
Abstract
The study investigated the protective effects of fisetin on chikungunya virus (CHIKV)-induced apoptosis hallmarks in Huh7 cells. Fisetin significantly reduced CHIKV RNA levels and viral infectivity, outperforming Z-VAD-FMK and cisplatin. At 30 µM, fisetin markedly decreased (by > 90%) the number of infectious viral particles at 24 and 48 h post-infection (hpi). Fisetin also hindered CHIKV-induced DNA fragmentation, with the lowest levels observed in CHIKV-infected cells treated with fisetin compared to other treatments. Immunoblot analysis revealed that fisetin inhibited caspase-mediated PARP cleavage and significantly reduced cleaved PARP levels, indicating decreased apoptosis. Additionally, fisetin diminished the expression of cleaved caspase-3 and HSP-27 proteins, while restoring HIF-1α protein levels, suggesting a protective role against CHIKV-induced apoptosis. The study highlights the potential of fisetin as an effective antiviral agent against CHIKV through the modulation of apoptosis and oxidative stress pathways. These findings underscore the therapeutic promise of fisetin for treating CHIKV-induced apoptosis and warrant further investigation to explore its clinical applications and optimize its use in antiviral therapy.
Similar content being viewed by others

Introduction
Chikungunya virus (CHIKV) is a mosquito-transmitted virus that has caused major outbreaks in numerous regions around the globe. It has been detected in more than 110 countries, spanning Asia, Africa, Europe, and the Americas1. Aedes mosquitoes, particularly Aedes aegypti and Aedes albopictus, serve as the primary vectors for CHIKV2. The virus typically induces fever, joint pain, rash, and other symptoms, which can persist for weeks or even months3. In severe cases, individuals may suffer from prolonged joint pain and other complications3. In November 2023, the FDA approved the first CHIKV vaccine, named Ixchiq, for individuals aged 18 and older who are at higher risk of exposure to the virus. The vaccine is administered as a single dose and contains a live, weakened form of the CHIKV4. There are still no specific antiviral drugs approved to treat chikungunya infection. Disease management mainly focuses on supportive care, including rest, hydration, and medications to alleviate pain and fever5.
CHIKV can cause severe joint arthritis through a mechanism involving programmed cell death, or apoptosis, by mainly targeting joint tissues, including synovial cells and fibroblasts6. The immune system reacts by activating immune cells like macrophages and T cells7. The virus induces apoptosis in infected cells via both intrinsic and extrinsic pathways, which involve caspases, the key players in executing cell death8. CHIKV-induced apoptotic cells release inflammatory mediators and debris, attracting more immune cells to the site of infection, thus creating a cycle of inflammation and tissue damage, contributing to arthritis symptoms9. Persistent inflammation and ongoing apoptosis can result in chronic joint pain and arthritis, even after the acute phase of the infection has passed. The continuous cell death and immune activity can lead to long-term joint tissue damage.
Although CHIKV is primarily associated with joint and musculoskeletal manifestations, accumulating clinical evidence indicates that hepatic involvement is also a frequent and significant feature of infection. Reports from Malaysia and other endemic regions have documented elevated hepatic transaminases and clinical hepatitis in CHIKV patients, suggesting hepatotropism and liver dysfunction as part of the systemic disease spectrum10,11. To investigate this hepatic pathology, we selected Huh7 cells, a widely used human hepatocyte-derived line that permits CHIKV replication and enables molecular interrogation of virus-induced apoptosis12. Prior in vitro studies have demonstrated oxidative stress, mitochondrial dysfunction, and caspase activation in CHIKV-infected hepatocytes, supporting their utility in modeling liver-specific responses13. While the relevance of other cell types—such as synoviocytes and macrophages—in chikungunya pathogenesis, we aim to delineate the hepatocellular mechanisms of CHIKV-induced apoptosis and evaluate the anti-apoptotic effects of fisetin in this context. Future studies incorporating synovial and immune cell models, along with in vivo systems, will be essential to generalize these findings across tissue compartments.
Other than that, histopathological analysis of human liver samples from CHIKV-infected patients revealed significant alterations, including cell swelling, microvesicular steatosis, and apoptosis, predominantly in the midzonal area14. In cynomolgus macaques, CHIKV establishes long-term infection in the liver and multiple tissues, with macrophages serving as the main cellular reservoirs15. The involvement of hepatocyte cell death in the disease process implicates the value and suitability of hepatocytes to be a model for studying CHIKV-induced apoptosis.
Fisetin, a flavonoid found in various fruits and vegetables, has shown promising potential as an antiviral agent against CHIKV. Fisetin has been demonstrated to inhibit CHIKV replication, reduce cytopathic effects, and decrease viral protein expression, thereby limiting the virus’s ability to multiply and damage host cells16,17. Fisetin modulates the excessive inflammation that often accompanies CHIKV infections, which can lead to severe joint pain and arthritis18. Fisetin also exhibits anti-apoptotic properties, preventing the over-activation of pathways that trigger cell death19,20,21. This can help protect joint tissues and reduce the severity of arthritis linked to CHIKV. Hence, in this study, we evaluated the protective capacity of fisetin against CHIKV-induced apoptosis.
Materials and methods
Cells and virus propagation
Human hepatocellular carcinoma (Huh7) cells [JCRB0403] were sourced from the Japanese Collection of Research Bioresources (JCRB) Cell Bank, part of the National Institutes of Biomedical Innovation, Health, and Nutrition (NIBIOHN, Osaka, Japan). The Huh7 cells were cultured in maintenance media consisting of RPMI-1640 medium (ATCC 30-2001) supplemented with 10% fetal bovine serum (FBS) (Gibco), 1x non-essential amino acids (NEAA) (Gibco), and 50 IU penicillin/streptomycin (Sigma-Aldrich). Cells were incubated at 37 °C with 5% CO2 and sub-cultured or cryopreserved upon reaching 80% confluency. The CHIKV used in this study was of the East/Central/South African genotype (accession number: MY/065/08/FN295485). Virus propagation was carried out in Huh7 cells using RPMI-1640 medium supplemented with 2% FBS, 1x NEAA, and 50 IU penicillin/streptomycin. Supernatants were harvested, titrated by plaque assay, and stored at − 80 °C for future use.
Flavonoid, inducer and inhibitor
Fisetin (Fig. 1) was obtained from INDOFINE Chemical Company and dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich) to create a 50 mM stock solution stored at − 20 °C for future use. The final concentration of DMSO used in this study is 0.06%, while the optimal concentration of DMSO for drug studies using hepatic cell lines is between 0.05% and 0.2%22. A similar range of DMSO was also used in the previous study17. For experiments, fisetin was diluted to a working concentration of 30 µM in serum-free RPMI-1640 medium and filtered using a syringe filter with a 0.2 μm pore size (Millipore).The fisetin concentration used in this study was selected based on prior dose-response studies demonstrating its antiviral efficacy in the post-infection stage and minimal cytotoxicity in Huh7 cells16,17. These earlier studies showed that fisetin inhibited CHIKV replication in a dose-dependent manner, with IC₅₀ values in the range of 20–35 µM and no significant cytotoxic effects observed up to 1.22mM. Z-VAD-FMK (InvivoGen), a pan-caspase inhibitor, and cisplatin (Tocris Bioscience), a potent pro-apoptotic agent, were prepared following the manufacturers’ instructions to a concentration of 20 µM and 130 µM, respectively. All concentrations of flavonoid, inducer and inhibitor used were non-toxic to the cells.

Chemical structure of fisetin, a flavonoid polyphenol, illustrated using ChemDraw. The molecular structure highlights key functional groups, including hydroxyl groups at positions 3, 7, 3’, and 4’ on the flavone backbone, which contribute to its antioxidant and bioactive properties.
Pre-treatment and post-treatment assay
The Huh7 cells monolayer in maintenance media was cultured in a 24-well plate (Corning). The experimental design included a range of control and treatment groups to allow meaningful comparisons: (i) untreated, uninfected cells (mock or negative control), (ii) uninfected cells treated with fisetin, Z-VAD-FMK, or cisplatin (to evaluate compound-specific effects), (iii) CHIKV-infected, untreated cells (positive control for infection), and (iv) CHIKV-infected cells treated with fisetin, Z-VAD-FMK, and cisplatin. The cells were incubated with Z-VAD-FMK (20 µM) and cisplatin (130 µM) for 1 h at 37 °C ± 5% CO2. Following the removal of the treatment, the cells were infected with CHIKV inoculum (MOI = 1), and the plate was then incubated at 37 °C ± 5% CO2 for 1 h. After 1 h, the inoculum was removed, and fisetin (30 µM) was added to the cells. The plate was then incubated for 24–48 h. The supernatants and cell lysates were collected to perform the downstream assays. This assay was performed in three biological replicates, while the downstream experiments were performed in three technical replicates using the samples collected from this assay to account for intra-assay variability, and the data were presented as mean ± standard deviation (SD) from these biological replicates.
Plaque-forming unit (PFU) assay
The supernatants from the pre-treatment and post-treatment assay were used to perform the plaque-forming unit (PFU) assay. A monolayer of Huh7 cells cultured in a 24-well plate was overlaid with diluted supernatants and incubated at 37 °C ± 5% CO2 for 1 h. Upon removal of the inoculum, the cells were overlaid with plaque media (2% FBS media with 0.8% high-viscosity carboxymethylcellulose sodium salt) and incubated for 48 h. After 48 h, the cells were washed with 1x PBS, fixed with 4% paraformaldehyde (PFA), and stained with 0.5% crystal violet.
Viral yield reduction assay
Viral RNA from the collected supernatants was extracted using QIAamp Viral RNA Mini Kit (Qiagen). The extracted viral RNA was amplified and quantified by targeting the E1 gene of CHIKV using the Step-OnePlus Real-Time PCR System (Applied Biosystems, Foster, CA, USA). This process utilized the SensiFAST SYBR Hi-ROX One-Step Kit (Bioline Meridian Bioscience, London, UK) according to the manufacturer’s instructions, with serial dilutions of known CHIKV concentrations as standards. The thermal cycling conditions were set at 45 °C for 10 min, 95 °C for 2 min, followed by 40 cycles of 95 °C for 5 s, 60 °C for 10 s, and 72 °C for 5 s. The amplified cDNA products were confirmed through melting curve analysis.
DNA fragmentation assay
The qualitative assessment of apoptosis was conducted by viewing the fragmentation of cellular DNA, which was visualized as oligonucleosomal-sized DNA fragments (DNA laddering). The DNA fragmentation assay involved extracting intracellular DNA following a previously established method. The cells were collected at 48 hpi and washed with PBS by centrifuging at 800xg for 10 min. The cells were then resuspended in Hanks’ Balanced Salt Solution (HBSS), mixed with 5 ml of ice-cold 70% ethanol, and stored at − 20 °C for at least 7 days. Following storage, the cells were centrifuged again at 800xg for 10 min, and the pellet was air-dried to remove any residual ethanol. The pellet was then resuspended and lysed in 50–100 µl of phosphate citrate buffer (prepared with 192 parts of 200 mM Na₂HPO₄ and 8 parts of 100 mM citric acid, pH 7.8). The cell lysate was incubated for 2 h at 37 °C, centrifuged at 3000 rpm for 10 min, and the supernatant was concentrated using a Speed-Vac concentrator. The concentrated material was resuspended in 10 µl of Tris-EDTA buffer, followed by the addition of 3 µl of 0.25% NP-40 and 5 µl of RNase A (1 mg/ml). This mixture was incubated at 37 °C for 90 min. Subsequently, 5 µl of proteinase K (1 mg/ml) was added, and the mixture was incubated overnight at 37 °C. Finally, 20–30 µl of Tris-EDTA buffer was added, the samples were vortexed, and incubated at 65 °C for 90 min to dissolve the DNA. The isolated DNA from both control and infected cells was then subjected to electrophoresis on a 1.5% agarose gel with a 1 kb DNA ladder as a standard.
Immunoblot assay
For Poly (ADP-ribose) polymerase (PARP) cleavage analysis, the cells were collected at 48 hpi, washed with PBS and lysed in PARP sample buffer containing 62 mM Tris-HCl (pH 6.8), 6 M urea, 10% glycerol, 2% SDS, 5% 2-mercaptoethanol, and bromophenol blue. To ensure efficient dissociation of PARP–DNA complexes, the samples were sonicated on ice for six 5-second cycles and then heated at 65 °C for 20 min. The proteins were then separated by SDS-PAGE, followed by PVDF membrane blotting and blocking. To optimize reagent usage and reduce costs, the PVDF membrane was carefully cut into appropriate sections before hybridization with antibodies. This approach ensured the efficient use of membranes, reagents, and antibodies without compromising experimental integrity. The blot was incubated overnight with an anti-PARP antibody (cleaved form; Merck), followed by staining with HRP-conjugated anti-mouse antibodies (Cell Signaling Technology). Clarity Western ECL substrate detection was performed according to the manufacturer’s instructions (Bio-Rad Laboratories), and the membranes were visualized using the Gel Doc XR + Molecular Imager (Bio-Rad Laboratories) with Image Lab version 3.0 software (https://www.bio-rad.com/en-my/product/image-lab-touch-software?ID=PJW3UUTU86LJ, Bio-Rad Laboratories). Further analysis of the images was conducted using the ImageJ processing software version 1.49 (https://imagej.net/ij/download.html, NIH). Protein loading was normalized using the consistent expression of GAPDH (Abcam).
Proteome profiler human apoptosis array kit
The collected cells were lysed and further processed following the manufacturer’s protocol for the detection of 35 human apoptosis-related proteins (Cat. No: ARY009, R&D Systems). Streptavidin-HRP and chemiluminescent detection reagents were applied, producing a signal at each capture spot that corresponded to the amount of protein bound. The arrays were visualized using the Gel Doc XR + Molecular Imager (Bio-Rad Laboratories) with Image Lab version 3.0 software (Bio-Rad Laboratories). Further analysis of the images was conducted using the ImageJ processing software version 1.49 (NIH).
Statistical analysis
Statistical analyses, including non-linear regression curves, one-way or two-way analysis of variance (ANOVA), and Dunnett’s post-test, were conducted using GraphPad Prism software version 9.0.0 (https://www.graphpad.com/updates/prism-900-release-notes, GraphPad Software). The data represented three independent experiments, with values expressed as the mean ± standard deviation (SD).
Results
Fisetin reduced CHIKV replication and infectivity
The impact of fisetin on CHIKV replication and infectivity was assessed using a viral yield reduction assay and a PFU assay. The viral yield reduction assay showed that fisetin treatment significantly reduced (p < 0.0001) CHIKV RNA levels at 24 (supernatant 95% CI: 570.4, 654.9; cell 95% CI: 15638, 161082) and 48 h (supernatant 95% CI: 30929, 37770; cell 95% CI: 100430, 106996) post-infection (hpi) compared to untreated controls (Fig. 2a and b and Supplementary Table S1). Notably, fisetin was more effective in lowering viral RNA levels than Z-VAD-FMK or cisplatin at both time points, suggesting that neither inhibiting nor inducing apoptosis before CHIKV infection could limit the replication or infectivity. The plaque-forming unit assay supported these findings, demonstrating that 30 µM fisetin markedly reduced the number of infectious viral particles produced at 24 and 48 hpi (Fig. 2c). In contrast, Z-VAD-FMK and cisplatin treatments showed high viral infectivity, indicating that fisetin’s antiviral activity was more pronounced.

Fisetin treatment significantly reduced CHIKV RNA levels in the supernatant (a) and intracellular (b), and the number of infectious viral particles at both time points (c), outperforming Z-VAD-FMK and cisplatin. The percentage reduction in viral RNA copy number calculation: The RNA copy number from the treated samples was subtracted from the untreated control (mock) RNA copy number. The resulting difference was then divided by the control RNA copy number to determine the relative decrease, and the value was multiplied by 100 to express the reduction as a percentage.
Fisetin hindered CHIKV-induced DNA fragmentation
DNA fragmentation is a cleavage pattern characteristic of apoptotic cells and serves as a reliable marker for identifying apoptosis. DNA fragmentation was analyzed at 48 hpi to assess the impact of different treatments on CHIKV-infected cells (Fig. 3). Intense DNA fragmentation, indicative of significant apoptosis, was observed in the CHIKV-infected group, the cisplatin-treated group, and the CHIKV-infected cisplatin-treated group. In contrast, CHIKV-infected cells treated with 20 µM Z-VAD-FMK showed reduced DNA fragmentation, suggesting partial inhibition of apoptosis. The most notable reduction in DNA fragmentation was seen in CHIKV-infected cells treated with 30 µM fisetin, which exhibited the lowest level of DNA fragmentation among all treatment groups. These findings indicate that while cisplatin promotes DNA fragmentation due to its pro-apoptotic nature, Z-VAD-FMK and fisetin effectively mitigate DNA damage, with fisetin providing the most substantial protective effect against CHIKV-induced apoptosis.

DNA fragmentation at 48 hpi showed significant apoptosis in CHIKV-infected and cisplatin-treated groups. Reduced DNA fragmentation was observed with Z-VAD-FMK, and the lowest levels were seen with fisetin treatment, indicating strong protection against CHIKV-induced apoptosis.
Fisetin hampered the later stage of apoptosis
Cleaved PARP serves as a common marker for identifying cells undergoing apoptosis. The levels of cleaved PARP were evaluated using immunoblot analysis to determine the extent of apoptosis in different treatment groups (Fig. 4and Supplementary Figure S1). A faint band representing cleaved PARP was observed in cells treated with cisplatin alone, indicating a moderate level of apoptosis. The CHIKV-infected cells treated with cisplatin displayed the most intense cleaved PARP band, signifying a high level of apoptotic activity. In uninfected cells treated with cisplatin, the band intensity for cleaved PARP was more intense compared to CHIKV-infected cells alone which was still visible, suggesting a partial apoptosis-inducing effect. Notably, no cleaved PARP bands were detected in CHIKV-infected cells treated with Z-VAD-FMK and fisetin, indicating a significant reduction in apoptosis and suggesting that fisetin effectively inhibits caspase-mediated PARP cleavage during CHIKV infection.

Cleaved PARP levels at 48 hpi, assessed via immunoblot, showed high apoptosis in CHIKV-infected cells. Z-VAD-FMK treatments reduced apoptosis, with fisetin showing the most significant reduction. Also shown in this figure is the relative quantification of the proteins analyzed using ImageJ. Cut PVDF membrane sections were used for antibody hybridization to optimize reagent consumption while maintaining experimental validity. The uncropped/original version can be visualized in the Supplementary Figure S1.
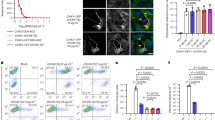
Fisetin diminished expression of caspase-3 and HSP-27 proteins
During CHIKV infection, cleaved caspase-3 and HSP-27 protein levels were elevated (p < 0.0001), while HIF-1α protein expression was decreased. These findings (Fig. 5and Supplementary Table S2) indicate that CHIKV infection induces apoptosis, as active caspase-3 degrades cellular proteins and causes morphological changes and DNA fragmentation characteristic of apoptosis. The increase in HSP-27 is likely due to its antioxidant role in mitigating cellular damage caused by the infection. In contrast, the reduction in HIF-1α is linked to excessive ROS-induced apoptosis. Treatment of CHIKV-infected cells with fisetin reversed the elevation (p < 0.0001) of cleaved caspase-3 (95% CI: 27690, 46629) and HSP-27 (95% CI: 49626, 56661) proteins while restoring HIF-1α protein levels. This suggests that fisetin protects cells from CHIKV-induced apoptosis.

During CHIKV infection, cleaved caspase-3 and HSP-27 protein levels were elevated, while HIF-1α protein expression was decreased. Fisetin treatment reversed these changes, indicating protection against CHIKV-induced apoptosis.
Discussions
Apoptosis is a defense mechanism that helps limit viral replication by eliminating infected cells. It plays a role in modulating the immune response. CHIKV infection can lead to mitochondrial dysfunction, endoplasmic reticulum stress, and activate the NF-κB pathway, triggering apoptosis23. By understanding the interaction between CHIKV and apoptotic pathways, researchers can develop strategies to enhance the immune response against the virus. Identifying key molecules involved in the apoptotic pathways can lead to development of new antiviral drugs. Flavonoids, with their ability to modulate apoptosis, can be explored as potential therapeutic agents against CHIKV. Fisetin exerts its anti-apoptotic effects through multiple molecular pathways, including the PI3K/Akt/mTOR pathway, which is known to regulate cell growth and survival19. Fisetin helps reduce oxidative stress, which can trigger apoptosis, by scavenging free radicals and enhancing antioxidant defenses24. Fisetin also has anti-inflammatory properties, which can indirectly contribute to its anti-apoptotic effects by reducing inflammation-induced cell death. Fisetin has shown potential in modulating antiviral responses, including those against CHIKV16,17. Leveraging the anti-CHIKV and anti-apoptotic activities of fisetin to develop a therapeutic antiviral treatment for CHIKV-induced apoptosis involves understanding and targeting specific molecular pathways. The findings from our viral yield reduction and PFU assays demonstrate that fisetin exerts a significant inhibitory effect on CHIKV replication and infectivity in Huh7 cells. Specifically, fisetin treatment resulted in a marked reduction in CHIKV RNA levels at both 24 and 48 hpi, with reductions reaching statistical significance (p < 0.0001). The viral yield assay indicated that fisetin decreased the intracellular and extracellular viral load more effectively than both Z-VAD-FMK and cisplatin, suggesting a robust antiviral effect that is not solely attributable to modulation of apoptosis pathways. These observations were corroborated by the PFU assay, which showed a substantial decline in the number of infectious viral particles upon fisetin treatment, highlighting its capacity to impair viral assembly or release. Interestingly, treatments with Z-VAD-FMK and cisplatin, although capable of affecting apoptosis, did not significantly reduce viral RNA or infectivity, indicating that the antiviral activity of fisetin is likely mediated through mechanisms beyond apoptosis inhibition.This is consistent with other studies that have explored the effects of these compounds on viral infections. For example, research on Z-VAD-FMK has shown that while it can inhibit apoptosis, it may not significantly impact viral replication25,26,27. Similarly, cisplatin, primarily known for its anti-cancer properties, has limited antiviral activity and can induce apoptosis in infected cells28. The findings of this study are consistent with previous research that highlights fisetin’s anti-apoptotic properties. For instance, a study by Adhami et al. 2012 demonstrated that fisetin inhibits apoptosis in cancer cells by modulating the PI3K/Akt/mTOR pathway29. This aligns with the current study’s observation that fisetin effectively reduces DNA fragmentation, a hallmark of apoptosis, in CHIKV-infected cells. The analysis of DNA fragmentation at 48 h post-infection (hpi) provides important insights into the apoptotic response of CHIKV-infected cells and the protective effects of different treatments. As expected, intense DNA fragmentation was observed in the CHIKV-infected group, reflecting the virus’s capacity to induce apoptosis. Treatment with cisplatin, a known pro-apoptotic agent, further enhanced DNA fragmentation, both in uninfected and infected cells, confirming its role in promoting apoptotic pathways. Conversely, treatment with 20 µM Z-VAD-FMK—the pan-caspase inhibitor—resulted in a notable reduction in DNA fragmentation in CHIKV-infected cells, indicating partial inhibition of apoptosis through caspase blockade. Most strikingly, cells treated with 30 µM fisetin exhibited the lowest levels of DNA fragmentation among all groups, demonstrating its potent anti-apoptotic effect. These findings suggest that fisetin effectively mitigates CHIKV-induced DNA damage and apoptosis, possibly through mechanisms involving its antioxidant and cell survival pathways. The differential effects of these treatments underscore fisetin’s potential as a cytoprotective agent capable of counteracting virus-induced apoptotic damage, which could have implications for preserving cellular integrity during viral infections. Previous studies, such as the one by Yang et al. 2006, have shown that cisplatin induces apoptosis through DNA damage and mitochondrial dysfunction30. The current study’s findings that cisplatin promotes DNA fragmentation are in line with its well-documented pro-apoptotic nature. The immunoblot analysis of cleaved PARP provides strong evidence for the modulation of apoptosis under different treatment conditions in CHIKV-infected cells. Notably, CHIKV infection induced a moderate level of apoptosis, as reflected by the cleaved PARP band, consistent with previous reports of viral-induced apoptotic activation. Treatment with cisplatin alone led to a higher increase in cleaved PARP, confirming its pro-apoptotic effects. When CHIKV-infected cells were treated with cisplatin, the elevation in cleaved PARP band intensity suggests that cisplatin partially influences apoptotic pathways in this context. The further attenuation of cleaved PARP bands in CHIKV-infected cells treated with Z-VAD-FMK, a pan-caspase inhibitor, indicates that caspase activity plays a significant role in virus-induced apoptosis, and its inhibition can decrease PARP cleavage. Most importantly, treatment with fisetin resulted in the absence of detectable cleaved PARP, demonstrating its potent capacity to suppress caspase-mediated apoptosis during CHIKV infection. These findings support the hypothesis that fisetin exerts a strong anti-apoptotic effect, likely through inhibition of caspase activation. The observed alterations in protein expression during CHIKV infection underscore the activation of apoptosis and cellular stress responses. Elevated levels of cleaved caspase-3 confirm that CHIKV induces apoptosis via caspase activation, leading to the degradation of critical cellular proteins and morphological features typical of apoptotic cells. The concurrent increase in HSP-27 likely reflects its role as a cellular antioxidant, being upregulated in response to oxidative stress to protect cells from damage caused by viral infection. Conversely, the decrease in HIF-1α expression aligns with the model where excess reactive oxygen species (ROS) generated during infection promote apoptosis, as HIF-1α is known to be inhibited under oxidative stress conditions. Importantly, treatment with fisetin effectively reversed these molecular changes: it significantly reduced levels of cleaved caspase-3 and HSP-27 and restored HIF-1α expression, indicating that fisetin can mitigate apoptosis and oxidative stress induced by CHIKV. The increase in HSP-27 protein levels during CHIKV infection supports the role of HSP-27 as an antioxidant that mitigates cellular damage31. The reduction in HIF-1α protein expression due to excessive ROS-induced apoptosis is also consistent with the findings of other studies that have shown the involvement of oxidative stress pathways in CHIKV-induced apoptosis32. Fisetin treatment reduces the elevated levels of cleaved caspase-3 and HSP-27 proteins while restoring HIF-1α protein levels, suggesting fisetin has a protective role against apoptosis induced by CHIKV. This aligns with the general understanding that antioxidants and anti-apoptotic agents can mitigate the cellular damage caused by viral infections. To the best of our knowledge, this is the first study to demonstrate that fisetin not only reduces CHIKV replication but also modulates virus-induced apoptotic hallmarks in hepatocellular carcinoma cells, highlighting its dual antiviral and cytoprotective roles. While fisetin demonstrates promising antiviral and cytoprotective effects in vitro, its clinical application faces significant challenges due to its poor bioavailability and rapid metabolism. Fisetin has low water solubility and is rapidly conjugated into glucuronides and sulfates in vivo, which substantially limits its systemic exposure following oral administration33,34. Pharmacokinetic studies have shown that fisetin has a short plasma half-life and low oral bioavailability, often necessitating high doses to achieve therapeutic plasma concentrations (33). These limitations could impact the translational potential of fisetin for antiviral therapy against CHIKV. Various strategies are being explored to overcome these barriers, including the development of fisetin-loaded nanoparticles, liposomes, and other novel drug delivery systems aimed at enhancing its stability, solubility, and bioavailability35,36. Although the present study demonstrates the efficacy of fisetin in vitro, future research should incorporate formulation approaches or chemical modifications to improve its pharmacokinetic profile to ensure clinical relevance.
Conclusion
This study demonstrates that fisetin significantly attenuates CHIKV-induced apoptosis in Huh7 cells by reducing DNA fragmentation, lowering cleaved PARP and caspase-3 expression, and restoring HIF-1α levels. These results underscore fisetin’s cytoprotective and antiviral potential in vitro. While promising, the findings are limited by the use of a hepatocellular carcinoma cell line, which may not fully represent CHIKV pathogenesis in vivo—particularly in joint and musculoskeletal tissues. To validate these observations, in vivo studies using relevant animal models are warranted. Moreover, incorporating targeted pathway inhibition or siRNA-based gene silencing approaches in future investigations could clarify the molecular mechanisms underlying fisetin’s modulation of CHIKV-induced apoptosis. Exploration of advanced delivery platforms, such as fisetin-loaded nanoparticles, may enhance its bioavailability and tissue targeting, potentially improving therapeutic outcomes. Defining its pharmacokinetics, optimal dosing strategies, and evaluating synergistic effects with existing antiviral or anti-inflammatory agents will be critical steps toward clinical translation. Finally, assessing fisetin’s efficacy against other medically relevant arboviruses with shared apoptotic features may further expand its potential in antiviral therapeutics.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Cai, L. et al. The research progress of Chikungunya fever. Front. Public. Health. 10, 1095549. https://doi.org/10.3389/fpubh.2022.1095549 (2023).
Higgs, S. & Vanlandingham, D. Chikungunya virus and its mosquito vectors. Vector Borne Zoonotic Dis. 15, 231–240. https://doi.org/10.1089/vbz.2014.1745 (2015).
Arroyo-Ávila, M. & Vilá, L. M. Rheumatic manifestations in patients with Chikungunya infection. P R Health Sci. J. 34, 71–77 (2015).
Ng, L. F. & Rénia, L. Live-attenuated Chikungunya virus vaccine. Cell 187, 813–813. https://doi.org/10.1016/j.cell.2024.01.033 (2024).
Soto-Garita, C., Carrera, J. P., López-Vergès, S. & Corrales-Aguilar, E. Advances in clinical diagnosis and management of Chikungunya virus infection. Curr. Treat. Opt. Infect. Dis. 10, 397–409. https://doi.org/10.1007/s40506-018-0172-x (2018).
Krejbich-Trotot, P. et al. Chikungunya virus mobilizes the apoptotic machinery to invade host cell defenses. FASEB J. 25, 314–325. https://doi.org/10.1096/fj.10-164178 (2011).
Lum, F. M. et al. Crosstalk between CD64 + MHCII + macrophages and CD4 + T cells drives joint pathology during Chikungunya. EMBO Mol. Med. 16, 641–663. https://doi.org/10.1038/s44321-024-00028-y (2024).
Nayak, T. K. et al. Regulation of viral replication, apoptosis and pro-inflammatory responses by 17-AAG during Chikungunya virus infection in macrophages. Viruses 9, 3. https://doi.org/10.3390/v9010003 (2017).
Amdekar, S., Parashar, D. & Alagarasu, K. Chikungunya virus-induced arthritis: role of host and viral factors in the pathogenesis. Viral Immunol. 30, 691–702. https://doi.org/10.1089/vim.2017.0052 (2017).
Sapuan, S. et al. Clinical manifestations of Chikungunya virus infection in malaysia: a retrospective cohort study. BMC Infect. Dis. 20, 693. https://doi.org/10.1186/s12879-020-05232-1 (2020).
Yactayo, S., Staples, J. E., Millot, V., Cibrelus, L. & Ramon-Pardo, P. Epidemiology of Chikungunya in the Americas. Bull. World Health Organ. 94, 441–442. https://doi.org/10.2471/BLT.16.175802 (2016).
Hapuarachchi, H. C. et al. Laboratory-confirmed dengue and Chikungunya co-infection: a case report and literature review. PLoS Negl. Trop. Dis. 13, e0007212. https://doi.org/10.1371/journal.pntd.0007212 (2019).
Lim, Y. S., Ng, C. T., Ng, K. P., Kamarulzaman, A. & Chan, Y. F. Chikungunya virus infection modulates apoptotic signaling pathways in human liver cells. Sci. Rep. 10, 11317. https://doi.org/10.1038/s41598-020-67922-3 (2020).
Ribeiro, Y. P. et al. Comparative analysis of human hepatic lesions in dengue, yellow fever, and chikungunya: revisiting histopathological changes in the light of modern knowledge of cell pathology. Pathogens 12, 680. https://doi.org/10.3390/pathogens12050680 (2023).
Labadie, K. et al. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J. Clin. Invest. 120, 894–906 (2010).
Lani, R. et al. Antiviral activity of selected flavonoids against Chikungunya virus. Antiviral Res. 133, 50–61. https://doi.org/10.1016/j.antiviral.2016.07.009 (2016).
Lani, R. et al. Fisetin modulates toll-like receptor-mediated innate antiviral response in Chikungunya virus-infected hepatocellular carcinoma Huh7 cells. Immuno 2, 703–719. https://doi.org/10.2139/ssrn.4166520 (2022).
Caglioti, C. et al. Chikungunya virus infection: an overview. New. Microbiol. 36, 211–227 (2013).
Zhou, C. et al. Biological effects and mechanisms of Fisetin in cancer: a promising anti-cancer agent. Eur. J. Med. Res. 28, 297. https://doi.org/10.1186/s40001-023-01271-8 (2023).
Sundarraj, K., Raghunath, A. & Perumal, E. A review on the chemotherapeutic potential of fisetin: in vitro evidences. Biomed. Pharmacother. 97, 928–940. https://doi.org/10.1016/j.biopha.2017.10.164 (2018).
Sung, B., Pandey, M. K. & Aggarwal, B. B. Fisetin, an inhibitor of cyclin-dependent kinase 6, down-regulates nuclear factor-κB-regulated cell proliferation, antiapoptotic and metastatic gene products through the suppression of TAK-1 and receptor-interacting protein-regulated IκBα kinase activation. Mol. Pharmacol. 71, 1703–1714. https://doi.org/10.1124/mol.107.034512 (2007).
Che Omar, R. N., Ismail, M. I., Noorden, M. S. A., Yusoff, R. M. & Hatta, F. H. Determining the population doubling time of HepG2 and Huh-7 cells and the toxic effect of dimethyl sulfoxide (DMSO). ASM Sci. J. 17, 1–11. https://doi.org/10.32802/asmscj.2022.1238 (2022).
Abraham, R. et al. Induction of cytopathogenicity in human glioblastoma cells by Chikungunya virus. PLoS One. 8, e75854. https://doi.org/10.1371/journal.pone.0075854 (2013).
Park, C. et al. Fisetin attenuated oxidative stress-induced cellular damage in ARPE-19 human retinal pigment epithelial cells through Nrf2-mediated activation of Heme oxygenase-1. Front. Pharmacol. 13, 927898. https://doi.org/10.3389/fphar.2022.927898 (2022).
Kim, M. S., Lee, J. A. & Kim, K. H. Effects of a broad-spectrum caspase inhibitor, Z-VAD (OMe)-FMK, on viral hemorrhagic septicemia virus (VHSV) infection-mediated apoptosis and viral replication. Fish. Shellfish Immunol. 51, 41–45 (2016).
van den Berg, E. et al. The caspase inhibitor zVAD increases lung inflammation in Pneumovirus infection in mice. Physiol. Rep. 3, e12332. https://doi.org/10.14814/phy2.12332 (2015).
Nava, V. E. et al. Sindbis virus induces apoptosis through a caspase-dependent, CrmA-sensitive pathway. J. Virol. 72, 452–459 (1998).
Wang, Y. F. et al. Cisplatin disrupts the latency of human herpesvirus 8 and induces apoptosis in primary effusion lymphoma cells. Cancer Investig. 30, 268–274 (2012).
Adhami, V. M. et al. Dietary flavonoid fisetin: a novel dual inhibitor of PI3K/Akt and mTOR for prostate cancer management. Biochem. Pharmacol. 84, 1277–1281 (2012).
Yang, Z. et al. Cisplatin preferentially binds mitochondrial DNA and voltage-dependent anion channel protein in the mitochondrial membrane of head and neck squamous cell carcinoma: possible role in apoptosis. Clin. Cancer Res. 12, 5817–5825 (2006).
Vidyasagar, A., Wilson, N. A. & Djamali, A. Heat shock protein 27 (HSP27): biomarker of disease and therapeutic target. Fibrogenesis Tissue Repair. 5, 1–7 (2012).
Dhanwani, R. et al. Characterization of Chikungunya virus infection in human neuroblastoma SH-SY5Y cells: role of apoptosis in neuronal cell death. Virus Res. 163, 563–572 (2012).
Sundarraj, K., Raghunath, A. & Perumal, E. A review on the chemotherapeutic potential of fisetin: in vitro evidences. Biomed. Pharmacother. 97, 928–940 (2018).
Touil, Y. S. et al. Fisetin disposition and metabolism in mice: identification of Geraldol as an active metabolite. Biochem. Pharmacol. 82, 1731–1739 (2011).
Szymczak, J. & Cielecka-Piontek, J. Fisetin—In search of better bioavailability—From macro to nano modifications: A review. Int. J. Mol. Sci. 24, 14158 (2023).
Khan, N., Syed, D. N., Ahmad, N. & Mukhtar, H. Fisetin: a dietary antioxidant for health promotion. Antioxid. Redox. Signal. 19, 151–162 (2013).
Acknowledgements
We would like to acknowledge the staff of Tropical Infectious Diseases Research and Education Centre (TIDREC) and the Department of Medical Microbiology, Faculty of Medicine, Universiti Malaya, for their unwavering support throughout this study.
Funding
This work was funded in part by grants from the Ministry of Higher Education, Malaysia, for niche area research under the Higher Institution Centre of Excellence (HICoE) program (MO002-2019 and TIDREC-2023) and the Ministry of Higher Education under Dana Langganan SUKUK Pakej Rangsangan Ekonomi Prihatin Rakyat (SUKUK PRIHATIN)-Fasa 2 (MO002-2021).We would like to acknowledge the staff of Tropical Infectious Diseases Research and Education Centre (TIDREC) and the Department of Medical Microbiology, Faculty of Medicine, Universiti Malaya, for their unwavering support throughout this study. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
Author information
Authors and Affiliations
Contributions
R.L. designed and conducted the experiments, analyzed the data, and wrote the main manuscript text. P.H. contributed to the experimental design, provided technical support, and assisted with data analysis. S.A.B. supervised the project, provided critical feedback, and reviewed the manuscript. All authors reviewed and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lani, R., Hassandarvish, P. & AbuBakar, S. Fisetin limits Chikungunya virus-induced apoptosis hallmarks in hepatocellular carcinoma cells. Sci Rep 15, 25140 (2025). https://doi.org/10.1038/s41598-025-09213-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-09213-6
