
Abstract
West Nile virus (WNV) is a mosquito-borne zoonotic flavivirus which often causes asymptomatic infection in humans but may develop into a deadly neuroinvasive disease. In this study, we aimed to investigate variables potentially associated with human WNV infection using human and mosquito WNV surveillance and monitoring datasets, established over 20 years, from 2003 to 2022, across the province of Ontario, Canada. We combined climatic and geographic data, mosquito surveillance data (n = 3010 sites), blood donation arboviral detection testing data in the human population, and demographic and socio-economic data from Canadian population censuses. We hypothesized that spatio-temporal indices related to mosquito vector habitat and phenology, in addition to human demographic and socio-economic factors, were associated with WNV infection in the human population. Our results show that high habitat suitability of the main WNV vector in southerly locations of this region, Cx. pipiens/restuans (IRR = 2.0), and variables related to lower income (IRR = 2.8), and shelter infrastructure spending (IRR = 0.7), were key risk factors associated with WNV infection among blood donors from 2003 to 2022 across Ontario (R2 = 0.67). These results may inform points of entry for practical intervention aimed at reducing risk of mosquito-borne pathogens in Canada.
Similar content being viewed by others


Introduction
Human infections with West Nile virus (WNV; family Flaviviridae) are most frequently mild or asymptomatic. Symptomatic infections result in an illness referred to as ‘West Nile fever’, which, in rare cases, particularly in older age groups, may develop into severe and sometimes fatal neuroinvasive disease1. The virus, first introduced to North America in 1999, rapidly spread across the continent, including Canada, in the following years2,3, and is now well established in much of North America4,5. In Canada, the virus was introduced in 2001, has seen an increase in number of cases over time with interannual fluctuations in the number of cases resulting in several epidemics in different provinces, and is now considered endemic (See “Surveillance of West Nile virus – Canada.ca” for more information). Detection tests for WNV have been regularly conducted on most blood donations since 2003, to avoid transmission from donors to recipients6. While the blood donor population isn’t absolutely synonymous with the general population, data from WNV detection tests on blood donations offer a representative snapshot of spatio-temporal variations in WNV infection risk in the human population7. Human case reporting in surveillance is also a useful source of information on WNV risk in North America8, despite having some limitations, including underreporting due to asymptomatic infections, misdiagnoses, and uncertainty as to the precise size of the exposed population9,10. While such limitations may be avoided by using active pathogen surveillance data in the human population11,12,13, data spanning a large area and timeframe are scarce.
Mosquito species of the genus Culex, such as Culex pipiens and Culex restuans, are recognized as important vectors maintaining the enzootic cycle of WNV among avian hosts in northeastern North America. Culex pipiens and Cx. restuans also occasionally feed on mammals, bridging transmission between birds and humans14,15,16,17,18,19. Aedes vexans is very common in northeastern North America, is competent for the transmission of WNV and is highly opportunistic in its blood feeding preferences, meaning it can also effectively bridge transmission between birds and humans20,21,22,23. Environmental factors that influence mosquito development and virus transmission are important for understanding the drivers of human WNV infection risk. Larval development time for Cx. pipiens and Cx. restuans mosquitoes is around two to three weeks while adult lifespan is approximately a month24. Development time for Ae. vexans is around one to two weeks25, and adult lifespan is approximately three weeks23. WNV infection can be detected using nucleic acid tests during viremia from the first day to about two weeks after bite by an infected mosquito, and using serological tests to detect antibodies from approximately a week after bite by an infected mosquito, to approximately six months after bite26. Exposure of humans, including blood donors, to infected mosquitoes depends on their abundance and infection prevalence. Mosquito abundance and infection prevalence vary seasonally and interannually according to long-term impacts of weather and climate affecting survival over winter and short-term effects of weather on mosquito reproduction and activity, and WNV replication in infected mosquitoes27,28,29,30,31,32. Interannual variations in cycles of WNV transmission amongst avian reservoirs, and mosquitoes changing their behaviour to include mammals as sources of blood meals, also impact seasonal and interannual variations in mosquito infection prevalence33. Together, this means that each year, weather over a period of weeks to months, combined with local environmental impacts on mosquito populations and WNV transmission, determine the risk of human infections, and of WNV outbreaks.
Current effects of climate change and land use change in Canada and elsewhere may contribute to an increase in the risk of zoonotic diseases, including mosquito-borne pathogens, such as WNV4,34,35,36,37,38,39,40. Therefore, there is a pressing need for an enhanced understanding of the environmental hazard posed by WNV for public health. Comprehensive temporal and spatial databases of a large variety of climatic, geographic and ecological factors are needed to correctly characterize the distribution and dynamics of mosquito vector populations and mosquito-borne disease transmission. Additionally, certain demographic and socio-economic factors may contribute to both mosquito vector density and exposure to mosquito-borne disease risk41,42,43,44,45,46,47. Indeed, lower socio-economic status seem to be strongly associated with mosquito-borne disease risk, through variation in population density48, quality of household infrastructure44, and education, potentially associated with level of risk perception46. Occupation49, age and sex50 may also influence exposure to mosquitoes. Therefore, regularly updated population censuses offer invaluable information to combine with other data sources to identify modifiable risk factors and better target interventions to reduce mosquito-borne disease risk.
In this study, we aimed to investigate variables potentially associated with human WNV exposure and infection using a large data collection, established over 20 years, from 2003 to 2022, across most of the province of Ontario, Canada. We combined climatic data, geographic data, mosquito surveillance data, arbovirus testing data from human blood donations, and demographic and socio-economic data from Canadian population censuses. We hypothesized that a mixture of spatial and temporal variables, impacting mosquito habitat and phenology, and demographic and socio-economic variables, which are associated with increased mosquito habitat and also increased exposure to mosquitoes, are the main predictors of WNV infection in the human population.
Results
Study area and duration
The study area in Ontario, Canada spanned more than 428,000 km2 with a northern limit (latitude ~ 50–51° N) that matched the extent of most datasets used in this study (Fig. 1). The study spans the years 2003 to 2022. Major urban population centers in our study area are situated in the southeastern section, where a warmer agricultural and residential landscape predominates, and which represents the most populated corridor in Canada from the cities of Windsor to Ottawa. The northern section is dominated by a mixture of forests, wild grasslands and wetlands, with colder temperatures, and ranging from dry in the west at the border with the province of Manitoba, to humid in the center and towards the border with the province of Quebec in the east (Fig. 1).

Spatial representations of land cover classes for the year 2013 according to Agriculture and Agri-Food Canada and United States Geological Survey (A); total daily precipitations averaged from 2003 to 2022 according to National Aerospace Space Agency (B); mean daily temperature averaged from 2003 to 2022 according to National Aerospace Space Agency (C); and projected habitat suitability index for Aedes vexans (D), Culex pipiens/restuans (E) and West Nile virus-infected mosquito vector pools (F). Habitat suitability indices were created using mosquito collection data from Public Health Ontario at 3,010 sampling locations in Ontario, Canada, from 2003 to 2022 (D-E), and a reduced dataset of 2,810 sampling locations from 2003 to 2022 excluding locations where the two vectors were not observed at all (F). Map was created using QGIS 3.34.10.
Mosquito collection data
Based on the available mosquito surveillance data from all 34 Ontario public health units, sampling sites (n = 3,010) were visited at least once, and at most 876 times, from 2003 to 2022. On average, each site was visited 48 times across the study period (Table 1). Sampling sites were visited mostly from May to October when both Cx. pipiens/restuans and Ae. vexans were most likely to be observed in the study area. Out of a total of 145,102 sampling site visits, 99.9% were made from May to October. Interestingly, 396 individuals of a West Nile virus (WNV) vector mostly found in western North America, Culex tarsalis, were observed. About twice as many Ae. vexans than Cx. pipiens/restuans individuals were collected, in total and on average per sampling site visit. Around 60% of sampling visits yielded Cx. pipiens/restuans and/or Ae. vexans (Table 1). Average abundance and occurrence per sampling site visit generally decreased across years for Ae. vexans, but not for Cx. pipiens/restuans (Fig. 2A,C). Average abundance and occurrence per sampling site visit increased from May to August, and then decreased from September to October (Fig. 2B,D). The number of mosquito pools tested for WNV was higher for Cx. pipiens/restuans due to higher priority of Cx. pipiens/restuans for WNV testing by authorities conducting the sampling, and the total number of positive pools was also higher for Cx. pipiens/restuans (Table 1). However, the total number of Cx. pipiens/restuans individuals tested was lower, which is due to fewer collected Cx. pipiens/restuans individuals for this species on average compared to Ae. vexans, and minimum infection rate (MIR) was much higher for Cx. pipiens/restuans (Table 1). There was an overall higher MIR infection rate from 2013 to 2022 (1.9), compared to 2003 to 2012 (1.3), where 2012 was the year with the highest MIR (7.9; Fig. 2A,C). The MIR was also much higher in August and September, compared to June, July and October (Fig. 2B,D). No WNV-positive mosquito pools were observed from November to May of any year.

Histograms showing average abundance across sampling visits (A,B) and average occurrence (% divided by 10) across sampling visits (C,D) of Aedes vexans and Culex pipiens/restuans, and minimum infection rate of West Nile virus (WNV), sampled and tested by Public Health Ontario at 3,010 sampling locations in Ontario, Canada, and number of blood donations to the Canadian Blood Services across Ontario that tested positive for WNV, for each year from 2003 to 2022 (A,C), and for each month from June to October (B,D).
Land cover and climatic data
Land cover data collated from Agriculture Canada and United States Geological Survey consisted of a total of 10 classes, namely two classes of humid cover (open water and wetlands), three classes of open cover (grasslands, croplands and exposed), one class of forested cover, and four classes of residential cover (containing varying degrees of vegetation, or green cover), at a resolution of 120 m. The predominance of each class varied considerably across the study area (Fig. 1A). The four most common landscape classes containing a mosquito sampling site were the medium green residential, high green residential, croplands and forested classes, and the three least common were non-vegetated residential, exposed, and wetlands (Table S1; Fig. 1A). The 20-year averaged total daily precipitation varied between 1.7 to 4.0 mm across the study area. The western part of the study area received less precipitation on average, while a few areas in the center and center-east received more precipitation on average (Fig. 1B). The 20-year averaged mean daily temperature varied between 0.7 and 10.4 °C, and the southern part of the study area was much warmer on average than the northern part, with the warmest area being the corridor from Windsor to Toronto in the extreme south near Lake Erie, and the coldest area being the center-north and northeast of the study area (Fig. 1C). Average total precipitation and mean temperature at cells containing a sampling site were 2.6 mm and 7.5 °C, respectively (Table S1).
Ecological niche modeling analysis
The correlation coefficient between the 20-year averaged total daily precipitation dataset and the 20-year averaged mean daily temperature dataset was low (r < 0.1). Overall, the receiver operating characteristic’s area under the curve (AUC) values were lower for Ae. vexans models compared to Cx. pipiens/restuans and WNV models. Out of 100 models, the best performing model had an AUC = 0.82 for Cx. pipiens/restuans, 0.79 for WNV, and 0.72 for Ae. vexans (Table 2). The ensemble model contained 100, 100 and 8 models, respectively for Cx. pipiens/restuans, WNV and Ae. vexans, with a mean AUC higher than 0.9 for mean habitat suitability index (HSI) values and committee averaging for all species (Table 2). Explanatory variable importance (on a scale of 0 to 1, obtained from 100 permutations of the ensemble niche model) of temperature in the ensemble model was higher than 0.6 and higher than the importance of the other two explanatory variables for both mosquito species and WNV. The importance of precipitation (0.62) was nearly the same as that of temperature for Ae. vexans, but was moderate (< 0.4) for WNV and low (< 0.2) for Cx. pipiens/restuans. Comparatively, land cover was of moderate importance for all species (< 0.4) (Table 2).
Projected HSI of the best performing model for all species led to an area of highest HSI value (1000, 924 and 883, for WNV, Ae. vexans and Cx. pipiens/restuans, respectively, with highest possible maximum value of 1000) concentrated in the extreme southern part of the study area for all species. It was centered around the corridor from Windsor to Toronto near Lake Erie (Fig. 1D–F) for both mosquito species, and was confined to major urban centers for WNV. Another area of high HSI was present for both mosquito species in the southeast around the cities of Kingston and Ottawa near Lake Ontario, where HSI was overall higher for Ae. vexans than for Cx. pipiens/restuans. Ae. vexans also displayed high HSI in areas where Cx. pipiens/restuans did not, particularly in the center and center-east of the study area north of Lake Huron, and in several areas in the extreme west of the study area west of Lake Superior (Fig. 1D,E).
Mean HSI and committee averaging showed high suitability of wetlands and two residential land cover classes, low green and medium green, for both mosquito species and WNV (Fig. 3A,D,G). Grasslands had high suitability for the two mosquito species, but low for WNV. High green residential showed high suitability only for Ae. vexans. Open water, forested, and exposed land cover classes showed low suitability for WNV (Fig. 3A,D,G). Other land cover classes either showed low confidence, as displayed by values near 0.5, or conflict between mean HSI and committee averaging values (Fig. 3A,D,G). Mid-range values of total daily precipitation, i.e. between roughly 2.5 and 3.0 mm, were associated with the highest suitability values for both mosquito species, and lowest values, below 2.5 mm, were associated with the highest suitability values for WNV, according to both mean HSI and committee averaging (Fig. 3B,E,H). Higher mean daily temperature values, i.e. above roughly 6 °C for both mosquito species and 9 °C for WNV, were associated with the highest suitability values, according to both mean HSI and committee averaging (Fig. 3C,F,I).

Graphs of habitat suitability index (black dots/lines) and committee averaging (red dots/lines) of the ensemble niche models for Aedes vexans (A–C), Culex pipiens/restuans (D–F) and West Nile virus (G–I), specifically for land cover classes (A,D,G), total daily precipitations averaged from 2003 to 2022 (B,E,H), and mean daily temperature averaged from 2003 to 2022 (C,F,I). Habitat suitability index and committee averaging values were created using mosquito collection data from Public Health Ontario at 3,010 sampling locations from 2003 to 2022 (A-F), and a reduced dataset of 2,810 sampling locations from 2003 to 2022 excluding locations where the two vectors were not observed at all (G-I).
Socio-economic and demographic data
Using Statistics Canada census datasets from 2016, we identified a total of 14 demographic and socio-economic variables of interest in our analyses, at the level of the census subdivision. One variable captured information on the sex ratio (percent male residents); three variables on age structure (percent residents younger than 15, percent residents older than 64, and percent residents older than 84); two variables on ethnicity (percent residents self-identified as immigrant, percent residents self-identified as Indigenous); one variable on population density (number of residents per km2); two variables on income (mean income, and percent of population earning less than $20,000), two variables on shelter infrastructure (percent shelters needing major repairs, percent residents spending 30% or more of their income on shelter); one variable on education (percent residents with no secondary education); and two variables on occupation (percent residents working in trades, and percent residents working with natural resources).
Blood donation arboviral testing data
A total of more than 6.5 million blood donations from more than 900,000 donors were tested by Canadian Blood Services across the province of Ontario, Canada, from 2003 to 2022 (Table 3). Many donations were coming from the same donor multiple times throughout the study period. We hereby use ‘donor’ to refer to each unique individual who donated blood either once or multiple times, and the term ‘donation’ to reflect each individual donation given. There were more donations from male than female donors, and more donations from donors aged 39 to 65 compared to younger and older (Table 3). However, there were slightly more individual female donors than males, and slightly more individual donors younger than 39 compared to 39 and older. Donors aged older than 65 represented the lowest number of individual donors, among all age groups (Table 3), which is expected due to age-related health issues being more prevalent in this age group that could prevent them for being able to donate blood.
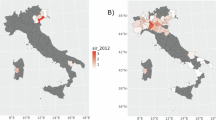
There was a total of 102 donations with a positive test result for WNV infection based on nucleic acid testing. The bulk of WNV-positive blood donations were from male donors and from donors aged 39 to 65. The cumulative WNV infection rate was slightly higher for male than female donors, and much higher for donors older than 65 than for other age groups (Table 3). The WNV infection rate was also much higher in some census subdivisions and some years compared to others. There was an overall higher WNV infection rate in southern Ontario. Some areas with high WNV infection rates, such as some areas in northwestern Ontario, have low population densities, and consequently, each case represents a significant percentage of the total population (Fig. 4). There was also an overall higher WNV infection rate from 2013 to 2022 (2.7 per 100,000), compared to 2003 to 2012 (0.8 per 100,000), where 2018 was the year with the highest WNV infection rate (10.6 per 100,000; Fig. 2A,C). The incidence of WNV infection amongst blood donors was also much higher in August and September, compared to July and October (Fig. 2B,D). No positive WNV blood sample was observed from November to June of any year.

Map of rates of West Nile virus (WNV) infection per 100,000 tested blood donations given to the Canadian Blood Services across Ontario, Canada, from 2003 to 2022. Map was created using QGIS 3.34.10.
Statistical WNV infection modeling analyses
At the analysis level of the census subdivision, we considered a total of 30 variables in zero-inflated negative binomial regression models to identify variables associated with WNV incidence: 14 demographic and socio-economic variables, 10 land cover variables, two climatic variables, two mosquito vector habitat suitability index variables, one WNV habitat suitability index variable, and census subdivision area (Table S2). Two groups of variables had r > 0.7. Population density and all residential land cover classes were highly correlated, and therefore we chose to only keep population density, and dropped the four residential land cover variables. Mean daily temperature and habitat suitability index of both Ae. vexans and Cx. pipiens/restuans were highly correlated, so we chose to keep only habitat suitability index of Cx. pipiens/restuans, and dropped the temperature variable and habitat suitability index of Ae. vexans. Six variables were dropped due to absence of significant association with WNV infection in univariable models (Table S2). We ran a multivariable model with the remaining 18 variables. After model selection, the final model contained thirteen variables, three of which had an IRR that was significantly different from 1 (Table 4). The proportion of households earning less than $20,000 (low income) had an IRR around 2.8 (Table 4), suggesting a strong positive association of this variable with human WNV infection. The proportion of residents spending 30% or more of their income on their shelter (high spending on shelter) had an IRR around 0.7 (Table 4), suggesting a strong negative association of this variable with WNV infection. Habitat suitability index of Cx. pipiens/restuans had an IRR value around 2.0 (Table 4), suggesting a strong positive association of this variable with WNV infection. All other variables did not display a significant association with WNV infection. The R2 value of the final model was around 0.67, which suggests strong statistical power in the final model.
At the analysis level of the individual blood donation, we considered a total of 4 variables in mixed-effects generalized linear regression models to identify factors associated with WNV infection: donor age and sex, and year and month of detection test (Table S2). No pair of variables had r > 0.7. One variable was dropped due to absence of significant association with WNV infection in univariable models (Table S2). The final model contained three variables, all of which had statistically significant slope coefficients, with values very close to 1 (Table 4). Donor age, year of detection test and month of detection test had a slope coefficient between 1.002 and 1.003 (Table 4), suggesting a positive association of these variables with WNV infection. The R2 value of the final model was lower than 0.001, which suggests poor statistical power in the final model, potentially stemming from extremely low variation in the outcome variable, i.e. small number of cases.
Discussion
Our study on spatiotemporal effects of climatic, geographic, ecological, demographic and socio-economic variables on West Nile virus (WNV) infection in the human blood donor population identified multiple modifiable and non-modifiable risk factors that may be practically useful to inform disease prevention and control efforts. First, we identified most regions of Southern Ontario along Saint Lawrence River, Lake Ontario, Lake Erie and south of Lake Huron, to be the main habitat for both Cx. pipiens/restuans and Ae. vexans, which is also where climate is warmest, somewhat more humid and mostly agricultural and urban. Habitat for WNV itself was much narrower, being confined to the Greater Toronto Area and the Windsor region. Some regions in Northern Ontario were also suitable for Ae. vexans. Positive WNV cases in the human blood donor population were mostly detected in Southern Ontario, with very few cases in Northern Ontario. Across years, the abundance and occurrence of Ae. vexans tended to decrease, but remained mostly unchanged for Cx. pipiens/restuans. We observed a peak during the month of August in abundance and occurrence of Cx. pipiens/restuans and Ae. vexans, the two main WNV vectors in our study area, and mosquito vector WNV infection prevalence. Positive mosquito vector pools and infection cases in the blood donor population mostly occurred in the second half of the study period, during the months of August and September. These results mostly support previous literature40,51,52,53,54,55,56,57. However, our study sheds light on the spatiotemporal interplay in abundance and/or occurrence between the two main WNV vectors in northeastern North America, and how this affects WNV infection in the vector populations and in the human population, using longitudinal surveillance data over a long timeframe and over a large study area. In addition, our study leverages blood donor testing data to provide spatiotemporally widespread arboviral detection in the Ontario population, which to date has not been investigated to such an extent in Canada, as opposed to other countries such as the United States7.
The importance of climatic and geographic variables in the ecological niche modeling analyses for WNV vector species showed similar results from a recent study in Eastern Ontario, which performed similar analyses at a smaller but overlapping spatial and temporal scale using mosquito surveillance data from 2011 to 202056. The previous study identified a mediocre to weak importance of all variables. Our study herein suggests a strong effect of temperature for both mosquito vector species and for WNV, with a certain survival threshold upon which suitability increased drastically, and a strong effect of precipitation for Ae. vexans, where average values were optimal, possibly due to a desiccation effect at low values and a flushing effect at high values58. This is to be expected given the larger climatic variation across this study area compared to the previous study. In this study, the threshold of temperature on habitat suitability for WNV was higher compared to that of the two mosquito species. Optimal amounts of precipitation were also lower compared to those of the two mosquito species. Interestingly, land cover had a moderate relative importance for both species, despite land cover classes being all relatively well represented in terms of sampling sites in the present study. Associations between habitat suitability and specific land cover classes were mostly similar between Ae. vexans, Cx. pipiens/restuans and WNV, except for grasslands which were unsuitable for WNV, and high levels of vegetation in urban landscape were not suitable for Cx. pipiens/restuans and WNV. Finally, urban landscapes with low to moderate levels of vegetation and wetlands were suitable for both mosquito species and for WNV, possibly due to their potential as productive breeding habitat. These results are largely supported by previous literature, where high temperatures, and vegetated urban and wetland cover were predictors of both WNV vector habitat and WNV transmission55,56,59,60,61,62.
Our statistical models identified habitat suitability index for Cx. pipiens/restuans as a strong predictor of WNV infection in the human blood donor population, which may also be expanded to two highly correlated variables: habitat suitability index for Ae. vexans and 20-year averaged mean daily temperature. Canada is currently experiencing effects of climate change on vector-borne disease risk, including WNV4,34,35,36,37,38,39,40. Given the strong effect of temperature on mosquito WNV vector habitat, which in turn affects WNV infection in the human population, rigorous surveillance of southerly locations is needed to effectively predict large upticks in human WNV cases across Canada.
Our statistical models also identified the proportion of low-income households and proportion of households where a large portion of the residents’ income is spent on shelter being positively and negatively, respectively, associated with WNV infection in the human population. Household wealth is widely known to be associated with mosquito vector density, potentially through greater perception of risk and access to mosquito control methods44,46,63,64. In a previous analysis at a smaller but completely overlapping spatial and temporal scale in the city of Ottawa, Ontario, from 2007 to 2014, proportion of 60-years-old-and-older shelters was associated with higher WNV risk. A similar result was also observed in Chicago during a WNV outbreak in 200264. This variable is possibly linked to availability of suitable habitats, i.e. breeding sites in suboptimal drainage systems65, used by WNV vector species. Population density and an urban environment were also previously associated with higher WNV risk in Ottawa, Ontario, from 2007–201455, and in Chicago and Detroit during a WNV outbreak in 200260. However, in our study herein there was no significant effect of either population density or residential land cover on WNV infection in the human population, despite using extensive and high-resolution datasets of both types of variables. Certain residential land cover classes do seem to affect habitat suitability of the two mosquito WNV vector species and WNV itself, albeit with a moderate or low importance compared to climatic variables, but this effect does not seem to translate to significantly higher WNV infection in humans, as demonstrated by our multivariable statistical models. These results either suggest limited power to identify an effect of population density or residential land cover by our multivariable statistical models, or that spurious associations in previous studies would have been better explained by alternative unmeasured variables that have been included in this study herein.
Our study has a few limitations. Nucleic acid tests we used have a possibility of cross-reacting with other members of the Japanese encephalitis serocomplex, such as Japanese encephalitis, Murray Valley encephalitis, Saint Louis encephalitis and Kunjin virus66,67,68,69 (leading to a modification to the “National case definition: West Nile virus – Canada.ca” in 2024), which is relevant for donors with a certain travel history or due to vaccination with Japanese encephalitic virus vaccine. Also, most data points in the mosquito sampling data and the human donor data are highly aggregated around urban and metropolitan areas of the study area. However, outcomes of these limitations on the main results of our analyses are likely to be minimal.
In conclusion, results from our study point to several modifiable risk factors that may be used as points of entry for practical intervention aimed at reducing risk of mosquito-borne pathogens in Canada, in a context of increasing mosquito-borne pathogen exposure and illness. For example, our study supports the need for government education campaigns and incentives facilitating renovations aimed at reducing mosquito habitat and/or exposure to mosquitoes, especially in areas with highly suitable habitat for Cx. pipiens/restuans in Southern Ontario, such as the “Prevention of West Nile virus – Canada.ca” program. Such programs and incentives would also be useful in practical intervention measures against Aedes albopictus, an invasive mosquito vector for several pathogens, including dengue virus, which is detected periodically in Southern Ontario. Our study is one of the few using arboviral detection tests over a large area and a large period in the human population to identify factors predictive of WNV infection. Such studies are not impeded by the same spatial, temporal and clinical (e.g., under reporting) biases as those using disease case reports, but are uncommon due to the sheer amount of work needed to produce representative databases.
Materials and methods
Study area and duration
The study area is situated across most of the province of Ontario, in Canada, at and below latitudes 50–51° N (Fig. 1). The study spans the years 2003 to 2022.
Mosquito collection data
Since 2002, Public Health Ontario has collected data on mosquito surveillance activities conducted by public health units in Ontario, which comprise mosquito capture and identification of 22 species/species groups. Capture is performed using the protocol as described in Talbot et al.56. Briefly, capture is performed using light traps designed by the Centers for Disease Control and Prevention (CDC; Atlanta, GA, USA) baited with dry ice for CO2 to increase collection effectiveness, and is performed at least weekly. Culex pipiens/restuans are prioritized over all other species, and Ae. vexans is also of high priority. RNA is extracted from each pool using RNeasy Mini Kit (Qiagen, Hilden, Germany). RNA extracts are tested by quantitative polymerase chain reaction for arbovirus presence70. We used all mosquito capture data available across the entire province of Ontario, Canada, comprising data from all 34 public health units, for the years 2003 to 2022. For each mosquito pool, we calculated the minimum infection rate of West Nile virus (WNV), which is the test outcome (positive = 1; negative = 0), divided by the number of mosquitoes present in the pool, multiplied by 1000.
Land cover and climatic data
We followed the approach of Talbot et al.56 to process land cover and climate data for subsequent ecological niche models. Land cover data for the year 2013 were obtained from Agriculture and Agri-Food Canada (AAFC) and the United States Geological Survey (USGS) with a resolution of 30 m across our study area. Land cover data for the year 2013 were chosen because it is approximately the mid-point of our study period from 2003 to 2022. Annual crop inventory data from AAFC71 comprise seven land cover classes: open water, wetlands, croplands, grasslands, forested, exposed and residential. Residential cover was subdivided into four categories, according to the normalized difference vegetation index (NDVI) from USGS, created using Landsat 8 data (collection 2, level 2, maximum 50% clouds) from May to October 2013, from USGS72,73. The goal of this procedure was to subdivide urban environments according to vegetation, or green cover, which may affect habitat selection by the studied species. Residential areas with NDVI < = 0.15 were referred to as non-vegetated, with NDVI > 0.15 and < = 0.30 as low green, with NDVI > 0.30 and < = 0.60 as medium green and with NDVI > 0.60 as high green residential cover. Data from AAFC and USGS used the same resolution with matching cell frames, and therefore merging of the two datasets could be performed manually.
We considered temperature and precipitation data across the entire study duration. Temperature and precipitation data were obtained from the National Aeronautics and Space Administration (NASA). Given their ease of access, we extracted data on annual total precipitation, and annual maximum and minimum temperature from the annual surface weather and climatological summaries from NASA74 with a resolution of 1,000 m. These data were averaged across the 20 years of the study duration. Then, precipitation data were divided by number of days in a year. The resulting variables were 20-year averaged mean daily temperature and 20-year averaged total daily precipitation for the total period across our study area.
Ecological niche modeling analysis
We followed the approach of Talbot et al.56. The two studied mosquito species were observed at least once in the vast majority of the 3,010 sampling locations over the study period (2,783 for Ae. vexans, or 92%, and 2,686 for Cx. pipiens/restuans, or 90%). At least one mosquito vector species was present in a total of 2,810 sites, for which a WNV detection test could be performed, and lead to at least one positive outcome in a total of 650 sites. Therefore, the number of visits in which each species was observed at least once at each site was calculated, and divided by the total number of visits at each site over the study period. The resulting value is the frequency of observed presence, and is a form of aggregated performance measure often used in species distribution models75, albeit likely over-simplifying the ecology of species involved76. Given the requirement of the approach of transforming frequency of observed presence into a binomial distribution, sites with 50% or more species occurrence for both mosquito species were classified as ‘presence’, and sites with less than 50% as ‘absence’. Sites with at least one positive test outcome for WNV were classified as ‘presence’, and others as ‘absence’.
The random forest algorithm was considered suitable for our analyses because the presence and absence of a species in a given sampling site visit are likely to be affected by the same sampling bias77,78. This decision tree-based approach performs as well as the maximum entropy approach79,80, and better than traditional regression-based approaches when using large datasets sampled over a long duration and a large spatial scale81. We performed the analyses using the ‘biomod2’ package82 in R 4.2.3 (R Development Core Team, Vienna, Austria). We projected all land cover and climatic datasets to Albers Conic Equal Area, which was the original projection of the land cover dataset from AAFC. All explanatory variables were resampled to a cell size of 120 × 120 m and set to be at the same cell frame to reduce spatial bias caused by unequal resolution with the mosquito dataset. We computed the Pearson’s correlation coefficient among climatic variables at sampling sites in R 4.2.3. Potential collinearity problems were considered if r > 0.783. We set the prevalence parameter to 0.5, meaning “presence” and “absence” distributions are considered in equal proportions in the analysis84. For each species, we trained 100 replicate models using 80% of data. To evaluate each model, we computed a receiver operating characteristic’s area under the curve (AUC) using the remaining 20% of data. Data was selected randomly in each model for training versus testing. We kept all other parameters at default values. We used all models with AUC above 0.7 to generate an ensemble niche model82. We generated response plots of the mean habitat suitability index (HSI) across models and committee averaging85, for each explanatory variable. We calculated variable importance for each explanatory variable, which varies from 0 to 1, using a procedure of 100 permutations from the ensemble niche model. Lastly, we created a projected HSI map from the model with the highest AUC for each mosquito species and for WNV.
Socio-economic data
Statistics Canada collects data on a large variety of socio-economic variables during the Census of Population every five years across Canada. Data for the years 2011 and 2016 were initially considered because they are near the mid-point of our study period from 2003 to 2022. Given their easier access and processing, we chose data for the year 2016, which we extracted using the Beyond 20/20 Professional Browser software. We selected all variables that relate to either sex, age, ethnicity, population density, income, shelter infrastructure, education, and occupation, for subsequent statistical WNV infection models. We used these variables in the form they are presented by Statistics Canada. We uploaded data at the level of the census subdivision, to match blood donation arboviral testing data structure.
Blood donation arboviral testing data
Across the study area, since 2003, Canadian Blood Services have tested blood donations across Ontario for WNV types I and II. These tests were performed on all blood donations from 2003 to 2015. As of December 2015, all donations from June to November were tested, but only donations from travelers to certain destinations were tested from December to May. These months usually see much less mosquito activity, and therefore likelihood of exposure to an infected mosquito is negligible. Groups of six donations were tested in 6-unit minipools. Positive minipools were then retested separately for each donation from the corresponding minipool, along with all donations from surrounding areas for the next two weeks. From June 2003, testing was performed using TaqScreen WNV test IUO (F. Hoffmann-La Roche AG, Basel, Switzerland). From June 2007, testing was performed using the IND cobas TaqScreen WNV test for use with the cobas s 201 system. From June 2008, testing was performed using the licensed cobas TaqScreen WNV test for use with the cobas s 201 system. From December 2017, testing was performed using the cobas® WNV – Nucleic acid test for use on the cobas® 6800/8800 Systems. At donor registration, the donor’s date of birth, sex and residential address are recorded. For confidentiality purposes, we used only the donor’s reported sex; year of birth, to obtain approximative age of the blood donor at the time of donation; census subdivision of residence from the Canadian Census of Population boundaries for the year 2016; and unique donor identifier. Blood donor data were used according to independent ethics approval by University of Ottawa, Canadian Blood Services and Public Health Agency of Canada. All methods were designed in compliance with a data sharing agreement between University of Ottawa and Canadian Blood Services. Informed consent was not necessary because data were of secondary use in this study and all donor identifiable personal information was removed by Canadian Blood Services prior to being sent to University of Ottawa for analysis.
To avoid sampling biases leading to spurious associations in our statistical WNV infection models, we chose to exclude all data from census subdivisions where fewer than 1000 WNV tests on blood donations were available, which are mostly located in sparsely populated areas mostly in northeastern parts of the study area, and north of the northern limit of the study area. This exclusion step lead to the removal of 126 subdivisions out of the total of 417 (30%) where blood donor data is available. In the remaining 291 subdivisions, we chose to exclude 4 census subdivisions where total number of residents was lower than 500, because most socio-economic variables were missing for confidentiality reasons. The final dataset contained 287 census subdivisions. Two of these census subdivisions, named “Kenora, Unorganized” and “Thunder Bay, Unorganized” had boundaries above the northern limit of the study area. However, the large majority of blood donations in these subdivisions were conducted in the south, near the Kenora and Thunder Bay townships, respectively, which is also where the large majority of the residents of these subdivisions live. For these reasons, we considered only the portion in these two subdivisions that lie within our study area (Figs. 1, 4).
Statistical WNV infection modeling analyses
To attain our main objective, we investigated the effect of a wide range of climatic, geographic, ecological, demographic and socio-economic variables on WNV infection in the human population, using data from WNV tests on blood donations. These factors were carefully chosen a priori to include variables most likely affecting WNV risk in the human population, including aspects related to mosquito occurrence, mosquito activity, and characteristics of residents linked to higher exposure to mosquito bites. We conducted these analyses at two different levels: one at the census subdivision, and one at the individual donation.
For the first level of analysis, we gathered all arboviral testing data from blood donations, for which residence information is available, from 2003 to 2022, and grouped them within the respective census subdivision of residence of the blood donor, aggregating all years together. We ran zero-inflated negative binomial regression models, using the ‘pscl’ package86 in R 4.2.3, where we used the number of positive cases within census subdivisions as outcome variable, and number of donations tested within census subdivisions as offset variable. We first ran univariable regression models for each explanatory variable separately, namely selected socio-economic variables, land cover variables (percent cover of each class in census subdivisions), climatic variables (20-year averaged daily total precipitations and 20-year averaged daily mean temperature, averaged across census subdivisions), habitat suitability index variables (for each mosquito vector species and for WNV, averaged across census subdivisions), and lastly, the census subdivision geographic area (in km2), which can be stochastically associated with number of positive cases. Prior to these analyses, we subtracted the mean and divided by the standard deviation of all values for numerical variables. We then ran multivariable models with all explanatory variables that displayed a significant effect in an univariable model.
For the second level of analysis, we gathered all arboviral testing data from blood donations, for which residence information is available, from 2003 to 2022, keeping information on the month and year of donation. We ran mixed-effects generalized linear regression models, using the ‘lme4’ package87 in R 4.2.3, where we used the test result as outcome variable, where 1 is positive and 0 is negative, and a hierarchical random-effects term, which is the unique donor number nested within the census subdivision of residence. We first ran univariable regression models for each explanatory variable separately, namely donor sex, donor age, month of test, and year of test. Prior to these analyses, we subtracted the mean and divided by the standard deviation of all values for numerical variables. We then ran multivariable models with all explanatory variables that displayed a significant effect in an univariable model.
For both analysis levels, we used Pearson’s correlation coefficient in R 4.2.3, to identify correlation among all selected variables, and dropped one variable from each pair of variables displaying r > 0.783. Next, we selected all variables displaying a significant univariable association (P < 0.05), with the outcome variable into a multivariable generalized linear regression analysis, using the same packages as previously described, in R 4.2.3. For the analysis at the level of the census subdivision, given the large number of variables included, we proceeded with a two-step model selection approach using the Bayesian Information Criterion (BIC), i.e. the Akaike Information Criterion using the logarithm of the number of observations as the k parameter. We computed a BIC value for the full model, which includes all variables, and for all combinations of the full model excluding one variable. A BIC value decrease of 2 or more is considered positive evidence for a variable displaying little effect on the response variable88. We ran a new model, but this time excluding all variables that, when dropped from the full model, caused a decrease of the BIC value of more than 2. We then reapplied the same model selection approach as a second step on this new model. Using this model selection approach, we ensured that only the most important variables were retained in the resulting final model. In both analysis levels, we computed the most appropriate model outputs (incidence rate ratios, i.e. the exponents of the slope coefficients, for the level of the census subdivision, and slope coefficients, for the level of the individual donation), 95% confidence intervals for the model outputs, and respective P values. Finally, we computed the R2 of the final model, using the ‘modelsummary’ package89 in R 4.2.3.
Data availability
Blood donor data from Canadian Blood Services is not available due to privacy reasons according to our data sharing agreement between University of Ottawa and Canadian Blood Services. All other data are freely available on the webpage of the relevant authority: Public Health Ontario (https://www.publichealthontario.ca/en/Data-and-Analysis/Using-Data/Data-Requests) for mosquito capture data, Statistics Canada (https://www150.statcan.gc.ca/n1/en/type/data) for socio-economic data, Agriculture and Agri-Food Canada (https://www.agr.gc.ca/atlas/apps/aef/main/index_en.html?AGRIAPP=23) and United States Geological Survey (https://earthexplorer.usgs.gov/) for geographic data, and National Aeronautics and Space Administration (https://www.earthdata.nasa.gov/) for climatic data.
References
Gubler, D. J. The continuing spread of West Nile virus in the western hemisphere. Clin. Infect. Dis. 45, 1039–1046 (2007).
Reisen, W. K. Ecology of West Nile virus in North America. Viruses 5, 2079–2105 (2013).
Chancey, C., Grinev, A., Volkova, E. & Rios, M. The global ecology and epidemiology of West Nile virus. BioMed. Res. Int. 2015, 1–20 (2015).
Chen, C. et al. Climate change and West Nile virus in a highly endemic region of North America. IJERPH 10, 3052–3071 (2013).
Shing, E. et al. The direct healthcare costs attributable to West Nile virus illness in Ontario, Canada: a population-based cohort study using laboratory and health administrative data. BMC Infect. Dis. 19, 1059 (2019).
Todoric, D. et al. An overview of the National West Nile Virus Surveillance System in Canada: A One Health approach. CCDR 48, 181–187 (2022).
Busch, M. P. et al. West Nile virus infections projected from blood donor screening data, United States, 2003. Emerg. Infect. Dis. 12, 395–402 (2006).
Murray, J. & Cohen, A. L. Infectious disease surveillance. In International Encyclopedia of Public Health (eds Murray, J. & Cohen, A. L.) 222–229 (Elsevier, 2017). https://doi.org/10.1016/B978-0-12-803678-5.00517-8.
Doyle, T. J. Completeness of notifiable infectious disease reporting in the United States: an analytical literature review. Am. J. Epidemiol. 155, 866–874 (2002).
Amodio, E., Zarcone, M., Casuccio, A. & Vitale, F. Trends in epidemiology: the role of denominator fluctuation in population based estimates. AIMS Public Health 8, 500–506 (2021).
Won, E. J. et al. Under-diagnosis of vector-borne diseases among individuals suspected of having Scrub Typhus in South Korea. PLoS One 18, e0286631 (2023).
Keita, A. K. et al. The detection of vector-borne-disease-related DNA in human stool paves the way to large epidemiological studies. Eur. J. Epidemiol. 30, 1021–1026 (2015).
Koliopoulos, P. et al. Multiplex-RT-PCR-ELISA panel for detecting mosquito-borne pathogens: Plasmodium sp. preserved and eluted from dried blood spots on sample cards. Malar. J. 20, 66 (2021).
Patrican, L. A. et al. Host-feeding patterns of Culex mosquitoes in relation to trap habitat. Emerg. Infect. Dis. 13, 1921–1923 (2007).
Hamer, G. L. et al. Host selection by Culex pipiens mosquitoes and West Nile virus amplification. Am. J. Trop. Med. Hyg. 80, 268–278 (2009).
Hamer, G. L. et al. Culex pipiens (Diptera: Culicidae): a bridge vector of West Nile virus to humans. J. Med. Entomol. 45, 125–128 (2008).
Ebel, G. D., Rochlin, I., Longacker, J. & Kramer, L. D. Culex restuans (Diptera: Culicidae) relative abundance and vector competence for West Nile virus. J. Med. Entomol. 42, 838–843 (2005).
Farajollahi, A., Fonseca, D. M., Kramer, L. D. & Marm Kilpatrick, A. “Bird biting” mosquitoes and human disease: a review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 11, 1577–1585 (2011).
Andreadis, T. G. The contribution of Culex pipiens complex mosquitoes to transmission and persistence of West Nile virus in North America. J. Am. Mosquito Control Assoc. 28, 137–151 (2012).
Tiawsirisup, S. et al. Vector competence of Aedes vexans (Diptera: Culicidae) for West Nile virus and potential as an enzootic vector. J. Med. Entomol. 45, 452–457 (2008).
Molaei, G. & Andreadis, T. G. Identification of avian- and mammalian-derived bloodmeals in Aedes vexans and Culiseta melanura (Diptera: Culicidae) and its implication for West Nile virus transmission in Connecticut, U. S. A. J. Med. Entomol. 43, 1088–1093 (2006).
Greenberg, J. A., Lujan, D. A., DiMenna, M. A., Wearing, H. J. & Hofkin, B. V. Identification of blood meal sources in Aedes vexans and Culex quinquefasciatus in Bernalillo County, New Mexico. J. Insect Sci. 13, 1–12 (2013).
Anderson, J. F., Main, A. J. & Ferrandino, F. J. Horizontal and vertical transmission of West Nile virus by Aedes vexans (Diptera: Culicidae). J. Med. Entomol. 57, 1614–1618 (2020).
Moser, S. K. et al. Scoping review of Culex mosquito life history trait heterogeneity in response to temperature. Parasites Vectors 16, 200 (2023).
Shaman, J. Using a dynamic hydrology model to predict mosquito abundances in flood and swamp water. Emerg. Infect. Dis. 8, 8–13 (2002).
Busch, M. P. et al. Virus and antibody dynamics in acute West Nile virus infection. J. Infect. Dis. 198, 984–993 (2008).
Dohm, D. J. & Turell, M. J. Effect of incubation at overwintering temperatures on the replication of West Nile virus in New York Culex pipiens (Diptera: Culicidae). ME 38, 462–464 (2001).
Dohm, D. J., O’Guinn, M. L. & Turell, M. J. Effect of environmental temperature on the ability of Culex pipiens (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 39, 221–225 (2002).
Roiz, D., Ruiz, S., Soriguer, R. & Figuerola, J. Climatic effects on mosquito abundance in Mediterranean wetlands. Parasites Vectors 7, 333 (2014).
Giménez, J. O., Fischer, S., Zalazar, L. & Stein, M. Cold season mortality under natural conditions and subsequent hatching response of Aedes (Stegomyia) aegypti (Diptera: Culicidae) eggs in a subtropical city of Argentina. J. Med. Entomol. 52, 879–885 (2015).
Eisen, L. et al. The impact of temperature on the bionomics of Aedes (Stegomyia) aegypti, with special reference to the cool geographic range margins. J. Med. Entomol. 51, 496–516 (2014).
Shocket, M. S. et al. Transmission of West Nile and five other temperate mosquito-borne viruses peaks at temperatures between 23 °C and 26 °C. eLife 9, e58511 (2020).
Ozdenerol, E., Taff, G. & Akkus, C. Exploring the spatio-temporal dynamics of reservoir hosts, vectors, and human hosts of West Nile Virus: a review of the recent literature. Int. J. Environ. Res. Public Health 10, 5399–5432 (2013).
Allan, B. F., Keesing, F. & Ostfeld, R. S. Effect of forest fragmentation on Lyme disease risk. Conserv. Biol. 17, 267–272 (2003).
Canadian Paediatric Society. West Nile virus in the context of climate change. Paediatr. Child Health 13, 399–400 (2008).
Kim, B. I. et al. A conceptual model for the impact of climate change on fox rabies in Alaska, 1980–2010. Zoonoses Public Health 61, 72–80 (2014).
Gardner, A. M., Lampman, R. L. & Muturi, E. J. Land use patterns and the risk of West Nile virus transmission in Central Illinois. Vector-Borne Zoonot. Dis. 14, 338–345 (2014).
Hoover, K. C. & Barker, C. M. West Nile virus, climate change, and circumpolar vulnerability. WIREs Clim. Change 7, 283–300 (2016).
Ludwig, A. et al. Increased risk of endemic mosquito-borne diseases in Canada due to climate change. CCDR 45, 91–97 (2019).
Rakotoarinia, M. R. et al. Effects of land use and weather on the presence and abundance of mosquito-borne disease vectors in a urban and agricultural landscape in Eastern Ontario, Canada. PLoS One 17, e0262376 (2022).
Elmqvist, T. et al. Urbanization, Biodiversity and Ecosystem Services: Challenges and Opportunities (Springer Netherlands, 2013). https://doi.org/10.1007/978-94-007-7088-1.
Li, Y. et al. Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship. PLoS Negl. Trop. Dis. 8, e3301 (2014).
Kjellstrom, T. et al. Urban environmental health hazards and health equity. J. Urban Health 84, 86–97 (2007).
Donnelly, M. A. P., Kluh, S., Snyder, R. E. & Barker, C. M. Quantifying sociodemographic heterogeneities in the distribution of Aedes aegypti among California households. PLoS Negl. Trop. Dis. 14, e0008408 (2020).
Little, E. et al. Socio-ecological mechanisms supporting high densities of Aedes albopictus (Diptera: Culicidae) in Baltimore, MD. J. Med. Entomol. 54, 1183–1192 (2017).
Whiteman, A. et al. Aedes mosquito infestation in socioeconomically contrasting neighborhoods of Panama City. EcoHealth 16, 210–221 (2019).
Dowling, Z. et al. Linking mosquito infestation to resident socioeconomic status, knowledge, and source reduction practices in suburban Washington, DC. EcoHealth 10, 36–47 (2013).
Fuentes-Vallejo, M. et al. Territorial analysis of Aedes aegypti distribution in two Colombian cities: a chorematic and ecosystem approach. Cad. Saude Publ. 31, 517–530 (2015).
Vonesch, N. et al. Emerging zoonotic viral infections of occupational health importance. Pathogens Dis. 77, ftz018 (2019).
Ellwanger, J. H., Cardoso, J. D. C. & Chies, J. A. B. Variability in human attractiveness to mosquitoes. Curr. Res. Parasitol. Vector-Borne Dis. 1, 100058 (2021).
Ripoche, M., Campagna, C., Ludwig, A., Ogden, N. H. & Leighton, P. A. Short-term forecasting of daily abundance of West Nile virus vectors Culex pipiens-restuans (Diptera: Culicidae) and Aedes vexans based on weather conditions in Southern Québec (Canada). J. Med. Entomol. 56, 859–872 (2019).
Giordano, B. V., Kaur, S. & Hunter, F. F. West Nile virus in Ontario, Canada: a twelve-year analysis of human case prevalence, mosquito surveillance, and climate data. PLoS One 12, e0183568 (2017).
Giordano, B., Turner, K. & Hunter, F. Geospatial analysis and seasonal distribution of West Nile virus vectors (Diptera: Culicidae) in southern Ontario, Canada. Int. J. Environ. Res. Public Health 15, 614 (2018).
Mallya, S. et al. Factors associated with human West Nile virus infection in Ontario: a generalized linear mixed modelling approach. BMC Infect. Dis. 18, 1–9 (2018).
Talbot, B., Ardis, M. & Kulkarni, M. A. Influence of demography, land use, and urban form on West Nile virus risk and human West Nile virus incidence in Ottawa, Canada. Vector-Borne Zoonot. Dis. 19, 533–539 (2019).
Talbot, B. et al. Ecological niche and positive clusters of two West Nile virus vectors in Ontario, Canada. EcoHealth 20, 249–262 (2023).
Talbot, B., Caron-Lévesque, M., Ardis, M., Kryuchkov, R. & Kulkarni, M. A. Linking bird and mosquito data to assess spatiotemporal West Nile virus risk in humans. EcoHealth 16, 70–81 (2019).
Chandra, G. & Mukherjee, D. Effect of climate change on mosquito population and changing pattern of some diseases transmitted by them. In Advances in Animal Experimentation and Modeling (eds Chandra, G. & Mukherjee, D.) 455–460 (Elsevier, 2022). https://doi.org/10.1016/B978-0-323-90583-1.00030-1.
Ludwig, A., Rousseu, F., Kotchi, S. O., Allostry, J. & Fournier, R. A. Mapping the abundance of endemic mosquito-borne diseases vectors in southern Quebec. BMC Public Health 23, 924 (2023).
Ruiz, M. O., Walker, E. D., Foster, E. S., Haramis, L. D. & Kitron, U. D. Association of West Nile virus illness and urban landscapes in Chicago and Detroit. Int. J. Health Geogr. 6, 10 (2007).
Johnson, B. J. et al. The roles of mosquito and bird communities on the prevalence of West Nile virus in urban wetland and residential habitats. Urban Ecosyst. 15, 513–531 (2012).
Skaff, N. K. & Cheruvelil, K. S. Fine-scale wetland features mediate vector and climate-dependent macroscale patterns in human West Nile virus incidence. Landsc. Ecol. 31, 1615–1628 (2016).
Talbot, B. et al. Determinants of Aedes mosquito density as an indicator of arbovirus transmission risk in three sites affected by co-circulation of globally spreading arboviruses in Colombia, Ecuador and Argentina. Parasites Vectors 14, 482 (2021).
Ruiz, M. O., Tedesco, C., McTighe, T. J., Austin, C. & Kitron, U. Environmental and social determinants of human risk during a West Nile virus outbreak in the greater Chicago area, 2002. Int. J. Health Geogr. 3, 8 (2004).
Ozdenerol, E., Bialkowska-Jelinska, E. & Taff, G. N. Locating suitable habitats for West Nile Virus-infected mosquitoes through association of environmental characteristics with infected mosquito locations: a case study in Shelby County, Tennessee. Int. J. Health Geogr. 7, 12 (2008).
Drews, S. J. et al. A case series of inactivated Japanese encephalitis virus vaccination associated with positive West Nile virus blood donor screening nucleic acid tests. Transfusion 60, 1097–1103 (2020).
O’Brien, S. F. et al. West Nile virus in 2006 and 2007: the Canadian Blood Services’ experience. Transfusion 50, 1118–1125 (2010).
Vamvakas, E. C., Kleinman, S., Hume, H. & Sher, G. D. The development of West Nile virus safety policies by Canadian Blood Services: guiding principles and a comparison between Canada and the United States. Transf. Med. Rev. 20, 97–109 (2006).
Postler, T. S. et al. Renaming of the genus Flavivirus to Orthoflavivirus and extension of binomial species names within the family Flaviviridae. Arch. Virol. 168, 224 (2023).
Lanciotti, R. S. et al. Rapid detection of West Nile virus from human clinical specimens, field-collected mosquitoes, and avian samples by a TaqMan reverse transcriptase-PCR assay. J. Clin. Microbiol. 38, 4066–4071 (2000).
Agriculture and Agri-Food Canada. Annual Crop Inventory. https://www.agr.gc.ca/atlas/apps/metrics/index-fr.html?appid=aci-iac&lang=en (2022).
Masek, J. G. et al. A Landsat surface reflectance dataset for North America, 1990–2000. IEEE Geosci. Remote Sensing Lett. 3, 68–72 (2006).
Vermote, E., Justice, C., Claverie, M. & Franch, B. Preliminary analysis of the performance of the Landsat 8/OLI land surface reflectance product. Remote Sens. Environ. 185, 46–56 (2016).
Thornton, M.M. et al. Daymet: daily surface weather data on a 1-km grid for North America, Version 4. 0 MB. https://doi.org/10.3334/ORNLDAAC/1840 (2020).
Halvorsen, R. A gradient analytic perspective on distribution modelling. Sommerfeltia 35, 1–165 (2012).
Rocchini, D. et al. A quixotic view of spatial bias in modelling the distribution of species and their diversity. NPJ Biodivers. 2, 10 (2023).
Sillero, N. & Barbosa, A. M. Common mistakes in ecological niche models. Int. J. Geogr. Sci. 35, 213–226 (2021).
Phillips, S. J. et al. Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecol. Appl. 19, 181–197 (2009).
Zhao, Z., Xiao, N., Shen, M. & Li, J. Comparison between optimized MaxEnt and random forest modeling in predicting potential distribution: a case study with Quasipaa boulengeri in China. Sci. Total Environ. 842, 156867 (2022).
Beeman, S. P., Morrison, A. M., Unnasch, T. R. & Unnasch, R. S. Ensemble ecological niche modeling of West Nile virus probability in Florida. PLoS One 16, e0256868 (2021).
Mondal, R. & Bhat, A. Comparison of regression-based and machine learning techniques to explain alpha diversity of fish communities in streams of central and eastern India. Ecol. Indic. 129, 107922 (2021).
Thuiller, W., Lafourcade, B., Engler, R. & Araújo, M. B. BIOMOD - a platform for ensemble forecasting of species distributions. Ecography 32, 369–373 (2009).
Akoglu, H. User’s guide to correlation coefficients. Turk. J. Emerg. Med. 18, 91–93 (2018).
Barbet-Massin, M., Jiguet, F., Albert, C. H. & Thuiller, W. Selecting pseudo-absences for species distribution models: how, where and how many?. Methods Ecol. Evol. 3, 327–338 (2012).
Araujo, M. & New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 22, 42–47 (2007).
Zeileis, A., Kleiber, C. & Jackman, S. Regression models for count data in R. J. Stat. Soft. https://doi.org/10.18637/jss.v027.i08 (2008).
Bates, D., Mächler, M., Bolker, B. & Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Soft. https://doi.org/10.18637/jss.v067.i01 (2015).
Raftery, A. E. Bayesian model selection in social research. Sociol. Methodol. 25, 111 (1995).
Arel-Bundock, V. modelsummary: Data and model summaries in R. J. Stat. Soft. https://doi.org/10.18637/jss.v103.i01 (2022).
Acknowledgements
We thank Canadian Blood Services (in particular Bill Ferguson), Public Health Ontario (in particular Curtis Russell), Statistics Canada, Agriculture and Agri-Food Canada, United States Geological Survey, and National Aeronautics and Space Administration, for access to blood donor, mosquito capture, socio-economic, geographic and climatic data used in this study. We also thank Maxime Rioux-Rousseau for their help collecting and processing geographic and climatic data sources. This project was supported by grants from the Canadian Institutes of Health Research and Natural Science and Engineering Research Council of Canada to M.A.K.
Funding
The funding was supported by Canadian Institutes of Health Research, PJT 166112, Natural Sciences and Engineering Research Council of Canada, RGPIN-2019-04730.
Author information
Authors and Affiliations
Contributions
B.T. contributed to the conception of the study, performed analyses, interpreted results, and wrote the manuscript draft. M.A.K. and A.L. contributed in the conception of the study, interpreted results and edited the manuscript draft. N.H.O., S.F.O. and S.J.D. contributed in the conception of the study and edited the manuscript draft.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Talbot, B., Ludwig, A., O’Brien, S.F. et al. Spatial and temporal analysis of West Nile virus infection in mosquito and human populations based on arboviral detection testing data. Sci Rep 14, 31343 (2024). https://doi.org/10.1038/s41598-024-82739-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-024-82739-3
This article is cited by
-
Emerging West Nile virus infections in Türkiye
European Journal of Clinical Microbiology & Infectious Diseases (2025)
