
Abstract
Shiga toxin-producing Escherichia coli (STEC) is a significant public health concern, with plasmids playing a key role in its pathogenicity. This study investigates the relationship between plasmid types, virulence genes, and host specificity in 96 STEC strains isolated from cattle, sheep, goats, and pigeons in Iran. We examined the distribution of plasmid-encoded virulence genes, including astA, cma, cba, rhsA1, rhsA2, rhsC, stcE, katP, toxB, espP, subAB, ihaG, LAA iha, saa, etpD, cif, and epeA, along with ehxA subtypes and plasmid replicon types using PCR and RFLP analysis. The most frequently detected virulence factors, rhsC and ihaG, were associated with ehxA subtype A and IncFIB/IncK/B plasmids. Pigeon-derived isolates had a unique virulence profile, including cif but lacking ehxA, suggesting distinct pathogenic mechanisms. stcE and etpD were exclusive to positive controls O157 isolates, linked to ehxA subtype B and IncFIB plasmids. The predominance of ehxA subtype A in ruminants highlights potential host-specific associations. The widespread presence of IncFIB plasmids suggests their broad role in STEC pathogenicity. These findings highlight the critical role of plasmid-encoded virulence factors in STEC pathogenesis and underscore the need for targeted surveillance and control strategies tailored to specific host populations.
Similar content being viewed by others
Introduction
The primary microorganisms responsible for food contamination are members of the Enterobacterales, with Escherichia coli being a significant species1. This bacterium is one of the most thoroughly researched and was among the first to have its genome sequenced2. It includes six intestinal pathotypes: enteropathogenic E. coli (EPEC), enterotoxigenic E. coli (ETEC), enteroaggregative E. coli (EAEC), enteroinvasive E. coli (EIEC), diffusely adherent E. coli (DAEC), and Shiga toxin-producing E. coli (STEC)1. These pathotypes are often implicated in foodborne outbreaks, with STEC being particularly critical for public health due to its potential to cause diarrhea, bloody diarrhea (hemorrhagic colitis), hemolytic uremic syndrome (HUS), and kidney damage3,4.
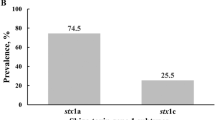
STEC have been found in several animal species, which act as natural reservoirs for these bacteria1. The primary reservoirs of STEC are cattle, sheep, and goats, which can harbor the bacteria in their gastrointestinal tracts without showing clinical signs1,5. Additionally, other animals like pigeons have also been implicated as potential carriers of STEC6,7. STEC’s disease-causing ability is linked to the production of Shiga-like toxins (Stx), which come in two main types, Stx1 and Stx2, encoded by the stx1 and stx2 genes8. stx1 has four variants: stx1a, stx1c, stx1d, and stx1e; stx2 is more diverse, with 15 variants: stx2a, stx2b, stx2c, stx2d, stx2e, stx2f, stx2g, stx2h, stx2i, stx2j, stx2k, stx2l, stx2m, stx2n, and stx2o9. Research has shown a link between the stx subtype and disease severity, with strains producing stx2, especially the stx2a subtype, being more associated with severe disease and HUS10. This knowledge has informed differential case management and public health strategies based on stx profile-derived STEC pathotype8,11. Additionally, the eae gene, located on the locus of enterocyte effacement (LEE), encodes intimin, an adhesin crucial for gut colonization. Furthermore, many disease-causing STEC strains, such as those belonging to O157 and serogroups O26, O103, O111, and O145, contain mobile genetic elements like plasmids, transposons, phages, and pathogenicity islands. These elements also play a significant role in the severity of the disease12.
STEC plasmids contain various genes involved in cell adhesion, proteases, toxins, and other potential virulence factors13,14,15. These combinations of virulence factors can increase the risk of severe symptoms like HUS16. Large plasmids encoding enterohemolysin (ehxA) are found in most STEC isolates, including E. coli O157 and non-O157 strains such as O26, O103, O113, and O145, all commonly associated with diarrheal disease and HUS17. The ehxA gene, part of the hemolysin operon on these plasmids, has six subtypes (A to F) and is used as a diagnostic marker for highly pathogenic STEC like the O157 serogroup16,18. Plasmid replicon typing is another method for classifying the plasmids based on the variations in replication loci14. To date, 17 different plasmid replicon types have been identified in E. coli, with the IncF family being among the most frequent in STEC strains that cause severe symptoms19.
Plasmids play crucial roles in the pathogenesis of STEC; often carrying key virulence genes such as ehxA, espP, katP, and etpD that contribute to the severity of human infections20,21. Despite increasing knowledge of plasmid-encoded factors, significant gaps remain in our understanding of the diversity of these plasmids, their evolutionary trajectories, and their functional associations with host specificity and pathogenic potential15,22,23. Notably, it remains unclear whether specific ehxA subtypes or replicon types are consistently associated with certain plasmid-borne virulence genes, or whether particular plasmids exhibit host-specific properties linked to animal reservoirs. Previous studies have characterized plasmids in select STEC lineages, but a comprehensive understanding of plasmid diversity across different hosts is still lacking22. Addressing these gaps is critical, as improved knowledge of the interplay between plasmid content, host origin, and virulence could enhance surveillance, support risk assessment, and inform targeted public health interventions24. In this study, we conducted a comprehensive analysis of STEC strains from various animal hosts to explore these crucial connections, aiming to advance our understanding of STEC pathogenesis and ultimately reduce the burden of STEC-related diseases.
Results
STEC isolates
Among the 96 STEC isolates analyzed in this study—including 50 previously collected and 46 newly obtained strains—the sources included cattle (35 isolates), sheep (34), pigeons (18), and goats (9), reflecting a diverse range of animal hosts. Regarding the genes stx1, stx2, eae, and ehxA, we found the following: stx1 was present in 29/96 (30.2%, p < 0.05) isolates, stx2 in 42/96 (43.7%, p < 0.05), both stx1 and stx2 in 25/96 (26.0%), eae in 30/96 (31.2%), and ehxA in 56/96 (58.3%). stx2 subtyping revealed that, among the 42 stx2-positive isolates, 18/42 (42.8%, p < 0.05) had the stx2f subtype only, 13/42 (30.9%) had the stx2a + stx2c + stx2d profile, 5/42 (11.9%) had stx2a + stx2c, 1/42 (2.3%) had stx2c + stx2d, and 5/42 (11.9%) did not have detectable stx2 subtypes. The strains were classified into the serogroups O5 (n = 13), O26 (n = 6), O80 (n = 2), O91 (n = 3), O103 (n = 12), O111 (n = 3), O113 (n = 13), and O128 (n = 9). For 35 isolates, the O-groups were not determined (Table S1 in supplementary materials).
Virulence genes
The analysis revealed that the most frequently identified virulence genes were rhsC (88.5%, 85/96, p < 0.05), IhaG (67.7%, 65/96, p < 0.05), and subAB (48.9%, 47/96). These were followed by cba (47.9%, 46/96, p < 0.05), LAA iha (42.7%, 41/96), cma (37.5%, 36/96), cif (30.2%, 29/96, p < 0.05), astA (29.1%, 28/96, p < 0.05), espP (23.9%, 23/96), saa (22.9%, 22/96), rhsA1 (12.5%, 12/96), epeA (11.4%, 11/96), katP (9.3%, 9/96), rhsA2 (6.2%, 6/96), and toxB (4.1%, 4/96) (Fig. 1). The genes stcE and etpD were not detected in any of the isolates, except for the positive control (O157:H7 strain Sakai), which contained both genes.

Distributions of the plasmid-encoded virulence genes, ehxA subtypes, and plasmid replicon types in different animal hosts (A, B, C) and stx2-positive isolates (D, E, F), respectively. The most prevalent virulence genes detected were IhaG in cattle (91.4%, 32/35), rhsC in sheep/goats (97.6%, 42/43), and cif in pigeons (100%, 18/18) isolates (A). Subtype A of the ehxA gene was the most common ehxA subtype in cattle (37.1%, 13/35) and sheep/goats (79%, 34/43), while pigeons tested negative for the ehxA gene (B). The highest rates of replicon types were found to be IncFIB in cattle (40%, 14/35), sheep/goats (74.4%, 32/43), and pigeons (88.8%, 16/18) isolates (C). Among the 42 stx2-positive isolates, rhsC (80.9%, 34/42) virulence gene (D), subtype A (28.5%, 12/42) of ehxA gene (E), and FIB (59.5%, 25/42) replicon type (F), were detected as the highest rate.
EhxA subtypes
By ehxA subtyping, of 56 ehxA-positive isolates, we identified the subtypes A (83.9%, 47/56, p < 0.05), C (12.5%, 7/56), and D (3.5%, 2/56) in the studied isolates (Fig. 2). ehxA subtype B was only detected in the positive control (E. coli O157:H7 295 EC-TMU). Subtypes E and F were not observed in any of the isolates. Regarding the association between ehxA subtypes and virulence genes, we found that subtype A was linked to three different patterns: rhsC, subAB, IhaG, LAA iha; rhsC, espP, subAB, IhaG, saa, epeA; and cma, cba, rhsC, subAB, IhaG, LAA iha. Additionally, subtype C was associated with the pattern cba, rhsA1, rhsC, katP, toxB, espP, IhaG, cif (Table 1).

Dissemination of ehxA subtypes, replicon types, and their associations in the 96 STEC isolates in Iran. A total of 56 ehxA-positive isolates were identified, with the majority (83.9%) classified as subtype A, followed by subtypes C (12.5%), and D (3.5%) (A). Among the 76 STEC isolates containing plasmids, the most common replicon type was FIB (82.8%), with additional types including IncK/B (27.6%), IncB/O (14.4%), IncP (10.5%), and others (B). The heat map illustrates the co-occurrence of ehxA subtypes and replicon types in these isolates (C).
Replicon types
Plasmid Inc typing revealed that, among the 76 STEC isolates harboring plasmids, the most common replicon type was FIB (82.8%, 63/76, p < 0.05), followed by IncK/B (27.6%, 21/76, p < 0.05), IncB/O (14.4%, 11/76), IncP (10.5%, 8/76), IncY (7.8%, 6/76), IncFIA (3.9%, 3/76), IncI1 (3.9%, 3/76), and IncL/M (1.3%, 1/76) (Fig. 2). Notably, some plasmids contained more than one replicon type. The virulence gene patterns associated with these replicon types were as follows: astA, cma, cba, rhsC, cif were most commonly detected in IncFIB plasmids; rhsC, subAB, IhaG, LAA iha were associated with IncK/B and IncFIB; rhsC, espP, subAB, IhaG, saa, epeA were linked to IncK/B; and cba, rhsA1, rhsC, katP, toxB, espP, IhaG, cif were found in IncFIB and IncI1 plasmids (Table 1; Fig. 3).

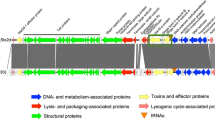
Features of the plasmids based on the most detected profiles in STEC isolates from cattle, sheep/goats (A), and pigeons (B).
Discussion
The highly variable nature of STEC plasmids prompted an examination of the correlations between their ehxA subtypes and replicon types with the virulence genes they carry. These plasmids harbor genes for adhesion, toxins, and other virulence factors, significantly influence the severity of illnesses such as HUS25. This investigation aims to elucidate the intricate relationship between specific plasmid types and virulence factors in STEC strains, with a particular focus on whether certain ehxA subtypes or plasmid replicon types are associated with the plasmid-encoded virulence genes, as revealed through a comprehensive analysis of STEC strains from various animal hosts.
The analysis revealed that the rhsC (88.5%, p < 0.05) and IhaG (67.7%, p < 0.05) genes were the most frequently identified virulence genes among the isolates. The rhsC gene is associated with the type VI secretion system (T6SS) in pathogenic E. coli, including STEC, which mediates bacterial competition and enhances pathogenicity by aiding intracellular survival against human macrophages26,27,28. Meanwhile, the IhaG gene encodes an adhesin crucial for STEC adherence to epithelial cells, facilitating toxin delivery, evasion of peristaltic clearance, and access to host nutrients29,30,31. These two genes were prevalent among the highly pathogenic stx2-positive isolates (p < 0.05), predominantly linked to plasmids containing ehxA subtype A (p < 0.05) and IncFIB/IncK/B replicons (p < 0.05). Notably, some alleles of the ihaG gene may also be encoded on the chromosome, indicating that its presence is not exclusively plasmid-mediated16. This finding underscores the significant role of plasmids harboring the rhsC gene and, in particular, those encoding the IhaG gene in the pathogenesis of STEC strains and emphasizes the necessity for specific approaches to reduce the dangers posed by these highly pathogenic types.
A notable presence of the cif virulence gene was found in pigeon-derived STEC isolates, with all such isolates testing positive for cif (p < 0.05). cif, a member of the cysteine protease superfamily, is known to inhibit cell-cycle progression in eukaryotic hosts, suggesting it provides specific advantages in the STEC pathogenic process32,33,34,35. This observation is consistent with findings by van Hoek et al., who also reported cif in all pigeon-derived STEC isolates, indicating a strong correlation between cif and the pigeon host36. Additionally, plasmids in these pigeon-STEC isolates were of the IncF family, specifically the IncFIB subtype, and notably, these plasmids lacked the ehxA gene, making ehxA subtyping unfeasible. This points to a distinctive plasmid profile in pigeon-derived STEC, emphasizing unique pathogenic mechanisms that may be involved in these isolates.
The identification of the virulence genes stcE and etpD was exclusive to positive controls O157 isolates. These genes, encoded on the plasmid O157 (pO157), play a crucial role in STEC pathogenesis. Specifically, O157 utilizes the etp type II secretion system to secrete the metalloprotease stcE, which cleaves the serpin C1 esterase inhibitor during human infection13,37. Consistent with previous studies, stcE and etpD were found only in O157 STEC isolates, highlighting their specific association with this serogroup38,39. Additionally, these genes were carried on STEC plasmids with ehxA subtype B and IncFIB replicon type, emphasizing the distinct plasmid profile of O157 strains and aligning with these reports by Lorenz et al., which identified ehxA subtype B specifically in O157 STEC strains18,40reinforcing the unique genetic and pathogenic characteristics of these isolates.
The prevalence of ehxA subtype A was significantly higher in STEC isolates from sheep (p < 0.05) and goats (p < 0.05) compared to other animal hosts. Due to limited data on this specific association, further research is strongly recommended to better understand the relationship between ehxA subtypes and different animal hosts. In terms of plasmid characteristics, the IncFIB replicon type was the most frequently detected across all studied animal hosts (p < 0.05), including cattle, sheep, goats, and pigeons. This high distribution of IncFIB plasmids in STEC strains aligns with previous studies highlighting their prevalence not only in STEC but also in other E. coli pathotypes, such as avian pathogenic E. coli (APEC), fecal E. coli (AFEC), and uropathogenic E. coli (UPEC)19,41,42. These findings underscore the widespread occurrence of IncFIB plasmids in E. coli strains and their potential role in the pathogenicity of various E. coli pathotypes, including STEC.
The most common plasmid-encoded virulence gene profile in pigeon isolates included the IncFIB (p < 0.05) and Frep (p < 0.05) replicons, along with the astA, cma, cba, rhsC, and cif (p < 0.05) virulence genes, but lacked the ehxA hemolysin gene. Conversely, the IncFIB/IncK/B replicons were predominantly found in sheep (p < 0.05), goat (p < 0.05), and cattle (p < 0.05) hosts, along with ehxA subtype A (p < 0.05) and various other virulence gene profiles. Interestingly, despite differing ecological niches, the plasmid-encoded virulence gene profiles in sheep, goat, and cattle hosts were more similar to each other than to those found in pigeons. Although more research is needed to confirm this, it points to a complex and surprising plasmid landscape. This highlights the importance of further studies to better understand plasmid diversity and its role in STEC pathogenicity across various animals. The similarity in plasmid profiles among sheep, goats, and cattle, compared to pigeons, suggests that these animals can be grouped by their plasmid-related traits, even though the plasmids themselves vary.
STEC carry large plasmids of various sizes, often 75–100 kb. These plasmids are highly heterogeneous, harboring different collections of genes43. Our study underscores the critical role of plasmid-encoded virulence factors in the pathogenicity of STEC strains across various animal hosts. We identified a significant prevalence of the rhsC and IhaG genes in highly pathogenic stx2 positive isolates, predominantly associated with plasmids containing the ehxA subtype A and IncFIB/IncK/B replicons. The exclusive detection of the cif gene in pigeon-derived isolates, with a unique plasmid profile lacking the ehxA gene, suggests distinctive pathogenic mechanisms in these strains. Similarly, the stcE and etpD genes were found solely in O157 isolates, linked to plasmids with the ehxA subtype B and IncFIB replicon type, reinforcing their specific association with this serogroup. Our findings also reveal that the ehxA subtype A is more prevalent in STEC isolates from sheep and goats, highlighting the need for further investigation into the relationship between ehxA subtypes and different animal hosts. The widespread occurrence of IncFIB plasmids across all studied hosts aligns with their known prevalence in other E. coli pathotypes, suggesting a broad role in E. coli pathogenicity. The observed similarities in plasmid-encoded virulence gene profiles among sheep, goat, and cattle hosts, despite their differing ecological niches, indicate a complex and heterogeneous plasmid landscape. These insights necessitate further research to fully elucidate the diversity of plasmid-encoded virulence factors and their implications for STEC pathogenicity, essential for developing targeted strategies to mitigate the risks posed by highly pathogenic STEC variants.
Methods
STEC isolates investigated
A total of 96 STEC strains were investigated in this study. Of these, 50 collection strains were obtained from various provinces in Iran, including Tehran, Razavi Khorasan, Semnan, Mazandaran, and Khuzestan. These isolates were collected from different animal hosts (cattle, sheep, goats, and pigeons) between 2015 and 2018, as documented in our previous studies6,44. In addition, 46 new STEC strains were included, which were isolated from 75 fecal samples from cattle, 70 from sheep, and 15 from goats in Razavi Khorasan province between April and June 2022 (strain characteristics are provided in Table S1).
DNA extraction
All E. coli isolates were confirmed by culturing on MacConkey agar and Eosin-Methylene Blue (EMB) agar, followed by biochemical tests. Each isolate was then cultured on Luria-Bertani (LB) agar and incubated for 24 h at 37 °C. After overnight incubation, total genomic DNA was extracted using a boiling method. Briefly, a loopful of confluent growth from the LB agar culture was suspended in 350 µL of molecular-grade water in sterile microtubes and boiled for 10 min. The samples were then centrifuged at 15,000×g for 15 min, and the supernatants were used as templates for end-point PCR assays. DNA samples were stored at − 20 °C until further analysis.
Strains’ characterisations
To confirm the STEC genotype for both collection and new strains, we utilized a multiplex PCR targeting stx1, stx2, eae, and ehxA, as previously described45. For detecting stx in pigeon isolates, a single primer pair was used to amplify stx2f7. Additionally, we performed three separate PCR assays to identify the key stx2 subtypes associated with human disease: stx2a, stx2c, and stx2d46. E. coli O157 strain (ATCC 35218) and E. coli O157 strain Sakai (ATCC BAA-460) served as positive controls, while the strain T5B-Ir (KJ397538) from the Ferdowsi University of Mashhad (FUM) collection was used as a control for stx2f.
Our study focused on 14 important serogroups associated with human diseases: O26, O45, O55, O80, O91, O103, O104, O111, O113, O121, O128, O145, O146, and O157. We used PCR to identify these serogroups specifically in new STEC isolates using the various panels described earlier47,48. Additionally, we double-checked the collection strains for the same serogroups. We also tested all isolates for the O80 and O146 serogroups using methods outlined in previous studies49,50. E. coli O157 (295 EC-TMU) and FUM collection strains were used as positive controls.
PCR for plasmid-encoded virulence genes
We conducted various PCR assays to detect the key plasmid-associated virulence genes in STEC strains, as outlined in previous studies26,29,38,51,52,53,54,55,56,57,58,59. The genes examined included: astA, Colicin M (cma), Colicin B (cba), T6SS (rhsA1, rhsA2, rhsC), stcE, katP, toxB, espP, subAB, IhaG, LAA iha, saa, T2SS (etpD), cif, and epeA (Table 2). E. coli O157:H7 strain Sakai (ATCC BAA-460) was used as a positive control for all PCR reactions.
EhxA subtyping
Six different genetic ehxA subtypes (A–F) were identified using PCR in conjunction with restriction fragment length polymorphism (RFLP) analysis. We performed PCR amplification of the entire ehxA gene (3 kbp) using primers described by Lorenz et al.18. For RFLP analysis, 9 µL aliquots of the ehxA PCR products were digested with 5 U of TaqI restriction enzyme (Thermo Fisher Scientific) and 1.1 µL of 10× restriction enzyme buffer, followed by incubation at 65 °C for 90 min18. The digestion mixture was then analyzed by DNA electrophoresis on a 2% agarose gel containing the safe stain dye Green Viewer. FUM collection strains and E. coli O157:H7 (295 EC-TMU) were used as the positive controls.
Plasmid replicon typing
The plasmid replicon types of the STEC isolates were determined using the method developed by Johnson et al.19. This method utilizes three multiplex PCR panels to identify 17 replicon types. The panels are as follows: panel 1: (B/O, FIC, A/C, P, T); panel 2: (K/B, W, FIIA, FIA, FIB, Y); and panel 3: (I1, Frep, X, HI1, N, HI2, L/M). We tested these panels on our new STEC isolates, while the collection strains were double-checked for their plasmid replicon types. FUM collection strains were used as positive controls. Figure 4 illustrates the workflow of the study.

This workflow outlines the plasmid profiling methodology for the 96 STEC strains, covering steps from sample collection to data interpretation.
Statistical analysis
The correlations between plasmid-encoded virulence genes, ehxA subtypes, and plasmid replicon types with studied animal hosts and stx2-positive isolates was illustrated using GraphPad Prism version 9.3.1 (https://www.graphpad.com/updates/prism-931-release-notes). The frequency comparisons between different groups were analyzed using Chi-square (χ2) and Fisher’s exact tests in SPSS 26.0 software (https://www.ibm.com/support/pages/downloading-ibm-spss-statistics-26). Results with a p-value of less than 0.05 were considered statistically significant and mentioned in the text.
Data availability
All data supporting the findings of this study are available within the manuscript and its supplementary information file. Raw data, are available from the corresponding author upon reasonable request.
References
Kaper, J. B., Nataro, J. P. & Mobley, H. L. T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2, 123–140 (2004).
Maldonado, N. et al. Whole-genome characterisation of Escherichia coli isolates from patients with bacteraemia presenting with sepsis or septic shock in spain: a multicentre cross-sectional study. Lancet Microbe. 5, e390–e399 (2024).
Thomas, C. M. et al. Shiga toxin-producing Escherichia coli O157:H7 outbreak associated with school field trips at a farm animal exhibit—Tennessee, September–October 2023. Zoonoses Public. Health. https://doi.org/10.1111/zph.13161 (2024).
Nemati, A. et al. Shiga Toxin-Producing Escherichia coli (STEC) in developing countries: A 10-Year review with global perspective. Microorganisms 13, 1529. https://doi.org/10.3390/microorganisms13071529 (2025).
Kalalah, A. A., Koenig, S. S. K., Bono, J. L., Bosilevac, J. M. & Eppinger, M. Pathogenomes and virulence profiles of representative big six non-O157 serogroup Shiga toxin-producing Escherichia coli. Front. Microbiol. 15 (2024).18:15:1364026
Tomeh, R., Nemati, A., Hashemi Tabar, G., Tozzoli, R. & Badouei, M. A. Antimicrobial resistance, β-lactamase genotypes, and plasmid replicon types of Shiga toxin-producing Escherichia coli isolated from different animal hosts. J. Appl. Microbiol. 135, lxae059 (2024).
Schmidt, H. et al. A new Shiga toxin 2 variant (Stx2f) from Escherichia coli isolated from pigeons. Appl. Environ. Microbiol. 66, 1205–1208 (2000).
Wang, X. et al. A comprehensive review on Shiga toxin subtypes and their Niche-Related distribution characteristics in Shiga-Toxin-Producing E. coli and other bacterial hosts. Microorganisms 12, 687 (2024).
McMahon, T. et al. Real-time PCR primers and probes for the detection of Shiga toxin genes, including novel subtypes. Int. J. Food Microbiol. 419, 110744 (2024).
Byrne, L., Adams, N. & Jenkins, C. Association between Shiga Toxin–Producing Escherichia coli O157:H7 Stx gene subtype and disease severity, england, 2009–2019. Emerg. Infect. Dis. 26, 2394–2400 (2020).
White, A. E., Jervis, R. H., Wilson, E. & Scallan walter, E. Improving occupational health surveillance for enteric infections. Zoonoses Public. Health. 71, 381–391 (2024).
Paton, J. C. & Paton, A. W. Pathogenesis and diagnosis of Shiga Toxin-Producing Escherichia coli infections. Clin. Microbiol. Rev. 11, 450–479 (1998).
Karama, M. et al. Molecular profiling and antimicrobial resistance of Shiga toxin-producing Escherichia coli O26, O45, O103, O121, O145 and O157 isolates from cattle on cow-calf operations in South Africa. Sci. Rep. 9, 11930 (2019).
Orlek, A. et al. Plasmid classification in an era of Whole-Genome sequencing: application in studies of antibiotic resistance epidemiology. Front. Microbiol. 8, 1–10 (2017).
Nemati, A., Gigliucci, F., Morabito, S. & Badouei, M. A. Virulence plasmids in edema disease: Insights from whole-genome analysis of Porcine O139:H1 Shiga toxin-producing Escherichia coli (STEC) strains. Front Cell. Infect. Microbiol 15 (2025).
Lorenz, S. C., Monday, S. R., Hoffmann, M., Fischer, M. & Kase, J. A. Plasmids from Shiga Toxin-Producing Escherichia coli strains with rare enterohemolysin gene (ehxA) subtypes reveal pathogenicity potential and display a novel evolutionary path. Appl. Environ. Microbiol. 82, 6367–6377 (2016).
Sim, E. M. et al. Added value of genomic surveillance of virulence factors in Shiga Toxin-Producing Escherichia coli in new South wales, Australia. Front. Microbiol. 12, 713724 (2021).
Lorenz, S. C., Fischer, M. & Kase, J. A. Improved PCR-RFLP method for the identification of Escherichia coli enterohemolysin (ehxA) subtypes. J. Microbiol. Methods. 100, 24–26 (2014).
Johnson, T. J. et al. Plasmid replicon typing of commensal and pathogenic Escherichia coli isolates. Appl. Environ. Microbiol. 73, 1976–1983 (2007).
Askari Badouei, M. et al. Distribution and molecular analysis of subtilase cytotoxin gene (subAB) variants in Shiga toxin-producing Escherichia coli (STEC) isolated from different sources in Iran. Vet. Ital. 60 (2024).
Karama, M. et al. Virulence characteristics and antimicrobial resistance profiles of Shiga Toxin-Producing Escherichia coli isolates from humans in South africa: 2006–2013. Toxins (Basel). 11, 424 (2019).
Leonard, S. R., Mammel, M. K., Rasko, D. A. & Lacher, D. W. Hybrid Shiga Toxin-Producing and enterotoxigenic Escherichia sp. Cryptic lineage 1 strain 7v harbors a hybrid plasmid. Appl. Environ. Microbiol. 82, 4309–4319 (2016).
Mangroliya, D., Adhyaru, H., Kabariya, J. & Ramani, V. Genomic insights into plasmid mediated AMR genes, virulence factors and mobile genetic elements in Raw milk Escherichia coli from gujarat, India. Sci. Rep. 15, 6320 (2025).
Nemati, A., Askari Badouei, M., Hashemi Tabar, G., Morabito, S. & Dadvar, A. Molecular and in Silico analyses for detection of Shiga toxin-producing Escherichia coli (STEC) and highly pathogenic enterohemorrhagic Escherichia coli (EHEC) using genetic markers located on plasmid, O Island 57 and O Island 71. BMC Vet. Res. 20, 413 (2024).
Brunder, W., Schmidt, H., Frosch, M. & Karch, H. The large plasmids of Shiga-toxin-producing Escherichia coli (STEC) are highly variable genetic elements. Microbiology 145, 1005–1014 (1999).
Vazquez-Lopez, J. & Navarro-Garcia, F. In silico analyses of core proteins and putative effector and immunity proteins for T6SS in enterohemorrhagic E. coli. Front Cell. Infect. Microbiol 10 (2020).
Navarro-Garcia, F., Ruiz-Perez, F., Cataldi, Á. & Larzábal, M. Type VI secretion system in pathogenic Escherichia coli: structure, role in virulence, and acquisition. Front. Microbiol. 10, 1965 (2019).
Wan, B. et al. Type VI secretion system contributes to enterohemorrhagic Escherichia coli virulence by secreting catalase against host reactive oxygen species (ROS). PLOS Pathog. 13, e1006246 (2017).
Colello, R. et al. Identification and detection of Iha subtypes in LEE-negative Shiga toxin-producing Escherichia coli (STEC) strains isolated from humans, cattle and food. Heliyon 5, e03015 (2019).
Colello, R. et al. Expression of hes, iha, and TpsA codified in locus of adhesion and autoaggregation and their involvement in the capability of Shiga toxin-producing Escherichia coli strains to adhere to epithelial cells. BMC Res. Notes. 16, 163 (2023).
Tarr, P. I. et al. Iha: a novel Escherichia coli O157:H7 Adherence-Conferring molecule encoded on a recently acquired chromosomal Island of conserved structure. Infect. Immun. 68, 1400–1407 (2000).
Samba-Louaka, A., Nougayrède, J. P., Watrin, C., Oswald, E. & Taieb, F. The enteropathogenic Escherichia coli effector Cif induces delayed apoptosis in epithelial cells. Infect. Immun. 77, 5471–5477 (2009).
Hsu, Y. et al. Structure of the cyclomodulin Cif from pathogenic Escherichia coli. J. Mol. Biol. 384, 465–477 (2008).
Loukiadis, E. et al. Distribution, functional expression, and genetic organization of cif, a Phage-Encoded type III-Secreted effector from enteropathogenic and enterohemorrhagic Escherichia coli. J. Bacteriol. 190, 275–285 (2008). Distribution.
Samba-Louaka, A. et al. Bacterial cyclomodulin Cif blocks the host cell cycle by stabilizing the cyclin-dependent kinase inhibitors p21 waf1 and p27 kip1. Cell. Microbiol. 10, 2496–2508 (2008).
Van Hoek, A. H. A. M. et al. Comparative genomics reveals a lack of evidence for pigeons as a main source of stx2f-carrying Escherichia coli causing disease in humans and the common existence of hybrid Shiga toxin-producing and enteropathogenic E. coli pathotypes. BMC Genom. 20, 271 (2019).
Lathem, W. W. et al. StcE, a metalloprotease secreted by Escherichia coli O157:H7, specifically cleaves C1 esterase inhibitor. Mol. Microbiol. 45, 277–288 (2002).
Lathem, W. W., Bergsbaken, T., Witowski, S. E., Perna, N. T. & Welch, R. A. Acquisition of stce, a C1 esterase Inhibitor–Specific metalloprotease, during the evolution of Escherichia coli O157:H7. J. Infect. Dis. 187, 1907–1914 (2003).
Maria, F. et al. Virulence profiling and molecular typing of Shiga Toxin-Producing E. coli (STEC) from human sources in Brazil. Microorganisms 8, 171 (2020).
Lorenz, S. C. et al. Prevalence of hemolysin genes and comparison of EhxA subtype patterns in Shiga Toxin-Producing Escherichia coli (STEC) and Non-STEC strains from clinical, food, and animal sources. Appl. Environ. Microbiol. 79, 6301–6311 (2013).
Salaheen, S. et al. Genomic diversity of antimicrobial-resistant and Shiga toxin gene-harboring non-O157 Escherichia coli from dairy calves. J. Glob Antimicrob. Resist. 33, 164–170 (2023).
Castro, V. S. et al. Inconsistent PCR detection of Shiga toxin-producing Escherichia coli: insights from whole genome sequence analyses. PLoS One. 16, e0257168 (2021).
Ohnishi, M. et al. Genomic diversity of enterohemorrhagic Escherichia coli O157 revealed by whole genome PCR scanning. Proc. Natl. Acad. Sci. 99, 17043–17048 (2002).
Badouei, M. A., Taban, H., Nemati, A., Santos, L. F. & Dos Molecular serotyping of Shiga toxin-producing Escherichia coli (STEC) of animal origin in Iran reveals the presence of important non-O157 seropathotypes. Vet. Res. Forum. 14, 267–274 (2023).
Paton, A. W. & Paton, J. C. Detection and characterization of Shiga toxigenic Escherichia coli by using multiplex PCR assays for Stx 1, Stx 2, eaea, enterohemorrhagic E. coli hlya, Rfb O111, and Rfb O157. J. Clin. Microbiol. 36, 598–602 (1998).
Scheutz, F. et al. Multicenter evaluation of a Sequence-Based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 50, 2951–2963 (2012).
DebRoy, C., Roberts, E. & Fratamico, P. M. Detection of O antigens in Escherichia coli. Anim. Heal Res. Rev. 12, 169–185 (2011).
Iguchi, A. et al. Escherichia coli O-Genotyping PCR: a comprehensive and practical platform for molecular O serogrouping. J. Clin. Microbiol. 53, 2427–2432 (2015).
Istituto Superiore di Sanità. Identification of the VTEC serogroups mainly associated with human infections by conventional PCR amplification of O-associated genes. EU Ref. Lab. E. coli 1–8 (2020).
Liu, Y., DebRoy, C. & Fratamico, P. Sequencing and analysis of the Escherichia coli serogroup O117, O126, and O146 O-antigen gene clusters and development of PCR assays targeting serogroup O117-, O126-, and O146-specific DNA sequences. Mol. Cell. Probes. 21, 295–302 (2007).
Paton, A. W. & Paton, J. C. Direct detection and characterization of Shiga toxigenic Escherichia coli by multiplex PCR for Stx 1, Stx 2, eae, ehxa, and Saa. J. Clin. Microbiol. 40, 271–274 (2002).
Leyton, D. L., Sloan, J., Hill, R. E., Doughty, S. & Hartland, E. L. Transfer region of pO113 from enterohemorrhagic Escherichia coli : similarity with R64 and identification of a novel Plasmid-Encoded autotransporter, EpeA. Infect. Immun. 71, 6307–6319 (2003).
Yamamoto, T. & Nakazawa, M. Detection and sequences of the enteroaggregative Escherichia coli heat-stable enterotoxin 1 gene in enterotoxigenic E. coli strains isolated from piglets and calves with diarrhea. J. Clin. Microbiol. 35, 223–227 (1997).
Setia, A., Bhandari, S. K., House, J. D., Nyachoti, C. M. & Krause, D. O. Development and in vitro evaluation of an Escherichia coli probiotic able to inhibit the growth of pathogenic Escherichia coli K881. J. Anim. Sci. 87, 2005–2012 (2009).
Bustamante, A. V., Sanso, A. M., Lucchesi, P. M. A. & Parma, A. E. Multiplex PCR assay for the detection of five putative virulence genes encoded in verotoxigenic Escherichia coli plasmids. Curr. Microbiol. 62, 1411–1415 (2011).
Amézquita-López, B. A., Quiñones, B., Lee, B. G. & Chaidez, C. Virulence profiling of Shiga toxin-producing Escherichia coli recovered from domestic farm animals in Northwestern Mexico. Front. Cell. Infect. Microbiol. 4, 1–9 (2014).
Tarr, C. L. et al. Molecular characterization of a serotype O121:H19 clone, a distinct Shiga Toxin-Producing clone of pathogenic Escherichia coli. Infect. Immun. 70, 6853–6859 (2002).
Wieler, L. H. et al. Enterohemorrhagic Escherichia coli (EHEC) strains of serogroup O118 display three distinctive clonal groups of EHEC pathogens. J. Clin. Microbiol. 38, 2162–2169 (2000).
Slanec, T., Fruth, A., Creuzburg, K. & Schmidt, H. Molecular analysis of virulence profiles and Shiga toxin genes in Food-Borne Shiga toxin-Producing Escherichia coli. Appl. Environ. Microbiol. 75, 6187–6197 (2009).
Acknowledgements
We would like to extend our deepest gratitude to our colleagues in the Escherichia coli research lab at Ferdowsi University of Mashhad for their invaluable support and contributions to this study. We would like to express our sincere thanks to Dr. Stefano Morabito for inspring us to work on this project and Dr. Federica Gigliucci (EU Reference Laboratory for STEC in Istituto Superiore di Sanità, Rome) for her expert guidance and valuable feedback, which greatly enhanced the quality of this research. We also thank Mrs. Sommaye Bagherzadeh from bacteriology lab at Ferdowsi University of Mashhad for her kind assistance to perform some laboratory experiments.
Funding
This project was supported by Ferdowsi University of Mashhad under the Grant No. FUM. 57299.
Author information
Authors and Affiliations
Contributions
A.N. conducted the experiments, data analyses, and drafted the manuscript and draw the figures. M.A.B. designed the study, supervised the project, revised the data analyses, and edited the manuscript. G.H. assisted on data analysis and critically reviewed the manuscript. M.H. carried out some experiments, reviewed, and edited the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
All procedures involving animals and their care in this study were approved (No. IR140357299) by Iran National Committee for Ethics in Biomedical Research. Moreover, a verbal informed consent was obtained from the owners of the companion animals. The research committee of Ferdowsi University of Mashhad reviewed and approved that all the study protocols were conducted in accordance with the related guidelines and regulations (No. FUM57299). The study was carried out in accordance with the ARRIVE guidelines (http://www.nc3rs.org.uk/page.asp? id=1357).
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Nemati, A., Askari Badouei, M., Hashemitabar, G. et al. Shiga toxin-producing Escherichia coli plasmid diversity reveals virulence potential and control opportunities in animal hosts. Sci Rep 15, 26562 (2025). https://doi.org/10.1038/s41598-025-11905-y
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-11905-y
Keywords
This article is cited by
-
Climate-driven co-evolution of antimicrobial resistance and virulence in Escherichia coli on dairy farms: unraveling adaptive genetic signatures with novel SSCP-PCR
World Journal of Microbiology and Biotechnology (2025)
