
Abstract
Adhesions and poor healing are major complications after Achilles tendon injury, and there is no effective solution to this problem. The purpose of this study is to determine whether the biomimetic peritoneurosis can solve the above problems in the process of Achilles tendon healing; This study investigated the adhesion and proliferation of tendon-derived stem cells (TDSCs) on dAM in vitro, as well as their tenogenic differentiation. The effects of Achilles tendon rupture on tendon regeneration were assessed in vivo by using an Achilles tendon rupture model in rats; Finally, from in vitro mRNA transcriptome sequencing combined with in vivo Achilles tendon protein to omics analysis to explore the repair mechanism of Achilles tendon rupture. Student’s t-tests were used to assess the significance of observed differences between the two experimental groups. Multiple groups were compared using one-way analysis of variances (ANOVAs), followed by post hoc Bonferroni comparisons. The main findings of this study are that cell adhesion, proliferation, and differentiation of TDSCs were enhanced by dAM. Implanted dAM + TDSCs significantly accelerated tendon regeneration in vivo. In addition, extracellular matrix-related differential genes and proteins were screened by mRNA transcriptometry in vitro and proteomic analysis of Achilles tendon in vivo, and ERK signaling pathway was further explored to participate in the repair of Achilles tendon rupture. The dAM-TDSCs composite biomimetic peritendinous membrane material can effectively promote the healing of Achilles tendon. It provides a new direction for the development of biomimetic peritendinous membrane materials.
Similar content being viewed by others
Introduction
The most substantial tendon in the human body and an essential component of daily bodily functions is the Achilles tendon, which is made up of the union of the soleus and gastrocnemius muscles1. As more people exercise and consume dangerous medicines (such as quinolones), the prevalence of Achilles tendon rupture has grown in several nations in recent years2,3. Currently, non-surgical, surgical, and tissue engineering treatments are the main options for acute Achilles tendon rupture. The main complications following Achilles tendon rupture healing, whether surgical or non-operative, are postoperative Achilles tendon adhesions, poor healing, and postoperative chronic pain. Studies have shown that Achilles tendon rupture has a comparable efficacy after surgical and non-surgical treatment4, and that there is almost no vascular distribution in Achilles tendon tissue5. Additionally, patients are now having issues with it6.
The use of tissue engineering to treat a severe Achilles tendonitis is one of the research hotspots, allowing for the restoration of the original structure and functionality of the Achilles tendon even if there is currently no universally accepted treatment plan for acute Achilles tendon rupture. Biologically active cytokines, seed cells, and cell scaffolds are the core components of tissue engineering materials7,8. To ensure successful integration within the host tissue, a bioactive scaffold must exhibit biocompatibility and mechanical properties comparable to native tissue. To facilitate cell adhesion, proliferation, and differentiation, the bioactive scaffold should possess a porous structure with varying pore sizes. This porous structure can also make it easier for cells to move nutrients and waste products. Second, the scaffold itself is abundant in growth factors and can more effectively encourage cell proliferation since it serves as the “soil” of seed cells. The target organ’s tissue repair rate should be equivalent to the pace at which the cell scaffold degrades, and the degradation products should be benign. Third, to more accurately mimic the condition of healthy tissue, the mechanical properties of the cell scaffold should be comparable to those of the target tissue. In order to be used for clinical treatment on a large scale, the optimal cell scaffold should also be widely available, easy to take materials, and simple to create9.
Amniotic membranes (AM), the innermost layer of the fetal membranes, are derived from the placenta of full-term newborns. The structure is a see-through membrane made up of an inner layer of epithelial cells situated on a basement membrane, which is linked to a thin connective tissue membrane by thread-like filaments10,11. The fibrous network structure of the amniotic membrane’s main components, type III, type IV, and type V collagen, contains the transformation of growth factors of active ingredients 1, including alkaline fibroblast growth factor, vascular endothelial growth factor, platelet-derived growth factor, and other growth factors, after the basal layer of the amniotic membrane has been separated and decellularized12. These characteristics make the acellular amniotic membrane a perfect carrier for stem cell transplantation because it can mimic the ecological environment of stem cells. encourages and promotes cell growth, attachment, and differentiation13,14. Furthermore, the use of amniotic membrane in tissue engineering and the creation of novel therapeutic approaches has grown significantly due to its anti-inflammatory, antimicrobial, antiangiogenic, and low immunogenic properties, as well as its relative ease of accessibility15. Amniotic membrane has been demonstrated to function not only as a physical barrier to prevent adhesions but also to promote Achilles tendon healing in tissue engineering experiments to repair tendons16,17.
While studies have shown that TDSCs have universal stem cell properties like clonal formation, pluripotency, and self-renewal ability and can differentiate into tendon cells and non-tendon cells under suitable conditions, they also play a significant role in tissue homeostasis and tendon repair as emerging seed cells. It is crucial for maintaining tendon stability and self-healing following injury18,19. After in vitro amplification and in vivo transplantation, isolated TDSCs produce tendon-like tissue, which is advantageous for Achilles tendon repair20.
In order to prepare a biomimetic peritendinous membrane material superior to amniotic membrane, an effective combination of decellularised amniotic membrane and TDSCs was performed in this study. The repair effect of biomimetic peritendinous membrane on acute Achilles tendon rupture was studied by means of gross healing score, tissue morphology analysis and biomechanical property test. In addition, transcriptomic analyses at the mRNA level in vitro combined with proteomic analyses of the Achilles tendon in vivo were used to tap into the repair mechanisms of the bionic peritendinous membrane material.
Materials and methods
Statement of ethics
The acquisition of amniotic membrane has been approved by the Ethics Committee of the First Affiliated Hospital of Ningbo University (ethical approval number: KS202211017). The Animal Research Ethics Committee of Ningbo University in China gave its approval for the animal experiments (ethical approval number: 13605). All experiments and methods were conducted in compliance with the Animal Research: Reporting of In vivo Experiments (ARRIVE) guidelines. All procedures were confirmed to comply with the relevant guidelines and regulations.
Preparation of decellularized amniotic membranes scaffolds
The study prospectively recruited participants who had recently given birth at the Affiliated Hospital of Ningbo University Medical School, and the recruitment period for the study began on November 23, 2022, and ended on November 23, 2023. All recruited participants provided written informed consent. It is worth noting that the participants in this study did not include minors. Fresh placentas were obtained from participants following childbirth. The AM were tested to exclude the possibility of carrying infectious diseases such as human immunodeficiency virus (HIV), hepatitis B virus (HBV), hepatitis C virus (HCV), chlamydia, syphilis, cytomegalovirus and other infections. Decellularized amniotic membranes (dAM) were prepared according to the method used in the previous experiments21. In brief, fresh amniotic membranes of fetal placental origin were first washed three times with 75% medical alcohol (E299578, aladdin, shanghai, China) and phosphate-buffered solution (P1010, Solarbio, Beijing, China) washed three times to remove residual stains and blood clots from the amniotic membranes, then the chorioallantoic membrane and basal metaplasia were bluntly separated from the amniotic membranes with a scalpel handle, and the amniotic membranes were washed again with PBS and placed in a centrifuge tube containing solution of penicillin and streptomycin (Pen-Strep) (200 U mL-1 in PBS) in a centrifuge tube and placed on a shaker for 48 h, changing the solution every 12 h. Afterwards, they were then submerged in Triton X-100 solution for 4 h at 37 degrees Celsius, rinsed with Pen-Strep solution submerged in trypsin solution for 10 h, and then obtained the dAM by submerging in DNA enzyme solution for 4 h and rinsing with PBS. Following a 72-hour freeze-drying process, the dAM was placed in a vacuum seal and kept in a refrigerator at 4 °C for further experiments.
Evaluation of decellularized amniotic membranes
Fresh and decellularized amniotic membranes were preserved by submerging them in 4% araformaldehyde, followed by dehydration in varying concentrations of alcohol (70, 80, 90, and 100%) and xylene, embedding them in paraffin, and cutting them into 5-mm-thick sections. Lastly, they were stained with hematoxylin and eosin dye for histopathological evaluation. The microstructure of the freeze-dried, decellularized amniotic membranes was examined using scanning electron microscopy (SEM, Hitachi TM3000, Japan). Cells are lysed using proteinase K followed by a mixture of Tris-HCl, ethylenediaminetetraacetic acid, sodium lauryl sulfate, and NaCl to extract DNA from the lysate. GenEluteTM Gel Extraction Kit (NA1111,Sigma, USA) is used to further purify the DNA that was obtained in the first step. A microUV-Vis spectrophotometer (NanoDropTM One, Thermo, USA) was used to measure and analyze the DNA concentrations in various sets of samples.
Isolation and culture of TDSCs
According to the previous method of our group22, TDSCs were cultured by isolation from the Achilles tendon of SD rats. The rats were anesthetized by injecting 2% (w/w) pentobarbital (30 mg/kg) into the peritoneal cavity, and the Achilles tendon was isolated in a sterile environment. Other tissues that remain in the periphery were removed, and the Achilles tendon was washed three times with PBS. Then the Achilles tendon was cut into fine tissue fragments in PBS solution using scissors and forceps, and the mixed solution was centrifuged (1000 r/min, 3 min), and the supernatant was aspirated, followed by the addition of complete medium diluted with 0.3% Collagenase type I (C8140, Solarbio, Beijing, China) in a cell culture incubator at 5% CO2, 20% O2, 37 °C, and 95% (referred to as the incubation environment) for 4 h, during which the centrifuge tubes were shaken for 1 min every 20 min. Afterwards, the digestion was terminated by adding an equal volume of low glucose Dulbecco’s modified Eagle medium with 10% fetal bovine serum (C0232, Beyotime Biotechnology, Shanghai, China), penicillin–streptomycin (B540734, Sangon Biotech Shanghai, China), and the mixed solution was filtered through a sieve (pore size: 70 μm) (#352350, Corning Falcon, USA). The supernatant was aspirated, and the cells were resuspended with low glucose medium and centrifuged (1500 r/min, 5 min). The cells were inoculated on culture dishes in a cell culture incubator at the incubation environment, and the medium was changed every 3 days.
Viability, proliferation, adhesion and differentiation of tendon derived stem cells on dAM
Cell viability and proliferation
The viability of TDSCs was assessed by culturing them in extracts with dAM as the test group and without dAM as the comparison group. A live/dead assay (PF00008, Proteintech, China) was carried out at day 1, 3, 5 and 7 after the cells were cultured in the extract. Following aspiration, the TDSCs were briefly washed by PBS liquor and 4% paraformaldehyde liquor (LJ61899500ML, Sinopharm, Shanghai, China), and 5 mL of normal saline was combined with Ethidium homodimer-I and Calcein AM. Staining the cells with 200 L dye solution, incubating them at 37 °C for 20 min, and taking pictures was achieved with an Olympus IX73 inverted fluorescence microscope.
The proliferation of cells on dAM was examined by CCK-8(GK10001, glpbio, shanghai, China). Freeze-dried decellularized amniotic membrane was cut into 1 unit area for experimental group, using a circular size of 7 mm in diameter (just covering the bottom of the 96-well plate) as 1 unit area. In the control group, 7 mm wafer prepared by type I collagen were used. All the wafers of the control group and the experimental group were placed in 96 - well plates and soaked in low-sugar medium for 24 h. The next day, 1 × 103 numbers of TDSCs were inoculated into each well and incubated in a cell culture incubator at the incubation environment for 1 day. The CCK − 8 reaction solution was incubated at 37 °C for 2 h. The reaction solution was aspirated and placed in the other 96 - well plate, and using an enzyme standard analyzer, the reaction solution’s absorbance at 450 nm was determined in each well. The residual test solution in the 96 - well plate was washed off with PBS solution. The cell proliferation were tested at 1, 3, 5, and 7 days by the same method.
Cell adhesion
Cells at a concentration of 1 × 10^3 cells/mL were placed on a dAM scaffold. Following incubation at a temperature of 37 °C for durations of 1 and 3 days, 4% paraformaldehyde was used to fix them. Then, they were stained by COL1 A1 (A16891, Abclonal, Wuhan, China).
Cell differentiation
A 96-well culture plate was seeded with cells at a density of 5 × 10^4 cells per milliliter. A new hole was drilled with the dAM the next day, where the cells would be further cultured. Hole groups with dAM were experimental, while hole groups with COL1 were control. In the three and seven day experiments, Scx, Tnc and Tnmd were measured by using qRT-PCR, and β-actin served as the housekeeping gene. Refer to Table 1 for the PCR primer sequences.
Primer sequences were given in 5’ to 3’ direction.
Analysis and sequencing of RNA data
During the assays, TRIzol reagent facilitated the extraction of total RNA from TDSCs and dAM & TDSCs (ET111-01-V2, transgen, Beijing, China). Following the instructions on the manufacturer’s website, paired-end libraries were prepared with the ABclonal mRNA-seq Lib Prep Kit (RK20350, ABclonal, China). In addition to purifying the PCR products with an AMPure XP system, the quality of the libraries was assessed on an Agilent Bioanalyzer 4150. In the final step, Illumina Novaseq 6000/MGISEQ-T7 was used to perform the sequencing. DESeq2 was used to analyze differential gene expression, identifying genes with expression changes over 1 and Padj values above 0.05 as significant. We utilized the cluster Profiler R software package for our analysis of GO function and KEGG pathway.
Assessments of in vivo repair
Animal experiment
In this experiment, 72 SD rats (age 8–10 weeks, weight 350 ± 50 g) were randomly divided into 3 groups of 24 rats each (n = 24). 2% (w/w) pentobarbital (30 mg/kg) was injected into the rat’s abdominal cavity, and the right lower limbs were treated during the experiment. Firstly, the hair on the right lower limbs were shaved, and a scalpel was used to make a longitudinal incision of about 20 mm along the long axis of the Achilles tendon to dissect and separate the Achilles tendon. The Achilles tendon was cut transversely at 5 mm proximal to the heel bone, and twenty four of the rats (n = 24) were sutured back in place by the Kessler method (control group, group C). Twenty four rats (n = 24) were sutured in the same way, and later wrapped in decellularized amniotic membrane and fixed with fine thread at both ends. The freeze-dried decellularized amniotic membranes were soaked in DMEM medium for 24 h in advance (amniotic membrane group, group H). The remaining twenty four rats (n = 24) were treated in the same way as group C, and then wrapped in decellularized amniotic membrane that had been inoculated with TDSCs (5 × 104 cells/mL) and fixed with fine wires at both ends (tendon derived stem cell group, group T). After the above operations were completed, the incisions were closed layer by layer in the same way, disinfected, wrapped with gauze and fixed in plaster for 1 week. Following surgery, the rats were housed in an environment with a 24–25 °C temperature and had unlimited from water and meals. Eight rats (n = 8) from each of the three groups were executed at 4, 8, and 12 weeks under overdose anesthesia, respectively. Rat Achilles tendon specimens were dissected and four rats (n = 4) were used for gross observation and histological analysis, while the other four rats were used for biomechanical analysis. Achilles tendon samples from week 12 (control group and dAM + TDSCs group) were immersed in RNAlater (AM7020, Solarbio, Beijing, China) for proteomic detection after mechanical testing.
Macroscopic analysis
All rat Achilles tendons were invited by three observers before being used for histological and biomechanical analyses. Healing of the Achilles tendon was observed visually without knowing the grouping. The injured Achilles tendon was observed for the presence of scar healing, the degree of adhesion to the surrounding tissues, and according to the Yang’s healing score23. (The specific scoring criteria are shown in Table S1)
Histological analysis
Dissected rat Achilles tendons were processed according to a standard procedure. They were first fixed with 4% paraformaldehyde for 12 h and then dehydrated by gradient alcohol (70%, 80%, 90%, and 95% anhydrous ethanol). Then they were immersed in liquid wax and sagittally embedded into wax blocks. Afterwards, the sections were cut into 5 μm-thick sections using a microtome, dewaxed in xylene and rehydrated in gradient alcohol so that they could be used for hematoxylin-eosin staining (HE), Masson’s trichrome reagent staining and immunohistochemical staining.
For immunohistochemical staining, the dewaxed and rehydrated sections were first washed with running water for 5 min and then immersed in a solution of sodium citrate heated to 90 °C for 5 min to repair the antigen. The solution was then heated to reach 90 °C again and the antigen was repaired for 5 min. The above cycle was repeated a total of four times and cooled to 37 °C. Then, the sections were rinsed with PBS solution and immersed in 3% H2O2 for 10 min to seal the endogenous peroxidase. Once completed, the sections were rinsed with PBS again, and the antigen was restored using a trypsin mixture (0.05% trypsin solution combined with 0.1% CaCl2 solution) for an hour at 37 °C. After 3 rinses with PBS solution, the non-specific antigens were continued to be blocked with 10% goat serum for 2 h at 37 °C. Afterwards, rabbit antibodies with 1:200 primary monoclonal antibodies [anti-collagen type I antibody (14695-1-AP, proteintech), anti-Tenomodulin (ab203676, Abcam)] were incubated for 12 h at 4 °C in a refrigerator. At the end of the incubation, the rabbits were rinsed five times with PBS solution. The above primary monoclonal antibody was recognized with fluorescein isothiocyanate (FITC)-labeled sheep anti-rabbit IgG (AS039, ABclonal, Wuhan, China; 1:100, secondary antibody, 37 °C, 2 h). Then it was rinsed again with PBS solution five times and stained with 2% diaminobenzidine (DA1016, Solarbio, Beijing, China) for 1 min. Finally, staining was done with hematoxylin for 3 min and running counter blue for 10 min.
For immunofluorescence staining, the steps of antigen repair were consistent with those of immunohistochemistry. The primary antibody were incubated overnight with phosphorylation-ERK (9298, CST, USA, 1:200). After being rewarmed to 37 °C, secondary antibody specific to rabbit IgG (AS039, ABclonal, Wuhan, China; 1:200) was used to incubate the samples. DAPI (C0060, Solarbio, Beijing, China) was used to stain nuclei.
For adhesion evaluation, three observers were likewise invited to observe microscopically and score the adhesions of Achilles tendon tissue according to Tang’s scoring rules without knowing the grouping24. (The specific scoring criteria are shown in Table S2)
Biomechanical analysis
The tendon tissue at the end of the gastrocnemius and flounder muscles and in the middle of the heel bone was amputated for biomechanical testing. The specimen to be tested should be shaved to remove as much tissue as possible, except for the Achilles tendon tissue. The ends of the specimen are clamped in each of the two clamps of the tensile testing device to ensure clamping. After zeroing the distance and tension, the test is started and the tension data of the Achilles tendon is recorded until the Achilles tendon is completely stretched. Four Achilles tendons in each group were used for biomechanical testing.
Proteomics
We performed proteomic analysis of the control group and dAM + TDSCs Achilles tendon samples at week 12. The samples are derived from the tissue after mechanical testing. Screening for differentially expressed proteins was based on an expression change of more than 1.5 - fold, P < 0.05. For Gene Ontology (GO) annotation, Blast2Go was employed, and the KEGG pathway annotation was carried out with the KEGG automatic annotation server software. Fisher exact test was used to compare the differential protein set and total proteins in the distribution of each GO classification or KEGG pathway and to perform enrichment analysis of GO annotation or KEGG pathway annotation set for differential proteins.
Statistical analysis
The numerical data were presented as mean standard deviation (SD). Student’s t-tests were used to assess the significance of observed differences between the two experimental groups. Multiple groups were compared using one-way analysis of variances (ANOVAs), followed by post hoc Bonferroni comparisons. A P value < 0.05 was considered statistically significant.
Results
Amniotic membrane decellularization is highly effective
The placenta removed from the mother is treated with decellularization and has a generally hyaline membrane-like shape. To identify the effect of decellularization of amniotic membranes, we compared the results of hematoxylin-eosin staining (HE), electron microscopic observation and DNA content measurement of amniotic membranes pre and post decellularization. It was observed that the amnion was mainly covered with epithelial cells and a few fibroblasts before decellularization. After decellularization, these two types of cells were significantly reduced, and the nuclear staining was also significantly reduced (Fig. 1A-B). Under the electron microscope, it was found that there were many round spherical cells arranged in a staggered pattern on the surface of the amnion before decellularization, while after decellularization, the amnion showed an irregular and loose meshwork scaffold structure (Fig. 1C-D). Following transplantation and culture for 1 and 3 days, TDSCs on the decellularized amniotic membrane were fixed and freeze-dried. Fluorescence microscopy revealed cellular adhesion to the amniotic membrane, and the number of cells increased on the third day (Fig. 1E-F). In addition, the TDSCs were observed to be attached to the amniotic membrane under electron microscopy, with multiple tentacles adhering to the reticular scaffold of the amniotic membrane. The amnion was also no longer porous, but was filled with extracellular matrix and showed a smooth dense structure (Fig. 1G). In the results of DNA content measurement, the DNA content was significantly lower after decellularization compared with that before decellularization, and there was a statistically significant difference (Fig. 1H).

Evaluation results of the decellularization effect of fresh amniotic membrane. (A-B) Amniotic tissue before and after decellularization HE staining observed under a microscope. (C-D) Amniotic tissue before and after decellularization observed under electron microscopy. (E-F) Morphology and distribution of TDSCs on the decellularized amniotic membrane observed via fluorescence microscopy. (G) Observation of TDSCs on decellularized amniotic membrane under scanning electron microscope. (H) Comparison of DNA content before and after decellularization of amniotic tissue. Bar: 50 μm. Statistical significance: ***P < 0.001.
dAM facilitated cell viability, proliferation and tenogenic differentiation of TDSCs
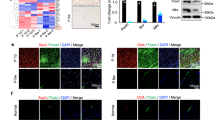
As shown in Fig. 2A, the TDSCs treated with dAM extract exhibited high viability on days 1, 3, 5 and 7. Living cells are marked in green, and dead cells are marked in red. CCK8 detection showed that dAM had a significant proliferative effect on TDSCs at day 5, and the trend decreased at day 7 (Fig. 2B). Compared to the Control group, Scx, Tnc, and Tnmd expression levels were higher in dAM loaded with TDSCs compared to Control (Fig. 2C).

dAM promoted TDSCs proliferation and the expression of tenogenic differentiation and extracellular matrix related genes. (A) A live/dead staining was performed on TDSCs in extracts of dAM at days 1, 3, 5 and 7. Calcein AM (green) and EthD-I (red) were used to identify live and dead cells. (B) The effect of dAM on TDSCs activity was detected by CCK8. (C) Scx, Tnc, and Tnmd expression levels in dAM loaded with TDSCs were higher than control group. Bar: 200 μm. Statistical significance: **P < 0.01, ***P < 0.001.
Microarray profiling of mRNA expressions linked to the extracellular matrix
In order to explore the possible molecular mechanisms underlying extracellular matrix-induced Achilles tendon repair, we performed mRNA microarray analyses of TDSCs-dAM. Comparing the Control group with the dAM group, there was a disparity in expression levels using Principal Component Analysis (PCA) (Fig. 3A). Using the volcano map, we were able to visualize the distribution of DEGs and the differences in expression regulation between the two groups (Fig. 3B). Significance of KEGG enrichment was calculated using Fisher’s exact test to determine signaling pathways associated with dAM groups using the KEGG analysis. The bubble scatter diagram in (Fig. 3C) shows the top 20 enriched KEGG pathways of DEGs, with ECM-receptor interaction being the most influential pathway. In addition, Focal adhesion is also involved in the function of TDSCs in the dAM group. There was also an analysis of DEGs associated with ECM-receptor interaction. In Fig. 3D, the top 15 DEGs with the greatest P values are presented, revealing that LAMC2, TNR, IBSP, TNN, HMMR, and SDC4 had elevated expression levels relative to the control group.

Microarray and bioinformatics analyses were used to examine differentially expressed genes regulated by dAM. Microarray analysis was conducted after 7 days of growing TDSCs in Petri dishes and dAM. (A) Two-dimensional representation of principal component analysis (PCA). (B) On the volcano map, blue dots highlight significantly down-regulated genes, red dots highlight significantly up-regulated genes, and grey dots indicate genes with no significant variation. (C) Enrichment analysis of differential genes, as well as essential downstream genes, from the Kyoto Encyclopedia of Genes and Genomes (KEGG). The right side of each bar represents the number of differential genes in the pathway. (D) In this chart, the Top 15 differential genes associated with ECM-receptor interaction are ranked with P value. Red genes are up-regulated; green genes are down-regulated.
Figure 3C was generated based on the KEGG pathway map25,26,27.
dAM-TDSCs promote healing of ruptured Achilles tendon
Analysis of hematoxylin-eosin (HE) staining results
In the HE-stained tissue sections, we can see that at 4 weeks, inflammatory cell infiltration was visible in all tissue sections. There were significant clusters of inflammatory cell aggregates with more neovascularization in group C and relatively less in groups H and T. In terms of healing evaluation, group C had a small amount of granulation tissue filling the defect in the area of Achilles tendon dissection, but still had discontinuous collagen fibers. However, in groups H and T, there was more granulation tissue filling than in group C, making the Achilles tendon injury area largely continuous. At 8 weeks, the healing of the Achilles tendon tissue was significantly better than at 4 weeks. The dissected area of the Achilles tendon was filled with tissue and achieved tissue continuity in all groups, but the extracellular matrix was less filled. Group C and H formed a disorganized arrangement of collagen fibers, whereas group T had a relatively regular arrangement of collagen fibers, and only group C still had scattered inflammatory cells. At 12 weeks, the inflammatory cells in group C disappeared, and a few blood vessels were still present. Disordered collagen fibrils were formed in the tissue, and the extracellular matrix was not tightly filled. Compared to group C, groups H and T had better filling of extracellular matrix. In addition, the group T had a tighter, more oriented collagen fiber arrangement than the group H, which was closer to the natural state of the Achilles tendon (Fig. 4A).
Analysis of Masson staining results
Masson staining can colorize collagen fibers and can assist us in better assessing Achilles tendon healing. At 4 weeks, all groups had sparser, discontinuous and disorganized collagen fibers. There was a scattered distribution of blood vessels, with relatively more collagen fibers in group T. At 8 weeks, the number of blood vessels in the tissue decreased and the collagen fiber staining became darker than before. There were large waves of collagen fiber formation filling the isolated area. Compared to group C, groups H and T were more filled, and group T had some degree of directional alignment compared to the remaining two groups. By 12 weeks, collagen production had increased in both groups, and group C still had a scattered distribution of blood vessels. The collagen fibrils were not oriented and the extracellular matrix was less filled, while the collagen fibril content and extracellular matrix filling were similar in the group H and T. But the group T had fewer cells in the more regularly arranged collagen fibrils (Fig. 4B).
Histologic scores of dAM-TDSCs were superior to those of other groups
In the microscopic tissue adhesion scores, higher adhesion degree scores were obtained in group C compared to group T at 4 and 8 weeks. Conversely, no notable difference was observed between group H and group T. At 12 weeks, although the T group score was further reduced, it was not statistically significant compared to the H and C groups (Fig. 4C). The healing scores from weeks 8 to 12 revealed that group T experienced superior healing compared to group C. No noticeable difference was found between group C and group H (Fig. 4D).
dAM + TDSCs group promoted the high expression of extracellular matrix protein in Achilles tendon
In immunohistochemical staining, type I collagen staining antibodies were darker over time in all groups, suggesting a gradual increase in type I collagen content. In contrast, group C stained lighter than the experimental group, while no significant difference was found between them (Fig. 4E). In the study of TNMD expression of genes related to Achilles tendon reconstruction, it was found that there was no significant difference in TNMD expression between the three groups at 4 weeks, but at 8 weeks and 12 weeks, the expression in T group was better than that of the other two groups (Fig. 4F).
dAM-TDSCs enhance Achilles tendon mechanical properties
In the evaluation of maximal load, all groups showed an increase in maximal load with increasing repair time. At 4 weeks, there was no difference in maximum load between three groups. At the 8-week mark, group C differed significantly in maximum load from groups H and T, which did not show any difference between themselves. Group T had a statistically significant difference with groups H and C at the 12-week mark, and groups C and H also differed significantly (Fig. 4G). In the test of elastic modulus at 4 weeks, groups H and T had comparable test values and were greater than group C. There was a distinct upward trend in groups H and T at 8 weeks, yet no statistical significance was found among the groups. By 12 weeks, group T showed an improved elastic modulus over groups H and C (Fig. 4H).

dAM-TDSCs promote healing of ruptured Achilles tendon. (A) Histological evaluation of Achilles tendon healing via HE staining. (B) Assessment of healing at Achilles tendon ruptures by Mason staining. (C) Adhesion score assessed by microscopic histological observation. (D) Healing score assessed by macroscopic observation. (E - F) Extracellular matrix-related factors, COL1 A1 and TNMD are assessed after treated with dAM-TDSC scaffolds 4 w, 8 w, and 12w. (G) The maximum load of 3 sets of Achilles tendons at 3-time nodes was obtained by stretching. (H) Schematic diagram of the elastic modulus of Achilles tendons in three groups at 3-time nodes. Bar: 50 μm. *: P < 0.05; **: P < 0.01; ***: P < 0.005.
Proteomic analysis
As shown in the Wayne diagram comparing the control and dAM + TDSC groups, there were 130 proteins specific to the control group 238 proteins specific to the dAM + TDSCs group for a total of 2452 proteins (Fig. 5A). The volcano plot revealed that the dAM + TDSCs group was found to be associated with elevated expression of proteins promoting protein synthesis and processing, and translocation when compared to control, including Col12a1, Xpo4, Tnc, Rpl22, Rps8, Rps9, Rpl7a, Rpl13, Mrps27, etc. (Fig. 5B). The top twenty GO functions showed that compared to the control group, the regulation of vascular permeability, transforming growth factor-beta secretion, ribosomal large subunit assembly and other signaling pathways related to vascular protein synthesis, processing and transport were activated (Fig. 5C). The KEGG map revealed that the two groups of differential proteins were involved in metabolic pathways such as Ribosome, ECM-receptor interaction, and Focal adhesion (Fig. 5D). GSEA showed that ribosome and protein processing, extracellular matrix receptor activity was boosted in the dAM + TDSCs group, while Tight junction, Focal adhesion signaling pathway was enhanced and HIF-1 signaling pathway was inhibited. This suggests that dAM + TDSCs may promote protein and extracellular matrix synthesis, processing, cell junction, inhibit inflammatory factor expression and pathological neovascularization within adhesion tissue, thereby promoting structured tendon repair (Fig. 5E-J).

Proteomic analysis of dAM + TDSCs group involved in the repair of Achilles tendon rupture. (A) Proteins between control and dAM + TDSCs groups Venn diagrams. (B) Volcano plot of differentially expressed proteins between dAM + TDSCs and control. In order to show the significant difference of proteins between the comparison groups, the significantly down-regulated proteins are marked in blue (FC < 1.5 and p < 0.05), the significantly up-regulated proteins are marked in red (FC > 1.5 and p < 0.05), and the proteins with no difference are marked in gray. (C) The differentially expressed proteins in the control and dAM + TDSCs groups were compared with all the proteins in the reference species by GO function annotation, and the resulting bubble plot was obtained. (D) KEGG pathway annotation and attribution bar chart of differentially expressed proteins between control and dAM + TDSCs groups. The horizontal axis represents the number of pathway protein annotations, and the vertical axis represents the KEGG annotation name. Different colors represent the seven branches of KEGG metabolic pathways. (E) GSEA plot of Ribosome. (F) GSEA plot of Protein processing in endoplasmic reticulum. (G) GSEA plot of ECM-receptor Interaction. (H) GSEA plot of Tight junction. (I) GSEA plot of Focal adhesion. (J) GSEA plot of HIF-1 signaling pathway.
Figure 5D was generated based on the KEGG pathway map25,26,27.
ERK signaling pathway is involved in the repair of Achilles tendon fracture
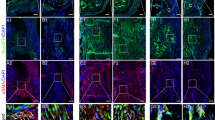
In the dAM group, p-ERK began to express at week 8. In dAM + TDSCs, p-ERK was expressed at the 4 th week, and this abundance was most obvious at the 12 th week. The results confirmed that the Biomimetic peritendinous membrane materials constructed by dAM-TDSCs enhanced Achilles tendon regeneration in vivo via activation of the ERK signaling pathway (Fig. 6A-B).

(A) Immunological staining of p-ERK in the damaged area at 4, 8 and 12 weeks. p-ERK was detected by IF staining (red). (B) Quantitative fluorescence analysis.
Discussion
The key discovery of this research was the application of TDSCs on a human-derived acellular amniotic membrane to treat acute Achilles tendon rupture. The human acellular amniotic membrane loaded with TDSCs not only promoted the adhesion, proliferation and early differentiation of tendon derived stem cells, but also promoted the early secretion of cell stroma related proteins from TDSCs, which additionally quickened the initial repair of a ruptured Achilles tendon.
A persistent medical problem is the investigation of how the body heals after an Achilles tendon rupture. According to recent research, the extracellular matrix of the Achilles tendon tissue is primarily a flat, round, columnar structure made of numerous parallel collagen fibers, and there is a layer of paratendon tissue surrounding the Achilles tendon rather than a true synovial membrane. The majority of the blood supply to the Achilles tendon is provided by this highly vascularized, monolayer, fatty reticular tissue called the paratendon28. In diseased tendon tissue, studies have discovered elevated cell counts, collagen confusion, and neovascularization. Additionally, abnormal changes in collagen fiber diameter, longitudinal division of collagen fibers, and collagen fiber decomposition can all be signs of abnormal collagen29,30. Achilles tendon repair is divided into two main pathways: endogenous repair and exogenous repair31. Endogenous repair plays a smaller role in repair because the number of resident Achilles tendon cells associated with endogenous repair is smaller and the repair effect is slower. The internal and external barriers produced by trauma disrupt the recruitment of wound surface epidermal cells and immune cells, and the involvement of exogenous repair is an important cause of fibrotic scarring and adhesion formation32. Therefore, inhibition of exogenous repair and promotion of endogenous repair can be effective in preventing adhesion healing, which is one of the generally favored views.
The use of sheet materials as a physical barrier separation can significantly inhibit exogenous repair. The materials commonly used today are divided into synthetic materials (polyethylene film, silicone film, cellophane) and natural biological materials (tissue around tendons, veins and fascial grafts)33. Both techniques can detach tissues from adhesions, but the former raises the likelihood of tendon injury and enduring foreign body residue due to its impermeability, which hinders normal nutrient delivery34. Therefore, resorbable biomaterials have become a hot research topic. The primary concern in developing a cell scaffold is that the various pore sizes within the porous structure must be biocompatible and elicit minimal immune response; Degradation should occur at the same rate as tendon tissue regeneration, and the resulting products must be harmless9. Under electron microscopy, the freeze-dried decellularized amniotic membrane is a lamellar mesh scaffold. It has good biocompatibility, permeability and degradability, and can serve as a physical obstruction, separating Achilles tendon tissue from the wound surface. Not only that, the rich growth factors contained in the extracellular matrix of the amniotic membrane are also retained after decellularization to promote the repair of Achilles tendon tissue35,36. The study by Nicodemo et al. demonstrated that wrapping the ruptured Achilles tendon with a decellularized amniotic membrane promoted tissue repair and lessened the inflammatory response37. Manti et al. also concluded after experiments that the amniotic membrane can successfully prevent the Achilles tendon and surrounding tissues from adhesion healing38. Also, they responded that the amniotic membrane is thinner, has low mechanical strength, and is more prone to tearing. Liu et al. combined synthetic material PCL nanofibers with amniotic membrane to strengthen the mechanical strength of the amniotic membrane. It was also found to be effective in isolating exogenous adherent tissue and promoting endogenous tendon healing39. Bi et al. made the initial discovery of TDSCs in humans and mice as emerging seed cells40, and it possess properties of universal stem cells. Tendon cells have a strong capacity to differentiate into TDSCs, and TNMD is a crucial regulator of tendon cell proliferation and takes part in collagen fiber maturation41. Additionally, it has characteristics of universal stem cells. Tendon cells have a potent ability to differentiate into TDSCs, and TNMD plays an important role in collagen fiber maturation and regulation of tendon cell proliferation19. The feasibility of implanting TDSCs tablets on tendons in vivo was confirmed by Komatsu I et al. after they prepared TDSCs tablets experimentally. At 2 and 4 weeks after implantation, the introduction of cell tablets into tendon defects significantly improved histological characteristics and collagen content, indicating that TDSCs tablets are effective in promoting tendon remodeling in the early stages of tendon healing42.
The experiments were designed to avoid adulteration with synthetic materials, thus having no impact on the degradability of the amniotic membrane. Furthermore, TDSCs were cultured on the amniotic membrane to further enhance the endogenous growth pathway of the amniotic membrane. Specifically, it was observed that the dAM promoted the better adhesion and spread of TDSCs in immunofluorescence and scanning electron microscopy, and CCK-8 experiment confirmed that the dAM promoted the proliferation of TDSCs. RT-qPCR analysis confirmed that amniotic membrane can promote the early differentiation of TDSCs and the expression of extracellular matrix. In the animal experiments, the adhesion score of group T was observed to be the smallest in both macroscopic and histological scores, which suggested that the amniotic membrane might effectively serve as a physical border to block the effects of exogenous repair and avoid adhesion formation. In the histological observation, we found that the healing rate of the Achilles tendon tissue rupture was faster in the group T with increasing time. The tissue fibers were more regularly aligned and the extracellular matrix filling was fuller. And in Masson staining, it could be found that the group T was richer in collagen compared to the other two groups. This trend was also reflected in the histological scores and was statistically significant. In immunohistochemical staining, type I collagen staining increased in depth with time, and the groups T and H did not differ from one another. which was better than the group C. The expression of extracellular matrix related protein TNMD was excellent in group T, and the abundance was higher than that in group C and group H, which meant that the synthesis of extracellular matrix was increased and could promote the early healing of Achilles tendon. In addition, at 12 weeks, the maximum load and elastic modulus of the group T were better than the other two groups, which indicated that the group T could effectively promote tissue healing.
In this experiment, dAM group provided an excellent growth environment for TDSCs, which was mainly manifested in transcriptome sequencing analysis at the mRNA level, which showed that extracellular matrix receptor interaction and Focal adhesion ranked 1 st and 8 th in KEGG in dAM group, respectively. Extracellular strom-related proteins LAMC2, TNR, IBSP, TNN, HMMR and SDC4 were increased in dAM group, which was significantly better than that in control group. Proteomic analysis of Achilles tendon samples in animal studies showed that regulation of vascular permeability, transforming growth factor-beta secretion and ribosomal large subunit assembly involved in the repair of a broken Achilles tendon, Ribosome, ECM-receptor interaction and Focal adhesion signaling pathways are also involved in the regeneration of ruptured Achilles tendon. These results were consistent with transcriptome sequencing at the mRNA level in vitro. As mitogens, growth factors, and cytokines are included in the extracellular stimuli that activate the ERK1/2 signaling pathway, it is capable of responding to a wide range of external stimuli43. Injured muscles were found to regenerate after activation of ERK1/2 by Hong et al.44. A pathway involving RAS and ERK controls cell growth, proliferation, regeneration, and differentiation, according to Trevor et al.45. In this experiment, p-ERK was expressed in dAM-TDSCs with the highest abundance. While transcriptomic and proteomic analyses highlighted ECM-receptor interaction as the predominant pathway regulated by dAM, the observed phosphorylation of ERK suggests a secondary role for ERK signaling in TDSCs differentiation. It is hypothesized that the collagen-rich scaffold of dAM engages integrin receptors (e.g., α2β1), triggering FAK/Src-mediated activation of the RAS-MAPK-ERK cascade. This pathway may amplify ECM-driven tenogenic differentiation by enhancing transcriptional activity of scleraxis (Scx) and tenomodulin (Tnmd). However, the precise hierarchy between ECM signaling and ERK activation requires further validation. Future studies employing integrin inhibitors (e.g., RGD peptides) will delineate whether ERK activation is mechanistically dependent on ECM-cell interactions.
Conclusions
In this experiment, the aponeurotic biomaterials were constructed by loading TDSCs on lyophilized decellularized amniotic membrane to repair the rat model of acute Achilles tendon rupture. Histological healing and mechanical recovery were superior to decellularized amniotic membrane. In conclusion, the decellularized amniotic biomimetic peritendinous membrane material loaded with TDSCs is conducive to promoting the healing of Achilles tendon and effectively preventing the adhesion of Achilles tendon. This study provides a new approach to the clinical challenges in the repair of acute Achilles tendon rupture. However, there are some limitations to this study; first, it is difficult to track the implanted tendon stem cells in vivo, and there is no way to know their exact fate in vivo. Second, the degradation of decellularized amniotic membrane in vivo has not been thoroughly studied. Third, although ERK phosphorylation was observed, its causal relationship with ECM signaling remains unproven. Fourth, the temporal sequence between ERK activation and ECM remodeling has not been fully determined. In the future, we will do further research and report on the above limitations.
Data availability
On reasonable request, the corresponding author will provide the datasets used and analyzed in this study.
References
Zielinska, N., LaPrade, R. F. & Olewnik, Ł. Morphological variations of the calcaneal tendon: clinical significance. J. Orthop. Surg, Res. 18 (1), 275 (2023).
Steinmann, S., Pfeifer, C. G., Brochhausen, C. & Docheva, D. Spectrum of tendon pathologies: triggers, trails and End-State. Int. J. Mol. Sci. 21(3), 844 (2020).
Teng, Z. L. et al. Epidemiological characteristics of patients operated for Achilles tendon rupture in Shanghai. Orthop. Surg. 14 (8), 1649–1655 (2022).
Wilder, J. H. et al. Rates of operative management for Achilles tendon rupture over the last decade and the influence of gender and age. Foot Ankle Spec. 16 (4), 377–383 (2023).
Grinstein, M. et al. A distinct transition from cell growth to physiological homeostasis in the tendon. eLife. 8, e48689 (2019).
Reda, Y., Farouk, A., Abdelmonem, I., El, O. A. & Shazly Surgical versus non-surgical treatment for acute Achilles’ tendon rupture. A systematic review of literature and meta-analysis, foot and ankle surgery: official. J. Eur. Soc. Foot Ankle Surg. 26 (3), 280–288 (2020).
Yang, X. et al. Construction of microunits by Adipose-Derived mesenchymal stem cells laden with porous microcryogels for repairing an acute Achilles tendon rupture in a rat model. Int. J. Nanomed. 15, 7155–7171 (2020).
Wu, R. et al. Bioactive silk Fibroin-Based hybrid biomaterials for musculoskeletal engineering: recent progress and perspectives. ACS Appl. Bio Mater. 4 (9), 6630–6646 (2021).
Liu, Q. W. et al. Characteristics and therapeutic potential of human Amnion-Derived stem cells. Int. J. Mol. Sci. 22(2), 970 (2021).
Dikmetas, O., Kapucu, Y., Cankaya, A. B. & Kocabeyoglu, S. Outcomes and success of amniotic membrane transplantation for the treatment of corneal diseases. Cutan. Ocul. Toxicol. 43 (1), 33–37 (2024).
Dua, H. S., Gomes, J. A., King, A. J. & Maharajan, V. S. The amniotic membrane in ophthalmology. Surv. Ophthalmol. 49 (1), 51–77 (2004).
Sang, R., Liu, Y., Kong, L., Qian, L. & Liu, C. Effect of acellular Amnion with increased TGF-β and bFGF levels on the biological behavior of tenocytes. Front. Bioeng. Biotechnol. 8, 446 (2020).
Gholami, K. et al. Decellularized amniotic membrane hydrogel promotes mesenchymal stem cell differentiation into smooth muscle cells. FASEB Journal: Official Publication Federation Am. Soc. Experimental Biology. 38 (16), e70004 (2024).
Chen, P. et al. Human amniotic membrane as a delivery vehicle for stem cell-based therapies. Life Sci. 272, 119157 (2021).
Jirsova, K. & Jones, G. L. A. Amniotic membrane in ophthalmology: properties, preparation, storage and indications for grafting-a review. Cell. Tissue Bank. 18 (2), 193–204 (2017).
Nicodemo, M. C., Arisawa, E. & Sant’anna, L. B. Lopes-Martins, photobiomodulation and amniotic membrane for treat tendon injury in rats. Anais Da Acad. Brasileira De Ciencias. 96 (suppl 1), e20231139 (2024).
Giacobazzi, M. et al. Primary repair of complete Achilles tear augmented with Amnion allograft wrap in college basketball player with a history of contralateral Achilles rupture: a case report. AME Case Rep. 7, 36 (2023).
Bi, Y. et al. Identification of tendon stem/progenitor cells and the role of the extracellular matrix in their niche. Nat. Med. 13 (10), 1219–1227 (2007).
Zhang, J. & Wang, J. H. The effects of mechanical loading on tendons–an in vivo and in vitro model study. PloS One. 8 (8), e71740 (2013).
Ni, M. et al. Tendon-derived stem cells (TDSCs) promote tendon repair in a rat patellar tendon window defect model. J. Orthop. Research: Official Publication Orthop. Res. Soc. 30 (4), 613–619 (2012).
Luo, Y. et al. Various administration forms of decellularized amniotic membrane extract towards improving corneal repair. J. Mater. Chem. B. 9 (45), 9347–9357 (2021).
Mao, X. et al. Enhancement of tendon repair using tendon-Derived stem cells in small intestinal submucosa via M2 macrophage polarization. Cells 11(17), 2770 (2022).
Yang, D. J., Chen, F., Xiong, Z. C., Xiong, C. D. & Wang, Y. Z. Tissue anti-adhesion potential of biodegradable PELA electrospun membranes. Acta Biomater. 5 (7), 2467–2474 (2009).
Tang, J. B., Shi, D. & Zhang, Q. G. Biomechanical and histologic evaluation of tendon sheath management. J. Hand. Surg. 21 (5), 900–908 (1996).
Kanehisa, M., Furumichi, M., Sato, Y., Matsuura, Y. & Ishiguro-Watanabe, M. KEGG: biological systems database as a model of the real world. Nucleic Acids Res. 53 (D1), D672–d677 (2025).
Kanehisa, M. Toward Understanding the origin and evolution of cellular organisms. Protein Science: Publication Protein Soc. 28 (11), 1947–1951 (2019).
Kanehisa, M. & Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28 (1), 27–30 (2000).
Sharma, P. & Maffulli, N. Tendon injury and tendinopathy: healing and repair, the journal of bone and joint surgery. Am. Volume. 87 (1), 187–202 (2005).
Okewunmi, J., Guzman, J. & Vulcano, E. Achilles tendinosis Injuries-Tendinosis to rupture (Getting the athlete back to Play). Clin. Sports Med. 39 (4), 877–891 (2020).
Riley, G. Tendinopathy–from basic science to treatment, nature clinical practice. Rheumatology 4 (2), 82–89 (2008).
Alimohammadi, M. et al. Mousavi Shaegh, electrospun nanofibrous membranes for preventing tendon adhesion. ACS Biomaterials Sci. Eng. 6 (8), 4356–4376 (2020).
Snedeker, J. G. & Foolen, J. Tendon injury and repair - A perspective on the basic mechanisms of tendon disease and future clinical therapy. Acta Biomater. 63, 18–36 (2017).
Hou, J. et al. Biomaterials strategies to balance inflammation and tenogenesis for tendon repair. Acta Biomater. 130, 1–16 (2021).
Liu, C., Yu, K., Bai, J., Tian, D. & Liu, G. Experimental study of tendon sheath repair via decellularized Amnion to prevent tendon adhesion. PloS One. 13 (10), e0205811 (2018).
Fénelon, M. et al. Applications of Human Amniotic Membrane for Tissue Engineering, Membranes 11(6), 387 (2021).
Duerr, R. A., Ackermann, J. & Gomoll, A. H. Amniotic-Derived treatments and formulations. Clin. Sports Med. 38 (1), 45–59 (2019).
Nicodemo, M. C. et al. Amniotic membrane as an option for treatment of acute Achilles tendon injury in rats. Acta Cirurgica Brasileira. 32 (2), 125–139 (2017).
Manti, N., Guvercin, Y., Mercantepe, T., Tumkaya, L. & Balik, M. S. Clinical and histologic evaluation of partial Achilles tendon injury repair with amniotic membrane in rats. J. Am. Podiatr. Med. Assoc. 112(1), 20-055 (2022).
Liu, C. et al. Regulation of ERK1/2 and SMAD2/3 pathways by using Multi-Layered electrospun PCL-Amnion nanofibrous membranes for the prevention of Post-Surgical tendon adhesion. Int. J. Nanomed. 15, 927–942 (2020).
Zhang, J. & Wang, J. H. Mechanobiological response of tendon stem cells: implications of tendon homeostasis and pathogenesis of tendinopathy. J. Orthop. Research: Official Publication Orthop. Res. Soc. 28 (5), 639–643 (2010).
Docheva, D., Hunziker, E. B., Fässler, R. & Brandau, O. Tenomodulin is necessary for tenocyte proliferation and tendon maturation. Mol. Cell. Biol. 25 (2), 699–705 (2005).
Komatsu, I., Wang, J. H., Iwasaki, K., Shimizu, T. & Okano, T. The effect of tendon stem/progenitor cell (TSC) sheet on the early tendon healing in a rat Achilles tendon injury model. Acta Biomater. 42, 136–146 (2016).
Meloche, S. & Pouysségur, J. The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transition. Oncogene 26 (22), 3227–3239 (2007).
Shi, H. et al. Myoprotective effects of bFGF on skeletal muscle injury in pressure-related deep tissue injury in rats. Burns Trauma. 4, 26 (2016).
Poitras, T. & Zochodne, D. W. Unleashing intrinsic growth pathways in regenerating peripheral neurons. Int. J. Mol. Sci. 23(21), 13566 (2022).
Acknowledgements
We thank all authors who contributed valuable methods and data and made them public. The authors acknowledge the Laboratory Animal Centre of Ningbo University for providing a superior breeding environment and operation room for the animal experiments in this study.
Funding
This research was funded by the National Natural Science Foundation of China under Grant number 82372385; Major Science and Technology project for Health of Zhejiang Province under Grant number WKY-ZJ-2445; Science and Technology Innovation 2025 Major Project of Ningbo under Grant number 2022Z133; Ningbo Health Science and Technology project under Grant number 2023Y08; International Cooperation Project of Ningbo City under Grant number 2023H013; Zhejiang Province Medical and Health Science and Technology Programme under Grant number 2025 KY1373.
Author information
Authors and Affiliations
Contributions
Xufeng Mao: methodology, formal analysis, investigation, resources, writing—original draft preparation, writing—review and editing, visualization, supervision, project administrationYulong Wang: software, formal analysis, investigation, writing—original draft preparation, project administrationXiqian Zhang: resources, writing—original draft preparation, visualization, supervisionZheyu Yao: software, data curation, Zheyang Yuan: resources, formal analysisLiwei Yao: investigation, project administrationLeining Wang: visualization, supervisionHaijiao Mao*: conceptualization, data curation, funding acquisition, project administration.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
The collection of samples used in this study was approved by the Ethics Committee of the First Affiliated Hospital of Ningbo University (ethical approval number: KS202211017). The Animal Research Ethics Committee of Ningbo University in China gave its approval for the animal experiments (ethical approval number: 13605). All participants were provided with written informed consent at the time of enrollment.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Mao, X., Wang, Y., Zhang, X. et al. Decellularised amniotic membrane-TDSCs composite promotes Achilles tendon healing. Sci Rep 15, 15556 (2025). https://doi.org/10.1038/s41598-025-00596-0
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-00596-0
