
Abstract
Osteosarcoma (OS) is a highly aggressive bone cancer in children and adolescents with extremely poor prognosis. Gambogic acid (GA) is a natural compound derived from the resin of Garcinia hanburyi trees and has been reported to possess multiple antitumor effects. However, the comprehensive mechanism of GA intervention in OS remains unclear. In this study, network pharmacology and experiment pharmacology were employed to elucidate the mechanism by which GA inhibited OS. Firstly, GA exhibited significant inhibitory effects on OS in vitro and in vivo. Then, the network pharmacological analysis and molecular docking predicted 9 targets involved in necroptosis and apoptosis induced by GA in OS. The qRT-PCR and western blotting verified that GA could increase the apoptosis-related mRNA and protein expression levels, such as cleaved caspase-3, cleaved PARP, indicating that GA could activate apoptosis. Furthermore, proteins related to necroptosis such as RIPK1, RIPK3, and p-RIPK3 were upregulated, while MLKL initially increased and then decreased, implying a dynamic regulation of necroptosis signaling. In summary, our results identified potential targets and pathways of GA to exert anti-osteosarcoma effect, suggesting that GA is a candidate drug for the treatment of osteosarcoma.
Similar content being viewed by others
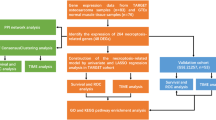


Introduction
OS is a malignant tumor that often occurs in children and adolescents. It is characterized by primary bone and easy to metastasize1,2. Nearly 75% of newly diagnosed OS patients are aged 15–25 years, and OS is slightly more common in males. Distant metastases occur in approximately 20% of OS cases, with 60–70% to the lung and 20–30% to the bone1. Currently, the treatment of primary OS is mainly chemotherapy, radiotherapy and surgery3. However, once OS progresses to an advanced stage, treatment options are very limited, and drug resistance is a key factor leading to treatment failure and tumor recurrence4. Generally, the use of large-dose chemotherapy is beneficial for treatment and five-year survival of OS, but may cause serious adverse reaction5. Therefore, it is of great importance to find potently safe and effective drugs against OS.
Traditional Chinese medicine has great potential in improving the quality of life of OS patients. GA is a natural compound derived from the resin of Garcinia hanburyi trees (Guttiferae)6. It has multipotent pharmacological activities, such as anti-inflammatory, antibacterial and antioxidant. Notably, it has also demonstrated strong anti-tumor activity on many human tumors, including liver, gastric, lung, pancreatic, ovarian and prostate cancer7. However, the comprehensive mechanism of GA in OS still remains unclear with limited research at the molecular level.
Network pharmacology is frequently applied for systematic screening potential bio-targets and predicting molecular mechanisms of drug monomers and compounds. Its unique multi-target efficacy, multi-pathways, and even multi-component interactions facilitate the study of the holistic nature of drugs on diseases8,9. In anti-OS studies, researchers have searched for potential targets and predicted molecular mechanisms through network pharmacological analyses. Previous studies have identified cyclin-dependent kinases (CDK1), tumor protein P53 (TP53), CASP3 and X-linked inhibitor of apoptosis protein (XIAP) as potential targets against osteosarcoma and verified the protein and mRNA expression levels in osteosarcoma cell lines or clinical patient samples10,11. Besides, network pharmacology has been also performed to explore the molecular mechanism of GA in endometrial cancer12.
In this study, we aimed to investigate the underlying antitumor mechanism of GA on osteosarcoma through network pharmacology and experiment pharmacology. Firstly, in vitro and in vivo antitumor effects of GA on OS were investigated. Then, network pharmacology was conducted to predict potent antitumor pathways, and the interaction between target genes and GA was explored via molecular docking. Furthermore, we verified the predicted core targets of GA on OS cell lines via experiment pharmacology. This study provides a theoretical basis for the clinical treatment of OS using GA.
Results
GA inhibited OS proliferation in vitro and in vivo
To assess the effect of GA on OS cell proliferation, we performed CCK8 and coony formation. The results of CCK8 showed that GA could inhibit proliferation of multiple osteosarcoma cell lines (143B, U2Os, MG63, HOS) in a concentration-dependent manner. The IC50 of GA on 143B, U2Os, MG63, HOS cells were 0.37 ± 0.02 μM, 0.32 ± 0.06 μM, 0.51 ± 0.02 μM, 0.60 ± 0.11 μM, respectively (Fig. 1A). Since U2Os and 143B cell lines were more sensitive to GA than other two cell lines, these two cell lines were selected for subsequent experiments. In the colony formation assay, we observed that 0.2 μM and 0.4 μM GA significantly suppressed colony formation of 143B cell after 7 days treatment, while 0.1 μM, 0.2 μM and 0.4 μM GA significantly suppressed colony formation of U2Os cell after 10 days of treatment (Fig. 1B, C). In the Fig. 1D, OS cells presented cytoplasmic vacuolation after treatment with GA (0.2, 0.4 and 0.6 μM). The above results indicated that GA could inhibit OS proliferation in vitro.

In vitro antitumor effect of GA on OS. (A) U2Os, 143B, MG63 and HOS cells were treated with different concentrations of GA (0.0625, 0.125, 0.25, 0.5, 1 and 2 μM) for 48 h, cell viability was detected by CCK8 assay and the IC50 values were calculated (n = 3, *P < 0.05). (B) 143B and U2Os cells were seeded in a 6-well plate at 100 cells/well for 7 days or at 400 cells/well for 10 days to form colonies after treatment with indicated concentrations of GA (0, 0.025, 0.05, 0.1, 0.2 and 0.4 μM). (C) Quantify analysis of colony formation (*P < 0.05, **P < 0.01, n = 3). (D) Morphological observation of 143B and U2Os treated with GA (0.2, 0.4, 0.6 μM) for 24 h.
We further investigated the in vivo antitumor effects of GA in OS. 1 mg/kg, 2 mg/kg GA and 3 mg/kg cisplatin all significantly reduced tumor weight and size (Fig. 2A, B), indicating that GA could suppress OS proliferation in vivo. Compared with control group, the body weight of mice in the 1 mg/kg GA group did not change significantly, while the body weight of mice in the 2 mg/kg GA group and 3 mg/kg cisplatin group decreased significantly (Fig. 2C).

GA inhibited OS growth in vivo. (A) Representative photographs of tumors after treatment with PBS, cisplatin (3 mg/kg), GA (1 mg/kg) and GA (2 mg/kg) (n = 5). (B) Tumor weight in each group (n = 5, #P < 0.05, cisplatin (3 mg/kg) vs solvent group (NC); *P < 0.05, GA treated groups vs NC). (C) Body weight variation of nude mice after different treatments (n = 7 or 8, ##P < 0.01, cisplatin (3 mg/kg) vs NC; **P < 0.01, GA treated groups vs NC).
Network pharmacological analysis
Collection of potential targets of OS and GA
A total of 3554 OS target genes were retrieved from the GeneCard and OIMI databases. 100 GA targets genes were obtained by searching the Swiss Target Prediction database. With the probability > 0 as the reference value, we selected 50 targets that intersected with the obtained OS target genes to obtain 32 common target genes and drew a Venn diagram (Fig. 3A). These 32 target common genes were namely IKBKB, TXN, TXN2, MCL1, carbonic anhydrase 2 (CA2), AGTR1, carbonic anhydrase 2 (CA2), carbonic anhydrase 9 (CA9), 3-oxo-5-alpha-steroid 4-dehydrogenase 1 (SRD5A1), cysteinyl leukotriene receptor 1 (CYSLTR1), 3-oxo-5-alpha-steroid 4-dehydrogenase 2 (SRD5A2), STAT3, STAT5B, STAT1, cathepsin D (CTSD), caspase-1 (CASP1), integrin beta-1 (ITGB1), thymidylate synthase (TYMS), tyrosinase (TYR), SERPINE1, FKBP1A, TNFRSF10A, arachidonate 5-lipoxygenase-activating protein (ALOX5AP), calcitonin receptor-like receptor (CALCRL), integrin alpha V (ITGAV), cholecystokinin type B receptor protein (CCKBR), receptor-type tyrosine-protein phosphatase S (PTPRS), CASP3, MME, prostaglandin E2 receptor EP1 subtype (PTGER1), integrin α4 (ITGA4) and integrin beta-3 (ITGB3), which were considered as potential therapeutic targets.

Collection of potential targets of GA and OS, and protein–protein interaction (PPI). (A) Vennu digram for searched targets in GA and OS. (B) PPI network. The collective proteins (or genes) were constructed a protein–protein interaction network. The nodes represented targets and the connected lines represented interactions between different target proteins. (C) Bar diagram for numbers of target proteins connected other target proteins. The diagram showed that the top 25 targets that the most quantity of linking with other targets. X-axis represents the numbers of connected other target proteins. Y axis represents the target proteins title.
Construction of PPI network
The PPI network was obtained by searching the common 32 target genes in the STRING database (Fig. 3B). The PPI network showed the complex interactions between targets, in which the nodes represent the targets and the connected lines represent interactions between different proteins. The histogram of the top 25 targets was finally obtained by importing the PPI network data from the STRING database into an edited R program. The results exhibited that the top 25 targets had the highest number of connections with other targets (Fig. 3C).
GO analysis and KEGG pathway enrichment analysis
In the GO analysis, the biological processes (BP), cellular components (CC) and molecular functions (MF) of the 32 target genes were analyzed. On the basis of BP, CC and MF, 213, 26, and 42 GO terms were obtained, respectively. The top 20 GO analyses are showed in Fig. 4A, B and C. In the KEGG analysis, 53 KEGG terms were acquired. The top 20 signaling pathways were presented as a bar plot (Fig. 4D). The results predicted that GA may have a significant impact on the apoptosis and necroptosis of OS. In light of the above PPI Network data (Fig. 3B, C), GA may potentially treat OS by acting on key targets involved in apoptosis and necroptosis, including CASP3, CASP1, STAT3, STAT1, MCL1, IKBKB, STAT5B, TNFRSF10A, and CTSD. Therefore, two critical pathways involved in apoptosis and necroptosis were selected for the following experiments.

GO and KEGG analyses of GA’s targeted genes. (A–C) Biological processes (BP) (A), cellular components (CC) (B) and molecular functions (MF) (C) of the 32 target genes. X-axis represents the different values of − log10(p). Y-axis represents the names of BP, CC and MF, respectively. (D) KEGG pathway enrichment. X-axis represents the ratio of enriched target proteins/background proteins. Y-axis represents the term of enriched pathways. The colors of the bar chart reflect the different values of − log10(p). (E) “GA-Target-Pathway-OS” Network.
Construction for “GA-target-pathway-OS” network
A “GA-Target-Pathway-OS” diagram was further constructed by importing the common target genes of GA and OS as well as the enriched KEGG pathways into the Cytoscape software. As shown in Fig. 4E, the pathway abbreviations in the network diagram are annotated as the specific pathway named in Table S1. The larger the shape of the target, the more pathways it represents its own influence on. The results showed that IKBKB and CASP3 affect most of the pathways in the top 20 most significant pathways.
Molecular docking between GA and the targets related to apoptosis and necroptosis
In order to verify whether GA acted on the above mentioned main targets, molecular docking between nine targets and GA was conducted. As shown in Fig. 5, GA could enter and bind the active pockets of the nine target proteins. The docking scores and amino acid residues were shown in Table 1. It was shown that the relevant targets of GA on apoptosis and necroptosis may be CASP3, CASP1, MCL-1, STAT3, STAT1, IKBKB, CSTD, STAT5B, and TNFRSF10A. Among them, MCL-1 has strong binding activity with GA, while STAT3, CTSD, STAT5B, STAT1 and CASP3, exhibits good binding activity. IKBKB, CASP1, and TNFRSF10A have certain binding activity with GA.

Molecular docking between GA and the targets related to apoptosis and necroptosis.
GA inhibited OS cell proliferation via inducing apoptosis
Tumors are new masses formed by unlimited cell proliferation, and induction of cell apoptosis is a direct antitumor method. As shown in Fig. 6A and B, with the increase of GA concentrations, the number of total cells decreased and the number of cells with intense blue fluorescence increased, indicating that the apoptosis rates increased after treatment with GA for 12 h, especially at the concentration of 0.6 μM. To further quantify these effects, flow cytometry (FCM) was employed to assess apoptosis rates in OS cells treated with varying GA concentrations. Compared with the control group, the apoptotic rates of 143B cells significantly increased to about 21.54% and 36.53% at GA concentrations of 0.4 and 0.6 μM, respectively (Fig. 6B and C). Similarly, the apoptotic rates of U2Os cells significantly increased to about 24.99% and 41.80% at the same GA concentrations (Fig. 6B and D). The above results manifested that GA induced apoptosis in both 143B and U2Os cells in a dose-dependent manner.

GA inhibited OS proliferation via inducing apoptosis. (A) The cell death of 143B or U2Os treated with GA were evaluated via Hoechst staining assay (Scale bars: 100 or 50 μm). (B) Flow cytometry was used to analyze apoptosis rate of 143B and U2Os treated with different concentrations of GA for 24 h. (C–D) Quantify analyses of apoptosis and necroptosis rate of 143B (C) and U2Os (D) cells via flow cytometry (*P < 0.05, n = 3).
The proteins involved in the GA-induced apoptosis process were detected by qRT-PCR and western blotting (WB). The apoptosis-related proteins CTSD, CASP3, and IKBKB, which modulate apoptotic signaling directly or indirectly13,14,15, exhibited dose-dependent upregulation at the mRNA level upon GA treatment (Fig. 7A and B). Furthermore, the ratio of Bax/Bcl-2, a key indicator of mitochondrial outer membrane permeabilization16, increased in a concentration-dependent manner (Fig. 7C–F). This indicated that GA could initiate apoptosis by inducing mitochondrial dysfunction. Subsequent caspase cascade activation led to the cleavage of Poly(ADP-Ribose) polymerase (PARP) and caspase-3, further confirming the occurrence of apoptosis. Consistent with these findings, IHC analysis of tumor tissues from nude mice revealed increased expression of cleaved PARP and cleaved caspase-3 (Fig. S1), reinforcing the pro-apoptotic role of GA in vivo.

mRNA transcription and protein expression of apoptosis induced by GA. (A–B) qRT-PCR measurement of apoptosis related protein mRNA expression in 143B and U2Os cells treated with indicated concentration of GA (*P < 0.05, **P < 0.01, n = 3). (C) WB analysis of Bcl-2, Bax, cleaved caspase 3 (clv. caspase 3) and cleaved PARP (clv. PARP) in 143B cell after treatment with GA for 24 h. (D) Quantitative analysis of the WB in 143B cell (*P < 0.05, **P < 0.01, n = 3). (E) WB analysis of Bcl-2, Bax, clv. caspase 3 and clv. PARP in U2Os cell after treatment with GA for 24 h. (F) Quantitative analysis of the WB in U2Os cell (*P < 0.05, **P < 0.01, n = 3).
GA may suppress OS growth via inducing necroptosis
Given that the PPI network and FCM data demonstrate that GA may induce necroptosis in OS, we investigated this mechanism using qRT-PCR and WB analysis. Necroptosis, a regulated form of programmed cell death, involves the recruitment of RIPK1, RIPK3, and MLKL to complex II. Upon activation, RIPK3 phosphorylates MLKL, a crucial step for initiating necroptosis, leading to MLKL oligomerization, pore formation, and subsequent cell membrane disruption14,17. Additionally, it has been reported that STAT5 and STAT1 play important roles in necroptosis via interferons (IFNs)-mediated pathway18,19. As illustrated in Fig. 8A and 8B, the mRNA expression levels of STAT1 and STAT5B positively correlated with GA concentrations, indicating that GA may activate the necroptosis. At the protein level, RIPK1, RIPK3 and phospho-RIPK3 expression increased with higher GA concentrations (Fig. 8C–E), while MLKL and phospho-MLKL (p-MLKL) levels decreased. This observation appeared inconsistent with the canonical necroptosis pathway, where p-MLKL, the executioner protein, typically accumulates. To further explore this discrepancy, we examined the temporal dynamics of p-MLKL and cleaved PARP. Time-course experiments revealed that p-MLKL initially increased but later declined, whereas cleaved PARP levels progressively rose (Fig. S2). These findings suggest that GA may initially induce necroptosis in OS cells but eventually shifts toward apoptosis as the dominant cell death mechanism in OS cells.

GA inhibited tumor growth via inducing necroptosis in OS. (A–B) qRT-PCR measurements of necroptosis related protein mRNA expression in 143B (A) and U2Os (B) cells treated with GA (*P < 0.05, **P < 0.01, n = 3). (C) WB analysis of RIPK3, phosphor-RIPK3 (p-RIPK3), RIPK1, MLKL and phosphor-MLKL (p-MLKL) expression in 143B cells after treatment with GA for 24 h. (D) Quantitative analysis of the WB in 143B cell (*P < 0.05, **P < 0.01, n = 3). (E) WB analysis of RIPK3, p-RIPK3, RIPK1, MLKL and p-MLKL expression in U2Os cells after treatment with GA for 24 h. (F) Quantitative analysis of the WB in U2Os (*P < 0.05, **P < 0.01, n = 3).
Discussion
Osteosarcoma is a highly malignant bone tumor with poor prognosis1. Despite significant improvements in survival rates in recent years, 80% of patients with OS already have local invasion and distant metastases when diagnosed, and this metastasis will cause death in 70% of patients with OS20,21. Moreover, high-dose chemotherapy can ameliorate 5-year survival in patients with early-stage non-metastatic osteosarcoma22. However, it may cause severe side effects and lead to drug resistance in patients23. Notably, chemotherapy resistance is a key factor for treatment failure and tumor recurrence. Therefore, the exploration of sensitive drugs for the treatment of OS has become one of the hot spots in current research.
GA was a molecule with a xanthone skeleton and has been reported to have anti-OS activity24,25. However, the underlying mechanism is still unknown. The existing research on its anti-osteosarcoma effect is still limited to inducing cell apoptosis, overcoming cisplatin resistance, and its effects on matrix metallopeptidase 9 (MMP-9), and metalloproteinase inhibitor 1 (TIMP-1)26,27,28,29. Therefore, more studies on the mechanisms and targets of anti-OS in vivo and in vitro are urgently needed. Monomer derived from traditional Chinese medicine or natural medicine often have multiple targets and signaling pathways that act on diseases for more beneficial treatment effects, but this challenges pharmacological research and exploitation30,31,32. Network pharmacology can be applied to explore the role of one monomer in complex diseases, thus screening for key target proteins that orchestrate the regulation of diseases33,34. In this study, we firstly analyzed the underlying mechanism of GA in OS based on network pharmacology and molecular docking. The results demonstrated that GA possibly induced apoptosis through IKBKB, MCL1, CTSD, TNFRSF10A, and CASP3, as well as necroptosis by acting on STAT3, STAT5B, STAT1, CASP1 (Figs. 3, 4 and 5). However, since PPI, GO, and KEGG analysis originate from the permutation and combination of targets, they are not very precise and often require further experimental screening and verification.
Necroptosis is an alternative mode of regulated cell death that could simulate apoptosis and necrosis features, typically triggered by TNF superfamily ligands, Toll-like receptor ligands or IFN signaling14,35. The IFNs/IFNR-JAK/STATs axis has been identified as a crucial regulator. IFNs transcriptionally activate the RNA-responsive protein kinase PKR through the JAK1/STAT-mediated pathway, which subsequently interacts with RIPK1 to initiate the assembly of the RIPK1-RIPK3 “necrosome” complex, ultimately triggering necroptosis35. Our findings revealed that GA upregulated STAT1 and STAT5B mRNA levels (Fig. 8A, B), strongly suggesting its potential to engage the IFN/STAT signaling axis. This observation aligns with prior reports that STAT1 and STAT5B are critical mediators of IFN-driven necroptosis in cancer18,19. Notably, GA induced the elevated expression of RIPK1, RIPK3, and p-RIPK3 (Fig. 8C), the unexpected decline in p-MLKL implied a non-canonical regulation of necroptosis execution. This apparent paradox may reflect a temporal competition between cell death pathways. The observed inverse correlation between declining p-MLKL and rising cleaved PARP (Fig. S2), implies a dynamic shift in cell death modalities-early necroptosis may give way to apoptosis during prolonged GA exposure. This phenomenon echoes findings that autophagy machinery p62/SQSTM1 could orchestrate a “death switch” between necroptosis and apoptosis36. Specifically, p62-mediated RIPK1 recruitment favors necrosome formation, whereas p62 inhibition redirects cell fate toward an apoptosis paradigm that could explain GA’s biphasic effects. Crucially, GA’s pro-apoptotic effects were independently confirmed through mitochondrial pathway activation, as evidenced by an elevated Bax/Bcl-2 ratio and levels of cleaved caspase-3/PARP, along with the upregulation of apoptotic mediators such as CTSD, IKBKB, and CASP3 (Fig. 7 and S1). This dual engagement of necroptotic and apoptotic machinery positions GA as a multifaceted cytotoxic agent capable of exploiting interconnected cell death networks in OS.
Conclusions
In summary, this study systematically elucidated the potential mechanism of GA against OS based on network pharmacology and experiment verification. Our data demonstrated that GA could inhibit osteosarcoma growth via apoptosis and necroptosis. The biphasic cell death regulation, characterized by an initial necroptotic response followed by apoptotic dominance, suggests GA’s unique pharmacological profile as a promising therapeutic candidate for OS treatment. Nevertheless, further studies are warranted to delineate the precise molecular crosstalk between GA-induced necroptosis and apoptosis, and identify the specific cellular targets mediating these effects. These investigations will provide a more comprehensive understanding of GA’s therapeutic potential and facilitate its clinical translation for OS management.
Materials and methods
Cell culture
Human osteosarcoma cancer cell line U2Os (CL-0236) was obtained from Procell Life Science & Technology (Hubei, China). 143B (CRL-8303), HOS (CRL-1543) and MG63 (CRL-1427) were purchased from Hunan Fenghui Biotechnology (Hunan, China). U2Os was cultured in Myco’s 5A Medium (Procell Life Technologies, PM150710) supplemented with 10% FBS (Gibco, 10,270–106) and 1% penicillin/streptomycin (Gibco, 15,140–122). 143B, and HOS were cultured in α-MEM Medium (containing NEAA) (Procell Life Technologies, PM150410) with 10% FBS (Gibco), and 1% penicillin/streptomycin (Gibco). MG63 was cultured in MEM Medium (Procell Life Technologies, PM150411) without NEAA supplemented with 10% FBS (Gibico) and 1% penicillin/streptomycin (Gibico). The cells were all cultured at 37 °C under a humidified atmosphere of 5% CO2.
Reagents and antibody
Antibodies for cleaved-PARP (AF7023), RIPK3 (AF7942), phospho-MLKL (AF7420), β-actin (AF7018), Bax (AF0120) and Bcl-2 (AF6139) were obtained from Affinity Biosciences LTD. MLKL (14,993), RIPK1 (3493), cleaved-caspase3 (9664), Phospho-RIPK3 (93,654) and the secondary antibodies with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG were obtained from Cell Signaling Technology (CST, Danvers, MA, USA). BCA protein assay kit (23,225) and Annexin V-FITC/PI apoptosis detection kit (MA0220-Nov-19G) were bought from Thermo Fisher (Waltham, MA, USA) and Meilun Biotechnology (Liaoning, China) respectively. TB Green Premix Ex Tap™ (RR420) and PrimeScript™ RT Master Mix (RR036A) were obtained from Takara. Primers were gained from Sangon Biotech (Shenzhen). Cisplatinum (MedChemExpress (MCE), CAS: 15663–27-1) was dissolved in PBS, gambogic acid (GA, CAS: 2752–65-0, HPLC ≥ 98%) and C16 (MedChemExpress (MCE), CAS: 608512–97-6) were dispersed in dimethyl sulfoxide (DMSO).
Cell viability assay
GA was diluted to 2 μM, 1 μM, 0.5 μM, 0.25 μM, 0.125 μM and 0.0625 μM with complete medium after dissolved in DMSO, respectively. 143B, MG63 cells were seeded in 96-well plates at a density of 2 × 103 cells/ well, while U2Os and HOS were seeded at a density of 2.5 × 103 cells/ well. The cells were incubated with GA for 48 h after overnight growth. Cell Counting Kit (CCK-8, MA0218-3-Jan-23G) (Meilun Biotechnology, China) was added and absorbance was measured at 450 nm on a microplate reader (Thermo Scientific, USA).
Morphological observation
GA was diluted to 0.6 μM with complete medium after dissolved in DMSO. 143B, U2Os cells were seeded in 96-well plates at a density of 4 × 103 cells/ well and 6 × 103 cells/ well, respectively. The cells were incubated with GA for 24 h after overnight growth.
Western blotting
The proteins were extracted from cells via RIPA lysis buffer including 1% PMSF and 1% phosphatase inhibitors. Protein concentrations were quantified using a BCA assay kit. Equal amount of proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene difluoride membranes (PVDF) membranes. The membranes were blocked for 15 min with quick blocking fluid. The proteins were banded with primary antibodies (1:1000) overnight at 4℃, and then were incubated HRP-conjugated second antibody (1:10,000) for 1.5 h. The protein signal of membrane were detected with ECL chemiluminescence reagents and were quantified according to the gray-scale values of related proteins by Image J.
qRT-PCR analysis
143B was seeded in a 6-well plate at a density of 2.5 × 105 cells/well for 24 h, and U2Os was seeded in a 6-well at a density of 3.5 × 105 for 24 h. Then both were treated with GA (0, 0.2, 0.4, 0.6 μM). The total RNA was extracted from the cells with Trizol reagent (Cat no.15596–02, Life Technologies). The mRNAs were detected by qRT-PCR assay. The primer sequences utilized as shown in Table S2.
Clone formation assay
143B and U2Os cells were seeded in a 6-well plate at density of 100 cells/well, 400 cells/well, respectively. After 24 h, 143B/U2Os were incubated with 0.4 μM, 0.2 μM, 0.1 μM, 0.05 μM, 0.025 μM, 0 μM GA for 7 days/10 days. Then cells were fixed with 4% paraformaldehyde for 30 min and stained with ammonium oxalate crystal violet dye for 15 min. After staining, the plates were rinsed with distilled water, dried and photographed.
Flow cytometry analysis of apoptosis
Cell death was detected by Annexin V-FITC/PI Apoptosis Dectection Kit (MA0220–Nov–19G, Meilun Biotechnology). 143B cells were seeded in a density of 1 × 105 cells/well overnight and treated with GA for 24 h. Then the cells were suspended in 1 × binding buffer and stained with Annexin V (5 μL) and PI (10 μL) for 15 min at room temperature in the dark. The apoptosis rates were detected with a flow cytometry (Beckman, CytoFLEX, MA, USA).
Hoechst staining
143B cells were seeded in a 24-well plate at a density of 4 × 104 cells/well. U2Os were seeded in a 24-well plate at a density of 6 × 104 cells/well. After overnight growth, cells were incubated with GA (0, 0.2, 0.4, 0.6 mM) for 12 h. Then the cells were fixed, washed with PBS and stained with Hoechst. Cell apoptosis was evaluated by fluorescence microscope.
In vivo xenograft tumor assay and IHC staining
BALB/c-nu male mice (6-week old) were purchased from Zhuhai Bes Test Life Technology (Zhuhai, China). Human 143B cells were inoculated into the back of mice (1 × 106 cells/mouse). The mice were randomly divided into four groups: Normal control (PBS with 1% DMSO treated, n = 7), positive control (cisplatin, 3 mg/kg treated, n = 8), gambogic acid low dose group (1 mg/kg, n = 8) and gambogic acid high dose group (2 mg/kg, n = 8). After continuous administration for 14 days, all mice were euthanized with an overdose of isoflurane, the tumors were excised and then fixed with 4% paraformaldehyde, and stained with cleaved-PARP and cleaved caspase 3 antibodies. Finally, the morphological changes of tissues were observed under an optical microscope.
Collection for target information of osteosarcoma and GA
Osteosarcoma targets were searched from the Gene Cards (https://www.genecards.org/) and OMIM (Online Mendelian Inheritance in Man) (https://omim.org/search/advanced/geneMap) websites. Only genes with a relevance score > 0.7 were included to prioritize high-confidence disease-related targets. Gambogic acid 3D structure was obtained from Pubchem (https://pubchem.ncbi.nlm.nih.gov/) database. Gambogic acid targets were searched from SwissTargetPrediction (http://www.swisstargetprediction.ch/) database and targets with a probability score > 0 were selected. Obtained osteosarcoma targets and Gambogic acid related targets were intersected by VennDisgram package in R version 4.1.2 to make Venn disgram obtaining 32 targets.
Gene ontology and KEGG enrichment analysis
The common targets were converted with the corresponding ID by RSQLite and org.Hs.eg.db in BiocManager packages in R version 4.1.2 to get a text named “id.text”. The text was applied to make a GO pathway enrichment analysis by stringi, ggplot2 and clusterProfiler, org.Hs.eg.db, enrichplot in BiocManager to get the GO barplot on biological processes (BP), cellular components (CC), molecular functions (MF) in R version 4.1.2. The KEGG barplot was obtained by colorspace, stringi and DOSE, clusterProfiler, pathview in BiocManager package to run some codes with the text in R version 4.1.2.
Network construction
The common targets were imported into the String (https://string-db.org/) website to get the PPI network and interactions data on protein interactions. The parameter was set as moderate confidence (0.400). The interactions data were made into a bar chart with taking the first 25 targets. The first 20 pathways (Table 2) and the targets in the pathways got from KEGG analysis were selected to get KEGG pathways to construct “GA-Target-Pathway-OS” network through Cytoscape 3.7.2.
Molecular docking verification
The apoptosis and necroptosis pathway-related targets were screened for subsequent molecular docking analysis. Nine targets Inhibitor of nuclear factor kappa-B kinase (IKBKB), myeloid cell leukemia 1 (MCL-1), cathepsin D (CTSD), tumor necrosis factor receptor superfamily member 10A protein (TNFRSF10A), CASP3, signal transducer and activator of transcription 3 (STAT3), signal transducer and activator of transcription 1 (STAT1), signal transducer and activator of transcription 5 (STAT5), caspase-1 (CASP1) were searched and downloaded 3D structure of the protein in RCSB PDB database (https://www.rcsb. org/). The 3D structure of drug was downloaded from the PubChem database (https://pubchem.ncbi.nlm.nih.gov/). Mastro 3.8 was used to dock the receptor protein with the drug molecule. Finally, PyMOL 3.7.0 software was used to show the active pocket sites where small drug molecule bind.
Statistical analysis
GraphPad Prism 8.0 software was applied for chart production (GraphPad Prism; San Diego, CA, USA). The data was analyzed by IBM SPSS Statistics 23 software. Data from every single experiment were applied by normality test. After passing the normality test, one-way ANOVA and Dunnett’s multiple comparisons tests were successively used for inter-group significance statistics. Otherwise, a non-parametric test is applied to the supplementary test. Each experiment was independently replicated at least three times. P < 0.05 was considered as significant difference.
Data availability
Data is provided within the manuscript or supplementary information files.
References
Gill, J. & Gorlick, R. Advancing therapy for osteosarcoma. Nat. Rev. Clin. Oncol. 18(10), 609–624 (2021).
Belayneh, R., Fourman, M. S., Bhogal, S. & Weiss, K. R. Update on osteosarcoma. Curr. Oncol. Rep. 23(6), 71 (2021).
Heng, M. et al. The role of chemotherapy and radiotherapy in localized extraskeletal osteosarcoma. Eur. J. Cancer 125, 130–141 (2020).
Chen, R., Wang, G., Zheng, Y., Hua, Y. & Cai, Z. Drug resistance-related microRNAs in osteosarcoma: Translating basic evidence into therapeutic strategies. J. Cell. Mol. Med. 23(4), 2280–2292 (2019).
Xiao, X. et al. HSP90AA1-mediated autophagy promotes drug resistance in osteosarcoma. J. Exp. Clin. Cancer Res. 37(1), 201 (2018).
Hatami, E., Jaggi, M., Chauhan, S. C. & Yallapu, M. M. Gambogic acid: A shining natural compound to nanomedicine for cancer therapeutics. Biochim. Biophys. Acta Rev. Cancer 1874(1), 188381 (2020).
Liu, Y., Chen, Y., Lin, L. & Li, H. Gambogic acid as a candidate for cancer therapy: A review. Int. J. Nanomed. 15, 10385–10399 (2020).
Hua, Y. et al. Deciphering the pharmacological mechanism of Radix astragali for allergic rhinitis through network pharmacology and experimental validation. Sci. Rep. 14(1), 29873 (2024).
Bakhsh, H. T. et al. Unveiling Lobophytum sp the neuroprotective potential of Parkinson’s disease through multifaceted mechanisms, supported by metabolomic analysis and network pharmacology. Sci. Rep. 14(1), 21810 (2024).
Wu, B. et al. Selected using bioinformatics and molecular docking analyses, PHA-793887 is effective against osteosarcoma. Aging (Albany NY) 13(12), 16425–16444 (2021).
Lai, H. T. et al. Insight into the interplay between mitochondria-regulated cell death and energetic metabolism in osteosarcoma. Front. Cell Dev. Biol. 10, 948097 (2022).
Xia, Z. & Tang, Z. Network pharmacology analysis and experimental pharmacology study explore the mechanism of gambogic acid against endometrial cancer. ACS Omega 6(16), 10944–10952 (2021).
Liaudet-Coopman, E. et al. Cathepsin D: newly discovered functions of a long-standing aspartic protease in cancer and apoptosis. Cancer Lett. 237(2), 167–179 (2006).
Ai, Y., Meng, Y., Yan, B., Zhou, Q. & Wang, X. The biochemical pathways of apoptotic, necroptotic, pyroptotic, and ferroptotic cell death. Mol. Cell 84(1), 170–179 (2024).
Thomas, G. S., Zhang, L., Blackwell, K. & Habelhah, H. Phosphorylation of TRAF2 within its RING domain inhibits stress-induced cell death by promoting IKK and suppressing JNK activation. Cancer Res. 69(8), 3665–3672 (2009).
Andreu-Fernández, V. et al. Bax transmembrane domain interacts with prosurvival Bcl-2 proteins in biological membranes. Proc. Natl. Acad. Sci. U S A 114(2), 310–315 (2017).
Liu, S. et al. MLKL polymerization-induced lysosomal membrane permeabilization promotes necroptosis. Cell Death Differ. 31(1), 40–52 (2024).
Chen, B. et al. Comprehensive analysis of mitochondrial dysfunction and necroptosis in intracranial aneurysms from the perspective of predictive, preventative, and personalized medicine. Apoptosis : An Int. J. Program. Cell Death 28(9–10), 1452–1468 (2023).
Sumaria, N. et al. Perinatal thymic-derived CD8αβ-expressing γδ T cells are innate IFN-γ producers that expand in IL-7R-STAT5B-driven neoplasms. Nat. Immunol. 25(7), 1207–1217 (2024).
Izadpanah, S. et al. Prospects for the involvement of cancer stem cells in the pathogenesis of osteosarcoma. J Cell Physiol 235(5), 4167–4182 (2020).
Zuo, D. et al. Inhibition of STAT3 blocks protein synthesis and tumor metastasis in osteosarcoma cells. J. Exp. Clin. Cancer Res. 37(1), 244 (2018).
Smeland, S. et al. Survival and prognosis with osteosarcoma: Outcomes in more than 2000 patients in the EURAMOS-1 (European and American Osteosarcoma Study) cohort. Eur. J. Cancer 109, 36–50 (2019).
Da, W. et al. A 10-year bibliometric analysis of osteosarcoma and cure from 2010 to 2019. BMC Cancer 21(1), 115 (2021).
Liu, Z. et al. Gambogenic acid induces cell death in human osteosarcoma through altering iron metabolism, disturbing the redox balance, and activating the P53 signaling pathway. Chem. Biol. Interact. 382, 110602 (2023).
Banik, K. et al. Therapeutic potential of gambogic acid, a caged xanthone, to target cancer. Cancer Lett. 416, 75–86 (2018).
Liu, L., Qi, X. J., Zhong, Z. K. & Zhang, E. N. Nanomedicine-based combination of gambogic acid and retinoic acid chlorochalcone for enhanced anticancer efficacy in osteosarcoma. Biomed. Pharmacother. 83, 79–84 (2016).
Xin, Z. F., Shen, C. C., Tao, L. J., Yan, S. G. & Wu, H. B. Gambogic acid inhibits invasion of osteosarcoma via upregulation of TIMP-1. Int. J. Mol. Med. 31(1), 105–112 (2013).
Zhao, W. et al. Viability inhibition effect of gambogic acid combined with cisplatin on osteosarcoma cells via mitochondria-independent apoptotic pathway. Mol. Cell. Biochem. 382(1–2), 243–252 (2013).
Zhao, W. et al. Gambogic acid inhibits the growth of osteosarcoma cells in vitro by inducing apoptosis and cell cycle arrest. Oncol. Rep. 25(5), 1289–1295 (2011).
Parthasarathy, A., Mantravadi, P. K. & Kalesh, K. Detectives and helpers: Natural products as resources for chemical probes and compound libraries. Pharmacol. Ther. 216, 107688 (2020).
Gao, S. et al. Rabdosia rubescens (Hemsl.) H. Hara: A potent anti-tumor herbal remedy - Botany, phytochemistry, and clinical applications and insights. J. Ethnopharmacol. 340, 119200 (2025).
Gao, S. et al. Unveiling polysaccharides of Houttuynia cordata Thunb.: Extraction, purification, structure, bioactivities, and structure-activity relationships. Phytomed. : Int. J. Phytother. Phytopharmacol. 138, 156436 (2025).
Wang, Y. et al. Erianin, the main active ingredient of Dendrobium chrysotoxum Lindl, inhibits precancerous lesions of gastric cancer (PLGC) through suppression of the HRAS-PI3K-AKT signaling pathway as revealed by network pharmacology and in vitro experimental verification. J. Ethnopharmacol. 279, 114399 (2021).
Gao, J., Yang, S., Xie, G., Pan, J. & Zhu, F. Integrating network pharmacology and experimental verification to explore the pharmacological mechanisms of aloin against gastric cancer. Drug Des. Dev. Ther. 16, 1947–1961 (2022).
Thapa, R. J. et al. Interferon-induced RIP1/RIP3-mediated necrosis requires PKR and is licensed by FADD and caspases. Proc. Natl. Acad. Sci. U S A 110(33), E3109–E3118 (2013).
Goodall, M. L. et al. The autophagy machinery controls cell death switching between apoptosis and necroptosis. Dev. Cell. 37(4), 337–349 (2016).
Acknowledgements
We are very grateful for the contributions of the Pubchem database, symMap database, the DisGeNET database and the GeneCards database that provide information on cancer research, as well as all colleagues involved in the study.
Funding
This work was supported by the Medical Scientific Research Foundation of Guangdong Province of China (A2022479 and A2024048), Guangdong Provincial Basic and Applied Basic Research Project (2023A1515011699), Science and Technology Major Project of Zhongshan City, Guangdong Province (2023B3016), the National Demonstration Pilot Project for the Inheritance and Development of Traditional Chinese Medicine-Huang Yongzhuo’s high level team.
Author information
Authors and Affiliations
Contributions
M.H. Z.Z. and C.Y. contributed to investigation, data curation, formal analysis. M.H., C.Y. and X.M.W. contributed to the original draft preparation. Z.Z., J.T. and G.R.L. contributed to conceptualization. X.J.H. and W.B.D. contributed to writing, review and editing. C.Y.Z. W.B.D. and D.W.G. contributed to conceptualization, funding acquisition and supervision. All authors have read and agreed to the published version of the manuscript. M.H. Z.Z. and C.Y. contributed equally.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
We confirmed that all animal experiments complied with the ARRIVE guidelines. The animal experiments were carried out in compliance with ethical standards and approved (approved protocol number: AEWC-2021024) by the Animal Ethics and Welfare Committee of Zhongshan Hospital of Traditional Chinese Medicine.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Hu, M., Zhang, Z., Yan, C. et al. Exploration of the antitumor effect and mechanism of gambogic acid on osteosarcoma through network pharmacology and experimental pharmacology study. Sci Rep 15, 21647 (2025). https://doi.org/10.1038/s41598-025-05308-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-05308-2
