
Abstract
Mechanical loading induces bone formation in young rodents, but mechanoresponsiveness is reduced with age. Glycolytic activity and mitochondrial dysfunction increase with age and may change bone mechanotransduction. To evaluate load-induced changes to bioenergetic activity in young and adult animals, we loaded the tibia of 10-wk and 26-wk female C57BL/6J mice and examined transcriptomic responses at the mid-diaphysis, and metaphyseal cortical shell and cancellous core. Across all biological processes, oxidative phosphorylation and mitochondrial pathways were most often enriched with loading and had contrasting enrichment in young and adult animals. Following loading, young animals had temporally-coordinated differential expression of mitochondrial-associated genes, with greatest expression at the mid-diaphysis. In adults, bioenergetic gene expression was lower compared to young animals. To assess individual contributions of glycolysis and pyruvate-mediated oxidative phosphorylation to load-induced bone formation in vivo, we inhibited each pathway therapeutically and loaded the tibia of young and adult female mice for 2 weeks. In both young and adult mice, loading increased cortical bone mass, but inhibition of oxidative phosphorylation reduced cortical area and moment of inertia in both loaded and control limbs. Conversely, load-induced improvements of adult cancellous bone depended on glycolysis. In summary, mechanical loading transcriptionally activated mitochondrial pathways in an age-specific manner and bioenergetic inhibition revealed unique metabolic programs for cortical and cancellous bone.
Similar content being viewed by others

Introduction
Osteoporotic fractures increase mortality1 and disproportionally affect women2,3. Despite the growing incidence of fractures globally2, anabolic therapies that increase bone mass remain limited4. The human skeleton is mechanosensitive; mechanical loading from exercise increases bone mass during growth but anabolic responses are reduced in aged bone5,6. In addition to eliciting changes in bone mass, loading increases transcriptomic activity in the growing skeleton as a function of mechanical strain7. Load-induced anabolic responses in murine bone are diminished in adulthood8 and continue to decline in old age9,10. Reduced mechanotransduction with aging may reflect age-related metabolic shifts in bone cells. In the skeleton, advancing age is correlated with changes to bioenergetic pathway activity at the protein- and tissue-level11. Metabolic shifts occur in whole bone with age; glycolysis increased and oxidative phosphorylation decreased in old mice compared with young animals12.
In vivo rodent loading models replicate the age-related changes in human bone mechanosensitivity8,13,14, thereby allowing for the discovery of biological pathways contributing to reduced mechanotransduction. To identify potential mechanisms causing reduced mechanotransduction, biological pathways responsible for tissue-level responses to mechanical loading can be assessed in an unbiased manner with gene expression. Load-induced transcriptional activation of bioenergetic pathways in whole bone samples differ between young and old animals9,15, highlighting potential age-related dysregulation in the cellular generation of ATP.
The metabolic profile of bone tissue reflects contributions of multiple cells, including osteoclasts, osteoblasts, and osteocytes. Both oxidative phosphorylation and glycolysis are used by mature osteoclasts16,17 and osteoblasts18,19, but little is known about the bioenergetic profile of osteocytes20,21. Stromal cells and osteoblasts collected from aged animals have increased mitochondrial dysfunction, a hallmark of aging, compared to young mice15. When mitochondrial dysfunction was increased in genetically manipulated mice, osteoblast number was reduced and osteoclast number was increased, resulting in reduced bone formation22.
Changes in bioenergetic activity can contribute to bone formation. The anabolic activity of parathyroid hormone may be partially attributed to the promotion of aerobic glycolysis in bone23,24, mirroring the effects of Wnt signaling on bioenergetic pathways25. Inhibiting glycolysis improves cortical bone architecture and increased bone strength in both young and old male mice26. Although glycolysis is only moderately activated by in vivo loading7, oxidative phosphorylation has been implicated in the load-induced transcriptional response of cortical bone7,9,27. Cortical and cancellous bone have unique transcriptomic responses to loading28, but little is known about their individual bioenergetic profiles. Therefore, we hypothesized that the load-induced transcriptional activation of bioenergetic pathways would vary with age and between cortical and cancellous bone. We further speculated that the load-induced bone formation in cortical and cancellous bone of young and adult animals would be uniquely affected by therapeutic inhibition of pyruvate-mediated oxidative phosphorylation or glycolysis.
Results
Load-induced transcriptional activation of oxidative phosphorylation differed with age and between cortical and cancellous bone
To identify novel pathways involved in the load-induced transcriptional response of bone, we performed an unbiased analysis using both previously published7 and unpublished gene expression data. We evaluated temporal differential gene expression and activated biological processes following in vivo tibial compression in 10-week-old (young) and 26-week-old (adult) female mice (Fig. 1A–C). The tibial mid-diaphysis, and metaphyseal cortical shell and cancellous core were analyzed separately. At each timepoint, young and adult mice had distinct skeletal transcriptomes (Fig. S1). Pathways enriched by tibial loading across multiple tissue segments and time points were considered highly activated (Figs. 1D, S2).

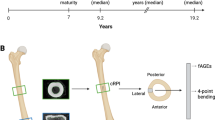
Mechanical loading transcriptionally activated oxidative phosphorylation in bone. (A) Schematic of mouse tibial loading configuration. (B) Cyclic loading waveform for loading at a 9 N magnitude and 4 Hz frequency. (C) Evaluation of load-induced transcriptional response: Short-term transcriptomic differences induced by loading were evaluated at 1-hour, 3-hours, or 24-hours after loading. Mice underwent daily loading for one week to examine later load-induced changes in gene expression. n = 4–6/group. Sequencing was validated at 1 week time point. n = 5/group (D) Oxidative phosphorylation (GO) was enriched at most time points in all tissue segments, and different with age. Glycolysis (GSEA Hallmark) was less often enriched but also different with age. Red = positively enriched with loading. Blue = negatively enriched with loading.
Among all pathways examined in our unbiased list of gene sets, metabolic pathways related to mitochondrial function and oxidative phosphorylation were most frequently enriched and had the greatest enrichment scores induced by loading7 (Figs. 1D, S2). Across all samples collected, enrichment of mitochondrial-associated pathways was greatest at the mid-diaphysis of young animals (Fig. S2). Oxidative phosphorylation was most often positively enriched, or upregulated, with loading at the mid-diaphysis in young animals (Fig. 1D). Following 1 wk of loading, oxidative phosphorylation enrichment at the mid-diaphysis was positive in young animals but negative in adults. Oxidative phosphorylation enrichment also was opposite between young and adult mice at 24 h post-loading in both the metaphyseal cortical shell and cancellous core. Glycolysis had limited enrichment across the samples, but enrichment was dissimilar between young and adult animals (Fig. 1D).
Differential expression of genes whose products are located within the mitochondria or involved in the electron transport chain was distinct between tissue segments and animal age
Evaluating specific genes involved in mitochondrial function and oxidative phosphorylation revealed further differences in gene expression between the tissue segments. Genes transcribing products that either localize to specific areas of the mitochondria or are associated with the electron transport chain (ETC), a sub-process of oxidative phosphorylation, were investigated. Load-induced differentially-expressed genes (DEG) were identified at all sub-mitochondrial compartments (intermembrane space, matrix, inner membrane, and outer membrane). The number of timepoints at which a given gene was differentially expressed was quantified to determine the frequency of that DEG; the most frequent DEG sub-localizing to the mitochondria were associated with either the mitochondrial matrix or mitochondrial inner membrane (MIM) (Fig. 2). The mid-diaphysis had most load-induced DEG related to the mitochondrial matrix and MIM whereas the metaphyseal cortical shell had the least, with an intermediate number expressed in cancellous bone.

Load-induced differential expression of genes encoding mitochondrial products was distinct between tissue segments and animal age. Most commonly differentially expressed genes whose products localize to sub-mitochondrial compartments were associated with mitochondrial matrix (Matrix) or mitochondrial inner membrane (M.I.M.), with fewer genes related to mitochondrial membrane (M.) and mitochondrial outer membrane (M.O.M.). Red = upregulated with loading. Blue = downregulated with loading. White = not significantly differentially expressed. * represents fold changes outside of the range displayed.
Load-induced differential expression of genes whose products localize to the mitochondria or are related to the ETC also was different between young and adult mice. At the mid-diaphysis, all MIM-associated DEG downregulated at 1 h in adults were also downregulated at 3 h in young animals (Fig. 2). In cortical segments, the proportion of load-induced DEG related to the MIM, normalized to the gene set, shifted with age; at the mid-diaphysis, the most MIM-associated DEG were present in young mice, whereas at the metaphyseal cortical shell the most MIM-associated DEG were present in adults (Table 1). At the mid-diaphysis, DEG related to the ETC were consistently downregulated at 3-h post-loading followed by upregulation after 1 wk of loading in young animals (Fig. S3). Adults lacked the coordinated temporal response of DEG associated with the ETC observed in young animals (Fig. S3).
Mitochondrial gene expression was spatially altered by loading
To validate the sequencing data, we examined in situ hybridization of genes related to mitophagy and osteoclast activity, Pink1 and Acp5 respectively. Pink1 encodes for a mitochondrial serine/threonine-protein kinase and Acp5 encodes for tartrate-resistant acid phosphatase. In the RNA sequencing data set, Pink1 was downregulated in the adult metaphyseal cortical shell at 1-wk post-loading (Fold change (FC) = -2.30). A second cohort of young and adult female mice underwent tibial loading for 1 week. Across all tissue locations, Acp5 was primarily expressed by bone-lining cells (Figs. 3A,B, S4A). At both the metaphyseal cortical shell and mid-diaphysis, loading decreased Pink1 expression in the posterior region (Fig. 3C,D). At the metaphysis, loading increased Pink1 expression in the anterior region. At the mid-diaphysis, adults had greater Pink1 and Acp5 expression compared with young animals. Loading increased Acp5 expression in the anterior region of the metaphyseal cortical shell, but decreased Acp5 expression in the posterior region of the mid-diaphysis. In cancellous bone, loading increased the percent of area stained for Pink1 (Fig. S4). The nature of these changes corresponded to the RNAseq-based changes measured.

In situ hybridization validated differences in mitophagy and osteoclast gene expression between loaded and control limbs in young and adult cortical bone. (A, B) Visualization of Pink1 (green) and Acp5 (yellow) in metaphyseal cortical shell and mid-diaphysis. Bone surface distinguished by white dotted line. (C, D) Quantification of percent area moderately stained for each probe at metaphyseal cortical shell and mid-diaphysis.
At all tissue segments, Pink1 expression most often co-localized with Acp5 expression, suggesting these transcripts were encoded by the same cell type, presumably osteoclasts (Fig. S5); over 70% of Pink1 expression was co-localized to Acp5. The co-localization of Acp5 and Pink1 expression differed with age, load, and region in cortical bone. At the metaphyseal cortical shell, loading increased co-localized expression of Acp5 and Pink1 in the anterior region of young animals (load*age*region: p = 0.032). At the mid-diaphysis, the percent area containing both Acp5 and Pink1 was greater in adults compared to young animals (age: p = 0.011). The co-localization of Pink1 and Acp5 in the posterior mid-diaphysis was decreased 3% with loading (load*region: p = 0.001), from 99 to 96%.
Compared to inhibition of glycolysis, inhibiting oxidative phosphorylation systemically increased bone resorption and altered whole bone bending properties
To evaluate the contributions of bioenergetic pathways on the response of the skeleton to loading, oxidative phosphorylation and glycolysis were individually inhibited with MSDC-016029 (MSDC) and sodium oxamate26 (SO), respectively, in young (10 wk) and adult (26 wk) female mice (Fig. 4A). Bodyweight did not change with bioenergetic inhibition (Fig. S6). Inhibiting oxidative phosphorylation increased systemic bone resorption. TRAcP 5b increased in the serum of MSDC-treated animals relative to SO and control groups (Fig. S7). Overall, bone turnover decreased with age (Fig. S7). Systemic markers of both bone resorption (TracP 5b) and formation (P1NP) were decreased in adults compared to young animals. Bioenergetic inhibition altered whole bone bending strength of the humerus but did not affect compressive behavior of the vertebra. Humerus whole bone bending stiffness increased with MSDC treatment relative to SO, but neither therapeutic inhibition differed from the control group (Fig. S8A). Therapeutic inhibition did not alter humeral ultimate moment (Fig. S8B). Neither age nor bioenergetic inhibition altered vertebral compressive stiffness and maximum load. Treatment did not alter humerus or vertebral morphology. Humeral length and vertebral height were greater in adults compared to young animals (p < 0.001 and p < 0.001, respectively), but humeral diameter and vertebral cross-sectional area did not differ with age.

Oxidative phosphorylation inhibition reduced cortical area and moment of inertia. (A) Evaluation of bioenergetic inhibition on load-induced bone formation: Four days prior to the initiation of tibial loading, glycolysis and oxidative phosphorylation were inhibited via sodium oxamate and MSDC-containing chow, respectively. Sodium oxamate and MSDC administration continued throughout loading. n = 7–10/group (B, C) Cortical area was increased with loading and decreased with oxidative phosphorylation inhibition. (D, E) Cortical maximum moment of inertia was increased with loading and reduced with oxidative phosphorylation inhibition. Letters indicate loading or loading*age interaction effects. # indicates treatment effects. * indicates age effects.
Compared to glycolysis inhibition, inhibiting oxidative phosphorylation reduced cortical area and moment of inertia of the tibia
To evaluate morphological differences in the tibia following bioenergetic inhibition during loading, we performed microcomputed tomography at the metaphysis and mid-diaphysis. Bioenergetic inhibition, loading, and age had similar effects on cortical bone at the metaphysis and diaphysis. Inhibiting oxidative phosphorylation reduced cortical area (Ct.Ar) at both sites relative to the control group (Fig. 4B,C). Metaphyseal cortical thickness (Ct.Th) was reduced in MSDC-treated animals compared to controls. Inhibition of oxidative phosphorylation reduced maximum moment of inertia (IMAX) at the metaphyseal cortical shell compared to inhibition of glycolysis, but neither treatment group differed from the control group (Fig. 4D). MSDC treatment reduced mid-diaphyseal IMAX compared to the control group (Fig. 4E). Relative to the control group, oxidative phosphorylation inhibition reduced minimum moment of inertia (IMIN) at both sites (Fig. S9). Neither treatment altered mid-diaphyseal Ct.Th. At both sites, treatment did not change cortical tissue mineral density (Ct.TMD).
Cortical bone mass was increased with loading and was greater in adult animals. Loading increased metaphyseal and mid-diaphyseal Ct.Ar, Ct.Th, IMAX, and IMIN in both young and adult animals (Fig. 4B–E). Ct.TMD at the metaphyseal cortical shell was reduced with loading in adult animals. Compared with young animals, adults had greater Ct.Th. and Ct.TMD at both the metaphysis and mid-diaphysis. At the mid-diaphysis, Ct.Ar, IMAX, and IMIN also were greater in adults compared with young animals (Fig. 4B–E). Comparing control limbs, adults had greater metaphyseal Ct.Ar, IMAX, and IMIN compared to young animals, indicating age-related differences at the metaphysis. Mid-diaphyseal marrow area (Ma.Ar) was not different with loading, treatment, or age. Independent of bioenergetic inhibition, loading and age both increased cortical bone mass and moment of inertia.
Load-induced bone formation in adult cancellous bone depended on glycolysis
The response of cancellous bone to loading varied with age and bioenergetic inhibition. Bone volume fraction (BV/TV) increased with loading in young animals, but not adults (Fig. 5A). Adult BV/TV was significantly lower compared to young animals. Trabecular number (Tb.N) and cancellous tissue mineral density (Tb.TMD) decreased with loading, whereas trabecular separation (Tb.Sp) increased (Fig. 5B,C). Adults had greater Tb.Sp and Tb.TMD, but lower Tb.N, compared to young animals. In general, loading increased trabecular thickness (Tb.Th) at both ages; however, inhibition of glycolysis via SO prevented load-induced increases in Tb.Th in adults (Fig. 5D).

Glycolysis inhibition altered metaphyseal cancellous response to loading. (A) Loading increased BV/TV in young animals. (B) Cancellous tissue mineral density increased with loading and was greater in adults compared to young animals. (C) Trabecular separation was increased with loading and higher in adults compared to young animals. (D) Trabecular thickness was increased with loading and reduced with oxidative phosphorylation inhibition.
Bioenergetic inhibition did not alter active osteoblast and osteoclast number
Bone cell activity was assessed with anti-pro-collagen immunohistochemistry (active osteoblasts) and TRAP staining (osteoclasts). Active osteoblast number per bone surface at the metaphyseal cortical shell and cancellous core were altered by loading and age, but not affected by treatment. Loading decreased active osteoblast number per bone surface in young animals (Fig. S10A,B). In the posterior region of the metaphyseal cortical shell, active osteoblast number per bone surface also decreased with loading. Within the posterior regions of the metaphyseal cortical shell, young animals had greater active osteoblast number per bone surface compared to adults. At the mid-diaphysis, active osteoblast number per bone surface did not differ with age, treatment, loading, or between anterior and posterior regions (Fig. S10C).
Osteoclast number per bone surface also was altered by loading and age, but not different between treatment groups. At the metaphyseal cortical shell and mid-diaphysis, osteoclast number per bone surface was greater in adults compared with young animals (p = 0.029 and p = 0.016, respectively). The anterior surface of cortical bone had greater osteoclast numbers per bone surface compared to the posterior surface (p < 0.001 at the metaphyseal cortical shell, p = 0.033 at the mid-diaphysis). Neither load nor treatment affected osteoclast number in cortical bone. Osteoclast number per bone surface did not change in cancellous bone with load, age, or treatment.
Bioenergetic inhibition affected lactate dehydrogenase expression at the adult mid-diaphysis
Sodium oxamate outcompetes pyruvate, its structural analog, to bind to lactate dehydrogenase and inhibit glycolysis; the bound SO-LDH complex is maintained within cells26,30,31. To assess glycolytic activity within the bone tissue, sections were stained for lactate dehydrogenase and the percent of positively-stained cells quantified (Fig. S10D–F). At the mid-diaphysis, SO increased the percent of LDH-positive cells in the control limbs of adults. At both the metaphyseal cortical shell and mid-diaphysis, the percent of LDH-positive cells was greater in the posterior compared with the anterior region. Load, treatment, or age did not affect the percent of LDH-positive cells at the metaphyseal cortical shell. The percent of LDH-positive cells did not differ with age, treatment, or loading in cancellous bone.
Bioenergetic inhibition affected bone formation indices in adult but not young animals
We used dynamic histomorphometry to assess bone formation indices in the metaphysis. Due to a lack of detectable double labels on the cortical surface in several adult mice, animals were analyzed separately by age. Bioenergetic inhibition affected bone formation indices in adults, but not young animals. MSDC treatment reduced endosteal mineralizing surface in adults, but not young animals (Fig. 6A, Table S1). Bone formation indices at the periosteal surface were not affected by bioenergetic inhibition (Table S1, 2). In adults, SO treatment reduced cancellous mineral apposition rate compared with control mice (Fig. 6B).

Bioenergetic inhibition reduced endosteal mineralizing surface and cancellous mineral apposition rate. (A) Oxidative phosphorylation inhibition reduced endosteal mineralizing surface. (B) Glycolysis inhibition reduced mineral apposition rate in cancellous bone. Loading increased cancellous mineral apposition rate.
Loading affected bone formation indices in both young and adult mice (Tables S1, S2). At the endosteal surface, loading reduced mineralizing surface at both ages. In young animals, loading reduced endosteal mineral apposition rate and bone formation rate. At both ages, loading increased periosteal mineralizing surface. In young animals, loading also increased periosteal mineral apposition rate, and bone formation rate. In both young and adult mice, loading increased cancellous mineral apposition rate and bone formation rate in adult mice.
Discussion
Development of improved treatments for osteoporosis requires better understanding of the anabolic mechanisms stimulating bone formation. In the present study, we have translated our transcriptional findings associated with load-induced bioenergetic pathway activation to demonstrate that bone morphology is governed by different bioenergetic requirements in cortical and cancellous bone. Based on a detailed analysis of differential gene expression related to mitochondrial function, highly expressed genes most often encoded proteins located in the mitochondrial matrix and MIM. Inhibition of oxidative phosphorylation via MSDC decreased cortical bone mass and moments of inertia compared to the control group. Conversely, cancellous bone microarchitecture only was affected by glycolysis inhibition, which prevented load-induced improvements in adults. The differences we measured at the transcriptional and tissue levels demonstrated that bioenergetic pathways play unique roles in the maintenance and anabolic response of both cortical and cancellous bone and may contribute to age-related changes in bone formation.
The two cortical bone sites examined had distinct load-induced transcriptional responses associated with bioenergetic pathways that may be associated with mechanical strain induced by tibial loading. A greater fraction of genes encoding mitochondrial products were differentially expressed with loading at the mid-diaphysis compared to the metaphyseal shell. The transcriptional response associated with mitochondrial function was greatest at the mid-diaphysis and lowest at the metaphyseal cortical shell in young animals, but not adults. These changes reflect the gradient of strain within the cortex32, suggesting that mechanical strain may govern mitochondrial-related transcriptional activity7. In addition to opposite trends in enrichment of bioenergetic processes between young and adult mice, enrichment scores increased at the metaphyseal cortical shell and decreased at the mid-diaphysis with age. With age, metaphyseal cancellous bone volume was reduced8, and load-sharing with the surrounding cortical bone increased, likely increasing cortical strain, which may account for increased mitochondrial transcriptional activity at the metaphyseal cortical shell in adult mice. Additionally, tibial bone material properties were different in adults compared to young animals8. Cortical bone strength has been correlated with differences in metabolite abundances33, directly linking the bioenergetic activity of bone cells to whole bone structural properties. Here, we demonstrated that inhibiting bioenergetic activity altered bone strength.
Bioenergetic demands change with age, and a better understanding of bioenergetic differences between young, growing animals and mature adults is required. With age, mitochondria produce reactive oxygen species that increase apoptosis in bone cells11. 13-month-old animals have increased glycolytic intermediates and decreased cell energy demand compared to 3-month-old animals12. Young animals (2.5 mo) form bone in response to mechanical load whereas adults (6 mo) have diminished tissue-level responses8,13,14. In our study, young (2.5 mo) and adult (6 mo) animals had opposite trends in load-induced transcriptional bioenergetic enrichment, suggesting that the onset of age-related differences in oxidative phosphorylation occurs by adulthood, before animals reach advanced age9,15. Bioenergetic demands likely change throughout the lifespan. Clinically, glucose uptake in bone varies throughout adulthood34, mirroring the age-related declines in skeletal glucose uptake reported in mice35.
Little is known about the bioenergetic state of osteocytes. Osteocytes located in high strain regions of cortical bone have more mitochondria than low strain regions36,37, implying increased oxidative phosphorylation activation with increasing tissue deformation. As the putative mechano-sensing cell of bone, osteocytes initiate essential cascades that lead to load-induced bone formation. Load-induced bone formation was altered by bioenergetic inhibition but was present in both treatment groups, suggesting that osteocyte signaling can persist when oxidative phosphorylation or glycolysis are individually inhibited36,38,39. The load-induced increases in bone mass observed across groups likely coincide with increased osteocyte number; mineralizing osteoblasts differentiate into osteocytes, thus potentially lowering the number of active osteoblasts recorded here following loading. In this study, the percentage of embedded bone cells that were LDH-positive was greater in the compressive region of cortical bone (posterior) compared to the tensile region (anterior). Loading modality may influence bioenergetic pathway activity in osteocytes, with greater glycolytic activity in regions of compression compared to tension. Increased glycolytic activity may result in faster ATP generation21, which may enable the enhanced tissue-level anabolic response to loading recorded in compressive compared with tensile regions40.
Clinically, bone formation differs in regions subject to tension or compression41,42, and may be driven by differences in bioenergetic gene expression or cell populations in these regions. In our study, expression of mitophagy gene Pink1 decreased with loading in the compressive region (posterior) but increased in the anterior region. Similarly, expression of osteoclast gene Acp5 was upregulated with loading within the tensile region of the metaphysis and downregulated within the compressive region of the mid-diaphysis. We recorded greater osteoclast numbers per bone surface in the anterior cortical regions (tensile) compared with posterior. The differences in cell populations in tensile and compressive regions may contribute to differences in regional bioenergetic profiles16,18,20,22,43.
Oxidative phosphorylation and glycolysis are essential processes for mature osteoblast18,20,43 and osteoclast16,22 function. Bioenergetic inhibition during this experiment did not affect active osteoblast or osteoclast number in our samples, as measured by immunohistochemistry and histology. However, the cell numbers do not reflect all activity or efficacy that is altered by bioenergetic activity. In some cell types, MSDC treatment enhances mitochondrial fitness44, which could alter cellular activity. Bone resorption marker TRAcP 5b increased in the serum of MSDC-treated animals, suggesting increased osteoclast activity. Oxidative phosphorylation is required for osteoclast differentiation but resorption depends on glycolysis16,45. Glycolysis is faster than oxidative phosphorylation and not limited to specific organelles and locations, enabling easier generation of ATP at the sites of resorption46,47. Inhibition of pyruvate-mediated oxidative phosphorylation may increase the glycolytic activity within cells, thereby improving the efficiency of osteoclast-mediated bone resorption. This increased efficiency may explain the decreased cortical bone area and moment of inertia when oxidative phosphorylation was inhibited.
Inhibition of glycolysis may affect the function of osteoblasts more than osteoclasts. Although both oxidative phosphorylation and glycolysis are active in osteoblasts, collagen synthesis relies more heavily on glycolysis18,19,20,43. When oxidative phosphorylation is inhibited in osteoblasts, glycolytic activities can increase to maintain ATP production43; however, when glycolysis is inhibited in osteoblasts, oxidative phosphorylation activity does not change and overall ATP levels decline43. Therefore, inhibiting glycolysis via SO likely affected the osteoblast’s ability to respond to mechanical loading in adult cancellous bone. In whole bone samples, older mice had more glycolytic intermediates compared to young animals, suggesting elevated glycolysis activity with age12. In our study, 2.5 weeks of bioenergetic inhibition did not change osteoblast numbers, but longer-term treatment may have negatively affected osteoblast differentiation18,38,43,48.
Several limitations affect the interpretation of our findings. Our detailed analysis transcriptional activity relating to mitochondrial function offered insights into the connection between bioenergetics and mechanical loading, but future advances in spatial transcriptomics and metabolomics will enable investigations of differences in regions experiencing tensile and compressive strains. Pink1 expression differed in regions exposed to tensile and compressive strains, suggesting that expression of genes related to mitochondrial function may vary circumferentially within cortical bone. Metabolic pathways active in human bone vary spatially49, and further examination of load-induced changes to spatial metabolomics is warranted50,51. In addition to limitations surrounding spatial analyses, our methods of bioenergetic inhibition limit the findings of our study. To investigate the load-induced transcriptional changes in mitochondrial function at the tissue-level, we examined contributions of major bioenergetic pathways by targeting pyruvate-mediated oxidative phosphorylation and glycolysis; amino acids and fatty acids also are known contributors to oxidative phosphorylation in mature osteoblasts20. Our therapeutics were systemically administered; targeting specific bone cell types would elucidate the mechanisms driving the morphology changes recorded here. Further, the doses for bioenergetic inhibition used in this study were selected based on previously published work26,29. Our tissue-level analyses of bioenergetic inhibition on bone formation were limited to two weeks of loading. Although our pretreatment period ensured inhibition upon the start of loading, extended treatment would account for any short-term compensation43 via increased activity of alternate bioenergetic pathways in bone cells. Longer term treatment with bioenergetic inhibitors may also yield more pronounced changes to bone morphology. Although cortical bone did not respond to 2.5 weeks of glycolysis inhibition, previously 8 weeks of SO treatment improved cortical bone architecture in young male mice26. Future examinations of metabolite abundances within bone tissue following inhibition of glycolysis or pyruvate-mediated oxidative phosphorylation could yield insight into potential compensation by other bioenergetic pathways33,52. At the mid-diaphysis, inhibition of glycolysis increased LDH-positive cell counts in control limbs of adult mice. Accumulation of pyruvate in muscle has been associated with increased LDH production53; similarly, SO-treated animals may have had elevated pyruvate due to the slow execution of oxidative phosphorylation, resulting in a compensatory increase in LDH as an attempt to re-establish glycolytic activities. In bone, glycolytic intermediates decrease with age12, suggesting that compared to older animals, young mice are less dependent on skeletal glycolysis. In our study, inhibiting glycolysis with sodium oxamate did not affect bone morphology or the percent of LDH positive cells in young mice. Finally, our small sample sizes for some outcome measures, such as histology quantification, may have precluded us from detecting significance or reliably identifying statistical outliers within our data.
In conclusion, we found that oxidative phosphorylation and mitochondrial genes were highly enriched with loading across cortical and cancellous bone of the mouse tibia in young and adult animals. Preservation of cortical bone mass relied on oxidative phosphorylation, with no effects on cancellous bone. Conversely, inhibiting glycolysis prevented load-induced improvements in adult trabecular thickness, indicating that glycolysis is necessary for cancellous bone formation in adults. Bioenergetic inhibition did not affect young cancellous bone, suggesting decreases in glycolysis may be compensated for by increasing oxidative phosphorylation and vice versa. The treatment differences were not explained by changes in osteoblast or osteoclast number, suggesting modifications in cellular activity or efficacy. This work demonstrated that mechanical loading transcriptionally altered bone mitochondrial function in an age-specific manner and established key differences in the bioenergetic programs of cortical and cancellous bone.
Materials and methods
Evaluation of load-induced transcriptional response: experimental design
To understand the load-induced transcriptional response of bone, 10-week-old (young) and 26-week-old (adult) C57Bl/6J female mice (Jackson Laboratory, Bar Harbor, ME, USA) were subjected to at least one bout of in vivo cyclic compression of the left tibia under anesthesia (2% isoflurane, 1.0 L/min, Webster). All experiments were performed under IACUC approval, in accordance with relevant guidelines, regulations, and ARRIVE guidelines. Right limbs served as contralateral control limbs. Compressive load was applied to the mouse tibia (triangular waveform, 9 N peak load, 4 Hz, 1200 cycles per bout) using a custom feedback controlled device (Fig. 1A,B)8,13,54. Normal cage activity was resumed following loading. Animals were housed 3–6 per cage with free access to water and chow (Envigo Teklad LM-485 7912, Indianapolis, IN, USA, unless diet is stated otherwise). Mice were not fasted prior to euthanasia.
Multiple time points following loading were examined. Early transcriptional responses following one bout of loading were assessed at 1-hour (n = 5 young animals, n = 4 adults), 3-hours (n = 4 young animals, n = 6 adults), or 24-hours (n = 4 young animals, n = 5 adults) (Fig. 1C). To evaluate later transcriptional changes, mice underwent one week of tibial loading, consistent with previous studies8,13,14,54 (n = 5 young animals, n = 4 adults). Following euthanasia with cervical dislocation, both tibiae were dissected, and soft tissue removed. Bone marrow was removed through centrifugation, and the metaphysis was separated into cortical and cancellous bone using a 1 mm biopsy punch (Miltex, Integra LifeSciences Corp, Plainsboro, NJ, USA)55. The cortical mid-diaphysis also was segmented and processed. Both loaded and contralateral limbs from each animal were included for analysis, resulting in six samples per mouse. Analysis of genes and pathways related to the skeleton, but not mitochondria, were previously published using the young cortical bone segments7. All other transcriptomic analyses presented here are original; the compartment-specific differences at the metaphysis and the adult data have not previously been reported. Animal work for both published7 and newly reported transcriptomic studies was performed during the same time period; mice were consistently euthanized in the morning to reduce circadian effects in the data set.
RNA sequencing
Bone tissue was collected and processed according to previously established methods7,28,55. Animals were included for analysis if all 6 tissue samples collected were high quality (RQN > 6.0). If a single sample did not pass quality checks, the mouse was excluded. Samples from 37 out of 66 mice were sequenced (3’ RNA sequencing, NextSeq500, Illumina, San Diego, CA, USA56, resulting in an average of 4.6 million reads per sample. 62% of reads were uniquely mapped to the mm10 genome using STAR alignment57. A paired design (edgeR58 determined differential expression between loaded and control limbs (|Fold-change| > 2, false discovery rate < 0.05). At each timepoint, variance was measured with principal component analysis (Fig. S1). Genes whose products localize to specific mitochondrial compartments were identified (MitoCarta3.059). Enriched biological processes were determined with Gene Set Enrichment Pre-Ranked Analysis60 (false discovery rate < 0.25) using previously established methods7. Gene sets examined included GSEA Hallmark gene sets and Ontology gene sets.
mRNA in situ hybridization
Using fluorescent in situ hybridization, mRNA expression of Pink1 and Acp5 was validated in young (n = 5) and adult (n = 5) C57B6J females following 1 week of loading (Jackson Laboratory, Bar Harbor, ME, USA). Following euthanasia, tibiae were fixed in neutral buffered formalin, decalcified in 10% EDTA and embedded in paraffin. Sagittal sections of 6-µm thickness were obtained. RNAscope© Multiplex Fluorescent Reagent Kit V2 (ACD, Newark, CA, USA) with custom bone reagent was applied with Pink1 and Acp5 probes. Probes were visualized with Opal dyes (Akoya Biosciences, Menlo Park, CA, USA) at FITC and TRITC spectra. DAPI stained cell nuclei. Images were obtained with a fluorescent slide scanner (Aperio Scanscope, Leica Biosystems, Danvers, MA, USA). The cancellous bone, metaphyseal cortical shell, and mid-diaphysis were individually analyzed, with anterior and posterior regions of the cortex evaluated separately. Stained area was quantified (HALO® Image Analysis Platform, Indica Labs, Albuquerque, NM, USA) and normalized by total bone area. Appropriate RNA quality and optimal permeabilization were validated using positive control probes recommended for RNAscope©. Polyubiquitin-C (Ppib), Ubiquitin C (Ubc), and Gapdh, a commonly-used control gene, were selected for their prevalent expression across cell types (Fig. S11).
Evaluation of bioenergetic inhibition on load-induced bone formation: experimental design
To evaluate the contributions of glycolysis and oxidative phosphorylation on the response of bone to loading, each individual pathway was therapeutically inhibited (Fig. 4A). Twice weekly intraperitoneal (IP) injections of sodium oxamate (SO) at a previously established dose of 100 mg/kg26 (Santa Cruz Biotechnology, Dallas, TX, USA) inhibited glycolysis by preventing the lactate dehydrogenase conversion of pyruvate to lactate, which forced pyruvate to enter the TCA cycle and participate in oxidative phosphorylation. Three SO-treated adults were excluded from the study due to excessive weight loss, resulting in n = 10 young animals and n = 7 adults. In a second treatment group, inhibition of the mitochondrial pyruvate carrier was achieved with MSDC-0160 (MSDC), preventing pyruvate contributions to oxidative phosphorylation and forcing increased glycolysis-mediated pyruvate conversions61. MSDC was delivered orally via chow (Research Diets, Inc., New Brunswick, NJ, USA) at a previously established dose to achieve full pharmacology and inhibition of the mitochondrial pyruvate career29 (30 mg/kg/day) (MedChem Express Monmouth Junction, NJ, USA). MSDC-treated animals also received IP injections of sterile PBS twice weekly (Alfa Aesar, Tewksbury, MA, USA) (n = 10 young animals, n = 10 adults). The final group served as controls and received twice weekly IP PBS injections and control chow (n = 10 young animals, n = 10 adults). Following a 4-day pretreatment period, animals underwent tibial loading for two weeks, consisting of 5 days of loading per week8,13,14,54. Mice were not fasted prior to euthanasia.
Serum measurements
Animals were euthanized by cardiac puncture. Blood was collected and stored at 4 ºC overnight. Serum was separated from the blood by centrifugation at 4000 rpm for 20 min. Systemic bone resorption and formation parameters were assessed via TRAcP 5b (IDS, Gaithersburg, MD, USA) and P1NP (IDS, Gaithersburg, MD, USA), respectively (n = 7–10/group).
Evaluation of whole bone structural properties
To evaluate systemic effects of bioenergetic inhibition on the whole bone structural behavior of sites with predominantly cortical and cancellous bone, we performed whole bone testing of the humerus and lumbar vertebra. Following euthanasia, samples were dissected from surrounding soft tissue, wrapped in PBS-soaked gauze, and stored at -20 ºC prior to testing. Morphology was measured from x-ray images of the bones (ImageJ 1.5162). To assess whole bone strength, one humerus per mouse was loaded to failure in three-point bending in the anterior-posterior direction at a rate of 0.1 mm/s using a span length (w) of 5.7 mm between the outer supports (MiniBionix 858, MTS Systems Corporation, Eden Prairie, MN, USA and load cell SSM-100, Transducer Techniques, Temecula, CA, USA). Force and displacement were sampled at 100 Hz. Humeral bending stiffness was calculated as kw3/48, for k = slope of the linear portion of the force-displacement curve63,64. Ultimate moment (Mmax) was calculated as Pmaxw/4, for Pmax = maximum load. To examine a predominantly cancellous structure, caudal surfaces of lumbar vertebra L4 were fixed with cyanoacrylate adhesive, and a compressive force was applied to the cranial surface at a rate of 0.05 mm/s until failure (MiniBionix 858, MTS Systems Corporation, Eden Prairie, MN, USA, and load cell MLP-25, Transducer Techniques, Temecula, CA, USA). Vertebral compressive stiffness and strength were determined from the force displacement data.
Microcomputed tomography
Following bioenergetic inhibition during loading, morphological differences in the tibia were evaluated with microcomputed tomography (µCT). Tibiae were dissected and immediately fixed in 4% paraformaldehyde. Following overnight fixation, tissues were washed and placed in 70% ethanol. Tibiae were scanned at the metaphysis and mid-diaphysis with isotropic voxel resolutions of 10 μm and 15 μm, respectively (µCT35, Scanco Medical AG, Bruttisellen, Switzerland); 55 kVp, 145 µA, 600 ms integration time). A calibration phantom was used. Thresholds for the mid-diaphysis, metaphyseal shell, and cancellous bone were 2385 HU, 2557 HU, and 1590 HU, respectively. Volumes of interest were defined as 10% of total tibial length beginning 50 μm distal to the growth plate for the metaphysis and 2.5% of total tibial length centered at the midshaft for the mid-diaphysis65. At the metaphysis cancellous bone was analyzed separately from the cortical shell. Cancellous bone outcome measures included bone volume fraction (BV/TV), trabecular thickness (Tb.Th), separation (Tb.Sp) and number (Tb.N), and cancellous tissue mineral density (Tb.TMD). At both the metaphysis and mid-diaphysis cortical outcome measures included cortical area (Ct.Ar), cortical thickness (Ct.Th), maximum and minimum moment of inertia (IMAX and IMIN), cortical tissue mineral density (Ct.TMD), and marrow area (Ma.Ar, diaphysis only).
TRAP staining and anti-pro-collagen immunohistochemistry
Bone remodeling was assessed at the metaphysis and cortical mid-diaphysis. Following µCT scanning, a subset of tibiae was decalcified in 10% EDTA for two weeks (n = 4–5/group) and sectioned using previously established methods65. Sections were stained with anti pro-collagen I specific antibodies or for tartrate resistant acid phosphatase (TRAP) using previously established methods65. The number of active osteoblasts (pro-collagen I) and positively-stained osteoclasts (TRAP) were quantified65 (ImageJ 1.5162). All cancellous surfaces were quantified together, and the anterior and posterior regions of endosteal cortical bone were evaluated separately. Due to complications during sectioning, the periosteal surface was not quantified.
Lactate dehydrogenase immunohistochemistry
Expression of lactate dehydrogenase (LDH) was evaluated by immunohistochemistry. Paraffin-embedded sections were stained for lactate dehydrogenase with recombinant anti-lactate dehydrogenase antibody (Abcam, #ab52488, Cambridge, United Kingdom). The number of LDH-stained cells within the bone matrix was normalized to total cell number (QuPath 0.3.266) (n = 4–5/group). Anterior and posterior regions of cortical bone were evaluated separately. All trabeculae present in cancellous bone were quantified together.
Dynamic histomorphometry
Dynamic bone formation was assessed during bioenergetic inhibition. All animals received calcein injections 12 and 3 days prior to euthanasia (20 mg/kg IP). After µCT scanning, a subset of tibiae (n = 3–5/group) were embedded in acrylosin and sectioned sagittally by the Bone Histology/Histomorphometry Laboratory (Yale University Department of Orthopaedics and rehabilitation, New Haven, CT, USA). An area of interest, consisting of a 2000 μm x 2000 μm square centered 500 μm below the epiphysis, was identified. The cancellous bone was analyzed, and the endosteal and periosteal surfaces of the metaphyseal cortical shell were evaluated separately (2 slides per animal, OsteomeasureXP, v3.3.0.2, Osteometrics, Decatur, GA, USA). Mineral apposition rate (MAR) was determined. The mineralizing surface normalized to the bone surface (MS/BS) and bone formation rate (BFR/BS) were calculated. One sample had no detectable single labeled surface in the cancellous bone, resulting in BFR/BS measurement of 0 μm/year and was excluded from cancellous MAR analysis (ND). A total of 13 samples had no detectable double-labeled surfaces at either the periosteal or endosteal surfaces and were excluded from MAR and BFR/BS analysis at the site lacking double labels (ND)67.
Statistical analysis
For in situ hybridization, we evaluated stained probe area in cancellous bone using a linear mixed-effects model with fixed effects of age (young or adult), loading (contralateral control or loaded), and their interaction term and a random mouse effect. In cortical bone, stained probe area was assessed using a linear mixed-effects model with fixed effects of age (young or adult), loading (contralateral control or loaded), cortical region (anterior or posterior), and their interaction term and a random mouse effect.
Serum markers of bone turnover and the mechanical properties of humeri and vertebrae were analyzed using a two-way ANOVA to assess the effects of age (young or adult), treatment (SO, MSDC, control) and their interaction terms. We analyzed µCT, dynamic histomorphometry, cancellous bone histology, and cancellous bone IHC data using a linear mixed-effects model with fixed effects of age (young or adult), treatment (SO, MSDC, control), loading (contralateral control or loaded), and their interaction terms and a random mouse effect. Histology and IHC at the endosteal surface of cortical bone were evaluated separately for the metaphyseal cortical shell and mid-diaphysis using a linear mixed-effects model with fixed effects of age (young or adult), treatment (SO, MSDC, control), loading (contralateral control or loaded), cortical region (anterior or posterior), and their interaction terms and a random mouse effect.
In assessing bone formation indices calculated by dynamic histomorphometry, several adult samples did not have double labeled surfaces and were classified as ND. Samples classified as ND were not included for cortical analysis of MAR and BFR/BS, resulting in n < 3 in control limbs. Therefore, young and adult mice were analyzed separately using linear mixed-effects models with fixed effects of treatment (SO, MSDC, control), loading (contralateral control or loaded), and a random mouse effect. Periosteal and endosteal MAR and BFR/BS were not evaluated in adult mice.
Significant interaction terms were investigated with post-hoc analysis. For factors that were not included in significant interactions, a single factor evaluation determined significance68. For all statistical tests, post-hoc analysis was performed using Tukey’s test for significant effects to assess differences across more than two groups, and a Student’s t-test to assess differences between two groups. Normality was confirmed visually for all data using histograms and QQ plots of the residuals. Significance was set at p < 0.05.
Data availability
Original RNA sequencing data created for this study is available in Gene Expression Omnibus, GSE210827 and GSE281942.
References
Kanis, J. A., Oden, A., Johnell, O., De Laet, C. & Jonsson, B. Excess mortality after hospitalisation for vertebral fracture. Osteoporos. Int. 15, 108–112 (2004).
Amin, S., Achenbach, S. J., Atkinson, E. J., Khosla, S. & Melton, L. J. Trends in fracture incidence: a population-based study over 20 years. J. Bone Min. Res. 29, 581–589 (2014).
Melton, L. J., Chrischilles, E. A., Cooper, C., Lane, A. W. & Riggs, B. L. Perspective how many women have osteoporosis? J. Bone Min. Res. 7, 1005–1010 (2009).
Foessl, I., Dimai, H. P. & Obermayer-Pietsch, B. Long-term and sequential treatment for osteoporosis. Nat. Rev. Endocrinol. 19, 520–533 (2023).
Jones, H., Priest, J., Hayes, W., Tichenor, C. & Nagel, D. Humeral hypertrophy in response to exercise. J. Bone Joint Surg. Am. Volume. 59, 204–208 (1977).
Warden, S. J. et al. Physical activity when young provides lifelong benefits to cortical bone size and strength in men. Proc. Natl. Acad. Sci. U.S.A. 111, 5337–5342 (2014).
Chlebek, C., Moore, J. A., Ross, F. P. & van der Meulen, M. C. H. Molecular identification of spatially distinct anabolic responses to mechanical loading in murine cortical bone. J. Bone Mineral. Res. 37, 2277–2287 (2022).
Lynch, M. E. et al. Tibial compression is anabolic in the adult mouse skeleton despite reduced responsiveness with aging. Bone 49, 439–446 (2011).
Galea, G. L. et al. Old age and the associated impairment of bones’ adaptation to loading are associated with transcriptomic changes in cellular metabolism, cell-matrix interactions and the cell cycle. Gene 599, 36–52 (2017).
Birkhold, A. I. et al. The influence of age on adaptive bone formation and bone resorption. Biomaterials 35, 9290–9301 (2014).
Manolagas, S. C. & Parfitt, A. M. What old means to bone. Trends Endocrinol. Metabolism. 21, 369–374 (2010).
Shum, L. C. et al. Cyclophilin D Knock-out mice show enhanced resistance to osteoporosis and to metabolic changes observed in aging bone. PLoS ONE. 11, 1–18 (2016).
Fritton, J. C., Myers, E. R., Wright, T. M. & van der Meulen, M. C. H. Loading induces site-specific increases in mineral content assessed by microcomputed tomography of the mouse tibia. Bone 36, 1030–1038 (2005).
Main, R. P., Lynch, M. E. & van der Meulen, M. C. H. Load-induced changes in bone stiffness and cancellous and cortical bone mass following tibial compression diminish with age in female mice. J. Exp. Biol. 217, 1775–1783 (2014).
Nandy, A. et al. Altered osteoblast metabolism with aging results in lipid accumulation and oxidative stress mediated bone loss. Aging Disease. 15, 767 (2024).
Lemma, S. et al. Energy metabolism in osteoclast formation and activity. Int. J. Biochem. Cell. Biology. 79, 168–180 (2016).
Williams, J. P. et al. Regulation of osteoclastic bone resorption by glucose. Biochem. Biophys. Res. Commun. 235, 646–651 (1997).
Guntur, A. R., Le, P. T., Farber, C. R. & Rosen, C. J. Bioenergetics during calvarial osteoblast differentiation reflect strain differences in bone mass. Endocrinology 155, 1589–1595 (2014).
Lee, W. C., Ji, X., Nissim, I. & Long, F. Malic enzyme couples mitochondria with aerobic glycolysis in osteoblasts. Cell. Rep. 32, 108108 (2020).
Riddle, R. C. & Clemens, T. L. Bone cell bioenergetics and skeletal energy homeostasis. Physiol. Rev. 97, 667–698 (2017).
Chlebek, C. & Rosen, C. J. The role of bone cell energetics in altering bone quality and strength in health and disease. Curr. Osteoporos. Rep. 21, 1–10 (2023).
Dobson, P. F. et al. Mitochondrial dysfunction impairs osteogenesis, increases osteoclast activity, and accelerates age related bone loss. Sci. Rep. 10, 1–14 (2020).
Esen, E., Lee, S. Y., Wice, B. M. & Long, F. PTH promotes bone anabolism by stimulating aerobic glycolysis via IGF signaling. J. Bone Min. Res. 30, 1959–1968 (2015).
DeMambro, V. E., Tian, L., Karthik, V., Rosen, C. J. & Guntur, A. R. Effects of PTH on osteoblast bioenergetics in response to glucose. Bone Rep. 19, 101705 (2023).
Esen, E. et al. WNT-LRP5 signaling induces Warburg effect through mTORC2 activation during osteoblast differentiation. Cell Metabol. 17, 745–755 (2013).
Hollenberg, A. M., Smith, C. O., Shum, L. C., Awad, H. & Eliseev, R. A. Lactate dehydrogenase inhibition with oxamate exerts bone anabolic effect. J. Bone Miner. Res. 35, 2432–2443 (2020).
Zaman, G. et al. Loading-related regulation of gene expression in bone in the contexts of Estrogen deficiency, lack of Estrogen receptor α and disuse. Bone 46, 628–642 (2010).
Kelly, N. H., Schimenti, J. C., Ross, F. P. & van der Meulen, M. C. H. Transcriptional profiling of cortical versus cancellous bone from mechanically-loaded murine tibiae reveals differential gene expression. Bone 86, 22–29 (2016).
Ghosh, A. et al. Mitochondrial pyruvate carrier regulates autophagy, inflammation, and neurodegeneration in experimental models of Parkinson’s disease. Sci. Transl. Med. 8, (2016).
Papaconstantinou, J. & Colowick, S. P. The role of glycolysis in the growth of tumor cells. J. Biol. Chem. 236, 285–288 (1961).
Lu, Q. Y., Zhang, L., Yee, J. K., Go, V. L. W. & Lee, W. N. Metabolic consequences of LDHA inhibition by epigallocatechin gallate and oxamate in MIA PaCa-2 pancreatic cancer cells. Metabolomics 11, 71–80 (2015).
Patel, T. K., Brodt, M. D. & Silva, M. J. Experimental and finite element analysis of strains induced by axial tibial compression in young-adult and old female C57Bl/6 mice. J. Biomech. 47, 451–457 (2014).
Welhaven, H. D. et al. The cortical bone metabolome of C57BL / 6J mice is sexually dimorphic. JBMR Plus 6, (2022).
Lu, W., Duan, Y., Li, K., Qiu, J. & Cheng, Z. Glucose uptake and distribution across the human skeleton using state-of-the-art total-body PET/CT. Bone Res. 11, 36 (2023).
Zoch, M. L., Abou, D. S., Clemens, T. L., Thorek, D. L. J. & Riddle, R. C. In vivo radiometric analysis of glucose uptake and distribution in mouse bone. Bone Res. 4, 16004 (2016).
Frikha-Benayed, D., Basta-Pljakic, J., Majeska, R. J. & Schaffler, M. B. Regional differences in oxidative metabolism and mitochondrial activity among cortical bone osteocytes. Bone 90, 15–22 (2016).
Jande, S. S. Fine structural study of osteocytes and their surrounding bone matrix with respect to their age in young chicks. J. Ultrasructure Res. 37, 279–300 (1971).
Schilling, K., Brown, E. & Zhang, X. NAD(P)H autofluorescence lifetime imaging enables single cell analyses of cellular metabolism of osteoblasts in vitro and in vivo via two-photon microscopy. Bone 154, 116257 (2022).
Boivin, G., Anthoine-Terrier, C. & Obrant, K. J. Transmission electron microscopy of bone tissue: A review. Acta Orthop. Scand. 61, 170–180 (1990).
Rooney, A. M., McNeill, T. J., Ross, F. P., Bostrom, M. P. G. & van der Meulen, M. C. H. PTH treatment increases cortical bone mass more in response to compression than tension in mice. J. Bone Mineral. Res. Jbmr 4728. https://doi.org/10.1002/jbmr.4728 (2022).
Rooney, A. M. et al. Loading modality and age influence teriparatide-induced bone formation in the human femoral neck. Bone 136, 115373 (2020).
Rooney, A. M. et al. Effects of teriparatide and loading modality on modeling-based and remodeling-based bone formation in the human femoral neck. Bone 157, 116342 (2022).
Komarova, S. V., Ataullakhanov, F. I. & Globus, R. K. Bioenergetics and mitochondrial transmembrane potential during differentiation of cultured osteoblasts. Am. J. Physiol. - Cell. Physiol. 279, 1220–1229 (2000).
Zhu, B. et al. Inhibition of the mitochondrial pyruvate carrier simultaneously mitigates hyperinflammation and hyperglycemia in COVID-19. Sci. Immunol. 8, eadf0348 (2023).
Zheng, C. X., Sui, B. D., Qiu, X. Y., Hu, C. H. & Jin, Y. Mitochondrial regulation of stem cells in bone homeostasis. Trends Mol. Med. 26, 89–104 (2020).
Lemma, S. et al. Energy metabolism in osteoclast formation and activity. Int. J. Biochem. Cell Biol. 79, 168–180 (2016).
Park-Min, K. H. Metabolic reprogramming in osteoclasts. Semin Immunopathol. 41, 565–572 (2019).
Misra, B. B., Jayapalan, S., Richards, A. K., Helderman, R. C. M. & Rendina-Ruedy, E. Untargeted metabolomics in primary murine bone marrow stromal cells reveals distinct profile throughout osteoblast differentiation. Metabolomics 17, 86 (2021).
Welhaven, H. D. et al. Metabolomic profiles of cartilage and bone reflect tissue type, radiography-confirmed osteoarthritis, and Spatial location within the joint. Biochem. Biophys. Res. Commun. 703, 149683 (2024).
Fujino, Y., Minamizaki, T., Yoshioka, H., Okada, M. & Yoshiko, Y. Imaging and mapping of mouse bone using MALDI-imaging mass spectrometry. Bone Rep. 5, 280–285 (2016).
Palmier, M. et al. Osteocyte gene expression analysis in mouse bone: optimization of a laser-assisted microdissection protocol. JBMR Plus 8, (2024).
Poudel, S. B. et al. Canagliflozin-induced adaptive metabolism in bone. Diabetes db240955 https://doi.org/10.2337/db24-0955 (2025).
Farhana, A. & Lappin, S. L. Biochemistry, Lactate Dehydrogenase (StatPearls Publishing, 2021).
Lynch, M. E. et al. Cancellous bone adaptation to tibial compression is not sex dependent in growing mice. J. Appl. Physiol. 109, 685–691 (2010).
Kelly, N. H., Schimenti, J. C., Ross, F. P. & van der Meulen, M. C. H. A method for isolating high quality RNA from mouse cortical and cancellous bone. Bone 68, 1–5 (2014).
Tandonnet, S. & Torres, T. T. Traditional versus 3′ RNA-seq in a non-model species. Genomics Data. 11, 9–16 (2017).
Dobin, A. et al. Ultrafast universal RNA-seq aligner. Bioinf. 29. STAR, 15–21 (2013).
Robinson, M. D., Mccarthy, D. J. & Smyth, G. K. EdgeR: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140 (2010).
Rath, S. et al. MitoCarta3.0: an updated mitochondrial proteome now with sub-organelle localization and pathway annotations. Nucleic Acids Res. 49, D1541–D1547 (2021).
Subramanian, A. et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. U.S.A. 102, 15545–15550 (2005).
Divakaruni, A. S. et al. Thiazolidinediones are acute, specific inhibitors of the mitochondrial pyruvate carrier. Proc. Natl. Acad. Sci. U.S.A. 110, 5422–5427 (2013).
Schneider, C. A., Rasband, W. S. & Eliceiri, K. W. NIH image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671–675 (2012).
Jepsen, K. J., Silva, M. J., Vashishth, D., Guo, X. E. & Van Der Meulen, M. C. H. Establishing biomechanical mechanisms in mouse models: practical guidelines for systematically evaluating phenotypic changes in the diaphyses of long bones. J. Bone Miner. Res. https://doi.org/10.1002/jbmr.2539 (2015).
Turner, C. H. & Burr, D. B. Basic biomechanical measurements of bone: a tutorial. Bone 14, 595–608 (1993).
Melville, K. M. et al. Female mice lacking Estrogen receptor-alpha in osteoblasts have compromised bone mass and strength. J. Bone Mineral. Research: Official J. Am. Soc. Bone Mineral. Res. 29, 370–379 (2014).
Bankhead, P. et al. QuPath: open source software for digital pathology image analysis. Sci. Rep. 7, 1–7 (2017).
Foldes, J., Shih, M. S. & Parfitt, A. M. Frequency distributions of tetracycline‐based measurements: implications for the interpretation of bone formation indices in the absence of double‐labeled surfaces. J. Bone Miner. Res. 5, 1063–1067 (1990).
Applied Linear Statistical Models. (McGraw-Hill Irwin, 2005).
Acknowledgements
We thank the Cornell CARE staff. Funding provided by NSF Grant #CBET-1605935, NIH R21-AR071587, and NSF GRFP (DGE-1650441). We thank Dr. Clifford Rosen and Dr. Anyonya Guntur for their help in experimental design and data interpretation. The sponsors had no role in the writing of the manuscript or in the decision to submit the manuscript for publication.
Author information
Authors and Affiliations
Contributions
Concept and design: CC, MCHM; Acquisition, analysis, and interpretation of data: CC, TJM, MH, MSR, MCHM; Drafting and critical revision of article: CC, TJM, MH, MSR, MCHM; Final approval of article: CC, TJM, MH, MSR, MCHM.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Chlebek, C., McNeill, T.J., Huang, M. et al. Bioenergetic programs of cancellous and cortical bone are distinct and differ with age and mechanical loading. Sci Rep 15, 19134 (2025). https://doi.org/10.1038/s41598-025-02141-5
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-02141-5
