hh, synonyms: Hedgehog-Patched (Hh-Ptch), Hedgehog-Patched-Smoothened (Hh-Ptch-Smo)), is also an evolutionarily conserved pathway of signal transmission from the cell membrane to the nucleus, that plays critical roles in development, regeneration of the lung epithelium, prostate epithelium, and exocrine pancreas cells. It is involved too in the maintenance of somatic stem cells and pluripotent cells important to tissue repair68 and is often dysregulated in neoplasias69,70,71. Downstream of activation of the Patch/Smo receptors, Gli1/2 are activated and translocated to the nucleus to act as transcription factors to Hh-dependent genes. Usnic acid is reported to mediate anti-proliferative effects via miRNA-dependent mechanisms that target Hdghog, TGF- and MAPK apoptosis pathways in human breast cancer cell lines72.
The Notch signaling pathway was activated by a protein kinase activator (PMA), but can also be activated by a specific Serine/threonine (Ser/Thr) protein kinase73. Numerous studies support a pro-oncogenic function for Notch signaling during neurogenesis, whose activity is inhibited by neuronal differentiation and tumor cell proliferation73. The modulation of the Notch pathway has been indicated that the modulation of the Notch pathway is essential in controlling the fate of cancer stem cells74 and angiogenesis75,76,77. Notch signaling has been shown to induce cell proliferation as well as resistance the chemotherapeutics78. Among the Notch-target genes, there is MYC79. The lichen extract and its metabolite were shown to reduce the activity of the genes Gli and CSL, main transcriptional factors of sonic hdghog, and notch signling pathways80. Similar to this study, we also observed strong inhibition of these two pathways by X. conspersa extract and compounds usnic acid and altraric acid, possibly due to the same mechanism.
The SMADs are intracellular signaling modulators that transmit signals from membrane receptors, such as the type I and type II serine/threonine-kinase receptors, which are in turn activated by inducer TGF-β (transforming growth factor beta) cytokines81. These receptors, upon activation, phosphorylate SMADs, whom then form transcriptional complexes that translocate to the nucleus to regulate target gene expression, functioning as a signaling device for cell regulation throughout the lifespan of the organism81.
STAT3 (signal transducers and activators of transcription) is a transcriptional modulator that is activated by a variety of receptors – such as the EGPR, FGFR, ILR, TLR and SIPR, among others. We induced its activation using cytokine IL-6, and transcription factors Ets, NF-κB, Ap-1, and E2F were activated by PMA. Stat3 is phosphorylated by the IL receptor82 and translocated to the nucleus, where it promotes the expression of prooncogenetic genes83,84. Stat3 is persistently activated in many malignancies, causing cell proliferation and angiogenesis and can promote oncogenesis. Inhibition of Stat3 in HeLa cell signaling caused by the hexane extract of X. conspersa is associated with the inhibition of growth and cell proliferation and with apoptosis85,86,87.
ETS (erythroblast transformation specific) are families of transcription factors associated with cancer88, where changes in gene expression and their deregulation leads to carcinogenesis (cancer, tumor formation) and/or apoptosis (programmed cell death)89. The deregulation of Ets induces cell growth, invasion, and migration in various tumor cells, leading to tumor progression, metastasis, and drug resistance. ETS is also a key factor in angiogenesis, i.e., the neoplastic ability to form blood vessels within a neoplastic tumor that causes the tumor to further increase in mass90. Song et al.91 have also reported strong inhibition of usnic acid in breast tumor angiogenesis and growth by suppressing VEGFR2-mediated AKT and ERK1/2 signaling pathways. This report, along with our observation, shows that X.conspersa extract and its compounds play a strong role in tumor angiogenesis.
NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells) is a transcription factor present in the cytoplasm of each cell. It plays a vital role in regulating the immune response to infection, and disturbances in its regulation are associated with tumor progression92,93,94,95,96. Usnic acid’s anti-inflammatory properties have already been reported in breast cancer97. Yildirim et al.97 and our observation of NFkB inhibition correlated with published data.
Activator protein 1 (AP-1) as a transcription factor controls many cellular processes including differentiation, proliferation, and apoptosis98, and regulates gene expression in response to a variety of stimuli, including cytokines, growth factors, stress, and bacterial and viral infections. Lichen extracts and their active metabolites viz., usnic acid decreased β-catenin mediated TopFlash activity and kitenin mediated AP-1 activity in a dose-dependent manner in non-small cell lung cancer models99.
E2F proteins are members of the family of transcription factors that control the expression of various genes involved in cell cycle regulation100. Among E2F transcriptional targets are cyclins, CDKs, replication proteins, and DNA repair. Inhibition of E2F by our test agents suggests their possible role in cell cycle regulation.
Because of several bioactivities, most notably the modulation of cancer signaling pathways, the secondary metabolites from lichen, usnic acid, and atraric acid present promising research potential to emerge as drugs in the future.
Conclusion
This study revealed that the secondary metabolites detected in X. conspersa extracts can be categorized into phenols, depsides, depsidones, and dibenzofurans. The predominant constituent in the hexane extract was usnic acid, accompanied by small amounts of placodiolic acid, other dibenzofurans, and a few phenols. In contrast, stictic acid was the main compound in the methanolic extract, followed by usnic and norstictic acids. The hexane extract exhibited notable anti-microbial potential, although the overall anti-microbial activity was relatively weak. However, the hexane extract demonstrated significant anti-parasitic action. Among the tested cell lines (VERO, FaDu, SCC-25, HeLa), the hexane extract from X. conspersa showed cytotoxic selectivity against FaDu (SI 7.36) and HeLa (SI 2.19) cells. Given that stictic and usnic acids exhibited higher activity against all studied cell lines compared to the extracts, it can be inferred that these compounds strongly contribute to the overall activity.
A detailed evaluation of the cancer-related signaling pathways in HeLa cells, conducted using two distinct methodologies (luciferase reporter assays), revealed that the hexane extract and usnic acid presented a similar profile of selective inhibition of Stat3, Smad, NF-κB, cMYC, and Notch pathways, confirming that usnic acid significantly contributes to the biological activity of the extract.
The authors acknowledge the limitations of the presented results, noting that the influence of X. conspersa extracts and selected secondary metabolites on cancer-related signaling pathways was assessed in only one cancer cell line. Future studies should expand this research to include other types of cancer cells. Additionally, despite using two independent methodologies for gene expression analysis, it is recommended that future studies incorporate other techniques, such as Western blotting. Further evaluation of the effects of X. conspersa extracts and isolated secondary metabolites on signaling pathways should also be conducted in tumor-bearing mice to assess their potential inclusion in preclinical studies.
Materials and methods
Plant material
The lichen samples of X. conspersa were collected from Krynickie village, Podlaskie Province, NE Poland in May 2018–2021. Plant material was identified by dr Sylwia Kiercul (lichenologist) and was deposited by Prof. Grażyna Łaska at the Herbarium of the Bialystok University of Technology. X. conspersa is a foliose lichen from the Parmeliaceae family, which was collected in open, sunny and dry habitats. It also grows on sunny siliceous rocks, in nitrogen-rich places, but also in more shaded and forested habitats, rarely on wood. Its upper surface is yellow-green, smooth, epruinose to densely isidiate101. The lower surface of the X. conspersa thalli is black with rhizines (simple to furcate) and a white colour of medulla with an algal layer.
The dry plant material (11.25 g) was extracted using the accelerated solvent extraction (ASE) technique (SpeedExtractor E-916, Buchi) with methanol and hexane p.a. grade solvents. The extraction time was 30 min., the temperature was 100 °C, and the pressure was 120 bar. The extracts were filtered through paper filters. Solvent was dried using a rotor evaporator. The obtained extracts were stored at -20 °C before analyses.
Authors confirm that the use of plants/plant parts in the present study complies with international, national and/or institutional guidelines.
Chemical characterization of the studied extracts
Identification of lichen metabolites was performed by means of liquid chromatography-mass spectrometry (LC-ESI-QTOF-MS). Metabolites were separated on Gemini® chromatographic column (3 μm i.d. C18 with TMS endcapping, 110 Å, 100 × 2 mm) supported by a guard column (Phenomenex Inc, Torrance, CA, USA), using Agilent 1200 Infinity HPLC chromatograph equipped with diode array detector (Agilent Technologies, Santa Clara, CA, USA). The solvents were pumped at the flow rate of 0.2 mL/min to perform the gradient of phase A (water with 0.1% formic acid (v/v)) and phase B (0.1% formic acid in acetonitrile (v/v)): 0 min, 0% B; 45 min, 90% B; 46 min, 95% B, 60 min, 95% B. In the case of hexane extract, the gradient was finished at 70 min, 95% B. The column was thermostated at 20 °C. Then, metabolites were detected on an Agilent 6530B QTOF spectrometer equipped with a Dual Agilent Jet Stream spray source (ESI) (Agilent Technologies, Santa Clara, CA, USA). The operating conditions were as follows: sheath gas temp: 325 °C, sheath gas flow: 12 L/min; drying gas temp: 300 °C, drying gas flow: 12 L/min; nebulizer pressure: 40 psig, capillary V (+): 4000 V, skimmer 65 V. The acquisition was performed in a negative ion mode, taking 2 spectra/s in a scan range from 100 to 1700 m/z. Collision-induced dissociation energy was set at 10 and 40 eV. The tentative identification was based on accurate masses and fragmentation patterns, supported by literature sources and the PubChem database43. DAD chromatograms were acquired in λ = 320 nm.
Cell culturing and experiments
Cell lines were acquired from the American Type Culture Collection (ATCC) and included VERO (CCL-81), FaDu (HTB-43), SCC-25 (CRL-1628), and HeLa (CCL-2). The VERO cells were cultured using Dulbecco Modified Eagle Medium (DMEM, Corning, Tewksbury, MA, USA) and cancer cells using Modified Eagle Medium (MEM, Corning), supplemented with antibiotics (Penicillin-Streptomycin Solution, Corning) and fetal bovine serum (FBS, Corning). Phosphate buffered saline (PBS), and trypsin were purchased from Corning, while MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) and DMSO (dimethyl sulfoxide) from Sigma (Sigma-Aldrich, St. Louis, MO, USA). Cells were incubated in a 5% CO2 atmosphere at 37 °C (CO2 incubator, Panasonic Healthcare Co., Tokyo, Japan). The X. conspersa extracts and selected bioactive compounds were dissolved (50 mg/mL) in DMSO (cell-culture grade, PanReac Applichem) to obtain the stock solutions for cytotoxicity testing.
Evaluation of cytotoxicity and anti-cancer potential
The cytotoxicity of X. conspersa extracts and selected bioactive compounds were evaluated in vitro on normal VERO cells and cancer-originating cell lines FaDu (hypopharyngeal squamous cell carcinoma), SCC-25 (tongue squamous cell carcinoma), and HeLa (cervical adenocarcinoma) using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) based protocol, following previously described methodology102. In short, the monolayers of cells in 96-well plates (Falcon, Corning) were treated with serial dilutions of extract or compound stock solutions for 72 h. After this, the media was removed, plates were washed with PBS, and 10% of MTT solution (5 mg/mL) in FBS-free cell media was added, and the incubation continued for the next 4 h. Subsequently, the precipitated formazan crystals were dissolved using SDS/DMF/PBS (14% SDS, 36% DMF, 50% PBS) solvent, and after overnight incubation at 37 °C, the Synergy H1 Multi-Mode Microplate Reader (BioTek Instruments, Inc. Winooski, Vermont, USA) with Gen5 software (ver. 3.09.07; BioTek Instruments, Inc.) was used for absorbance acquisition (540 and 620 nm). Data was exported to GraphPad Prism (version 7.04) to calculate the CC50 values (50% cytotoxic concentration) from dose-response curves. Moreover, anticancer specificity was evaluated and presented as the selectivity indexes (SI) (SI = CC50VERO/CC50Cancer, SI > 1 indicates a potential anticancer selectivity). The cytotoxicity of DMSO at the concentrations present in serial dilutions of tested samples was also evaluated.
The anti-microbial assay
The anti-microbial activity of the X. conspersa extracts was tested against 10 microorganisms. The experiment involved five standard strains of fungi Candida albicans ATCC 90,028, C. glabrata ATCC 90,030, C. krusei ATCC 6258, Aspergillus fumigatus ATCC 204,305 and Cryptococcus neoformans ATCC 90,113, and five standard strains of bacteria Staphylococcus aureus ATCC 29,213, methicillin-resistant S. aureus ATCC 33,591 (MRS), Escherichia coli ATCC 35,218, Pseudomonas aeruginosa ATCC 27,853, and Mycobacterium intracellulare ATCC 23,068. The microorganisms were from the American Type Culture Collection (Manassas, VA). Susceptibility testing was performed using a modified version of the CLSI (formerly NCCLS) methods103,104,105,106. M. intracellulare was tested using a modified method of Franzblau et al.107. Samples were serially diluted in 20% DMSO/saline and transferred in duplicate to 96-well flat-bottom microplates. Microbial inocula were prepared by correcting the OD630 of microbe suspensions in incubation broth to afford the final target inocula. In each assay, drug controls were used for the fungi Amphotericin B (ICN Biomedicals, Ohio) and the bacteria Ciprofloxacin (ICN Biomedicals, Ohio). The readings were made for organisms before and after incubation at either 530 nm using the Biotek Powerwave XS plate reader (Bio-Tek Instruments, Vermont) and for A. fumigatus and M. intracellulare at 544 ex/590 em using the Polarstar Galaxy Plate Reader (BMG LabTechnologies, Germany). To determine minimum fungicidal or bactericidal concentrations 5 µL were removed from each clear well, transferred to agar, and incubated. The MFC/MBC (the lowest test concentration that kills the organism) was also established.
The anti-leishmanial assay
The study of anti-leishmanial properties involved assessing the ability to inhibit the growth of Leishmania donovani, a fly-borne protozoan that causes visceral leishmaniasis. L. donovani strain LSB-51.1 (MHOM/S.D./00/Khartoum) as promastigotes and amastigotes were tested according to the methodology Callahan et al. with modifications108. We also tested amastigotes in differentiated, non-dividing human acute monocytic leukemia cells (THP1) following the methodology of Jain et al.48. L. donovani promastigotes were cultured in RPMI-1640 medium (without sodium bicarbonate and sodium pyruvate) with 10% fetal bovine serum (FBS), pH 7.4 at 26oC. Axenic amastigotes were cultured in the same medium, in which the medium was acidified to pH 5.5, and increased incubation temperature to 32oC. THP1 cells were maintained in RPMI-1640 medium with 10% FBS, pH 7.4 at 37oC in the atmosphere of 5% CO2, which were subcultured twice a week. For anti-leishmanial assay promastigotes and amastigotes were seeded in 96-well plates at an initial concentration equivalent to the early log phase (2 × 105 cells/ml) and allowed to multiply for 60 h in case of promastigotes or for 90 h in case of amastigotes. The metabolism of L. donovani was checked by adding 10 µg of Alamar blue (AbD Serotec catalog no BUF012B) into each well and overnight incubation. The fluorescence was measured at 544 nm excitation and 590 nm emission on Fluostar Galaxy fluorometer (BMG LabTechnologies). The activity assay against intracellular L. donovani amastigotes in THP1 cells was done with the guidelines of Jain et al.48. All experiments were repeated at least three times in triplicate.
Hexane extract from X. conspersa was initially tested in a primary assay at 20 µg/mL and percent inhibitions (% inh.) were calculated relative to negative and positive controls. Samples showing ≥ 50% inhibition were secondary tested. In the secondary assay, all the samples were dissolved to 2 mg/mL and 20 mg/mL. Samples at 20 mg/mL were tested at 40, 8.0, and 1.6 µg/mL, and samples at 2 mg/mL were tested at 20, 4.0, and 0.8 µg/mL, and IC50 values were reported. All IC50 was calculated using the XLFit fit curve fitting software (Microsoft Office, 2019). The drug controls pentamidine and amphotericin B were used as positive controls.
Anti-trypanosomal assay
The anti-trypanosomal tests checked samples for their ability to inhibit Trypanosoma brucei protozoan parasites transmitted by the bite of the tsetse fly, responsible for human trypanosomiasis and animal trypanosomiasis in cattle109. The methanolic and hexane extracts were dried and dissolved in DMSO at a concentration of 20 mg/mL. Blood stage forms of T. brucei (bloodstream form, Strain 427) were from Frederick S. Buckner, Department of Medicine, University of Washington, Seattle, Washington, USA. Parasites were grown in Iscove’s Modified Dulbecco’s Medium (IMDM) medium supplemented with 10% FBS. The culture was maintained at 37oC in 5% CO2. The activity tests were performed according to Jain et al.110. Briefly, the 2 days parasites in the exponential phase were diluted with IMDM medium to 5000 parasites/mL. The maximum permissible limit of DMSO in the assay was 0,5%. For primary assay (single concentration of 20 µg/mL in duplicate) extract dilutions (1 mg/mL) were prepared from the stock extracts (20 mg/mL) in IMDM medium. Each well in 96 well microplate received 4 µL of diluted extract sample and 196 µL of the culture volume. After incubation at 37oC in 5% CO2 for 48 h Alamar blue reagent (10 µL) (AbD Serotec, catalog no BUF012B) was added to each well and the plates were incubated further overnight. The fluorescence was measured at 544 nm excitation and 590 nm emission on a Fluostar Galaxy fluorometer (BMG LabTechnologies). Pentamidine and α-difluoromenthylornithine (DFMO) were tested as control110. The extracts that showed more than 90% inhibition of T. brucei growth in the primary assay were subjected to a secondary assay. Active extracts were tested at concentrations ranging from 20 − 0,8 µg/mL. IC50 values were computed from dose-response.
Cytotoxicity assay against transformed human monocytic (THP1) cells
Active extracts were also tested for toxicity in differentiated THP1 cells at 10 µg/mL concentration110. THP1 from ATCC was grown in RPMI-1640 medium with 10% FBS. The culture was maintained at 37 oC in 5% CO2. A 4-day-old culture of THP1 cells was diluted with RPMI medium to 2.5 × 105 cells/mL. To the culture was added phorbol 12-myristate 13-acetate (PMA) at 25 nq/mL concentration for the transformation of the cells to adherent macrophages48. Each well in 96 well microplates received 200 µL culture and plates were incubated at 37 oC in a 5% CO2 incubator overnight. Plant extracts were diluted in separate plates in RPMI medium and added to medium with THP1, which were again incubated in the same condition for 48 h. After 48 h 10 µL of Alamar blue reagent was added to each well and the plates were incubated further overnight. Standard fluorescence was measured on a fluorometer at 544 nm ex, 590 nm em. Cytotoxicity assay was done for active extracts that showed more than 90% inhibition in the primary assay. Active extracts were tested at concentrations ranging from 20 − 0,8 µg/mL IC50 values were computed from dose-response.
Transfection and luciferase reporter gene assays
HeLa cells (ATCC, CCL-2, Bethesda, MD, USA) were maintained in Dulbecco’s modified Eagle’s medium (DMEM) (Gibco Life Technologies, Grand Island, NY, USA) containing 10% fetal bovine serum (Atlanta Biologicals Inc., Atlanta, GA, USA) and 1% penicillin/streptomycin in a humidified atmosphere of 5% CO2 at 37 °C. HeLa cells were plated in 384 well-white opaque plates at the density of 4300 cells/30 µL of growth medium with 10% FBS and 1% Pen/Strep. After 24 h, the growth medium was replaced with DMEM containing 1% FBS. The cells were transfected with the appropriate plasmid DNA(s) using an X-tremeGENE HP transfection reagent (Roche Applied Science, Indianapolis, IN, USA). The battery of 13 inducible FFLuc reporter gene vectors where expression is driven by enhancer elements (inducer) that bind to specific transcription factors (Table S3-S4). After 24 h of transfection, the test agents were added to the transfected cells, followed 30 min later by an inducing agent (IL-6 for Stat 3, TGF-β for Smad, m-wnt3a for Wnt and PMA for AP-1, NFkB, E2F, Myc, ETS, Notch and Hedgehog). No inducer was added for FoXO, miR-21, Ras, and PMA. After 4–6 h of induction, the cells were lysed by the addition of a One-Glo luciferase assay system (Promega Corporation, Madison, WI, USA). The light output was detected in a Glomax Multi + detection system with Instinct Software (Promega Corporation, Madison, WI, USA). This luciferase assay determines if the test agent was able to inhibit the activation of cancer-related signaling pathways. In the case of FoxO, mi-R21, and Ras enhanced luciferase activity by the test agents was assessed.
Gaussia Luciferase reporter plasmids used in the study contain (1) A genetic element specific to the investigated pathway followed by Gaussia Luciferase (GLuc), a secreted luciferase that generates an up to 100-fold higher bioluminescent signal than Firefly (FFLuc) and Renilla luciferases111; and (2) A secreted alkaline phosphatase (SEAP) gene under a universal promoter (pCMV) for normalization of gene/protein expression and secretion. Both (Gluc and SEAP) are secreted outside the cell to the cell culture media from which it can be readily measured without the need for cell lysis. HeLa cells (ATCC, CCL-2, Bethesda, MD, USA) were seeded on 6-well plates. The next day cells were transfected with reporter plasmids using TurboFect (Thermo Fisher Scientific, Waltham, MA, USA) – two wells per reporter, and incubated for 24 h. In the next step, cells were detached and seeded on 96 well plates. The day after, media was replaced with media containing PMA for 24 h and then media was replaced with media containing extracts for 24 h. After that time media was collected and Gluc as well as SEAP measurements were conducted.
Gaussia Luciferase (GLuc) detection: The substrate for GLuc, coelenterazine, was purchased from Selleck Chemicals (Germany), dissolved in PBS to the final concentration of 20 µM, incubated for 30 min/RT in the absence of light, and used in a 1:1 ratio with cell culture media/supernatant on the 96-well flat white plate. Detection of secreted GLuc was performed using a Biotek Synergy H1 microplate reader with an integration time of 1000 ms and then analyzed. The results were normalized with Secreted alkaline phosphatase (SEAP) assay.
SEAP analysis: The assay was performed according to the manufacturer protocol. In brief QUANTI-Blue (QB) Solution (InvivoGen (France) has been prepared by combining QB reagent and QB buffer in a 1:1 ratio in a 50 mL falcon tube. After 10 min of incubation, 90 µL QB Solution was mixed with 10 µL of the sample on the 96-well transparent plate and incubated for 30 min at 37oC. After the incubation period optical density has been measured at 640 nm on the Tecan M200Pro microplate reader. The results were then analyzed with GraphPad Prism software.
This QUANTI-Blue assay protocol enables the quantification of SEAP in cell culture media through the formation of a colored product, measured at 620–655 nm. This method ensures reproducibility and accuracy in assessing analysis.
Statistical analysis
GraphPad Prism was used for the statistical evaluation of cytotoxicity results. Statistical significance of cytotoxicity towards cancer-originating cell lines was evaluated regarding results obtained for non-cancerous VERO cells based on 2-way analysis of variance (ANOVA) followed by Dunnett’s multiple comparisons test, where p < 0.05 was regarded significant and p < 0.01 as highly significant.
Data availability
All data generated or analyzed during this study are included within the article (and its supplementary information files).
References
Atanasov, A. G. et al. Natural products in drug discovery: advances and opportunities. Nat. Rev. Drug Discov.. 20, 200–216. https://doi.org/10.1038/s41573-020-00114-z (2021).
Chaachouay, N. & Zidane, L. Plant-derived natural products: a source for drug discovery and development. Drugs Drug Candidates 3, 184–207. https://doi.org/10.3390/ddc3010011 (2024).
Mia, M. A. R. et al. The efficacy of natural bioactive compounds against prostate cancer: Molecular targets and synergistic activities. Phytother. Res. 37, 5724–5754 (2023). https://doi.org/10.1002/ptr.8017. PMID: 37786304.
El-Nashar, H. A. S., Mostafa, N. M., Eldahshan, O. A. & Singab, A. N. B. A new antidiabetic and anti-inflammatory biflavonoid from Schinus polygama (cav.) Cabrera leaves. Nat. Prod. Res. 36, 1182–1190. https://doi.org/10.1080/14786419.2020.1864365 (2020).
Nasim, N., Sandeep, I. S. & Mohanty, S. Plant-derived natural products for drug discovery: current approaches and prospects. Nucleus 65, 399–411. https://doi.org/10.1007/s13237-022-00405-3 (2022). PMID: 36276225, PMCID: PMC9579558.
Huneck, S. The significance of lichens and their metabolites. Naturwissenschaften 86, 559–570 (1999).
Hale, M. E. A synopsis of the Lichen Genus Xanthoparmelia (Vainio) Hale (Ascomycotina, Parmeliaceae). Smithson. Contrib. Bot. 74, 1–250 (1990).
Hawksworth, D. L., Blanco, O., Divakar, P. K., Ahti, T. & Crespo, A. A first checklist of parmelioid and similar lichens in Europe and some adjacent territories, adopting revised generic circumscriptions and with indications of species distributions. Lichenologist 40, 1–21 (2008).
Elix, J. A. & Stocker-Wörgötter, E. Biochemistry and secondary metabolites in Lichen Biology (ed. Nash III, T. H.) 104–133 Cambridge, (2008).
Mastan, A., Sreedevi, B. & Pramoda Kumari, J. Evaluation of the in vitro antioxidant and antibacterial activities of secondary metabolites produced from lichens. Asian J. Pharm. Clin. Res. 7, 193–198 (2013).
Lumbsch, H. T. Protocols in Lichenology in Springer Lab. Manual (eds. Kranner, I., Beckett, R. P. & Varma A. K.) 281–295 Berlin, (2002).
Lauterwein, M., Oethinger, M., Belsner, K., Peters, T. & Marre, R. In vitro activities of the lichen secondary metabolites vulpinic acid, (+)-usnic acid, and (–)-usnic acid against aerobic and anaerobic microorganisms. Antimicrob. Agents Chemother. 39, 2541–2543 (1995).
Lohézic-Le Devehat, F. et al. Stictic acid derivatives from the lichen Usnea articulata and their antioxidant activities. J. Nat. Prod. 70, 1218–1220 (2007).
de Amo, G. et al. HPLC isolation of antioxidant constituents from Xanthoparmelia spp. J. Pharm. Biomed. Anal. 53, 165–171 (2010).
Bézivin, C., Tomasi, S., Lohézic-Le Dévéhat, F. & Boustie, J. Cytotoxic activity of some lichen extracts on murine and human cancer cell lines. Phytomed 10, 499–503 (2003).
Łaska, G., Kiercul, S., Stocki, M., Bajguz, A. & Pasco, D. S. Cancer chemopreventive actvity of secondary metabolites isolated from Xanthoparmelia conspersa lichen. Planta Med. 81, 59 (2015).
Opanowicz, M. Ecological role of lichen secondary metabolites. Bot. News 46, 35–44 (2002).
Müller, K. Pharmaceutically relevant metabolites from lichens. Appl. Microbiol. Biotechnol. 56, 9–162 (2001).
McEvoy, M., Nybakken, L., Solhaug, K. A. & Gauslaa, Y. UV triggers the synthesis of the widely distributed secondary lichen compound usnic acid. Mycol. Prog 5, 221–229 (2006).
Molnár, K. & Farkas, E. Current results on biological activities of lichen secondary metabolites: review. Z. Naturforsch 65, 157–173 (2010).
Kanigowski, P. et al. The lichen family Parmeliaceae in Poland. Xanthoparmelia species containing usnic acid. Herzogia 29, 108–119 (2016).
Tay, T., Türk, A. Ő., Türk, H. & Kivanę, M. Evaluation of the antimicrobial activity of the extract of the lichen Ramalina farinacea and its (+)- usnic acid, norstictic acid and protocetraric acid constituents. Z. Naturforsch 56, 384–388 (2004).
Ranković, B. & Mišić, M. The antimicrobial activity of the lichen substances of the lichens Cladonia furcata, Ochrolechia androgyna, Parmelia Caperata, and Parmelia Conspersa. Biotech. Biotchnol. Equip. 22, 1013–1016 (2008).
Mayer, M. et al. C. L. Usnic acid: a non-genotoxic compound with anti-cancer properties. Anti-Cancer Drugs 16, 805–809 (2005).
Studzińska-Sroka, E. Lichens as a Source of Bioactive Compounds (University of Pharmacognosy (Poznan, 2013).
Fazio, A. T. et al. Lichen secondary metabolites from the cultured lichen mykobionts of Teloschistes chrysophthalmus and Ramalina celastri and their antiviral activities. Z. Naturforsch 62, 543–549 (2007).
Yilmaz, M., Türk, A. Ő., Tay, T. & Kivanę, M. The antimicrobial activity of the lichen Cladonia foliacea and its (+)-usnic acid, atranorin and fumarprotocetraric acid constituents. Z. Naturforsch. 59, 249–254 (2004).
Perry, N. B. et al. Antimicrobial, antiviral and cytotoxic activity of New Zealand lichens. Lichenologist 31, 627–636 (1999).
Honda, N. K. et al. Antimycobacterial activity of lichen substances. Phytomed 17, 328–332 (2010).
Sharma, A. K., Sharma, M. C. & Dobhal, M. P. Phytochemical constituents from different species of Parmelia Genus: a review. Der Chem. Sin. 4, 1–11 (2013).
Callaway, E. Deal done over HeLa cells line. Nature 500, 132–133 (2013).
Landry, J. J. M. et al. The genomic and transcriptomic landscape of a HeLa cell line. G3 (Bethesda) 8, 1213–1224. https://doi.org/10.1534/g3.113.005777 (2013).
Forment, J. V., Abderrahmane, K. & Jackson, S. P. Chromothrypsis and cancer: causes and consequences of chromosome shattering. Nat. Rev. Cancer 12, 663–670 (2012).
Korbel, J. O. & Campbell, P. J. Criteria for inference of chromothripsis in cancer genomes. Cell 152, 1226–1236 (2013).
Zaki, M. A. et al. Cytotoxicity and modulation of cancer-related signaling by (Z)- and (E)–3,4,3′,5′-tetramethoxystilbene isolated from Eugenia BrigidaJ. Nat. Prod. 76, 679–684 (2013).
Łaska, G. et al. Extracts from Pulsatilla patens target cancer-related signaling pathways in HeLa cells. Sci. Rep. 11, 1–16. https://doi.org/10.1038/s41598-021-90136-3 (2021).
Balachandran, P. et al. Crosstalk of cancer signaling pathways by cyclic hexapeptides and anthraquinones from Rubia CordifoliaMolecules 26, 735 (2021).
Łaska, G. et al. Pulsatilla vulgaris inhibits cancer proliferation in signaling pathways of 12 reporter genes. Int. J. Mol. Sci. 24, 1–28. https://doi.org/10.3390/ijms24021139 (2023).
Consortium of North American Lichen Herbaria. (2023). http://lichenportal.org/portal/taxa/index.php? taxon = 54736.
Salgado, F. et al. Secondary metabolite profiling of species of the genus Usnea by UHPLC-ESI-OT-MS-MS. Molecules 23, 54. https://doi.org/10.3390/molecules23010054 (2018).
Musharraf, S. G., Kanwal, N., Thadhani, V. N. & Choudharya, M. I. Rapid identification of lichen compounds based on the structure–fragmentation relationship using ESI-MS/MS analysis. Anal. Methods 7, 6066 (2015).
Jin, Y. et al. UHPLC-Q-TOF-MS/MS-oriented characteristic components dataset and multivariate statistical techniques for the holistic quality control of Usnea. RSC Adv. 8, 15487–15500 (2018).
PubChem Database. (2024). https://pubchem.ncbi.nlm.nih.gov
Fidel, P. L., Vazquez, J. A. & Sobel, J. D. CaGlabrataabrata: review of epidemiology, pathogenesis, and clinical disease with comparison to C. albicans. Clin. Microbiol. Rev. 12, 80–96. https://doi.org/10.1128/CMR.12.1.80 (1999).
Virella, G. Microbiology and infectious diseases (1st Polish edition, ed. Heczko, P.) Publisher Urban and Partner (Wrocław, (2015).
Kolenda, R., Burdukiewicz, M. & Schierack, P. A systematic review and meta-analysis of the epidemiology of pathogenic Escherichia coli of calves and the role of calves as reservoirs for human pathogenic E. ColiFront. Cell. Infect. Microbiol. 5, 23 (2015).
Croxen, M. A. et al. Recent advances in understanding enteric pathogenic Escherichia coliClin. Microbiol. Rev. 26, 822–880 (2013).
Jain, S. K., Sahu, R., Walker, L. A. & Tekwani, B. L. A parasite rescue and transformation assay for antileishmanial screening against intracellular Leishmania donovani amastigotes in THP1 human acute monocytic leukemia cell line. J. Vis. Exp. 70, e4054. https://doi.org/10.3791/4054 (2012).
Gillespie, P. M. et al. Status of vaccine research and development of vaccines for leishmaniasis. Vaccine 34, 2992–2995. https://doi.org/10.1016/j.vaccine.2015.12.071 (2016).
Kędzierski, L. Leishmaniasis vaccine: where are we today? J. Glob Infect. Dis. 2, 177–185. https://doi.org/10.4103/0974-777X.62881 (2010).
Handman, E. Leishmaniasis: current status of vaccine development. Clin. Microbiol. Rev. 14, 229–243. https://doi.org/10.1128/CMR.14.2.229-243.2021 (2001).
Paulnock, D. M., Freeman, B. E. & Mansfield, J. M. Modulation of innate immunity by African trypanosomes. Parasitology 137, 2051–2063 (2010).
McGettrick, A. et al. Trypanosoma Brucei metabolite indole pyruvate decreases HIF-1α and glycolysis in macrophages as a mechanism of innate immune evasion. Proc. Natl. Acad. Sci. U S A 113, E7778–E7787 (2016).
De Carvalho, E. A., Andrade, P. P., Silva, N. H., Pereira, E. C. & Figueiredo, R. C. Effect of usnic acid from the lichen Cladonia substellata on Trypanosoma Cruzi in vitro: an ultrastructural study. Micron 36, 155–161. https://doi.org/10.1016/j.micron.2004.09.003 (2005).
Derici, M. K., Cansaran-Duman, D. & Taylan-Özkan, A. Usnic acid causes apoptotic-like death in Leishmania major, L. infantum and L. tropica. 3 Biotech. 8, 384, DOI: (2018). https://doi.org/10.1007/s13205-018-1409-6
Geran, R. I., Greenberg, N. H., Macdonald, M. M., Shumacher, A. M. & Abbott, B. J. Protocols for screening chemical agents and natural products against animal tumors and other biological systems. Cancer Chemother. Rep. 3, 1–103 (1972).
Łaska, G. et al. Phytochemistry and biological activities of Polemonium caeruleum L. Phytochem Lett. 30, 314–323. https://doi.org/10.1016/j.phytol.2019.02.017 (2019).
Kocovic, A. et al. Phytochemical analysis, antioxidant, antimicrobial, and cytotoxic activity of different extracts of Xanthoparmelia stenophylla lichen from Stara Planina, Serbia. Plants 11https://doi.org/10.3390/plants11131624 (2022).
Yeash, E. A., Letwin, L., Malek, L., Suntres, Z. & Knudsen, K. Christopher L. P. Biological activities of undescribed north American lichen species. J. Sci. Food Agric. 97, 4721–4726. https://doi.org/10.1002/jsfa.8340 (2017).
Ozturk, S. et al. Lichens exerts an anti-proliferative effect on human breast and lung cancer cells through induction of apoptosis. Drug Chem. Toxicol. 44, 259–267. https://doi.org/10.1080/01480545.2019.1573825 (2019).
Shrestha, G., El-Naggar, A. M. & St. Clair, L. L. O’Neill, K. L. Anticancer activities of selected species of north American lichen extracts. Phytother Res. 29, 100–107. https://doi.org/10.1002/ptr.5233 (2015).
Cavin, L. G. et al. Transforming growth factoralpha inhibits the intrinsic pathway of c-Myc-induced apoptosis through activation of nuclear factor-kappab in murine hepatocellular carcinomas. Mol. Cancer Res. 3, 403–412 (2005).
Chanvorachote, P., Sriratanasak, N. & Nonpanya, N. C-myc contributes to malignancy of lung cancer. A potential anticancer drug target. Anticancer Res. 40, 609–618. https://doi.org/10.21873/anticanres.13990 (2020).
Kumar, K., Mishra, J. P. & Singh, R. P. Anti-cancer efficacy and mechanisms of usnic acid. Indian J. Pharm. Biol. Res. 7, 1–4 (2019).
Maubant, S. et al. Transcriptome analysis of Wnt3a-treated triple-negative breast cancer cells. PloS One 10, e0122333. https://doi.org/10.1371/journal.pone.0122333 (2015).
Katoh, M. Regulation of WNT3 and WNT3A mRNAs in human cancer cell lines NT2, MCF-7, and MKN45. Int. J. Oncol. 20, 373–377 (2002).
Majchrzak-Celińska, A. et al. Krajka-Kuźniak, V. Lichen secondary metabolites inhibit the Wnt/β-catenin pathway in glioblastoma cells and improve the anticancer effects of Temozolomide. Cells 11, 1084 (2022).
Skoda, A. M. et al. The role of the hedgehog signaling pathway in cancer: a comprehensive review. Bosn J. Basic. Med. Sci. 18, 8–20. https://doi.org/10.17305/bjbms.2018 (2018).
Xu, Y. et al. The role of hedgehog signaling in gastric cancer: molecular mechanisms, clinical potential, and perspective. Cell. Commun. Signal. 17, 157. https://doi.org/10.1186/s12964-019-0479-3 (2019).
Riaz, S. K. et al. Involvement of hedgehog pathway in early onset, aggressive molecular subtypes and metastatic potential of breast cancer. Cell. Commun. Signal. 16https://doi.org/10.1186/s12964-017-0213-y (2018).
Yang, Y. et al. Elucidating hedgehog pathway’s role in HNSCC progression: insights from a 6-gene signature. Sci. Rep. 14, 4686. https://doi.org/10.1038/s41598-024-54937-6 (2024).
Değerli, E., Torun, V. & Cansaran-Duman, D. Mir-185-5p response to usnic acid suppresses proliferation and regulating apoptosis in breast cancer cell by targeting Bcl2. Biol. Res. 53, 1–14 (2020).
Rizzo, P. et al. Rational targeting of notch signaling in cancer. Oncogene 27, 5124–5131. https://doi.org/10.1038/onc.2008.226 (2008).
Viatour, P. et al. Notch signaling inhibits hepatocellular carcinoma following inactivation of the RB pathway. J. Exp. Med. 208, 1963–1976. https://doi.org/10.1084/jem.201101982 (2011).
Van Engeland, N. C. A. et al. Vimentin regulates notch signaling strength and arterial remodeling in response to hemodynamic stress. Sci. Rep. 9, 12415. https://doi.org/10.1038/s41598-019-48218-w (2019).
Antfolk, D. et al. Selective regulation of Notch ligands during angiogenesis is mediated by vimentin. PNAS 114, DOI: (2017). https://doi.org/10.1073/pnas.1703057114
Zalpoor, H. et al. The roles of metabolic profiles and intracellular signaling pathways of tumor microenvironment cells in angiogenesis of solid tumors. Cell. Commun. Signal. 20, 186. https://doi.org/10.1186/s12964-022-00951-y (2022).
Kałafut, J. et al. Rivero-Müller, A. Optogenetic control of Notch1 signaling. Cell. Commun. Signal. 20, 67. https://doi.org/10.1186/s12964-022-00885-5 (2022).
Falo-Sanjuan, J. & Bray, S. J. Decoding the Notch signal. Develop Growth Differ. 62, 4–14. https://doi.org/10.1111/dgd.12644 (2020).
Poulsen-Silva, E., Gordillo-Fuenzalida, F., Atala, C., Moreno, A. A. & Otero, M. C. Bioactive lichen secondary metabolites and their presence in species from Chile. Metabolites 13, 805. https://doi.org/10.3390/metabo13070805 (2023).
Massagué, J. TGF-beta in cancer. Cell 134, 215–230 (2008).
Tkach, M. et al. P42/p44 MAPK-mediated Stat3Ser727 phosphorylation is required for progestin-induced full activation of Stat3 and breast cancer growth. Endocr. -Relat Cancer 20, 197–212. https://doi.org/10.1530/ERC-12-0194 (2013).
Carpenter, R. L. & Lo, H. W. STAT3 target genes relevant to human cancers. Cancers 6, 897–925. https://doi.org/10.3390/cancers6020897 (2014).
Klampfer, L. Signal transducers and activators of transcription (STATs): novel targets of chemopreventive and chemotherapeutic drugs. Curr. Cancer Drug Targets 6, 107–121 (2006).
Mirzaei, H. et al. A new candidate for melanoma therapy? Int. J. Cancer 139, 1683e95. https://doi.org/10.1002/ijc.30224 (2016).
Bhattacharyya, S. et al. Curcumin prevents tumor-induced T cell apoptosis through Stat-5a-mediated Bcl-2 induction. BioMed. Cent. J. Biol. Chem. 282, 15954e64 (2007).
Terlikowska, K., Witkowska, A., Zujko, M., Dobrzycka, B. & Terlikowski, S. Potential application of curcumin and its analogues in the treatment strategy of patients with primary epithelial ovarian cancer. Int. J. Mol. Sci. 15, 21703e22 (2014).
Qi, T. et al. Function and regulation of the PEA3 subfamily of ETS transcription factors in cancer. Am. J. Cancer Res. 10, 3083–3105 (2020).
Oikawa, T. & Yamada, T. Molecular biology of the ets family of transcription factors. Gene 303, 11–34 (2003).
Hollenhorst, P. C., McIntosh, L. P. & Graves, B. J. Genomic and biochemical insights into the specificity of ETS transcription factors. Annu. Rev. Biochem. 80, 437–471 (2011).
Song, Y. et al. Usnic acid inhibits breast tumor angiogenesis and growth by suppressing VEGFR2-mediated AKT and ERK1/2 signaling pathways. Angiogenesis 15, 421–432 (2012).
Nouri, M. et al. Transient Sox9 exspression facilitates resistance to androgen-targeted therapy in prostate cancer. Clin. Cancer Res. 26, 1678–1689, DOI: (2020). https://doi.org/10.1158/1078-0432
Vlahopoulos, S. A. Aberrant control of NF-κB in cancer permits transcriptional and phenotypic plasticity, to curtail dependence on host tissue: molecular mode. Cancer Biol. Med. 14, 254–270. https://doi.org/10.20892/j.issn.2095-3941.2017.0029 (2017).
Hoesel, B. & Schmid, J. A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 12, 86. https://doi.org/10.1186/1476-4598-12-86 (2013).
Pahl, H. Activators and target genes of Rel/NF-κB transcription factors. Oncogene 18, 6853–6866. https://doi.org/10.1038/sj.onc.1203239 (1999).
Deka, K. & Li, Y. Transcriptional regulation during aberrant activation of NF-κB signalling in cancer. Cells 12, 788. https://doi.org/10.3390/cells12050788 (2023).
Yildirim, M. et al. Anti-inflammatory effects of usnic acid in breast cancer. Russ J. Bioorg. Chem. 48, 110–114 (2022).
Tewari, D. et al. Targeting activator protein 1 signaling pathway by bioactive natural agents: possible therapeutic strategy for cancer prevention and intervention. Pharmacol. Res. 128, 366–375. https://doi.org/10.1016/j.phrs.2017.09.014 (2018).
Yang, Y. et al. K. inhibitory activity of (+)-usnic acid against non-small cell lung cancer cell motility. PloS ONE 11, e0146575 (2016).
Chen, H. Z., Tsai, S. Y. & Leone, G. Emerging roles of E2Fs in cancer: an exit from cell cycle control. Nat. Rev. Cancer 9, 785–797 (2009).
Wójciak, H. Lichens, Mosses and FernsWarsaw, (2013).
Świątek, Ł. et al. Chemical characterization of different extracts of Justicia secunda Vahl and determination of their anti-oxidant, anti-enzymatic, anti-viral, and cytotoxic properties. Antioxidants. 12https://doi.org/10.3390/antiox12020509 (2023).
National Committee for Clinical Laboratory Standards. Reference method for broth dilution antifungal susceptibility testing of yeasts: approved standard. Wayne, PA: National Committee for Clinical Laboratory Standards. NCCLS. 22, 15 (2002).
National Committee for Clinical Laboratory Standards. Reference method for broth dilution antifungal susceptibility testing of yeasts: approved standard. Wayne, PA: National Committee for Clinical Laboratory Standards. NCCLS. 22, 16 (2002).
National Committee for Clinical Laboratory Standards. Performance standards for antimicrobial disk susceptibility tests: approved standard. Wayne, PA: National Committee for Clinical Laboratory Standards. NCCLS 23, 18. (2003).
National Committee for Clinical Laboratory Standards. Performance standards for antimicrobial disk susceptibility tests: approved standard. Wayne, PA: National Committee for Clinical Laboratory Standards. NCCLS 26, 2. (2006).
Franzblau, S. G. et al. Rapid, low-technology MIC determination with clinical Mycobacterium tuberculosis isolates by using the microplate Alamar Blue assay. J. Clin. Microbiol. 36, 362–366 (1998).
Callahan, H. L., Portal, A. C., Devereaux, R. & Grogl, M. An axenic amastigote system for drug screening. Antimicrob. Agents Chemother. 41, 818–822 (1997).
Hoet, S., Opperdoes, F., Brun, R. & Adjakidje´, V. Quetin-Leclercq, J. In vitro antitrypanosomal activity of ethno pharmacologically selected Beninese plants. J. Ethnopharmacol. 91, 37–42 (2004).
Jain, S. K., Jacob, M., Walker, L. A. & Tekwani, B. L. Screening north American plant extracts in vitro against Trypanosoma Brucei for discovery of new antitrypanosomal drug leads. BMC Complement. Altern. Med. 16, 131. https://doi.org/10.1186/s12906-016-1122-0 (2016).
Remy, I. & Michnick, S. W. A highly sensitive protein-protein interaction assay based on Gaussia luciferase. Nat. Methods. 3, 977–979 (2006).
Elix. J. A. A Catalogue of Standardized Chromatographic Data and Biosynthetic Relationships for Lichen Substances. Third Edition Canberra, (2014).
Acknowledgements
This study was supported by a grant nr WZ/WB-IIS/4/2023 and by grants nr Ds28 from the Ministry of Science and Higher Education of Poland and in part by a contract from the University of Mississippi Medical Center Cancer Institute Agreements #10001549 to P.B. and #64580110000 to D.S.P.
Author information
Authors and Affiliations
Contributions
G.Ł. conceptualization; G.Ł., E.S., Ł.Ś., P.B., J.Cz., A.R.M. methodology; G.Ł., E.S., Ł.Ś., P.B., D.S.P., J.Cz. B.L.T. investigation; G.Ł., E.S., P.B., M.MT., Ł.Ś., A.R.M., J.Cz. contributed to writing-original draft preparation and editing; G. Ł., E.S., Ł.Ś., A.R.M., D.S.P., P.B. funding. All authors reviewed the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Łaska, G., Sieniawska, E., Świątek, Ł. et al. Evaluating the impact of Xanthoparmelia conspersa extracts on signaling in HeLa cells and exploring their diverse biological activities. Sci Rep 14, 28531 (2024). https://doi.org/10.1038/s41598-024-73599-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-73599-y
