
Abstract
The cooperative binding of molecular agents onto a substrate is pervasive in living systems. To study whether a system shows cooperativity, one can rely on a fluctuation analysis of quantities such as the number of substrate-bound units and the residence time in an occupancy state. Since the relative standard deviation from the statistical mean monotonically decreases with the number of binding sites, these techniques are only suitable for small enough systems, such as those implicated in stochastic processes inside cells. Here, we employ a general-purpose grand canonical Hamiltonian description of a small one-dimensional (1D) lattice gas with either nearest-neighbor or long-range interactions as prototypical examples of cooperativity-influenced adsorption processes. First, building upon previous work on finite-size one-dimensional Ising-type models, we elucidate how the strength and sign of the interaction potential between neighboring bound particles on the lattice determine the intensity of the fluctuations of the mean occupancy and the nature of bound particle-particle correlations. We present our theoretical results, which extend beyond standard analysis, in a novel, physically transparent form. We explore simple limiting cases of the parameter space in greater detail, examine the complete probability distribution functions for occupation, and relate these to the shape and strength of the fluctuations. Second, we leverage these relationships to compare the theoretical predictions of our model to data from single molecule experiments on bacterial flagellar motors (BFM) of Escherichia coli. In this way, we find evidence that cooperativity controls the mechano-sensitive dynamical assembly of the torque-generating units, the so-called stator units, onto the BFM and thereby arrive at an estimate of the stator-stator interaction potential from our fluctuation analysis. Furthermore, we attempt to clarify the link between occupation fluctuations and the adaptability of the BFM. Finally, we conclude that the system resides in a sweet spot of the parameter space (phase diagram) with characteristics suitable for a smoothly and widely adaptive system, while minimizing fluctuations.
Similar content being viewed by others
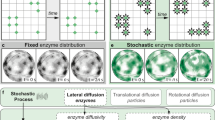
Introduction
Cooperativity is a pervasive phenomenon that emerges in biological systems at different time and length scales, and levels of complexity1: from the molecular scale2,3,4,5 to multiple-cell organization6, extending to the emergence of collective behavior in many-body systems. Biologically important problems concerning the cooperative properties of regulatory enzymes7 and ligand binding on biological membranes8, as well as, more recently, cell adhesion9, have been extensively and successfully studied using simple one and two-dimensional Ising-type physical models10.
We focus here on cooperative processes involving the adsorption of ligands onto a substrate disposing of a limited number of binding sites11. Our main goal is to develop a general method for assessing how short- and long-range interactions between substrate-bound ligands affect stochastic fluctuations in the number of adsorbed units. In the presence of cooperativity, small system size effects strongly influence the amplitude of the stochastic fluctuations and the shape of the equilibrium probability distribution function (PDF) describing substrate occupancy. By studying these characteristic signatures of cooperativity, we propose (1) a criterion to determine whether any given adsorption system exhibits cooperative or anti-cooperative behavior and (2) a method to quantify the amplitude of the ligand-ligand interaction potential.
To this end, we base our study on a minimal finite size 1D lattice gas model with interacting units in equilibrium with a thermal bath and chemical reservoir, described using a general-purpose grand canonical Hamiltonian with short- or long-range interactions. With it, we study how fluctuations at equilibrium, described by the standard deviation of the occupancy, are influenced by the model parameters, namely the ligand binding energy, the ligand-ligand interaction potential, and the chemical potential of the bulk reservoir. To get closer to observable quantities, we invert the problem by determining the key formulae relating the occupancy standard deviation and PDF to the experimentally accessible average occupancy of the system. In this way, we are able to quantify how, at fixed average occupancy, increasing cooperativity leads to an increase in fluctuations and how increasing anti-cooperativity leads to a decrease. In the process, we highlight the saturation of fluctuations for both strong cooperativity and strong anti-cooperativity, as well as a subtle difference between fluctuations near half-filling for systems with even and odd binding sites in the case of strong anti-cooperativity. This difference arises due to frustration and is an expected consequence of the system’s constraints. Finally, we apply the model by comparing our theoretical predictions with experimental data for the bacterial flagellar motor (BFM) of E. coli12,13, a macro-molecular complex that has previously been found to exhibit cooperative binding of the torque-generating (stator) units14. In this previous work, although evidence for cooperativity was uncovered in stator unit assembly dynamics, no estimate of the stator-stator interaction potential was proposed, which is our goal here. More precisely, we aim to achieve this goal by analyzing previously published single-molecule data15,16 through the lens of cooperativity, using the present theoretical framework as a foundation (see Section “Experimental data” for details on the data and experimental setup). In Methods Section “Effective Hill coefficient”, we make the connection between our model-based approach and phenomenological approaches based on effective Hill coefficients11,17.
We also discuss the concept of motor adaptability to changes in external conditions and set up a general framework to assess its desired level depending on how the motor is expected to function. Our fluctuation analysis provides evidence for cooperativity by leading to an estimate of the stator-stator interaction potential between 1 and 2 \(k_{\text {B}}T\) for the short-range model, a result that is coherent with this system’s expected smooth adaptive nature to external stimuli18,19,20,21,22. We contrast this moderate cooperative behavior found for stator unit binding to the BFM with the highly cooperative behavior of rotor-switching units that are believed to be behind the rapid stochastic switching in BFM rotational direction in the context of bacterial chemotaxis17,23.
Results
In this section, in the spirit of Skoge et al.24,25,26 and Tu27, who studied the dynamics of cooperativity in chemical sensing to better understand bacterial chemotaxis, we employ a minimal prototypical one dimensional lattice gas model of cooperative particle binding on a substrate of finite size. We then obtain an analytical expression for the fluctuations of the mean occupancy at equilibrium as a function of the relevant model parameters, including the number of binding sites and the interaction potential between bound particles, which acts as a proxy for system cooperativity. Finally, we apply these results to investigate the characteristic signatures of cooperativity by establishing a general criterion for its manifestation and then propose a procedure to estimate the interaction potential from experimental data.
Notably, a fluctuation analysis of the type used here was previously performed by Liu and Dilger28, utilizing finite-size one- and two-dimensional Ising models to investigate cooperative phenomena in ligand-gated ion channels. Modeling of this adsorption system is complicated, in part because of geometrical questions, because it is not clear if the ion channel network can be approximated by a regular Ising like lattice (disorder could play an important role), which could explain why their attempt to quantify ion channel cooperativity was inconclusive. Our work complements and extends the work of Liu and Dilger by providing a comprehensive theoretical analysis: we present the theoretical results in a physically transparent form, explore simple limiting cases in greater detail, and examine the full probability distribution functions (PDFs) for occupation.
Most of the applications of the 1D Ising model, or its generalizations, to the study of cooperativity in biological systems are restricted to the simplifying limit of very large (or infinite) system size for which the type of fluctuation and correlation analysis we perform is not pertinent (fluctuations vanish in this limit and correlations cannot extend over the whole system), see e. g. the recent work on the cooperative binding of cofilin to actin filaments29. Here, in contrast, as in the work of Liu and Dilger on fluctuations in ligand binding to ion channels28 and Rice et al.30 on cooperative correlations in cardiac thin filament activation, we explicitly focus on the effects that the finite size of the system has on both fluctuations and correlations, the amplitudes and shapes of which can provide valuable insights into the internal configuration of the system.
Our intentional use of a minimalist model strips down the complexity to the bare essentials required to isolate and examine the fundamental roles of cooperativity and anti-cooperativity in characterizing fluctuations. Although derived from a minimal model, the core insights of our study serve as universal principles still applicable to refined and more biologically adapted models. The type of model adopted here is equivalent to the Ising-type models successfully employed to understand the role of (strong) cooperativity in bacterial chemotaxis, although our exact detailed analytical treatment of finite size systems goes beyond the usual treatments proposed for the Ising-type models in this context28,31. Despite the simplicity of the model, the small system size leads to a panoply of physical behavior and key insight into the functioning of a complex biological motor.
Lattice gas model
Consider a periodic 1D lattice with L binding sites in contact with a heat and particle reservoir of constant temperature T and effective chemical potential \(\mu\). We assign to each binding site i (\(i=1,\ldots , L\)) a binary variable \(\varphi _i\): \(\varphi _i=1\) means that a particle (ligand) occupies the i-th site; otherwise, \(\varphi _i=0\). The array \(\Phi =\{ \varphi _i \} = \left( \varphi _1,\ldots ,\varphi _L\right)\) uniquely defines one of the \(2^L\) possible microscopic configurations (or microstates). The relative occupancy in a given microstate is \(\varphi = L^{-1} \sum _{i=1}^L\varphi _i\). We say that the system is in mesostate N if the number of particles on the lattice, i. e. the occupancy of the system equals \(N = L\varphi\). There are \(C^L_N = L! /\left[ N! \left( L-N\right) !\right]\) microstates \(\Phi\) consistent with the occupancy N; in this case, we say that the mesostate N has multiplicity \(C^L_N\).
The energy of the system depends on the specific microscopic configuration \(\Phi\) according to the grand canonical effective 1D short-range interaction Hamiltonian:
Since we choose periodic boundary conditions, \(i= L+1\) corresponds to \(i=1\). Furthermore, we introduce \(\beta = 1/(k_{\text {B}}T)\) with the Boltzmann constant \(k_{\text {B}}\), rendering the two adjustable control parameters of our model, J and \(\mu\), dimensionless. The effective chemical potential, \(\mu\), is considered to be made up of two parts, \(\mu = \mu _\textrm{r} - \varepsilon\), where \(\mu _\textrm{r}\) is the chemical potential of the external particle reservoir and \(\varepsilon < 0\) is the binding energy of particle adsorption onto the substrate. The nearest-neighbor interaction J is attractive for \(J>0\) (cooperativity) and repulsive for negative values (anti-cooperativity). For fixed J the number of bound particles can be modulated by varying \(\mu\) through \(\mu _\textrm{r}\) and/or \(\varepsilon\). When \(J=0\) we recover the usual non-interacting (Hill–Langmuir) model.
In the Methods Section “Infinite range (IR) model”, we will compare the predictions of this short-range model with those of the infinite-range model in which all pairs of particles interact with uniform strength, irrespective of their relative distances on the lattice. As detailed below, this model effectively serves as a proxy for the more general long-range lattice gas model.
Mean occupancy and its standard deviation
The Hamiltonian Eq. (1) falls into the universality class of the 1D short-range lattice gas model. Hence, we can map our model to the 1D Ising model (see32,33 for comprehensive overviews). It is, therefore, possible to derive analytical expressions for the grand partition function and both the first and second moments of the relative occupancy in thermodynamic equilibrium by employing the transfer matrix formalism. In what follows, we will express our theoretical results in terms of relative quantities, such as relative occupancy, even when absolute values are experimentally available. This standardized approach allows for consistent qualitative and quantitative comparison across systems of different (finite) size to which our general-purpose approach can be applied, simplifies the analysis, and enhances the applicability across different experimental conditions.
By taking the derivative of the grand partition function \(\Xi\) with respect to \(\mu\), we can calculate the mean relative occupancy at equilibrium, \(\langle \varphi \rangle = L^{-1} \sum _{i=1}^L\langle \varphi _i\rangle =(L \Xi )^{-1}\partial _\mu \Xi\):
where \(X = \left( J+\mu \right) /2\), and \(\xi\) is the equilibrium correlation length of the system (given in number of lattice sites), which can be rewritten in terms of J and \(\mu\) as follows (we refer to Section “1D short-range lattice gas (SRLG)” for more details):
The mean equilibrium occupancy equals \(\langle N\rangle = L\langle \varphi \rangle\).
We focus on small systems with around ten binding sites. As discussed below, such systems will turn out to possess the important feature that there should be enough theoretical resolution in the standard deviation to discriminate between different values of J for couplings within the bio-physically expected range (\(0< J <5\)). For purposes of illustration in what follows, unless otherwise stated, we shall use \(L=13\). This particular choice will allow us to both illustrate the general behavior of such small systems and compare with greater ease the theoretical results presented here to the experimental data when we later, in Section “Application to the bacterial flagellar motor (BFM)”, apply our model to the study of the recruitment of torque-generating units (or stator units) onto the BFM of E. coli, which has recently shown to recruit up to \(\sim 13\) stator units under high-load conditions15, updating earlier results from previous studies34,35. It should be kept in mind that our theoretical results are valid for any value of L and that our conclusions for small systems apply to any sufficiently small value of L. In particular, while the present work considers single-molecule bead assay data from E. coli, our analysis can be readily extended to the BFMs of other bacterial species, which are known to have a different, though still limited, maximum number of torque-generating stator units36,37. This generalization is facilitated by the fact that we express our results in terms of relative, rather than absolute, occupancies.
We propose that the operational definition of “small system” be simply those systems for which fluctuations, as measured by the standard deviation of relative occupancy, are within experimental resolution and can be used to extract the sign and amplitude of the interaction strength within the usual biophysical range. A more careful rendering of this definition can be found in Section “System size, fluctuations, and experimental precision”. Such a definition is not contingent on the amplitude of (anti)cooperativity and therefore can encompass even the non-interacting (Hill–Langmuir) model.
Figure 1a depicts the mean relative occupancy from Eq. (2) as a function of the two control parameters of the system: J and \(\mu\). The half-filling (HF) contour line \(\langle \varphi \rangle =0.5\) cuts the plot diagonally, meaning that the lattice is, on average, half-filled whenever \(J=-\mu\) (or \(X=0\)) holds. Below (above) that line, the system’s mean occupancy is lower (higher) than 50 %. Contour lines run together close to the plot’s bottom right sector diagonal, corresponding to positive J and negative \(\mu\). In this sector defined by \(X\approx 0\), for \(J \gg 1\), the mean occupancy for a fixed positive value of the interaction potential becomes extremely sensitive to slight chemical potential variations. Systems characterized by parameters in that range behave like biological switches, nearly jumping from zero to full occupancy with small changes in \(\mu\). On the contrary, more adaptive systems, that is, systems whose occupancy varies more smoothly from zero to full occupancy as a function of \(\mu\), would be located in the central and upper left sector of Fig. 1a.

Contour plots of the mean relative occupancy in equilibrium \(\langle \varphi \rangle\) (left), and the standard deviation of the relative occupancy in equilibrium \(\sigma = \sqrt{\langle \varphi ^2 \rangle - \langle \varphi \rangle ^2}\) (right), as functions of the dimensionless interaction potential J and chemical potential \(\mu\), according to expressions from Eq. (2) and (4), for a system of size \(L=13\). Contour plots of the mean relative occupancy and its standard deviation for an even system size, \(L = 14\), are presented in Fig. 14 and 15.
For the short-range lattice gas model from Eq. (1), the standard deviation \(\sigma\) of the occupancy at equilibrium can be expressed in terms of the derivative of the mean relative occupancy with respect to the chemical potential as follows (cf. Section “1D short-range lattice gas (SRLG)”): \(\sigma = \sqrt{\langle \varphi ^2 \rangle - \langle \varphi \rangle ^2} = L^{-1/2} \sqrt{ \partial _\mu \langle \varphi \rangle }\). Applying this result to Eq. (2), we obtain a compact analytical expression for \(\sigma\) as a function of \(\mu\), J, and L:
The dependency of \(\sigma\) on J and \(\mu\) for fixed L is depicted in Fig. 1b. In what follows, we will use \(\sigma\) as a measure of fluctuations, hence the stochastic nature of the system.
In Fig. 2, we compare the average relative occupancy, \(\langle \varphi \rangle\), and the standard deviation, \(\sigma\), as functions of \(\mu\) for two vertical cuts in the above contour plots: \(J = 0\) (Hill–Langmuir) and \(J = 2\). We note that with increasing J the \(\langle \varphi \rangle\) curve sharpens and shifts to lower values of \(\mu\) and that the maximum in \(\sigma\) increases in amplitude and also shifts to lower values of \(\mu\). These shifts are due to the displacement with increasing J of the value of \(\mu\) at half-filling, \(\mu _\textrm{HF}(J) = - J\), where the slope of the \(\langle \varphi \rangle\) curve and the amplitude of the standard deviation are both largest.

Comparison between the average relative occupancy (solid line) and the standard deviation (dashed line) as functions of \(\mu\), for two values of cooperativity (\(L = 13\)): \(J = 0\) (Hill–Langmuir, blue) and \(J = 2\) (purple).
System behavior at half-filling
Fluctuations increase as \(J > 0\) grows. Along the half-filling diagonal (\(X=0\)) in Fig. 1b, they saturate at a finite global maximum as J goes to infinity.
By evaluating Eqs. (4) and (3) for \(X=0\), one obtains an explicit formula for the standard deviation at half-filling (HF) as a function of L and J:
with
which represents the maximum equilibrium correlation length at fixed J for any value of mean occupancy.
In the Hill–Langmuir limit (\(J=0\)), the correlation length vanishes, and the standard deviation at HF becomes \(1/(2\sqrt{L})\). For low couplings (\(0 \le J <2\)), the correlation length increases sub-linearly with J and is accurately given by \(\xi _\textrm{HF} \approx -1/\ln (J/4)\). In this same limit, the standard deviation increases exponentially with J, \(\sigma _\text {HF}(J)/\sigma _\text {HF}(0) \approx \text {e}^{J/4}\), an approximation valid for \(J<4\), when \(L \approx 10\) (see Fig. 16). This increase can be interpreted as a widening of the probability distribution function of states of a given N around the average value \(\langle N \rangle\), where the probability is a maximum. In the high cooperativity limit \(J \gg 1\), \(\xi _\textrm{HF}\) shows an asymptotic exponential growth, \(\xi _\textrm{HF} \sim \frac{1}{2}\text {e}^{J/2}\), which is the analog of the well-known zero temperature critical behavior of spin chains38; and, consequently, for fixed L, \(\sigma _\text {HF}\) tends to 1/2 when the correlation length surpasses L/2 and the system evolves into a bimodal one (for which only the zero and full occupancy states contribute, see below). We remark that although for fixed J, \(\sigma _\text {HF}\) vanishes as \(L^{-1/2}\) when \(L\rightarrow \infty\), as expected, in the limit where L is kept fixed and J tends to \(+\infty\), the standard deviation at HF reaches the finite (maximum, or saturation) value of one-half.
Fluctuations decrease as \(J < 0\) becomes more negative. Along the half-filling diagonal (\(X=0\)) in Fig. 1b (\(L=13\)), they saturate at a finite value as \(J \rightarrow - \infty\). This is a general result for systems with an odd number of sites. In this strong anti-cooperative limit, for small enough systems, it turns out that fluctuations depend sensitively on whether the number of lattice sites is even or odd. For L even, \(\sigma _\text {HF}\) tends to zero; for odd L, it tends to 1/(2L). This leads to the striking result that in the limit of strong anti-cooperativity, a fluctuation analysis near HF could clearly detect the difference between small systems with even and odd numbers of sites, provided that the experimental precision is high enough.
Fluctuations of the mean occupancy and their dependency on \(\mathinner {\langle {\varphi }\rangle }\) and J
Often, the experimental parameter one can measure (or control indirectly, as is the case for the BFM by varying the external load, cf. Section “Application to the bacterial flagellar motor (BFM)”) is not the chemical potential of the reservoir \(\mu _\text {r}\), but the mean number of bound ligands at equilibrium, \(\langle N \rangle\). Therefore, it is insightful to plot, in the spirit of28, the standard deviation \(\sigma\) as a function of the mean relative occupancy \(\langle \varphi \rangle\), which, in general, can be done using a parametric plot.
To unfold this relation, we express \(\langle \varphi \rangle\) and \(\sigma\), for different but fixed values of J, as functions of \(\mu\) and plot this family of curves in Fig. 3a,b for an odd and even number of lattice sites, respectively (the corresponding contour plots for the case \(L=14\) are shown in Figs. 14 and 15, analogously to Fig. 1a,b for \(L=13\)). Each curve for a given J represents the set of points of coordinates \(\left( \langle \varphi \rangle =f_J(\mu ), \,\sigma =g_J(\mu ) \right)\) with \(\mu\) spanning from \(-15\) to 30, which turns out to be a suitable one-dimensional domain to complete the plots. We have chosen the interaction potential J to take values from \(-10\) to 10 in steps of size 1. The standard deviation shows a left-right particle-hole symmetry with respect to the system half-filling: \(\langle \varphi \rangle _\text {HF}= f_J(-J)=1/2\), which is a manifestation of the underlining particle-hole symmetry of the system under study. The system has zero fluctuations when \(\langle \varphi \rangle =0\) (empty substrate) or \(\langle \varphi \rangle =1\) (full substrate), which is easy to understand because a distribution of states with non-zero probability around these average values would be incompatible with the average values themselves. For non-negative values of J, the maxima of fluctuations appear at half-filling, taking on the value of \(1/(2 \sqrt{L})\) for \(J=0\) and 1/2 for \(J\rightarrow +\infty\), and the \(\sigma\) vs. \(\langle \varphi \rangle\) curves converge asymptotically towards the limiting \(J\rightarrow +\infty\) one. More subtly, results for the anti-cooperative limit \(J\rightarrow -\infty\) depend on whether L is odd or even.

Parametric plots of the standard deviation of the relative occupancy, \(\sigma\), versus the mean relative occupancy, \(\langle \varphi \rangle\), for different values of the interaction potential: from \(J=-10\) (bottom curve) to \(J=10\) (upper curve) in steps of 1. Plots (a) and (b) correspond to systems of size \(L=13\) and \(L=14\), respectively.
A key result of the above analysis and one of our major conclusions is that for cooperative systems of size \(L\approx 10\) and mean relative occupancies not too close to 0 or 1, in the range \(0<J<7\) the curves of \(\sigma\) vs. \(\langle \varphi \rangle\) are spaced sufficiently far enough apart that the standard deviation is suited for estimating J. The same conclusion can be drawn for anti-cooperative systems of size \(L\approx 10\) and mean relative occupancies close to 1/2 in the range \(-3<J<0\). These results are illustrated in Fig. 3a for \(L=13\) and Fig. 3b for \(L=14\). Outside these ranges, the standard deviation saturates to limiting values, and therefore experimental data near these limits could only be used to put bounds on the value of J. In the absence of prior knowledge of the system size, the characteristic non-monotonic shape of the parametric plot in Fig. 3b can be exploited to estimate L. For instance, for non-negative values of J, the maximum in occupancy fluctuations corresponds to half-filling.
We will investigate the cooperative and anti-cooperative regimes in more detail in what follows, taking care to study those limits for which we were able to derive simplified exact or approximate analytical expressions for \(\sigma (\langle \varphi \rangle )\).
Fluctuations in the zero-cooperativity limit: the Hill–Langmuir case
In the absence of cooperativity (\(J=0\)), which corresponds to the case of a simple adsorption process with on-site volume exclusion interactions only, the free energy calculated from Eq. (1) reduces to an expression determined entirely by the mesostate (the occupancy) N: \(\beta \mathcal {H}_0(\Phi | N)=-\mu N\). The correlation length from Eq. (3) vanishes; hence each adsorbing site acts as an independent particle trap. Thus we recover the Hill–Langmuir adsorption model at equilibrium for which \(\langle \varphi \rangle\) takes the following typical sigmoid form as a function of \(\mu\):
The corresponding standard deviation becomes:
which can be reinterpreted to obtain
This latter expression exemplifies the general result that, in the thermodynamic limit, \(L\rightarrow \infty\), \(\langle \varphi ^2 \rangle \rightarrow \langle \varphi \rangle ^2\), and hence, as expected, the standard deviation (of the relative occupancy) tends to zero as \(L^{-1/2}\) (self-averaging property at equilibrium in the thermodynamic limit).
In Section “1D short-range lattice gas (SRLG)”, we show using a Gaussian approximation for the PDF that for \(J=0\) the standard deviation is inversely proportional to the curvature (in absolute value) of the entropy of mixing, which reaches a minimum at HF. Therefore, the standard deviation for \(J=0\) reaches a maximum there. More generally, one can show that for \(J>0\) not too large, a Gaussian approximation for the PDF is accurate and therefore the standard deviation is inversely proportional to the curvature (in absolute value) of an effective free energy, which reaches a minimum at HF.
Fluctuations in the strong cooperativity limit
Away from HF, although \(\xi\) remains finite in the large J limit, it can still be large compared with L/2 for not-too-large values of L, provided that one is not too close to zero or full filling (see Fig. 17). Taking the limits \(J \rightarrow \infty\) and \(\mu = 2 X - J \rightarrow -\infty\) simultaneously, but keeping L finite and X constant, allows Eqs. (2) to be simplified to
and \(\langle \varphi ^2 \rangle _\infty = \langle \varphi \rangle _\infty\), where \(\xi _\infty (X) = 1/(2 |X|)\), leading to an infinite J standard deviation of
which can be reinterpreted to obtain
The maximal standard deviation at half-filling is due to the large width of the distribution of states with non-zero probability compatible with \(\langle \varphi \rangle = 1/2\) (see Section “Probability distribution function (PDF) of the occupancy and its dependency on \(\langle \varphi \rangle\) and J”). This limit is easy to understand in terms of a bimodal system: only the empty and full states contribute because the probability of intermediate states is suppressed by the presence of energetically costly domain walls (as will be discussed in detail below). The bimodal system, therefore, behaves as an effective Hill–Langmuir model with a system size equal to 1, as can be seen by taking \(L=1\) in Eqs. (9). The purely bimodal model, developed in the context of chemical sensing with applications in bacterial chemotaxis, is commonly referred to as the Monod–Wyman–Changeux (MWC) model7.
Fluctuations for intermediate finite cooperativity
We have seen in Fig. 3 how, at fixed average occupancy, increasing cooperativity leads to an increase in fluctuations. A simple, insightful, and accurate way of understanding this increase can be formulated by starting from the previously derived result for the Hill–Langmuir model and introducing the concept of block domains. When \(J=0\), all sites are decoupled, and the standard deviation is non-zero for a finite-size system simply because of statistical fluctuations (absence of self-averaging). When \(J>0\), neighboring sites are coupled and can be grouped (approximately) into correlated block domains of size \(b(\xi , L) > 1\), leading to
where \(L_\textrm{eff} \equiv L/b\). When \(1< 2 \xi < L\), correlations lead to block domains that we take to be of size \(b = 2 \xi\). The above approximation allows the system to be described by a reduced number, \(L_\textrm{eff}\), of independently fluctuating block domains of size b. Since b should also tend to 1, when \(\xi \rightarrow 0\) (zero cooperativity, Hill–Langmuir); and to L, when \(2\xi /L \gg 1\) (strong cooperativity, bimodal behavior), a convenient interpolation formula between the above three cases is
By comparing the predictions of this approximation with the exact results at half-filling, where the standard deviation is a maximum, one can see that this approximation is extremely accurate (see Fig. 16).
Although the choice Eq. (14) for b leads to an accurate approximation for \(\sigma\) over the whole parameter range, it has the inconvenience for practical applications that, because \(\xi\) depends explicitly on \(\mu\), it is not a function of L, \(\langle \varphi \rangle\), and J alone. To remedy this, a useful and reasonably good global interpolation formula for \(\sigma\) as an explicit function of L, J, and \(\langle \varphi \rangle\) can be obtained by adopting a choice for \(L_\textrm{eff}\) [\(L_\textrm{eff}^{-1/2} \rightarrow 2 \sigma _\textrm{HF}(L, J)\) in Eq. (13)] suggested by an approximate analysis of the Hill coefficient (see Eq. (84), Section “Effective Hill coefficient”). This approximation, which emerges from a simple physically clear approach, is exact at half-filling for all couplings and provides an upper bound for all occupancies that becomes exact at weak and strong coupling. Starting from a weak coupling cumulant expansion, we can find another useful approximate interpolation formula for \(\sigma\) as an explicit function of L, J, and \(\langle \varphi \rangle\):
(see Section “Infinite range (IR) model”). For \(L \approx 10\), the window of maximum accuracy for this approximation for \(J<2\) is \(0.2< \langle \varphi \rangle < 0.8\). As we will see below, it encompasses the experimental data window for the system to which we apply our method (the BFM).
The two approximations are complementary (see Figs. 13 and 20) since Eq. (15) provides a lower-bound that is most accurate at low coupling and Eq. (84) provides an upper bound that is a better approximation at strong coupling.
Exact results for the standard deviation can be obtained by writing the mean square relative occupancy at equilibrium,
in terms of the 2-point correlation function, \(C_{ij} =\langle \varphi _i \varphi _j \rangle\) (with \(C_{ii} = \langle \varphi _i^2 \rangle = \langle \varphi _i \rangle = \langle \varphi \rangle\)).
Equation (16) can be used as a starting point to gain deeper physical insight into the behavior of the standard deviation \(\sigma\) than the previous result obtained directly from the grand partition function because it becomes possible to express \(\sigma\) as an explicit function of \(\langle \varphi \rangle\) and the correlation length \(\xi\), see Eq. (36).
For \(J>0\), the standard deviation is bounded by the zero and infinite cooperativity values, which can be calculated from the limiting forms of \(C_{ij}\). In the absence of cooperativity (\(J=0\)), \(C_{ij}\) factorizes for \(i\ne j\): \(C_{ij} \rightarrow \langle \varphi _i \rangle \langle \varphi _j \rangle = \langle \varphi \rangle ^2\). In this \(J=0\) limit, \(\langle \varphi ^2 \rangle\) can then be obtained directly from Eq. (16) and \(\sigma\) simplifies to \(\sigma _0\) [Eq. (9)]. In the other limit of infinite cooperativity (\(J\rightarrow \infty\)) and strong correlations, \(\xi\) as a function of \(\langle \varphi \rangle\) diverges at HF and away from HF for not too large L remains larger than L/2 (see Fig. 17). From Eq. (30) we see that for L finite in this limit \(C_{ij} \rightarrow \langle \varphi _i^2 \rangle = \langle \varphi \rangle\), and \(\sigma\) saturates at \(\sigma _\infty\) [Eq. (12)].
Fluctuations in the strong anti-cooperativity limit
By comparing the \(L=13\) and 14 cases (Fig. 3a,b), we observe that for small enough anti-cooperative systems, there is a clear difference between even and odd system sizes. This difference clearly manifests itself near HF, and arises because of frustration for odd L.
For negative values of J (anti-cooperativity) and L even the curves inflect and tend for \(J \ll -1\) towards a characteristic limiting shape with global maxima near \(\langle \varphi \rangle = 0.2\) and 0.8 and a zero at half-filling (see Fig. 10). A restricted grand partition function approach (presented in Section “Probability distribution function of the occupancy”), which retains only non-overlapping particle-hole pairs, leads to a simple but accurate approximation in the limit \(J\rightarrow -\infty\) for L even (see Fig. 10):
The corresponding standard deviation for \(1/2 \le \langle \varphi \rangle \le 1\) can be obtained by exploiting particle-hole symmetry: \(\sigma (\langle \varphi \rangle ) = \sigma (1-\langle \varphi \rangle )\).
In this limit of strong anti-cooperativity (\(J\rightarrow -\infty\)), the correlation length becomes complex, \(1/\xi \simeq i \pi + 2 \cosh (X) \text {e}^{-|J|/2}\). Hence, at HF, \(1/\xi \simeq i \pi\), leading directly to an oscillating two-point correlation function that describes a sequence of non-overlapping particle-hole pairs (anti-ferromagnetic order in Ising spin language):
where we have taken the real part of \(C_{i,i+r}\) and assumed that \(r/L \ll 1\). \(C_{i,i+r}^\textrm{HF}\) alternates between 0 and 1/2, which reflects alternating perfect anti-correlations (nearest neighbors) and correlations (next nearest neighbors) at HF in this limit of strong anti-cooperativity.
Probability distribution function (PDF) of the occupancy and its dependency on \(\langle \varphi \rangle\) and J
We can extract the equilibrium probability distribution function (PDF) of the occupancy [corresponding to the effective Hamiltonian given in Eq. (1)] for fixed J and \(\mu\), \(P(N; J, \mu )\) through exact enumeration and study how it depends on the interaction potential J. To study \(P(N; J, \langle \varphi \rangle )\) for fixed J and \(\langle \varphi \rangle\), the experimentally accessible quantity, we varied J but kept the mean occupancy fixed (by adjusting \(\mu\) to any given choice of J).
Taking \(L = 13\), Fig. 4 shows how the PDF behaves for three different positive values of the interaction potential and three characteristic values of the mean relative occupancy: \(\langle \varphi \rangle = 1/3,\,1/2,\) and 2/3, respectively. The correlation length increases with J, and so does the standard deviation; consequently, the PDF profile broadens and flattens. On the one hand, finite-size effects set in when the tails of the PDF touch the system boundaries. On the other hand, when the typical size of a highly correlated particle domain, \(\approx 2\xi\), becomes comparable to the substrate size, L, finite-size effects dominate, and the PDF saturates to its strong interaction bimodal form.

Discrete equilibrium probability distribution of the number of bound particles, \(P\left( N\right)\), for different values of the (dimensionless) interaction potential: \(J=0\) (orange), 3 (blue), and 5 (green). Histograms are obtained from an exact enumeration using the 1D Hamiltonian from Eq. (1) for a system of size \(L=13\) for the average occupancies \(\langle N \rangle = \langle \varphi \rangle L = 4.33,\,6.5,\) and 8.67 (\(\langle \varphi \rangle = 1/3,\, 1/2,\) and 2/3), respectively.
PDF in the zero-cooperativity limit: the Hill–Langmuir case
In the absence of cooperativity, we expect N to be normally distributed with standard deviation \(L \sigma _0\), provided the system is large enough, and the position of the mean is far enough from the system boundaries: \(P_0\left( N; \langle \varphi \rangle \right) \equiv P\left( N; 0,\langle \varphi \rangle \right) \approx \mathcal {N}\left( \langle N\rangle , L\sigma _0\right)\) for \(L\gg 1\) (strictly speaking, the variable N being discrete, the probability follows a binomial distribution. But the latter is well approximated by the normal distribution in the large L limit). The half-filling case with \(J=0\) in Fig. 4b approximates this Gaussian behavior well because the average value, \(\langle N\rangle\), is far enough from the boundaries that the tails of the distribution do not reach the boundary values, 0 and L (the distribution decays fast enough to the left and right of the average value). More quantitatively, for any value of J, finite-size effects set in when \(\langle N\rangle < 2L \sigma\) or \(L-\langle N\rangle < 2L \sigma\) (i.e., when the tails of the PDF touch the system boundaries).
PDF in the strong cooperativity limit
The PDF broadens and flattens for intermediate positive values of the interaction potential. For higher values, e. g. \(J\ge 5\), the PDF saturates by accumulating at the boundaries at
(here, \(\delta _{ij}\) is the Kronecker delta) and the system becomes well described by an effective two-state (bimodal) system (this is the limit where there exists only one effective block domain). This result is intuitively plausible and can be obtained by retaining in the full partition sum only the empty and full states (see Section “Probability distribution function of the occupancy” for more details). In this limit, finite-size effects are always dominant, and the standard deviation for the occupancy N can easily be calculated directly from the PDF. As expected, it saturates at \(L\sigma _\infty\) (\(=L/2\) at half-filling). Owing to the simple form of this limiting PDF, it is easy to calculate all moments of N, leading to \(\langle N^m \rangle _\infty = L^m \langle \varphi \rangle _\infty\) (or \(\langle \varphi ^m \rangle _\infty = \langle \varphi \rangle _\infty\)) and therefore explicit expressions for low order standardized moments, such as the skewness and kurtosis, see Section “Probability distribution function of the occupancy”. This simplification occurs because, with the correlation length, the energy cost needed to create domain walls also increases with J. Hence, a strongly correlated system favors microscopic configurations that minimize the number of domain walls, selecting the mesostates corresponding to either an empty or a fully occupied substrate. This tendency to select extremal occupancies is a characteristic signature of the presence of strong cooperativity effects in small-size systems. Furthermore, for a parameter choice corresponding to an occupancy expectation value different from half-filling, the distribution is skewed, as in Fig. 4a,c, with an asymmetry that increases with the value of the interaction potential.
PDF in the strong anti-cooperativity limit
The situation is more complicated for strong anti-cooperativity (\(J \ll -1\)) because the system cannot, in general, be reduced to a simple one or two-state system, except near zero and half-filling. Furthermore, near HF, the reduction depends on whether L is even or odd (see Fig. 5). In the following, we will focus on the left half of the plots in Fig. 5, i. e. on occupancies from \(N=0\) to L/2, the right half being a reflection of the left through HF owing to the particle-hole symmetry of the system.
For L even, the PDF has a single peak at \(N=0\) at zero filling and at \(N=L/2\) at half-filling, leading to vanishing standard deviations in these two limiting cases (see Fig. 3b). At HF, there is only one allowed state, consisting of L/2 non-overlapping particle-hole pairs [\((\varphi _i, \varphi _{i+1}) = (1,0)\)] (with a two-fold degeneracy, because the particles can all be on either the even or odd sites).
For L odd, the PDF still has a single peak at \(N=0\) at zero filling, but since no single microstate corresponds to HF, there are in this case two peaks with equal (50%) weight at \(N = (L-1)/2\) and \((L+1)/2\), leading to a non-vanishing standard deviation, \(\sigma _{-\infty }^\textrm{HF} = 1/(2L)\), (see PDF for \(L=13\) in Fig. 5). For L odd, particle-hole pairs cannot cover the whole system, and defects must appear to fulfill the HF constraint, either an extra hole (\(N = (L-1)/2\)) or an extra particle (\(N = (L+1)/2\)).
PDF near half-filling: summary
We can summarize the situation as follows. At (or close to) HF, the four different cases studied give rise to very different results for the fluctuations described by the standard deviation and can be considered as clearly distinguishable signatures of different types of particle-particle correlations in small-size systems. At half-filling, the standard deviation takes on the following values in decreasing order: (i) for strong cooperativity (\(J\rightarrow \infty\)), \(\sigma = 1/2\); (ii) for no cooperativity (\(J=0\)), \(\sigma = 1/(2\sqrt{L})\); (iii) for strong anti-cooperativity (\(J\rightarrow -\infty\)) and L odd, \(\sigma = 1/(2L)\); and (iv) for strong anti-cooperativity (\(J\rightarrow -\infty\)) and L even, \(\sigma = 0\).

Discrete equilibrium probability distribution of the number of bound particles, \(P\left( N\right)\), for \(J=-10\) and three different mean occupancies: \(\langle \varphi \rangle = 1/3,\, 1/2,\) and 2/3, respectively. Histograms show the result of exact enumeration based on the 1D Hamiltonian from Eq. (1) for a system of size \(L=13\) (a) and \(L=14\) (b).
In contradistinction to the strong cooperativity limit where the two-state approximation is valid over the whole range of relative occupancy (from zero to full-filling) for both L even and odd, the strong anti-cooperativity limit is more complicated because the one or two-state approximation is only valid for zero and half-filling. Although more than two occupancy states are involved between these limits, it is clear that strong anti-cooperativity favors the formation of particle-hole pairs. Therefore for even L, the system can be approximated by a system of non-overlapping particle-hole pairs. For L odd, two extra defect states must be included (an extra particle or extra hole). This restricted grand partition function approach, where only the relevant states participating in the strong cooperativity and strong anti-cooperativity limits are retained, is developed in Section “Probability distribution function of the occupancy” of the Methods.
Application to the bacterial flagellar motor (BFM)
The theoretical framework outlined in preceding sections is well adapted for examining (anti)co-operativity-mediated adsorption on substrates with a limited number of binding sites—a fundamental mechanism underlying various critical biological processes. This includes enzyme regulation, chemical sensing, and signal transduction, as well as molecular binding dynamics7,8,39. A prime example where our methodology finds application is in (homo)oligomeric ring structures. These small-sized, ligand-activated protein structures are widespread across biological systems, and their regulatory mechanisms often involve allosteric interactions between binding sites. Our work complements the study presented in40, where it was possible to achieve nearly perfect experimental control and thereby span the whole spectrum of mean occupancies. In contrast, by putting the main emphasis on the analysis of fluctuations, we designed our theoretical approach to work under less controlled experimental conditions, where experimental variables, such as bead sizes in single-molecule experiments, are limited. This is a common scenario in biological research, where perfect control over all variables is often not feasible. Nonetheless, among the diverse suitable applications, we specifically target the stator unit assembly dynamics within the BFM as a primary illustrative application of our theoretical framework.
In what follows, we aim to determine whether cooperativity between BFM stator units, mediated by a (dimensionless) interaction potential J, plays a role in their dynamical assembly at the periphery of the rotor.
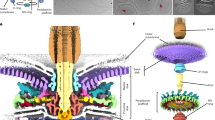
Like most microorganisms, bacteria live in fluid environments with low Reynolds numbers, making them experience a viscous force much larger than the inertial ones41. They have subsequently evolved a variety of compatible motility mechanisms that have been widely studied42,43,44,45,46,47. One type of motility involves rotating one or several flagella that propel bacteria through aqueous media48,49,50,51,52,53,54,55,56. The BFM, whose schematic structure is shown in Fig. 6a, is a transmembrane macromolecular complex that consumes the electrochemical potential across the inner bacterial cell membrane to generate torque and set the flagellum in rotary movement12,13,18,52,57,58. One of the most compelling properties of the BFM is its ability to change both its conformation and stoichiometry depending on the external medium, allowing it to change the direction of rotation and the magnitude of torque produced17,19,59,60. Its mechanisms of adaptation to external stimuli have been widely studied, becoming a model molecular machine to investigate properties such as mechano-sensitivity15,17,20,60,61,62, chemotaxis63,64,65 and dynamic subunit exchange66.

(a) Schematic structure of the flagellar motor, shown in cross section, showing the flagellum (pink), hook (purple), L-P ring (yellow, orange), rod (cyan), MS ring (red), C ring (green), and stators (blue). The inset at the top right shows a top down schematic of the stators assembling at the periphery of the rotor, where \(\mu _\text {r}\) is the chemical potential and J is the nearest-neighbor interaction. (b) The bead assay technique to measure the single motor activity.
Torque is generated by inner membrane ion channel complexes called stator units (depicted in blue in Fig. 6a) which dynamically bind and unbind to the peptidoglycan (cell wall) at the periphery of the rotor. When unbound, they are inactive and passively diffuse in the inner membrane. In their bound state, anchored to the peptidoglycan, the ion channels are activated, and, through a mechanism not yet fully understood, apply torque to the rotor67,68,69,70,71,72,73,74. The inset of Fig. 6a shows a top-down view of the BFM, illustrating the dynamic recruitment of stator units in analogy with the minimal SRLG model described in Section “Lattice gas model”. In this representation, the C ring is modeled as a lattice with a fixed number L of binding sites in contact with a reservoir of stator units, treated as an ideal gas characterized by a chemical potential \(\mu _\text {r}\). Precise measurements of the temporal evolution of the angular velocity of the motor of E. coli, a direct proxy for the number of bound stator units, have shown that the system can recruit up to \(L \approx 13\) stator units15 and that the system is mechanosensitive in that stator unit stoichiometry scales with the external torque induced by the viscous drag acting on the flagellum turning inside an aqueous medium55.
In recent years, observations have indirectly suggested that the stator unit recruitment dynamics of the BFM may be consistent with cooperativity among stator units. The first piece of evidence comes from the occupancy dwell time analysis of the BFM remodeling process in tethered cell experiments14. In this study, the dwell time in the zero occupancy state is considerably longer than all other non-zero occupancy states, which suggests that once a stator unit is bound, the affinity for successive bindings increases. The second piece of evidence comes from experimental observation that the stator unit adsorption rate onto the rotor depends non-monotonically on the number of bound units75. The third line of evidence comes from protein structure studies of the stator units employing cryoelectron microscopy, which led to the proposal of a molecular framework for BFM mechanosensitivity76. This framework is coherent with the catch-bond model introduced by15, and it is also compatible with incorporating cooperativity through an effective long-range interaction mechanically mediated by the rotor. The authors suggest a dual-state mechanism for adsorbed stator units whose switching from an inactive (non-torque-producing) to an active (torque-producing) state is triggered by the forces exerted on the stator units by the movement of the rotor. This framework offers insight into the longer dwell time for the zero occupancy reported by14: the delay can be attributed to the initial unit’s engagement with the rotor and the cell wall in a still-inactive state, lacking the motor torque necessary to activate the unit and produce motor rotation.
Much can be learned about the dynamics of the BFM using bead assay measurements. In our experiments performed on E. coli, we attach a microparticle (bead) to the ‘hook’ (the extracellular portion that joins the motor to the flagellum) (cf. Fig. 6b). We use bacterial strains with genetically deleted cheY. This modification inhibits the production of CheY-P and ensures that the BFM rotates only in the CCW direction, effectively decoupling stator unit dynamics from the CW-CCW switching mechanism. By tracking the bead’s off-axis rotation, we can calculate the angular velocity, \(\omega\), and the torque produced, \(\tau\) (from the relation \(\tau = \gamma \omega\), where \(\gamma\) is the drag coefficient that increases with the bead diameter, yet without obeying a simple power law because of the correction due to rotation near the cell wall77). Both \(\omega\) and \(\tau\) are direct proxies for the number of bound stator units. We can (indirectly) control the mean number of bound stator units at steady state, \(\langle N \rangle\), by varying the bead size and hence the viscous load, because the binding of stators to the BFM is mechanosensitive (see, e.g.,15, and references therein). We can thus measure the temporal evolution of the number of bound stator units on individual motors, as well as the fluctuations around mean occupancy (for more information on the experimental setup, see Section “Experimental data” in the Methods and references15,16).
We describe the mechanosensitive binding and unbinding of stator units in the stationary angular velocity regime of the BFM in terms of our adsorption model [see Eq. (1)], the rotor being a small-size substrate with periodic boundary conditions onto which up to \(L = 13\) stator units can bind at fixed equally spaced positions. Our working hypothesis, in line with previous studies, is that as long as we only focus on the occupancy of the stator units in the (non-equilibrium) motor stationary state this quantity fluctuates around a fixed mean value as if the stator subsystem were effectively at equilibrium. In this picture, we account for the presence of the unbound (inactive) stator units diffusing freely in the inner membrane by imposing an external reservoir chemical potential, \(\mu _\textrm{r}\), which is taken to be constant, as we expect depletion effects to be negligible. We incorporate the mechanosensitivity into the model in an average way by assuming that the stator unit binding energy, \(\varepsilon\), depends on the viscous load (in our case dependent on the size of the bead and the viscosity of the surrounding medium). Load-induced changes in \(\varepsilon\) will naturally lead to load-dependent average occupancies and fluctuations, as observed experimentally. Furthermore, we assume that the interaction parameter J remains fixed (independent of the load) and check a posteriori if this assumption is consistent with the data.
As explained previously, we attempt to use the fluctuations in the average number of bound stator units in the stationary angular velocity regime to determine whether or not cooperativity is at play in the BFM. Figure 7 shows a comparison between the theoretical standard deviation curves, already presented in Fig. 3a, and the experimental standard deviations from five different applied viscous loads, corresponding to three different beads with different diameters that are indicated in the legend (the values of the experimental data are presented in Table 1). One can see that the experimental data fall between the values of \(J = 0.5\) and 2, leading to the conclusion that: (i) a constant (load-independent) cooperativity parameter (J) is a reasonable working hypothesis; (ii) a moderate level of cooperativity in the system, \(J \approx 1.21 \pm 0.22\), obtained by fitting the model to the experimental data, is consistent with the experimental observations, see Fig. 7. The standard error of the nonlinear fit is 0.22 and the coefficient of determination is \(R^2\) = 0.98. Since \(R^2\) is close to its maximum value of 1 and the relative error is less than 20%, we can conclude that there is a good fit of the model to the standard deviation data and therefore that the model is “robust” enough to provide a reliable estimate of J; and (iii) the estimated value of J is coherent with what is expected for typical biological systems exhibiting moderate cooperativity.

Parametric plot of the standard deviation of the relative occupancy, \(\sigma\), versus the mean occupancy, \(\langle \varphi \rangle\), comparing the experimental values with the theory for different values of cooperativity J (\(L=13\)). The experimental data correspond to different applied viscous loads with three different micro-particles (beads) whose diameters are indicated in the legend. The different shapes indicate that the data come from three different types of experiments, corresponding to the steady state stage: (i) after the resurrection of the motor (circles), (ii) after the stall of the motor (squares), and (iii) in the purely steady state (diamonds). Information on the experimental data is detailed in Section “Experimental data” and further elaborated upon in15,16. The dotted black line is the fit to the experimental data of the theoretical standard deviation obtained from Eq. (79). The gray shaded area is the 90% confidence window and the best-fit value is \(J = 1.21 \pm 0.22\), where 0.22 is the standard error of the nonlinear fit.
To delve into further detail, we can also compare the probability distributions of the occupancy shown in Section “Probability distribution function (PDF) of the occupancy and its dependency on \(\langle \varphi \rangle\) and J” with our experimental ones. Figure 8 shows a comparison between the probability distribution from experiments (black dots, see Table 2) and the probability distributions obtained by exact enumeration for different values of J. Even though, in this analysis of our previously published, but limited, experimental data, we don’t observe a precise fit with any of the chosen values, it is clear that the PDFs for the range of J estimated from the standard deviation are coherent with our experimental results. Our results suggest that the interaction potential has a non-zero positive value qualitatively in accordance with the value range deduced from the above fluctuation analysis. (We refer to Fig. 18 in the Methods for a direct comparison of the experimental PDFs and the theoretical PDFs calculated using the best-fit value of the dimensionless interaction potential, \(J=1.21\).) This qualitative agreement between the evolution of the experimental PDFs with increasing average relative occupancy and the predictions of the underlying lattice gas model with moderate cooperativity therefore lends credence to our choice of minimalist adsorption model. Moreover, as further explained in the Methods section, we observe theoretically that the system becomes bimodal (with only the completely full and empty states having significant probabilities) for slightly higher values of the interaction potential, \(J \gtrsim 5\), behavior that is not seen in the experimental PDF data. We see from Section “Probability distribution function (PDF) of the occupancy and its dependency on \(\langle \varphi \rangle\) and J” (Fig. 18c) that for the chosen system size (\(L=13\)), \(J\approx 3\) is a threshold value marking a transition from low cooperativity PDFs having a maximum centered on the average occupancy to high cooperativity PDFs exhibiting a bimodal form with local maxima at zero and full filling. In the latter case, microstates with fillings close to the average occupancy have low probabilities, and the motor would undergo discontinuous jerky motion. Such a bacteria would fluctuate between an immobile state (zero stator unit occupancy) and maximum speed (full stator unit occupancy). In the absence of cheY the BFM rotates only in the CCW direction and we expect the stator unit dynamics (now decoupled from the CW-CCW switching mechanism) to be appropriate for a highly and smoothly adaptive molecular machine. High values of stator cooperativity would, therefore, not be expected since such a motor would not be able to adapt smoothly to environmental variations.

Discrete equilibrium probability distribution of bound stator units, P(N), for the experiments (black dots with error bars corresponding to one standard deviation of uncertainty) and different values of the interaction potential, \(J = 0\) (blue), 1 (orange), 2 (green), obtained from exact enumeration of states from the effective Hamiltonian; here, \(L=13\). The mean equilibrium occupancy of each distribution is indicated on top of the three plots, which corresponds to a given external load imposed by beads of different diameters indicated in the legend.
Discussion and Conclusion
Theoretical insights into cooperative ligand binding on small substrates
We have focused on cooperative processes involving the adsorption of ligands onto a substrate disposing of a limited number of binding sites. By presenting a general method (based on a minimalist 1D periodic lattice gas) for recognizing and assessing characteristic signatures of cooperativity or anti-cooperativity in the stochastic occupancy fluctuations, we propose both a criterion to determine whether any given adsorption system exhibits cooperative or anti-cooperative behavior and a method to quantify the amplitude of the ligand-ligand interaction potential.
Despite their derivation from a minimal model, the core insights of our study—namely, the functional dependence of fluctuations on cooperativity, the trade-off between adaptability and fluctuation minimization, and the intricate relationship between system size, fluctuation amplitude, and the extractable information within a given experimental resolution—serve as universal principles that extend to more refined models. In lattice gas models, however, for which most results are based on the Ising model and designed for large systems where the thermodynamic limit applies, results often become complex when it comes to finite lattices and, even for the most stripped down model—such as the 1D SRLG presented in Section “Lattice gas model”—lack simple intuitive interpretation. Therefore, we have made a conscious effort to present our theoretical findings for finite size systems in a physically transparent manner.
First, in Section “Mean occupancy and its standard deviation”, we analyzed how the mean relative occupancy in equilibrium, \(\langle \varphi \rangle\), and its fluctuations, \(\sigma\), taken to be the relevant experimentally accessible quantities, depend on the model parameters: the (dimensionless) effective chemical and the interaction potential, \(\mu\) and J, respectively, as well as the system size L. We have used our analytical results for \(\langle \varphi \rangle\) and \(\sigma\) to draw parameter space phase diagrams illustrating different types of system behaviour depending on the values of \(\mu\) and J. Second, to deepen our physical understanding of these quantities Section “Fluctuations of the mean occupancy and their dependency on \(\mathinner {\langle {\varphi }\rangle }\) and J” presents analytical asymptotic expressions for different scenarios, including strong cooperativity, zero cooperativity (Hill–Langmuir), and strong anti-cooperativity. Based on these results, we elucidated the parametric relationship between the standard deviation and expected lattice occupancy for a wide range of model parameters (\(\mu\) and J) and various lattice sizes L. This relationship is the key to estimating the interaction potential by fitting the model to the experimental data (cf. Fig. 7). Additionally, the difference in fluctuation signatures for lattices with L even and odd shown in Fig. 3 highlights the effects of anti-cooperativity and illustrates how these effects diminish as system size increases (cf. Fig. 19). In Section “Probability distribution function (PDF) of the occupancy and its dependency on \(\langle \varphi \rangle\) and J”, we reconstructed the occupancy probability distribution through exact enumeration of the microstates, focusing mainly on how the finite size of the lattice affects the distribution’s shape; moreover, we refer the reader to Section “Probability distribution function of the occupancy” for an extensive analysis of the approximated PDFs. From these insights, we demonstrated that the variance of a finite-size system can be explicitly expressed by Eq. (30) using the correlation length \(\xi\) and the expected occupancy \(\langle \varphi \rangle _{\text {TL}}\) in the thermodynamic limit, thus clearly linking behaviors observed in finite and infinite systems. Finally, the insights gained by comparing the SRLG with the IRLG, detailed in Section “Infinite range (IR) model”, enabled us to derive the interpolation formula Eq. (79), designed to bridge the gap between theoretical predictions and practical observations.
In the process, knowing that in the thermodynamic limit relative occupancy fluctuations (or standard deviation) vanish, we have addressed the following essential questions: (i) what is a sufficiently “small” system for studying fluctuations? (ii) what model parameter values allow a system to smoothly “adapt” to external conditions?
We address the first question in detail in Section “System size, fluctuations, and experimental precision” of the Methods, by proposing a criterion for how small a system must be for the relative fluctuations to be experimentally detectable. There, we propose an operational definition: we consider a system “small” if the lines of the contour plots (\(\sigma\) against \(\langle \varphi \rangle\)) are “spaced apart enough” compared to the experimental error so that a reliable estimation of J is possible (see in particular Fig. 11).
In what follows, we address the second question and discuss its implication for BFM stator unit dynamics.
Stator unit occupancy of the BFM: moderate cooperativity allows for high adaptability by keeping fluctuations low
We compared the theoretical results for both the standard deviation and the probability distribution function of stator unit occupancy with experimental data obtained for the BFM. We concluded that a moderate value of cooperativity, \(\approx 1 \,k_{\text {B}}T\) (i.e., \(J \approx 1\) with a standard error of about \(20\%\)), for the short-range (SR) model is not only coherent with the experimental data but also expected from the characteristic smooth adaptability of the motor to changing external loads [for the infinite-range (IR) model, using the mapping (73) derived in Section “Infinite range (IR) model”, the corresponding value is \(J_\textrm{IR} = 2 J_\textrm{SR}/(L-1) \approx 0.20\)]. For the characteristic size of the BFM (\(\approx 10\) binding sites), slightly higher values of cooperativity would lead to a motor that is bimodal in the occupancy and, therefore, would exhibit switch-like behavior for the produced torque not compatible with the required motor characteristics. The highly cooperative bimodal behavior found at strong coupling does, however, appear to describe the rapid stochastic switching in BFM rotational motion17,23,31, a mechanism that plays an important role in the run-and-tumble strategy adapted to the chemical sensing essential in bacterial chemotaxis (suppressed in our experiments by using bacterial strains with a cheY deletion).
Within the framework that we have developed, we are now in a position to propose, using the BFM as an illustrative example, a general principle of motor adaptability depending on whether the motor under investigation should respond smoothly to external stimuli or behave like a two-state switch. As stated earlier, we suppose that for the BFM the binding energy \(\varepsilon\) depends on the load (and therefore the bead size). A change in load would therefore lead directly to a change in the effective chemical potential \(\mu\) and therefore a modification of the average occupancy and standard deviation (fluctuations).
For the short-range model, these modifications to \(\mu\) (extracted from the experimental data, see Tables 3 and 4) are shown in Fig. 9 on the occupancy and standard deviation contour plots for each of the three studied values of J (for \(L=13\)). By positioning the effective chemical potential \(\mu\) window in this way we observe that for \(1< J < 2\) it is situated in a sweet spot suitable for a motor that responds smoothly to environmental changes with the ability to cover a wide range of occupancies spanning half-filling while minimizing the amplitude of the fluctuations.
For the BFM the occupancy is directly related to the motor speed and therefore, near this sweet spot, the BFM can smoothly adjust its speed in response to external stimuli with a minimum of fluctuations. In retrospect the bead sizes used in the experiments were clearly chosen to see an effect because a much higher load would have forced the system into nearly full filling (exactly the case in the stall experiments reported in16); a much lighter load would have pushed the system to nearly zero filling (similar to what was done using another technique in the resurrection experiments). The highly cooperative switch-like behavior (such as observed for the BFM rotational direction changes17,31, suppressed here) would place this system in the strong coupling regime on the right-hand side of the contour plots (\(J>4\)).
For the motor to respond smoothly to external stimuli and cover a wide range of occupancy with minimum of fluctuations we therefore see that moderate positive values of cooperativity \(1< J < 2\) are optimal given the \(\mu\) window imposed by the system characteristics. Although the occupancy range would increase at higher values of J, this positive effect would be counterbalanced by a strong increase in fluctuations because the system would be pushed into the switch-like operation regime. On the other hand, for lower and even negative values of J, the fluctuations would (favorably) be diminished in amplitude, but at the cost of severely restricting the accessible range of occupancy (and therefore motor speed for the BFM).
By examining Fig. 9, we observe that if a two-state switch-like motor operation were sought after in order to cover a wider range of average relative occupancy, then higher values of J would be the best choice; the price to pay would be a strong increase in fluctuations, provided that the observation time window be wide enough.

Values of the effective chemical potential \(\mu\) extracted from the experimental data for different values of the interaction potential \(J=1,2,3\) (\(L=13\)). We situate them on the contour plots of \(\langle \varphi \rangle\) and \(\sigma\) in order to find the interval, or window, of chemical potential for which the BFM would work at fixed J. Experimental values for J and \(\mu\) are provided in Table 3 for the mean relative occupancy and in Table 4 for the standard deviation.
If a motor with weak fluctuations and a restricted range of occupancies were sought after then negative values of J (anti-cooperativity) would be the best choice. In fact, this type of motor would be relatively immune to changes in external stimuli and therefore exhibit a relatively constant speed (provided the speed were still directly related to occupancy, as for the BFM).
More generally, our work contributes to a deeper understanding of how biological systems optimize their functioning depending on system requirements and how each system deals with the trade-off between an increase in signal gain from cooperativity and the concomitant increase in fluctuations. Despite the loss in signal-to-noise ratio in the presence of cooperativity24,25,26,27, there are chemical sensing systems that benefit from cooperativity in order to optimize other system requirements. In the case of stator binding to the BFM, we propose that the presence of moderate cooperativity permits the system to access a wider occupancy window, without paying a too high price in enhanced fluctuations. In the case of bacterial chemotaxis, Skoge et al. suggest that E. coli bacteria optimize their drift velocity arising from run-and-tumble motion at the expense of the sensory detection signal-to-noise ratio. Other biological systems, such as the eukaryotic chemotaxis one, seem to favor the optimized signal-to-noise ratio coming from the absence of chemical receptor cooperativity.
We have not addressed here the biochemical origins of stator-stator interactions, although one can imagine that stator units interact at short range much like proteins, either directly or through allosteric pathways, and that long-range interactions could also be due to allosteric effects. We plan to address this and other open questions in future work. In the present context the major important open question in modeling the BFM concerns how to integrate cooperativity into kinetic models that already allow one to account for the relaxation time asymmetry between stall and resurrection16. The goal is to arrive at a model that can describe in a unified way both BFM stator number fluctuations and relaxation time asymmetries.
Methods
1D short-range lattice gas (SRLG)
Analytical solutions for the equilibrium state can be obtained using the transfer matrix formalism32,33,78. According to it, the grand partition function of the system can be written as:
where T is the transfer matrix, which for the system we are working on, is given by
In the case of periodic boundary conditions, Eq. (20) can be simplified to
where
are the eigenvalues of the transfer matrix and \(X = \frac{1}{2}(J + \mu )\). From this result (22) for the grand partition function, one can obtain the mean relative occupancy, \(\langle \varphi \rangle\), and the mean square relative occupancy, \(\langle \varphi ^2\rangle\), as functions of \(\mu\) and J by employing known relations between thermodynamic quantities (i.e., by taking derivatives of \(\Xi\) with respect to the effective chemical potential \(\mu\)). The standard deviation \(\sigma\) can then be derived as a function of \(\mu\) and J from these two averages.
The transfer matrix formalism can also be used to calculate the occupancy-occupancy correlation function, which provides another route to \(\langle \varphi ^2\rangle\) (and therefore \(\sigma\)), but now as an explicit function of \(\langle \varphi \rangle\) and the correlation length, \(\xi \equiv 1/\ln \left( \lambda _+/\lambda _-\right)\) [already presented in Eq. (3)]. This second form for \(\sigma\) lends itself to a clearer physical picture of how the standard deviation interpolates between the zero and infinite cooperativity limits.
According to the grand canonical formalism, the mean relative occupancy of a system is given by the expression
where \(\Omega = -L\ln {\Xi }\) is the grand potential corresponding to the grand partition function.
Using the chain rule, we can find that
Replacing
in the previous equation, we obtain Eq. (2).
Similarly, we can determine the mean square relative occupancy by taking another derivative:
By carrying out this calculation, we obtain:
Denoting \(\xi\) as the correlation length makes physical sense because correlations decay exponentially on a scale \(\xi\), as can be seen by inspecting the correlation function obtained using the transfer matrix formalism:
where \(v_- = \text {e}^{\mu /2}(\lambda _- - 1)\) and \(K =(\lambda _+^L + \lambda _-^L)(1 + v_-^2)^2\). By using the definition of \(\xi\) presented above, the products of \(\lambda _+^r \lambda _-^{-r}\) and \(\lambda _+^{-r}\lambda _-^r\) can be rewritten as \(\text {e}^{\pm r/\xi }\). This result for the correlation function, Eq. (29), can be put into a more physically transparent form,
This result, which illustrates how the correlation function can be written explicitly in terms of \(\langle \varphi \rangle\), \(\xi\), and L, allows the exponential decay on a scale \(\xi\) to be made clear. Here \(\langle \varphi \rangle _\textrm{TL}\) is the average relative occupancy in the thermodynamic limit (TL), defined to be an explicit function of \(\langle \varphi \rangle\) and \(\xi\):
This result for the correlation function, Eq. (30), is clearly periodic and tends to the correct TL when \(L\rightarrow \infty\):
By performing the sum depicted in Eq. (16) using Eq. (29) we obtain for the mean square relative occupancy an expression that explicitly depends on the eigenvalues and the correlation length as follows:
It can be shown that Eq. (33) is equivalent to Eq. (28), which provides a check on the thermodynamic result. In the calculation leading to Eq. (28), the correlation length appears from the fraction \((\lambda _+^L - \lambda _-^L)/(\lambda _+^L + \lambda _-^L)\) which can be rewritten as \(\tanh (L/(2\xi ))\).
With both Eqs. (2) and (28), we can obtain an analytical expression for the standard deviation from the expression \(\sigma = \sqrt{\langle \varphi ^2 \rangle - \langle \varphi \rangle ^2}\). The result obtained in this way is equivalent to the one obtained more efficiently in the main text using the thermodynamic relation \(\sigma = L^{-1/2}\sqrt{\partial _\mu \langle \varphi \rangle }\).
To make explicit the \(\xi\) dependence we now calculate \(\sigma\) using the connected correlation function, given by \(\langle \varphi _i \varphi _j \rangle - \langle \varphi \rangle ^2\), in the double sum leading to \(\sigma ^2\):
The sum in Eq. (35) can be evaluated using Eq. (30). to obtain a physically transparent expression for the variance, \(\sigma ^2\), explicitly in terms of \(\langle \varphi \rangle\), \(\xi\), and L:
where \(\langle \varphi \rangle _\textrm{TL}\) is given by Eq. (31) and
This form for the variance allows the weak and strong cooperativity limits to be clearly identified, since \(F \rightarrow 0\), when \(J\rightarrow \infty\) and \(F \rightarrow -1 + 1/L\), when \(J\rightarrow 0\) (in the latter limit, \(\langle \varphi \rangle _\textrm{TL} \rightarrow \langle \varphi \rangle\)). More generally, for fixed J, \(\xi\) becomes an implicit function of \(\langle \varphi \rangle\) and can be plotted parametrically, as in Fig. 17 for infinite J.
The thermodynamic limit corresponds to \(L \rightarrow \infty\), for which \(\tanh {(L/2\xi )} \rightarrow 1\), therefore the average relative occupancy simplifies to
and \(\langle \varphi ^2 \rangle _\textrm{TL} = \langle \varphi \rangle ^2_\textrm{TL}\). Hence the standard deviation in the thermodynamic limit, \(\sigma _\textrm{TL}\), is zero and self-averaging holds.
Probability distribution function of the occupancy
For high positive values of J, e. g. \(J\ge 5\), the PDF saturates by accumulating at the boundaries to
and the system becomes well described by an effective two-state system. The standard deviation for the occupancy N therefore saturates at \(L\sigma _\infty\) (\(=L/2\) at half-filling), where we recall that \(\sigma _\infty = \sqrt{\langle \varphi \rangle - \langle \varphi \rangle ^2}\), and it becomes easy to calculate all moments of N: \(\langle N^m \rangle = L^m \langle \varphi \rangle\) (or \(\langle \varphi ^m \rangle = \langle \varphi \rangle\)). The moments are defined by
and the standard moments by \(\mu _n/\sigma ^n\), where \(\sigma = \sqrt{\mu _2}\) is the standard deviation. In the strong cooperativity limit, one can therefore find simple explicit expressions for low order standard moments, such as the skewness \(\gamma = \mu _{3}/\sigma ^3\),
and kurtosis, \(\kappa = \mu _{4}/\sigma ^4\),
These results for the effective two-state system are very different from those predicted for a Gaussian PDF.
For strong anti-cooperativity (\(J \ll -1\)), the system near zero and half-filling can be reduced to a simple one or two-state system. Near HF, the reduction depends on whether L is even or odd (see Fig. 5). The PDF always has a single peak at \(N=0\) at zero filling. For even L in the strong anti-cooperativity limit, the PDF also has a single peak at \(N=L/2\) at half-filling,
(see Fig. 5) leading to a vanishing standard deviation. For L odd at half-filling, on the other hand, there are two peaks with equal (50%) weight at \(N = (L-1)/2\) and \((L+1)/2\),
leading to a non-vanishing standard deviation of 1/2 for the occupancy N. Unlike for L even, for L odd non-overlapping particle-hole pairs cannot cover the whole system, and a defect (non-particle-hole pair) must appear, either an extra hole [\(N = (L-1)/2\)] or an extra particle [\(N = (L+1)/2\)], to arrive at HF.
The restricted grand partition function approach consists in keeping in the sum over all possible microstates \(\{ \varphi _i \}\) only those states that survive in the studied limit. Before presenting this approach, we first recall the exact calculation in the absence of cooperativity (\(J=0\)).
For \(J=0\) the partition sum can be organized into a sum over states with a fixed occupancy N with the Boltzmann factor \(\exp [-\beta \mathcal {H}_0(N)]=\text {e}^{\mu N}\) multiplied by a multiplicity (or binomial coefficient) \(C^L_N = L! /\left[ N! \left( L-N\right) !\right]\) (related to the configurational entropy) that gives the number of microstates \(\Phi\) consistent with occupancy N:
From \(\Xi _0\) we immediately obtain the expected \(J=0\) results:
and
By introducing the usual (configurational) entropy of mixing,
the multiplicity \(C^L_N\) can be accurately approximated (using Stirling’s formula) for \(L \gg 1\) and \(0< \varphi = N/L < 1\) by
where
is the dimensionless entropy of mixing. The entropy of mixing is a non-monotonic (concave) function of \(\varphi\) that goes to 0 for zero and full filling and reaches a maximum at HF.
In Eq. (49) for \(L \gg 1\), the main variation comes from the exponential, and it is possible under certain conditions to replace \(\varphi\) in the prefactor by \(\langle \varphi \rangle\) without loss of accuracy. The PDF for \(J=0\) can then be accurately approximated by
where
is an effective free energy for \(\varphi\) and we have written
and
as functions of \(\langle \varphi \rangle\).
For \(L \gg 1\), we can use the saddle point approximation to obtain a simple Gaussian form for \(P_0(N; \langle \varphi \rangle )\): by expanding \(\beta \mathcal {F}_0 (\varphi ; \langle \varphi \rangle )\) around \(\varphi = \langle \varphi \rangle\) (the position of its minimum) to second order in \((\varphi - \langle \varphi \rangle )\) we find
where the variance
is related to the second derivative of the entropy of mixing, or curvature, evaluated at \(\langle \varphi \rangle\). Since \(-s''_{\textrm{mix}}(\langle \varphi \rangle )\) has a minimum at half-filling, \(\sigma _0\) has a maximum there.
The Gaussian approximation for \(P_0(N; \langle \varphi \rangle )\) is (approximately) correctly normalized because the sum of the exact PDF over N is equal to 1, which is equivalent to the Gaussian approximation times L integrated over \(\varphi\) from \(-\infty\) to \(+\infty\) about its maximum value at \(\langle \varphi \rangle\). The Gaussian approximation is itself clearly only valid when the tails of the distribution are far enough from the extremal values (\(\varphi = 0\) and 1), which is the case for the three \(J=0\) PDFs displayed in Fig. 4 (for \(\langle \varphi \rangle = 0.31,\,0.5,\) and 0.62). For these three cases the Gaussian approximation is extremely accurate. More generally a PDF calculated using the full approximation for \(C_N^L\), Eq. (49), should be accurate over the whole range of \(\varphi\) except close to the extremal values.
In the strong cooperativity limit (\(J \rightarrow +\infty\)), only the empty and full states need be retained in the restricted grand partition function, \(\Xi _{+\infty }\), because any state with \(0< N < L\) will necessarily have domain walls and these states will be suppressed via the Boltzmann factor by \(\text {e}^{-J}\) (with respect to the fully occupied state) for each lost nearest-neighbor interaction. This leads to
where \(X = (J+\mu )/2\) is considered to be kept fixed as \(J\rightarrow + \infty\) and \(\mu \rightarrow -\infty\). From \(\Xi _{+\infty } (X;\mu )\) we obtain
which allows us to read off the expected results,
and
directly from \(\Xi _{+\infty }\) [within this approximation the PDF \(P_{+\infty } (N; \langle \varphi \rangle\)) vanishes for all other values of N].
For strong anti-cooperativity (\(J \ll -1\)), we must treat the L odd and even cases differently. In both these cases, the statistical physics is very different from the strong cooperativity (\(J \gg 1\)) limit where only two states need to be retained.
We start with the simpler L even case for which the system can be approximated by non-overlapping, and therefore non-interacting particle-hole pairs (we choose the convention where the position of the particle-hole pair is determined by the position of the particle making up the particle-hole pair). The restricted grand partition function for \(0 \le \langle \varphi \rangle \le 1/2\) in this particle-hole pair approximation is
where the upper limit on the sum of L/2 reflects the maximum number of non-overlapping pairs on a lattice of size L (even) and the multiplicity \(D^{L}_{N_d}\) counts the number of distinct ways of putting \(N_d\) pairs on a periodic lattice of size L. Since calculating \(D^{L}_{N_d}\) is a non-trivial combinatorial problem, we simplify matters by considering two disjoint lattices of size L/2 consisting of the odd sites for the first and the even sites for the second. If we neglect mixed particle-hole pair microstates, i.e., those that have pairs on both lattices, we obtain the following approximate restricted particle-hole pair grand partition function
where \(C^{L/2}_{N_d}\) is the usual binomial coefficient that counts the number of distinct ways of placing \(N_d\) particles on a lattice of size L/2. In passing from Eqs. (61) to (62), we have replaced \(D^{L}_{N_d}\) by \(2 C^{L/2}_{N_d}\) and therefore simplified the problem to that of two independent non-interacting quasi-particle (i.e., particle-hole pair) systems [cf. Eq. (45))]. Following the above discussion for the \(J=0\) case, we can immediately conclude that \(\langle \varphi \rangle =\langle N_d/L \rangle \approx \langle \varphi \rangle _d^\textrm{even}\), where
and therefore \(\sigma _{-\infty }^\textrm{even} \approx \sigma _d^\textrm{even}\), where
The replacement of \(D^{L}_{N_d}\) by \(2 C^{L/2}_{N_d}\) is exact for the one particle-hole pair state (\(N_d =1\)) and at half-filling, (\(N_d = L/2\)), two cases for which there are no allowed mixed-states (in the latter case because of the high filling of pairs). Although we over-count the (unique) empty state by a factor of 2 (in order to put \(\Xi _{d}^\textrm{even}\) in the free particle form) and under-count the number of particle-hole pair states for \(1< N_d < L/2\), for which there are allowed mixed-states, the physics captured by the approximation \(\sigma _{d}^\textrm{even}\) is sufficiently faithful to that of the original system to be able to account well for the main quantity of interest, namely the standard deviation in the strong anti-cooperativity limit for L even (see Fig. 10).

Comparison between the approximation for the very strong anticooperativity regime \(\sigma _d^\textrm{even}\) and the exact result given by the parametric plot with \(\langle \varphi \rangle\) and \(\sigma\) (see even case in Fig. 3b) for \(J = -10\).
Inspection of Figs. 3b and 10 shows that the approximate form \(\sigma _d^\textrm{even}\) accounts well for the exact behavior, going to 0 at zero and HF and reaching a maximum at 1/4 filling, not far from the exact maximum position. Because of the approximations inherent in obtaining \(\sigma _d^\textrm{even}\), however, the approximate result cannot capture the weak lack of symmetry about 1/4 filling, i.e., the observed skewness towards lower fillings seen in Figs. 3b and 10. This skewness can be seen even more clearly in the PDFs for \(L=14\) and \(\langle \varphi \rangle = 1/3\) presented in Fig. 5. It arises because for \(1< N_d < L/2\) the approximation \(2 C^{L/2}_{N_d}\) underestimates the exact particle-hole pair multiplicity \(D^{L}_{N_d}\) more severely for lower fillings than for higher ones (below HF). The reason being that at higher fillings it becomes more and more difficult to have allowed mixed states.
The roughly Gaussian shape for the PDF with \(L=14\) presented in Fig. 5 for \(\langle \varphi \rangle = 1/3\) can be understood using the approximate particle-hole pair approach that maps the problem onto free quasi-particles (particle-hole pairs). In this case we can adapt the results for the \(J=0\) case to estimate the particle-hole pair PDF for \(J \rightarrow -\infty\) and L even,
using the Gaussian approximation:
where we have used
and
The Gaussian PDF approximation is very accurate compared with \(P_d^\textrm{even}(N; \langle \varphi \rangle )\) for \(L=14\) and \(\langle \varphi \rangle = 1/3\), although \(P_d^\textrm{even}(N; \langle \varphi \rangle )\) itself underestimates the exact maximum by about 30% and cannot capture the skewness observed in the exact results (see Fig. 5). The particle-hole pair approximation does reproduce accurately, however, the width of the exact PDF.
In the strong anti-cooperativity limit (\(J \rightarrow - \infty\)) for L odd, the system cannot be approximated by non-overlapping particle-hole pairs at HF. We need to account for a defect in order to approach HF. The restricted grand partition function for \(0 \le \langle \varphi \rangle \le 1/2\) in this particle-hole pair approximation is
where \((L-1)/2\) is the maximum number of non-overlapping pairs on a lattice of odd size L and the multiplicity \(O^{L}_{N_d}\) counts the number of distinct ways of putting \(N_d\) pairs on a periodic lattice of odd size L. The last two terms are the defect contributions, an extra hole or particle not making up a particle-hole pair that costs an energy \(-J\) for an extra particle and that leads to a multiplicity L counting the number of ways to place an extra hole or particle on a lattice made up of \((L-1)/2\) pairs in sequence.
Since calculating \(O^{L}_{N_d}\) is a non-trivial combinatorial problem, we follow a procedure similar to the one used in the even case and simplify the problem by considering two disjoint lattices of size \((L+1)/2\). If we neglect mixed particle-hole pair microstates for \(0 \le N_d \le (L-3)/2\), i.e., those that have pairs on both lattices, we obtain the following approximate restricted particle-hole pair grand partition function
which can be evaluated by completing the binomial sum and then subtracting the added terms:
where \(C^{(L+1)/2}_{N_d}\) counts the number of distinct ways of placing \(N_d\) particles on a lattice of size \((L+1)/2\). \(\Xi _{d}^\textrm{odd}\) correctly counts the multiplicity of the \((L\pm 1)/2\) states by construction. Although this approximation overcounts the one particle-hole pair state by 1 (\(L+1\) instead of L) and overcounts the empty state by a factor of 2, we believe that it captures the essential physics of the strong anti-cooperativity case for L odd. The above approximation, \(\Xi _{d}^\textrm{odd}\), can be used to calculate \(\langle \varphi \rangle _{d}^\textrm{odd} = (L \Xi _{d}^\textrm{odd})^{-1}\partial _\mu \Xi _{d}^\textrm{odd}\) and \(\sigma _{d}^\textrm{odd} = \sqrt{L^{-1} \partial _\mu \langle \varphi \rangle _{d}^\textrm{odd}}\), an approximate results that could then be compared with the exact ones. We will not, however, pursue this approach any further here.
Infinite range (IR) model
In what follows, we will use the infinite-range (IR) as a proxy for the more general long-range (LR) behavior. As detailed below, the IR results are qualitatively equivalent to those of the SR upon the mapping between the two models proposed in Eq. (73), which is exact to the lowest order in the coupling constant (dimensionless interaction potential). Similarly, the coupling constant in the LR, \(J_\textrm{LR}\), which would explicitly depend on the effective range of the interactions, would always be bound by the SR and IR couplings: \(J_\textrm{SR}< J_\textrm{LR} < J_\textrm{IR}\). Therefore, we can confidently state that we do not expect the LR case to show qualitatively different behavior aside from interpolating between the two other limiting cases.
The mapping between the SR and IR Lattice Gas can be established by performing a perturbative cumulant expansion of the grand partition function:
where the last line gives the first order cumulant expansion. The subscript 0 indicates a statistical average with respect to the non-interacting (Hill–Langmuir) model (ideal Lattice Gas). By calculating \(\langle H_\textrm{int}\rangle _0\), we obtain \(\langle H_\textrm{int}\rangle _0 = -J_\textrm{SR} L \langle \varphi \rangle _0^2\) for the SR model and \(-\frac{1}{2}J_\textrm{IR} L (L-1) \langle \varphi \rangle _0^2\) for the IR model, results that suggest the following mapping between the two models:
This mapping is exact to lowest order in the coupling constant.
We adopt the following strategy to avoid carrying out an explicit calculation of the standard deviation \(\sigma\) directly within the cumulant approximation, which is cumbersome. The weak coupling results for the SR model can be found by expanding the exact parametric results for \(\langle \varphi \rangle (\mu )\) and the variance \(v(\mu ) = \sigma ^2(\mu )\) in powers of J. To first order in \(J_\textrm{SR}\) we find
We can then use the above mapping between the SR and LR models to get a result that is exact to first order in \(J_\textrm{IR}\) for the IR model:
One can check the above method for SR model at half-filling:
It would be useful to find a good interpolation formula for \(\sigma\) as an explicit function of L, J, and \(\langle \varphi \rangle\) for both the SR and IR models. A reasonably good interpolation scheme can be set up by recognizing that the exact result for the standard deviation at half-filling for small J is to a very good approximation,
(where \(\sigma _\infty ^2 = 1/4\) at HF), which shows that it’s best to use an exponential resummation of the small J perturbation expansion.
Comparing the exact SR results with the above approximation shows that an even better approximation is to take the 5/2 root mean (a heuristic choice),
to get a smooth interpolation. The above 5/2 root mean square average leads to our best approximation for \(\sigma _\textrm{SR}\):
This is a convenient form (see Fig. 20) because the dependence on \(\langle \varphi \rangle\) (through \(\sigma _\infty\)), \(J_\textrm{SR}\), and L is made explicit (since \(\sigma _0=L^{-1/2} \sigma _\infty\)). The crossover to the large J saturation limit is centered on
\(J_\textrm{co}\) depends on L and filling \(\langle \varphi \rangle\), increasing slowly with L and reaching a minimum at half-filling for fixed L. The convergence of \(\sigma\) as a function of \(\langle \varphi \rangle\) to the large J saturation limit is uniform only for \(J < J_\textrm{co}\).
The approximation (79) is very accurate for all fillings (except possibly very close to zero and full-filling) as long as \(L^{-5/2} \ll 1\) (the case for \(L \approx 10\)) and \(J < J_\textrm{co}\) (see Fig. 20). For \(L^{-5/2} \ll 1\) and \(J > J_\textrm{co}\), this approximation is accurate only within a window centered on half-filling. Because of the factor \(\sigma _\infty ^2\) in the argument of the exponential, convergence to \(\sigma _\infty\) is fastest at HF and becomes slower and slower as moves away from HF to zero and full filling. For \(L \approx 10\), the window of accuracy for \(J<2\) is \(0.2< \langle \varphi \rangle < 0.8\), which encompasses the experimental data window.
The interpretation of the above results is in accordance with our previous understanding: as long as the system is far enough from zero and full-filling, increasing J leads to a flattening of the Gaussian PDF with \(\sigma\) growing exponentially with J until the system reaches the crossover region, \(J \approx J_\textrm{co}\), before saturating at \(\sigma _\infty\) for \(J \gg J_\textrm{co}\). When the system reaches the crossover region the Gaussian form is no longer accurate because one (or two) of the wings starts to touch one (or two) of the boundaries, before morphing into a bimodal (two-state) system.
The corresponding result for the IR case can be obtained using the mapping (73). We note in passing that an approximate Gaussian effective free energy approach can be developed for the IR model along the lines of the one developed above for the non-interacting (Hill–Langmuir) model. This result can be used to find an extended Gaussian approximation for the IR model,
where the variance \(\sigma _\textrm{IR}\) can be approximated using the approximation for the SR model (79) and the mapping (73).
System size, fluctuations, and experimental precision
By proposing a pragmatic operational definition, we clarify here what we mean by a small system. Before doing so we note that if we were to attempt to qualify the size of an adsorption system by the ratio between the number of binding sites, L, and the size of a correlated domain (twice the correlation length), we would only be characterizing the Hill–Langmuir nature of the fluctuations, occurring when \(2\xi /L \ll 1\), or the bimodal nature of the fluctuations, occurring when \(2\xi /L \gg 1\), and not the feasibility of extracting the cooperativity from the experimentally measured relative occupancy fluctuations at equilibrium.
We propose here that a system be considered a small cooperative one if fluctuations, as quantified by the standard deviation of relative occupancy, are within experimental resolution and can be used to extract, using the theoretical framework that we have developed, the amplitude of the interaction strength, J, and that this strength be within the usual biophysical range, \(0 \le J \le J_\textrm{max} \approx 10\), for adsorption systems.
To quantify our capacity to resolve J theoretically, we therefore propose the following criterion: the derivative of \(\sigma\) with respect to J, \(\sigma _\text {HF}'(J;L)\), must be larger than a given value \(\Delta \sigma\), whose choice depends on the measurement precision, over a J range starting at 0. In what follows, for the application to the BFM, we shall choose \(\Delta \sigma = 0.02\), which is compatible with the experimental error for the data provided in Section “Application to the bacterial flagellar motor (BFM)”.
This choice is motivated by comparing the parametric plots in Fig. 3 to the experimental data (see Fig. 7). One can estimate the coupling J from the data when these fall in regions of the parametric plots where the lines corresponding to different values of J are spaced far enough apart compared to the experimental error. The highest resolution for positive values of J, and fixed \(L\ge 3\), is achieved at half-filling (HF), and for a value \(J^* \approx 2 \ln (L/2)\) that maximizes the derivative of the standard deviation. These maxima correspond to the peaks of the curves in Fig. 11a where we plot \(\sigma _\text {HF}'(J;L)\) as a function of J for different but fixed values of L and compare it with the threshold value \(\Delta \sigma =0.02\).
We observe that for the choice of \(\Delta \sigma =0.02\), \(L^* \approx 39\) is the maximum system size for which there is a feasibility window (\(0< J < 8\)) starting at 0 for extracting the cooperativity from the experimental data. This threshold value \(L^*\) can be recovered analytically by using the approximation Eq. (79) for the standard deviation, which is highly accurate near half-filling provided \(L \gg 1\), to find \(\sigma _\text {HF}'(J=0; L) \approx 1/(8 L^{1/2})\). One can then solve \(\sigma _\text {HF}'(J=0; L) = \Delta \sigma = 0.02\) to obtain \(L^* = 1/(64 \Delta \sigma ^2)\). The same approximation leads to \(\sigma _\text {HF}''(J=0; L) \approx 1/(32 L^{1/2})\). These approximations show that both the first and second derivatives of \(\sigma _\text {HF}\) at zero coupling (\(J=0\)) decrease as \(L^{-1/2}\). For high coupling, \(J \gg J_\text {max}\), the system assumes a bimodal character and the standard deviation saturates at the infinite coupling limit.
We conclude, based on our definition and our choice of \(\Delta \sigma\), that a system is small if \(L \le 39\). We also observe that for any system of finite size with \(L > 39\), there is always a feasibility window for sufficiently strongly cooperative systems, \(J_{\textrm{min}}< J < J_\textrm{max}\), characterized by a peak at a value of \(J = J^*\) that increases logarithmically with increasing L, but with \(J_{\textrm{min}} > 0\). Consequently, for \(L \gg 39\), the standard deviation at HF is insensitive to J both at low coupling, where the system acts as a Hill–Langmuir one, and high coupling, where the system becomes bimodal.

(a) Derivative of the standard deviation at half-filling, \(\sigma _\text {HF}'(J;L) \equiv \text {d}\sigma _\text {HF}/\text {d}J\), plotted for systems of size \(L=10,\, 39,\, 10^2,\,10^3,\,10^4\), respectively. The horizontal dashed gray line represents the threshold value \(\Delta \sigma =0.02\). (b) Contour plot of \(\sigma _\text {HF}'(J;L)\) for positive values of J and \(L\ge 3\). The red contour lines correspond to \(\sigma _\text {HF}'(J;L) = \Delta \sigma =0.02\).
The (semi-log) contour plot in Fig. 11b gives the isolines of \(\sigma _\text {HF}'(J)\) as a function of the interaction potential J and the system size L, where the latter, for the sake of simplicity, is treated as a continuous variable. Recall the asymptotic behavior of the correlation length at half-filling for large values of the interaction potential38: \(J \sim 2 \ln \left( 2 \xi _\text {HF}\right)\). Hence, if we were to plot the logarithm of \(\xi _\text {HF}\) instead of J on the horizontal axis, we would get a qualitatively equivalent contour diagram. The two red contour lines correspond to the constant value \(\Delta \sigma =0.02\). The region between them indicates the set of parameters J and L for which \(\sigma _\text {HF}'(J)>\Delta \sigma\). We finally conclude that a system of size \(L=13\), like the one considered in the main text to describe the BFM, is well within the feasibility window for small systems.
Effective Hill coefficient
Following the analysis in11 on the logarithmic sensitivity of kinetics schemes, we can also quantify cooperative behavior by introducing an effective Hill coefficient which, for our system, takes the following form:
Here, we have exploited the relation \(\partial _\mu \langle \varphi \rangle = \sigma ^2 L\).
The effective Hill function defined above takes on a maximal value \(H_\text {eff}^*\) at half-filling:
where \(\sigma _\text {HF}^2\) is the square of the relative standard deviation at half-filling whose explicit dependency on J is captured by Eq. (5). This expression tends rapidly to zero for negative values of J (anticooperativity). Moreover, for \(J=0\), the coefficient equals 1, as expected for the Hill–Langmuir case. Finally, by increasing the interaction potential, \(\sigma _\text {HF}\) tends to 1/2, and so \(H^*_\text {eff}\rightarrow L\). This result is in agreement with the rule that at equilibrium, the Hill coefficient is expected not to exceed the maximal number of ligands that can bind simultaneously, which, in the case of an adsorption process, corresponds to the number of binding sites, L. Fig. 12 shows the sigmoidal allure of the Hill coefficient, bounded from above by the system size and below by 0.
It should be noted, however, that in11 it has been demonstrated that this upper bound for the effective Hill coefficient may be surpassed when the system is out-of-equilibrium. In such cases, the new limit is determined not by the size of the lattice, but by the number of states of the kinetic scheme whose exit rates increase with the concentration of the ligands.

The effective Hill coefficient at half-filling from Eq. (83) as a function of the interaction potential J for a system of size \(L=13\).
Note that Eq. (83) depends only on J, and not on \(\mu\). Therefore, by approximating \(H_\text {eff}\approx H^*_\text {eff}\) in Eq. (82) and solving for \(\sigma ^2\), we obtain an approximate, explicit analytic expression for the fluctuation amplitude as a function of the mean occupancy \(\langle \varphi \rangle\) and the interaction potential J:
As shown in Fig. 13, this expression holds for non-negative J, and becomes exact at half-filling. It overestimates the fluctuations but performs well for intermediate occupancies. The discrepancy with respect to the exact curve for low and high values of the occupancy levels—increasing for intermediate values of J—vanishes in both limits \(J\rightarrow 0\) and \(J\rightarrow \infty .\)

Comparison between the exact result for the standard deviation (solid lines) and the approximate expression (dashed lines) that depends explicitly on \(\langle \varphi \rangle\), J, and L [Eq. (84)].
Supplemental plots
See Figs. 14, 15, 16, 17, 18, 19 and 20.

Contour plot of the mean relative occupancy in equilibrium, \(\langle \varphi \rangle\), as a function of the dimensionless interaction potential J and chemical potential \(\mu\) for a system of size \(L = 14\).

Contour plot of the standard deviation of the relative occupancy at equilibrium, \(\sigma = \sqrt{\langle \varphi ^2 \rangle - \langle \varphi \rangle ^2}\), as a function of the dimensionless interaction potential J and chemical potential \(\mu\) for a system of size \(L=14\).

Standard deviation of the mean occupancy at half-filling, \(\langle \varphi \rangle =1/2\), as a function of the dimensionless interaction potential J for a system of size \(L=13\) (blue) and \(L=14\) (orange). For \(J \ge 0\) the prediction of the block domain approximation, Eq. (13), is indistinguishable from the exact result.

Correlation length, \(\xi\), in the limit of \(J\rightarrow \infty\). It diverges at half-filling and is mainly higher than \(L/2 = 6.5\) (indicated by the red line), except near zero and full fillings.

Discrete equilibrium probability distribution of bound stator units, P(N), from the experiments (colored dots with error bars corresponding to one standard deviation of uncertainty; from left to right increasing load: 300 nm, 500 nm and 1300 nm beads) and the best-fit value of \(J = 1.21\), obtained by fitting Eq. (79) to the experimental data. The mean equilibrium occupancy of each distribution is indicated on top of the three plots, which correspond to a given external load imposed by the different bead sizes.

Anti-cooperativity signatures for even and odd lattices for different system sizes: Parametric plots of the standard deviation of the relative occupancy, \(\sigma\), versus the mean occupancy, \(\langle \varphi \rangle\), for increasing values of the system size L. In each plot, the values of the dimensionless interaction potential J span from \(-10\) to 10. The signature differences between L even and odd, visible for negative values of the interaction potential J, which are due to particle-hole defects in the case of odd L compared to the case of even L, diminish as the system size increases.

Comparison between the exact result for the standard deviation (solid lines) and the approximate expression (dashed lines) that depends explicitly on \(\langle \varphi \rangle\), J, and L [Eq. (79)].
Experimental data
The experimental methods used to probe the activity of single flagellar motors are described in detail in15,16. Briefly, E. coli cells lacking flagellar filaments and containing an endogenously biotinylated hook were adhered to a coverslip. The hook can bind to the surface of a straptividin-conjugated micron-sized bead. The slightly off-axis rotation of the bead in the liquid is tracked and provides a proxy for the angular rotation of the motor. The binding (unbinding) of individual stator units in the motor complex gives a sudden increase (decrease) in angular speed, enabling the quantification of the stator occupancy.
Each experimental trace15,16 was sampled at 10 Hz to avoid temporal correlations due to the hook of the BFM, which acts as a torsional elastic link between the motor and attached particle. For a given viscous load, the mean and standard deviation of the relative occupancy of stator units were calculated for each time point across all traces, yielding \(\langle \varphi (t) \rangle\) and \(\sigma (t)\), respectively; the mean of \(\langle \varphi (t) \rangle\) over all time points gave \(\langle \varphi \rangle\), and the mean of \(\sigma (t)\) over all time points gave \(\sigma\) (Fig. 7, Table 1). This approach was used because the experimental time traces were short, rendering them insufficient to assess the intracellular stationarity of single-cell traces. By calculating the mean and standard deviation in this manner, we assume that the intercellular variability is insignificant, which remains to be tested. Consequently, our method provides an upper estimate of the cooperativity J. Future studies probing individual motors for extended periods will enable a more accurate calculation of stator cooperativity.
Data availability
The present article does not present new experimental data. The analysis was conducted using previously published single-molecule data within the theoretical framework developed in the present study (see Section “Experimental data” for details on the data and experimental setup). The data analyzed originate from articles [15] and [16]. The data availability statements contained in the above articles remain valid. For further information regarding the data or analysis, please contact A.L.N. (ashley.nord@cbs.cnrs.fr) and F.P. (francesco.pedaci@cbs.cnrs.fr).
References
Whitty, A. Cooperativity and biological complexity. Nat. Chem. Biol. 4(8), 435–439 (2008).
Ogata, R. T. & McConnell, H. M. Mechanism of cooperative oxygen binding to hemoglobin. Proc. Natl. Acad. Sci. 69(2), 335–339 (1972).
Dill, K. A., Fiebig, K. M. & Chan, H. S. Cooperativity in protein-folding kinetics. Proc. Natl. Acad. Sci. 90(5), 1942–1946 (1993).
Tielrooij, K. J., Garcia-Araez, N., Bonn, M. & Bakker, H. J. Cooperativity in ion hydration. Science 328(5981), 1006–1009 (2010).
Williamson, J. R. Cooperativity in macromolecular assembly. Nat. Chem. Biol. 4(8), 458–465 (2008).
West, S. A., Diggle, S. P., Buckling, A., Gardner, A. & Griffin, A. S. The social lives of microbes. Annu. Rev. Ecol. Evol. Syst. 38, 53–77 (2007).
Monod, J., Wyman, J. & Changeux, J.-P. On the nature of allosteric transitions: A plausible model. J. Mol. Biol. 12(1), 88–118 (1965).
Changeux, J.-P., Thiéry, J., Tung, Y. & Kittel, C. On the cooperativity of biological membranes. Proc. Natl. Acad. Sci. 57(2), 335–341 (1967).
Blom, K. & Godec, A. Criticality in cell adhesion. Phys. Rev. X 11, 031067 (2021).
Hill, T. L. Cooperativity Theory in Biochemistry. Steady-State and Equilibrium Systems (Springer, 1985).
Owen, J. A. & Horowitz, J. M. Size limits the sensitivity of kinetic schemes. Nat. Commun. 14(1), 1280 (2023).
Berg, C. H. The rotary motor of bacterial flagella. Annu. Rev. Biochem. 72(1), 19–54 (2003).
Nirody, J. A., Sun, Y.-R. & Lo, C.-J. The biophysicist’s guide to the bacterial flagellar motor. Adv. Phys. X 2(2), 324–343 (2017).
Ito, K. I., Nakamura, S. & Toyabe, S. Cooperative stator assembly of bacterial flagellar motor mediated by rotation. Nat. Commun. 12, 3218 (2021).
Nord, A. L. et al. Catch bond drives stator mechanosensitivity in the bacterial flagellar motor. Proc. Natl. Acad. Sci. USA 114(49), 12952–12957 (2017).
Perez-Carrasco, R. et al. Relaxation time asymmetry in stator dynamics of the bacterial flagellar motor. Sci. Adv. 8(12), eabl8112 (2022).
Bai, F. et al. Conformational spread as a mechanism for cooperativity in the bacterial flagellar switch. Science 327(5966), 685–689 (2010).
Fukuoka, H., Wada, T., Kojima, S., Ishijima, A. & Homma, M. Sodium-dependent dynamic assembly of membrane complexes in sodium-driven flagellar motors. Mol. Microbiol. 71, 825–35, 02 (2009).
Tipping, M. J., Delalez, N. J., Lim, R., Berry, R. M. & Armitage, J. P. Load-dependent assembly of the bacterial flagellar motor. mBio 4(4), e00551-13 (2013).
Castillo, D. et al. The c-terminal periplasmic domain of motb is responsible for load-dependent control of the number of stators of the bacterial flagellar motor. Biophysics 9, 173–181, 12 (2013).
Wadhwa, N., Phillips, R. & Berg, H. C. Torque-dependent remodeling of the bacterial flagellar motor. Proc. Natl. Acad. Sci. USA 116(24), 11764–11769 (2019).
Nirody, J. A., Nord, A. L. & Berry, R. M. Load-dependent adaptation near zero load in the bacterial flagellar motor. J. R. Soc. Interface 16(159), 20190300 (2019).
Bai, F., Minamino, T., Wu, Z., Namba, K. & Xing, J. Coupling between switching regulation and torque generation in bacterial flagellar motor. Phys. Rev. Lett. 108, 178105 (2012).
Skoge, M. L., Endres, R. G. & Wingreen, N. S. Receptor-receptor coupling in bacterial chemotaxis: evidence for strongly coupled clusters. Biophys. J . 90(12), 4317–4326 (2006).
Skoge, M., Meir, Y. & Wingreen, N. S. Dynamics of cooperativity in chemical sensing among cell-surface receptors. Phys. Rev. Lett. 107(17), 178101 (2011).
Skoge, M., Naqvi, S., Meir, Y. & Wingreen, N. S. Chemical sensing by nonequilibrium cooperative receptors. Phys. Rev. Lett. 110(24), 248102 (2013).
Tu, Y. Quantitative modeling of bacterial chemotaxis: signal amplification and accurate adaptation. Annu. Rev. Biophys. 42, 337–359 (2013).
Liu, Y. & Dilger, J. P. Application of the one-and two-dimensional ising models to studies of cooperativity between ion channels. Biophys. J . 64(1), 26–35 (1993).
Cao, W., Taylor, E. W. & De La Cruz, E. M. Cooperative ligand binding to a double-stranded ising lattice–application to cofilin binding to actin filament. PNAS Nexus 2, 1–13 (2023).
Rice, J. J., Stolovitzky, G., Tu, Y. & de Tombe, P. P. Ising model of cardiac thin filament activation with nearest-neighbor cooperative interactions. Biophys. J . 84(2), 897–909 (2003).
Duke, T., Le Novère, N. & Bray, D. Conformational spread in a ring of proteins: a stochastic approach to allostery. J. Mol. Biol. 308(3), 541–553 (2001).
Hill, T. L. Statistical Mechanics. Principles and Selected Applications (McGraw-Hill, 1956).
McCoy, B. M. & Wu, T. T. The Two-Dimensional Ising Model (Courier Corporation, 2014).
Reid, S. W. et al. The maximum number of torque-generating units in the flagellar motor of Escherichia coli is at least 11. Proc. Natl. Acad. Sci. 103(21), 8066–8071 (2006).
Morimoto, Y. & Minamino, T. Structure and function of the bi-directional bacterial flagellar motor. Motor. Biomol. 4, 217–234 (2014).
Chang, Y. et al. Molecular mechanism for rotational switching of the bacterial flagellar motor. Nat. Struct. Mol. Biol. 27(11), 1041–1047 (2020).
Drobnič, T. et al. In situ structure of a bacterial flagellar motor at subnanometre resolution reveals adaptations for increased torque. Nat. Microbiol. 10, 1–18 (2025).
Godreche, C. & Luck, J. Response of non-equilibrium systems at criticality: exact results for the Glauber–Ising chain. J. Phys. A Math. Gen. 33(6), 1151 (2000).
Duke, T. & Bray, D. Heightened sensitivity of a lattice of membrane receptors. Proc. Natl. Acad. Sci. 96(18), 10104–10108 (1999).
Li, W. et al. Thermodynamic coupling between neighboring binding sites in homo-oligomeric ligand sensing proteins from mass resolved ligand-dependent population distributions. Protein Sci. 31(10), e4424 (2022).
Purcell, E. M. Life at low Reynolds number. Am. J. Phys. 45(1), 3–11 (1977).
Kearns, D. A field guide to bacterial swarming motility. Nat. Rev. Microbiol. 8, 634–644 (2010).
Spormann, A. M. Gliding motility in bacteria: Insights from studies of Myxococcus xanthus. Microbiol. Mol. Biol. Rev. 63(3), 621–641 (1999).
Henrichsen, J. Twitching motility. Annu. Rev. Microbiol. 37(1), 81–93 (1983).
Martínez, A., Torello, S. & Kolter, R. Sliding motility in mycobacteria. J. Bacteriol. 181(23), 7331–7338 (1999).
Dvoriashyna, M. & Lauga, E. Hydrodynamics and direction change of tumbling bacteria. PLoS ONE 16, 1–26, 07 (2021).
Wadhwa, N. & Berg, H. Bacterial motility: Machinery and mechanisms. Nat. Rev. Microbiol. 20, 161–173 (2022).
Nakamura, S. & Minamino, T. Flagella-driven motility of bacteria. Biomolecules 9(7), 279 (2019).
Grognot, M. & Taute, K. M. More than propellers: How flagella shape bacterial motility behaviors. Curr. Opin. Microbiol. 61, 73–81 (2021).
Brown, M. T. et al. Flagellar hook flexibility is essential for bundle formation in swimming Escherichia coli cells. J. Bacteriol. 194(13), 3495–3501 (2012).
Gabel, C. V. & Berg, H. C. Speed of the flagellar rotary motor of Escherichia coli varies linearly with protonmotive force. Proc. Natl. Acad. Sci. USA 100(15), 8748–51 (2003).
Hosking, E. R., Vogt, C., Bakker, E. P. & Manson, M. D. Escherichia coli MotAB proton channel unplugged. J. Mol. Biol. 364(5), 921–937 (2006).
Sowa, Y., Homma, M., Ishijima, A. & Berry, R. M. Hybrid-fuel bacterial flagellar motors in Escherichia coli. Proc. Natl. Acad. Sci. USA 111(9), 3436–3441 (2014).
Ryu, W. S., Berry, R. M. & Berg, H. C. Torque-generating units of the flagellar motor of Escherichia coli have a high duty ratio. Nature 403(6768), 444–7 (2000).
Reid, S. W. et al. The maximum number of torque-generating units in the flagellar motor of Escherichia coli is at least 11. Proc. Natl. Acad. Sci. USA 103(21), 8066–71 (2006).
Fung, D. C. & Berg, H. C. Powering the flagellar motor of Escherichia coli with an external voltage source. Nature 375, 809–812 (1995).
Kojima, S., Nonoyama, N., Takekawa, N., Fukuoka, H. & Homma, M. Mutations targeting the C-terminal domain of FliG can disrupt motor assembly in the Na(+)-driven flagella of Vibrio alginolyticus. J. Mol. Biol. 414(1), 62–74 (2011).
Biquet-Bisquert, A., Labesse, G., Pedaci, F. & Nord, A. L. The dynamic ion motive force powering the bacterial flagellar motor. Front. Microbiol. 12, 659464 (2021).
Lele, P. P., Branch, R. W., Nathan, V. S. J. & Berg, H. C. Mechanism for adaptive remodeling of the bacterial flagellar switch. Proc. Natl. Acad. Sci. 109(49), 20018–20022 (2012).
Lele, P. P., Hosu, B. G. & Berg, H. C. Dynamics of mechanosensing in the bacterial flagellar motor. Proc. Natl. Acad. Sci. USA 110(29), 11839–44 (2013).
Hu, X., Margadant, F. M., Yao, M. & Sheetz, M. P. Molecular stretching modulates mechanosensing pathways. Protein Sci. 26(7), 1337–1351 (2017).
Wadhwa, N., Tu, Y. & Berg, H. C. Mechanosensitive remodeling of the bacterial flagellar motor is independent of direction of rotation. Proc. Natl. Acad. Sci. USA 118(15), e2024608118 (2021).
Sourjik, V. & Wingreen, N. S. Responding to chemical gradients: Bacterial chemotaxis. Curr. Opin. Cell Biol. 24(2), 262–268 (2012) (Cell regulation).
Ma, Q., Sowa, Y., Baker, M. & Bai, F. Bacterial flagellar motor switch in response to chey-p regulation and motor structural alterations. Biophys. J . 110, 1411–1420, 03 (2016).
Yuan, J. & Berg, H. Ultrasensitivity of an adaptive bacterial motor. J. Mol. Biol. 425, 02 (2013).
Tusk, S. E., Delalez, N. J. & Berry, R. M. Subunit exchange in protein complexes. J. Mol. Biol. 430(22), 4557–4579 (2018) (Plasticity of Multi-Protein Complexes).
Santiveri, M. et al. Structure and function of stator units of the bacterial flagellar motor. Cell 183(1), 244–257 (2020).
Deme, J. et al. Structures of the stator complex that drives rotation of the bacterial flagellum. Nat. Microbiol. 5, 1553–1564 (2020).
Zhu, S. et al. Conformational change in the periplamic region of the flagellar stator coupled with the assembly around the rotor. Proc. Natl. Acad. Sci. USA 111(37), 13523–13528 (2014).
Kojima, S. et al. Stator assembly and activation mechanism of the flagellar motor by the periplasmic region of motb. Mol. Microbiol. 73(4), 710–8 (2009).
Tipping, M. J., Steel, B. C., Delalez, N. J., Berry, R. M. & Armitage, J. P. Quantification of flagellar motor stator dynamics through in vivo proton-motive force control. Mol. Microbiol. 87(2), 338–347 (2013).
Kojima, S. et al. The helix rearrangement in the periplasmic domain of the flagellar stator b subunit activates peptidoglycan binding and ion influx. Structure 26(4), 590–598 (2018).
Terahara, N. et al. Na-induced structural transition of motps for stator assembly of the flagellar motor. Sci. Adv. 3(11), 4119 (2017).
Lin, T.-S., Zhu, S., Kojima, S., Homma, M. & Lo, C.-J. Flil association with flagellar stator in the sodium-driven vibrio motor characterized by the fluorescent microscopy. Sci. Rep. 8(1), 11172 (2018).
Wadhwa, N., Sassi, A., Berg, H. C. & Tu, Y. A multi-state dynamic process confers mechano-adaptation to a biological nanomachine. Nat. Commun. 13(1), 5327 (2022).
Rieu, M., Krutyholowa, R., Taylor, N. M. & Berry, R. M. A new class of biological ion-driven rotary molecular motors with 5: 2 symmetry. Front. Microbiol. 13, 948383 (2022).
Leach, J. et al. Comparison of faxén’s correction for a microsphere translating or rotating near a surface. Phys. Rev. E 79(2), 026301 (2009).
Kardar, M. Statistical Physics of Fields (Cambridge University Press, 2007).
Acknowledgements
The authors thank Anaïs Biquet-Bisquert for fruitful discussions. This work was financially supported by the ANR FlagMotor project grant ANR-18-CE30-0008 of the French Agence Nationale de la Recherche and by the LabEx NUMEV (ANR-10-LABX-0020) within the I-Site MUSE (ANR-16-IDEX-0006).
Author information
Authors and Affiliations
Contributions
M.-J.F.-O., J.P., and N.-O.W. conceived the research and derived the theoretical (analytical and numerical) results. A.L.N. and F.P. designed and carried out the experiments. M.-J.F.-O. compared the theoretical results to the experimental BFM data with assistance from A.L.N. and F.P. M.-J.F.-O., J.P., A.L.N., F.P., and N.-O.W. wrote the manuscript, with contributions to the manuscript and conceptual development of the theoretical framework from A.P., F.G., J.D., and J.-C.W.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Franco-Oñate, MJ., Parmeggiani, A., Dorignac, J. et al. Signature of cooperativity in the stochastic fluctuations of small systems with application to the bacterial flagellar motor. Sci Rep 15, 32109 (2025). https://doi.org/10.1038/s41598-025-14570-3
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-14570-3
