
Abstract
Mosquitoes pose serious public health risks, especially in densely populated cities. Although mosquito populations are closely linked to meteorological factors, the cumulative effects of these factors remain understudied from an ecological perspective. This study investigates mosquito population patterns in relation to cumulative meteorological factors, analyzing daily data collected nightly (18:00–07:00) from May to October in 2012, 2013, and 2015 at 21 monitoring sites across different urban habitats in Seoul, South Korea. Results revealed that cumulative temperature and precipitation significantly influence mosquito populations, with marked differences observed between waterside and non-waterside habitats. Mosquito populations in waterside areas were more sensitive to temperature fluctuations and the number of days with precipitation compared with those in non-waterside areas. Although higher minimum temperatures increased mosquito abundance, extreme variations led to declines. Cumulative precipitation generally had a negative impact on mosquito populations, whereas low precipitation showed a positive correlation. Notably, the number of days with precipitation was more influential than cumulative precipitation. These findings emphasize the importance of habitat types and cumulative meteorological factors in shaping urban mosquito populations, offering insights for future research and control measures.
Similar content being viewed by others

Introduction
Meteorological factors, including temperature and precipitation, play a crucial role in shaping the occurrence and distribution of mosquitoes1,2. Temperature affects various aspects of mosquito biology, including development rate, reproduction, and survival3 while precipitation creates breeding sites, as standing water serves as an ideal habitat for mosquito larvae4,5,6.
The influence of temperature on mosquito populations is well-documented, with minimum temperature being particularly critical7. Understanding minimum temperature effects is essential for predicting mosquito abundance and distribution, as well as for implementing control measures1. Lim et al.8 showed that the abundance of mosquito species, such as Culex pipiens and Aedes vexans, peaks at 23–26 °C but decreases at extremely high temperatures. Additionally, changes in temperature and precipitation patterns affect mosquito-borne disease transmission dynamics7,9 with higher temperatures and reduced precipitation linked to increased disease rates10. Therefore, understanding these meteorological influences is pivotal for managing mosquito populations and mitigating associated mosquito-borne disease risk.
Although the direct effects of temperature and precipitation on mosquito populations are well-established11,12 the cumulative or delayed effects of these factors on mosquito occurrence patterns remain understudied. Shaman and Day13 demonstrated that precipitation frequency markedly impacts mosquito population dynamics and mosquito-borne disease transmission rates. However, prior research has mainly focused on immediate correlations between meteorological variables and mosquito abundance, often neglecting potential delayed effects14. Examining these cumulative impacts can provide a deeper understanding of how weather conditions influence mosquito dynamics over time, enhancing our ability to predict and manage mosquito-borne diseases effectively9,15.
Globally, mosquito-related public health issues are steadily increasing. Indeed, many countries face major health challenges due to mosquitoes16,17 with South Korea similarly affected. Malaria, eradicated in South Korea in 1979, re-emerged in 1993, with case numbers rising to 294 in 2021 and 747 in 202318,19. Similarly, Japanese encephalitis, first reported in 1947, averaged about 10 annual cases from 2001 to 2009, but case numbers increased to 23 in 2021 and 17 in 202319,20. Numerous studies have explored mosquito distribution in South Korea21,22 and examined relationships between mosquito populations and environmental conditions, including meteorological and spatial factors8,23,24,25. However, few studies have quantitatively assessed associations between mosquito abundance and environmental conditions in urban areas, particularly regarding cumulative meteorological effects.
Mosquitoes pose a significant public health threat, particularly in densely populated urban areas where environmental conditions facilitate their proliferation26,27. Climate change, the urban heat island effect, and water accumulation in artificial structures can extend mosquito activity periods and increase breeding opportunities26. These conditions heighten the risk of mosquito-borne diseases, such as dengue, chikungunya, and malaria, which may spread rapidly in cities with high population density and extensive transportation networks17,24.
Seoul, the largest metropolitan area in South Korea, has a population exceeding 9 million as of 2025 28. The city contains a diverse range of urban habitats, from dense commercial and residential districts to extensive parklands and riverine areas. The hot, humid summers and pronounced urban heat island effect create favorable conditions for mosquito proliferation. In addition, Seoul’s active public health policies, including long-term mosquito surveillance and vector control programs, provide extensive datasets and an institutional framework that facilitate comprehensive research on urban mosquito ecology. These factors make Seoul a highly relevant and representative setting for such studies.
This study investigated how temperature and precipitation impact mosquito populations across different habitat types, aiming to inform more effective control and disease management strategies. Specifically, we analyzed mosquito occurrence patterns in relation to cumulative meteorological values over specific periods and evaluated these patterns across various urban habitats in metropolitan Seoul, South Korea.
Results
Daily mosquito abundance fluctuated, with high values predominately in summer and autumn (Fig. 1). Throughout the study, annual precipitation in all areas showed a yearly decrease (1343.7 mm, 1012.6 mm, and 531.8 mm in 2012, 2013, and 2015, respectively), whereas temperature remained relatively stable (average temperature: 22.6 ℃, 22.7 ℃, and 22.7 ℃ in 2012, 2013, and 2015, respectively). Mosquito abundance was consistently higher in waterside habitat (WH) compared with non-waterside habitat (NH) across the study period. Mean annual abundance for WH/NH was 66.2/13.3, 213.8/59.7, and 141.0/27.2 in 2012, 2013, and 2015, respectively. No distinct precipitation or temperature differences were observed between habitat types.

Changes in mosquito abundance (a, b), precipitation (c, d), and temperature (e, f) by year in waterside (a, c, e) and non-waterside (b, d, f) areas. Values shown are averages for each area. Gray lines in temperature plots represent minimum and maximum temperatures.
Mosquito abundance was closely associated with cumulative temperature and precipitation factors. Abundance patterns were notably shaped by cumulative minimum temperature, with variations across habitat types (Fig. 2). Overall, WH areas exhibited higher mosquito abundance compared with NH areas, although abundance in both habitats decreased beyond a specific temperature threshold. In WH areas, mosquito abundance increased with daily minimum temperature up to around 25 ℃ before decreasing, whereas abundance in NH areas changed little across the same range. For cumulative minimum temperatures, abundance in both habitats increased for 5-day and 10-day accumulations, peaking at approximately 120 ℃ and 250 ℃, respectively, before declining. For 30-day accumulations, abundance varied widely but generally decreased above 700 ℃.

Changes in mosquito abundance as a function of cumulative minimum temperature over 1, 5, 10, and 30 days. Data and error bars represent means and standard errors, respectively. Solid lines with gray shaded areas are polynomial regression lines with 95% confidence intervals. A third-order polynomial was best for 30 days; a fourth-order polynomial was best for the other periods.
Daily precipitation and cumulative precipitation over five days showed a negative linear relationship with mosquito abundance in both habitat types (Fig. 3). When the cumulative precipitation period exceeded 10 days, mosquito populations declined under both low and high precipitation conditions. In WH areas, the decline was more pronounced at high precipitation levels than in NH areas, particularly above ~ 300 mm for 10-day and ~ 480 mm for 30-day accumulations.

Changes in mosquito abundance as a function of cumulative precipitation over 1, 5, 10, and 30 days. Data and error bars represent means and standard errors, respectively. Solid lines with gray shaded areas are polynomial regression lines with 95% confidence intervals, respectively. A first-order regression was best for daily and 5-day cumulative precipitation; a fourth-order polynomial fit was best for the other periods.
The relationship between the number of rainy days and mosquito abundance differed between habitats (Fig. 4). In WH areas, abundance peaked at approximately six rainy days out of 10 and 11 rainy days out of 30. In contrast, abundance in NH areas increased steadily with the number of rainy days across all examined time windows.

Changes in mosquito abundance as a function of the number of days with precipitation over 1, 5, 10, and 30 days. Data and error bars represent means and standard errors, respectively. Solid lines with gray shaded areas are polynomial regression lines with 95% confidence intervals, respectively. A third-order regression was best for 5- and 30-day periods; a fourth-order polynomial fit was best for 10-day period.
We used 3D surface plots to examine how mosquito abundance responded to combinations of two meteorological variables (Fig. 5). In WH areas, abundance was highest under moderate cumulative precipitation (~ 100–150 mm) combined with higher cumulative minimum temperatures. In NH areas, abundance increased almost linearly with the number of rainy days, while cumulative minimum temperature had little effect except under low precipitation. Across habitats, cumulative minimum temperature generally had a stronger influence on abundance than the number of rainy days (Fig. 5c and f).

Surface plots illustrating the relationship between cumulative meteorological factors and mosquito abundance. Five-day cumulative meteorological factors are shown: cumulative precipitation and the number of days with precipitation (a, d), cumulative minimum temperature and cumulative precipitation (b, e), and cumulative minimum temperature and the number of days with precipitation (c, f) in waterside (a–c) and non-waterside (d–f) areas. Smooth surfaces were created using generalized additive models.
CCM analysis in the forward direction (meteorological factors to mosquito abundance) confirmed that daily and cumulative meteorological factors influenced mosquito abundance in both habitat types (p < 0.05; Fig. 6). Except for the daily meteorological variables in WH areas, all variables exhibited an increase in cross map skill (\(\:{\uprho\:}\), calculated as the Pearson correlation coefficient between reconstructed and observed time series) as the library size (number of subsampled data points) increased. In WH areas, the effect of minimum temperature was initially strongest for up to five days, whereas after 10 days, precipitation had the greatest influence. In NW areas, the minimum temperature among daily meteorological factors, precipitation among cumulative variables over five days, and the number of days with precipitation over 10 days had the greatest effects on mosquito abundance. Additionally, as the number of days increased, the \(\:{\uprho\:}\) value for minimum temperature decreased in WH areas but increased in NH areas (Supplementary Fig. S1). Supplementary Fig. S1 presents bidirectional CCM results, including the reverse direction (mosquito abundance to minimum temperature), in which \(\:{\uprho\:}\) values, although not all negative, converged toward zero, indicating no predictive skill. For precipitation and the number of days with precipitation, \(\:{\uprho\:}\) values increased with the number of days for both habitat types.

Causality analysis of cumulative meteorological factors in relation to mosquito abundance based on convergent cross mapping. The y-axes represent the cross map skill (\(\:{\uprho\:}\)). Meteorological factors by cumulative days in waterside habitat areas (a–d): daily (a) and 5 (b), 10 (c), and 30 (d) cumulative days. Meteorological factors by cumulative days in nonwaterside areas (e–h): daily (e) and 5 (f), 10 (g), and 30 (h) cumulative days. Abbreviations: Tmin, minimum temperature; Pday, number of days with precipitation; Prec, precipitation.
Discussion
In our study area, Culex pipiens pallens accounted for the vast majority of captured mosquitoes (> 90% in similar surveys1. Accordingly, mosquito abundance in this study was interpreted primarily as a proxy for the abundance of Cx. pipiens pallens. This assumption provides ecological and spatial consistency in our interpretation, as the species’ average flight distance (~ 400 m) informed the habitat classification radius used in our analysis. By focusing on this dominant species, we ensure that the observed associations with meteorological variables and habitat characteristics are interpreted within a coherent ecological framework.
Habitat type based on land cover is highly influential on mosquito abundance29,30,31. The present study revealed distinct differences in mosquito abundance and responses to meteorological factors between WH and NH areas, with the former showing greater sensitivity to meteorological factors. These differences are attributed to variations in environmental factors across the habitat types. WH areas are directly exposed to such factors, including standing water in exposed locations, e.g., puddles, making them more reactive to temperature and precipitation changes. In contrast, NH areas tend to have sheltered, more stable environments, e.g., stagnant water in flowerpots and septic tanks inside buildings, where conditions remain less affected by outside weather changes. Consequently, mosquito population dynamics across these habitats are influenced by different ecological factors, such as water availability, humidity levels, temperature regulation, and food resources. Thus, the observed variation in mosquito abundance between the two habitat types likely results from these complex interactions influenced by land cover and exposure levels.
Temperature is a key factor for mosquito development, directly affecting their life cycle and boosting mosquito population growth as it rises3,15,32. Among various temperature metrics, the minimum temperature plays a critical role, serving as a threshold for survival, development, and activity in insects, including mosquitoes16,33. In this study, daily mosquito abundance increased with rising daily minimum temperatures in both habitat types (Fig. 2). However, extremely high cumulative minimum temperatures led to a decrease in mosquito abundance, particularly in WH areas. Mosquito abundance in these areas responded more strongly to cumulative minimum temperatures compared with that in NH areas, likely owing to greater exposure to fluctuating environmental conditions. A previous laboratory study has shown that temperature fluctuations, compared with constant conditions of the same mean, can significantly alter mosquito development, survival, and reproduction, thereby influencing population growth34. This finding supports our observation that WH populations, which experience greater environmental variability, are more sensitive to changes in minimum temperature. Under extreme heat, this heightened sensitivity, combined with habitat reduction caused by drought, may lead to sharp declines in mosquito numbers in this area.
CCM analysis in the forward direction (cumulative minimum temperature to mosquito abundance) revealed a significant causal relationship (p < 0.05) between cumulative minimum temperature and mosquito abundance (Fig. 6). Reverse-direction analyses are provided and consistently showed no predictive skill. The \(\:{\uprho\:}\) value increased with longer cumulative periods, although it decreased in WH areas for 30-day cumulative minimum temperatures compared with shorter periods (Supplementary Fig. S1). This variation suggests that in WH areas, the relationship between temperature and mosquito abundance over 30 days differs from that in other areas and across intervals. These findings indicate that bioenvironmental relationships across habitat types and time are likely caused by specific environmental factors in WH areas, e.g., urban waterside conditions, suggesting the need for further research on mosquito populations in urban environments with such habitats.
Precipitation is also a key driver of mosquito habitat conditions6 directly impacting breeding sites and indirectly affecting resources, such as food availability35making it essential to mosquito occurrence12,36. The effects of precipitation on mosquito populations vary across studies, with some suggesting that increased rainfall expands mosquito habitats and boosts abundance6,37 whereas others indicate that heavy precipitation can wash away larvae and inhibit adult mosquito activity. In the present study, mosquito abundance generally decreased with increasing precipitation levels in both habitat types (Fig. 3). However, for cumulative precipitation over 10 days, a positive correlation emerged between precipitation and mosquito abundance when rainfall levels were low. In contrast, in WH areas, mosquito populations declined sharply with high cumulative precipitation. The CCM results (Fig. 6) showed a stronger causal relationship between cumulative precipitation and mosquito populations, particularly in WH areas, indicated by higher ρ values as the number of cumulative days increased.
Numerous studies have demonstrated that precipitation and humidity significantly influence mosquito populations29,38. The number of days with precipitation reflects environmental changes, capturing the frequency and duration of rainfall, key factors in mosquito breeding and survival. Therefore, this metric may be more relevant than cumulative precipitation in predicting mosquito abundance. Consistent with this, we found that mosquito abundance increased with the number of rainy days (Fig. 4) and was considerably lower when the number of days with rainfall was low, even when cumulative precipitation was high. The CCM analysis further indicated that in WH areas, mosquito populations were more strongly influenced by the number of days with precipitation than by cumulative rainfall, particularly when the sample (library) size increased (Fig. 6). Similarly, in NH areas, > 10 days of precipitation showed a stronger causal relationship with mosquito abundance when sample sizes were larger.
Our findings align with previous studies suggesting that climate change, urban heat island effects, and water accumulation in artificial structures can promote mosquito proliferation in urban settings25,26. In the study area, the observed increase in mosquito abundance following rainfall events supports the hypothesis that stagnant water in construction sites, discarded containers, and other urban structures serves as key breeding habitats. This pattern indicates that rising urban temperatures and extended warm seasons may further enhance the potential for disease transmission, consistent with earlier projections.
This study revealed that cumulative temperature and precipitation markedly influence mosquito abundance, with significant differences observed between habitat types in urban environments. Mosquito populations in WH areas, directly exposed to environmental changes, showed greater sensitivity to meteorological factors, particularly temperature and the number of days with precipitation. Increased minimum temperatures raised mosquito abundance in both habitat types, whereas extremely high or low cumulative minimum temperatures reduced abundance, with WH areas showing the sharpest decline. Mosquito abundance also decreased with increasing cumulative precipitation, although a positive correlation emerged at lower precipitation levels, particularly with cumulative precipitation over 10 days. Additionally, the cumulative number of rainy days appeared to be a more predictive variable for mosquito abundance compared with cumulative precipitation, especially with a larger sample size.
These findings highlight the importance of cumulative meteorological factors and habitat type in understanding urban mosquito population dynamics. In our study area, Cx pipiens pallens accounted for over 90% of captured mosquitoes in previous field surveys1and abundance was therefore interpreted primarily as a proxy for this species. While this focus ensured ecological consistency in linking meteorological variables and habitat characteristics, it may limit the generalizability of the findings to other mosquito species. For instance, the subdominant species Aedes koreicus (~ 5% of captures) is more tolerant of colder temperatures and capable of exploiting diverse urban habitats, and may respond differently to temperature and precipitation patterns39. In addition, relative humidity, which is an important factor for mosquito development and survival, was not included due to the lack of spatially and temporally matched data. This may have influenced our results, particularly regarding temperature and precipitation effects, and should be addressed in future studies to provide a more comprehensive understanding of meteorological influences on urban mosquito dynamics.
Urbanization and climate change are expected to increase mosquito prevalence in cities and related public health threats; therefore, these results offer insights relevant to mosquito-related urban ecology, public health, and control measures. In particular, the identification of seasonal peaks and differences between NH and WH clusters could help public health authorities recognize critical periods of heightened mosquito activity, thereby supporting the optimization of intervention timing within the existing DMS-based operational framework. This underscores the need for further research and targeted management approaches in urban settings.
Materials and methods
Data collection
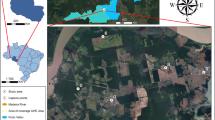
Mosquito abundance data were collected from the Yeongdeungpo-gu Public Health Center in Seoul, South Korea1,2 using daily measurements from 21 monitoring sites via the Digital Mosquito Monitoring system (DMS; Environmental Technology & Development, South Korea; Fig. 7). The DMS operated nightly from 18:00 to 07:00 from May to October over three years (2012, 2013, and 2015), achieving a 93% accuracy rate in mosquito counts when comparing transmitted data to actual numbers2. Further DMS details were reported in the previous study1. That work used data from 2011, 2012, 2013, and 2015. In the present study, the year 2011 was excluded because the survey that year was conducted only in the cluster 1(i.e., NH in this study), with no corresponding data for the WH. As the primary objective here was to compare NH and WH, including 2011 would have been unsuitable for comparative and causal analyses. The year 2014 was also excluded because numerous unexpected values were observed in the dataset due to problems with the DMS calibration, as reported in our previous study1. In the DMS data, mosquito species were not identified; however, data collected by the Seoul Metropolitan Government using light traps during 2011–2017 showed that Cx. pipiens pallens was the predominant species1.

Location of 21 monitoring sites (A–U) and 4 automatic weather stations (AWSs). Monitoring sites were classified into waterside and non-waterside types based on land cover1. Land cover ratios were calculated within a 400 m radius of each site. The figure was created using QGIS v3.22.9 (https://qgis.org), and the satellite imagery was provided by V-World, Republic of Korea (https://www.vworld.kr), and is used under the Korea Open Government License Type 1.
Meteorological data, including daily average, minimum, and maximum temperatures and daily precipitation, were sourced from automatic weather stations (AWSs) operated by the Korea Meteorological Administration (http://www.kma.go.kr). Data from the nearest AWSs (AWS 405: A, B, and C; AWS 410: O, P, Q, R, S, T, and U; AWS 418: K and L; AWS 510: D, E, F, G, H, I, J, and N) were used for each monitoring site. Land cover data (urban area, agricultural land, forest, grassland, barren land, wetland, and water) around each DMS site were obtained from the Environmental Geographic Information Service in South Korea (https://egis.me.go.kr).
Data organization and analysis
In this study, monitoring sites were classified into waterside habitats (WH) and non-waterside habitats (NH) following the approach of previous study1 (Fig. 1). That research applied a hierarchical cluster analysis to landscape descriptors (proportion of urban areas, agricultural areas, water, stream, and grass areas, and distance to rivers), revealing two distinct clusters: one characterized by higher proportions of water, stream, and grass areas (WH) and the other by higher proportions of urban and agricultural areas and greater distances from rivers (NW). Based on this framework, each site in our dataset was assigned to WH or NW according to the proportion of waterside and grass areas within a 400-m radius of each monitoring site on a digital map. Sites A, B, and R, despite their proximity to rivers, were classified as NW because their proportion of waterside and grass areas was similar to that of sites G, H, I, J, K, and L. Although the rivers adjacent to A, B, and R are narrower than the Han River, the classification was based on land-cover composition within the analysis radius rather than river width, as mosquitoes predominantly breed in stagnant zones along river margins2 making river width a minor factor for this purpose.
Mosquito and environmental data were grouped into these two categories such as WH an NH (Fig. 1). Meteorological data were accumulated over periods ranging from 1 to 30 days. Given the high correlation between temperature data (both r > 0.80, p < 0.001), daily minimum temperature was emphasized owing to its importance for mosquito development. Additionally, the number of days with precipitation was derived from precipitation data. Mosquito abundance data were preprocessed using a common logarithm: log10(abundance + 1).
These data were used to assess cumulative meteorological effects on mosquito occurrence. Trend lines showing changes in mosquito abundance based on cumulative meteorological factors were fitted with the best regression model, selected from the first- to fourth-order polynomials. Temperature and precipitation were averaged at intervals of 1℃ and 10 mm, respectively. Furthermore, three-dimensional surface plots of mosquito abundance versus cumulative meteorological variables were generated. The smooth surface of mosquito abundance in relation to two meteorological factors was generated using generalized additive models (GAMs) with restricted maximum likelihood.
Convergent cross mapping (CCM), a method for identifying causal linkages in complex dynamic systems40,41 was used to assess the impact and causality of accumulated meteorological factors on mosquito abundance. Our primary analyses focused on the forward direction of causality (meteorological factors to mosquito abundance), as this direction is most relevant to our ecological hypothesis. Reverse-direction analyses were conducted only for supplementary purposes to confirm the absence of causality in the opposite direction (Supplementary material Fig. S1).
All analyses were performed in R (version 4.4.1)42 using the “stats” package42 for polynomial regression, the “plot3D”43 and “mgcv” packages44 for surface plotting with GAMs, and the “rEDM” package for CCM45.
Data availability
The analysis code used in this study is publicly available at https://github.com/dleotjd520/UrbanMosq. Sample datasets are provided in the repository to illustrate the analysis workflow. The original datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request and with appropriate permissions.
References
Lee, D. S., Lee, D. Y. & Park, Y. S. Interpretable machine learning approach to analyze the effects of landscape and meteorological factors on mosquito occurrences in Seoul, South Korea. Environ. Sci. Pollut. Res. 30, 532–546. https://doi.org/10.1007/s11356-022-22099-5 (2023).
Kwon, Y. S. et al. Modeling occurrence of urban mosquitos based on land use types and meteorological factors in Korea. Int. J. Environ. Res. Public Health. 12, 13131–13147. https://doi.org/10.3390/ijerph121013131 (2015).
Beck-Johnson, L. M. et al. The importance of temperature fluctuations in understanding mosquito population dynamics and malaria risk. Royal Soc. Open. Sci. 4, 160969. https://doi.org/10.1098/rsos.160969 (2017).
Webb, C., Doggett, S. & Russell, R. A Guide To Mosquitoes of Australia (Csiro Publishing, 2016).
Wilke, A. B. B., Medeiros-Sousa, A. R., Ceretti-Junior, W. & Marrelli, M. T. Mosquito populations dynamics associated with climate variations. Acta Trop. 166, 343–350. https://doi.org/10.1016/j.actatropica.2016.10.025 (2017).
Jacob, B. G., Gu, W., Caamano, E. X. & Novak, R. J. Developing operational algorithms using linear and non-linear squares estimation in Python® for the identification of culex pipiens and culex restuans in a mosquito abatement district (Cook county, Illinois, USA). Geospat. Health. 3, 157–176 (2009).
Bellone, R. & Failloux, A. B. The role of temperature in shaping mosquito-borne viruses transmission. Front. Microbiol. 11, 584846. https://doi.org/10.3389/fmicb.2020.584846 (2020).
Lim, A. Y., Cheong, H. K., Chung, Y., Sim, K. & Kim, J. H. Mosquito abundance in relation to extremely high temperatures in urban and rural areas of Incheon metropolitan city, South Korea from 2015 to 2020: an observational study. Parasites Vectors. 14, 559. https://doi.org/10.1186/s13071-021-05071-z (2021).
Ruiz, M. O. et al. Local impact of temperature and precipitation on West nile virus infection in culex species mosquitoes in Northeast Illinois, USA. Parasites Vectors. 3, 19–19. https://doi.org/10.1186/1756-3305-3-19 (2010).
Giesen, C. et al. The impact of climate change on mosquito-borne diseases in Africa. Pathog Glob Health. 114, 287–301. https://doi.org/10.1080/20477724.2020.1783865 (2020).
Morin, C. W., Comrie, A. C. & Ernst, K. Climate and dengue transmission: evidence and implications. Environ. Health Perspect. 121, 1264–1272. https://doi.org/10.1289/ehp.1306556 (2013).
Baril, C. et al. The influence of weather on the population dynamics of common mosquito vector species in the Canadian prairies. Parasites Vectors. 16, 153 (2023).
Shaman, J. & Day, J. F. Reproductive phase locking of mosquito populations in response to rainfall frequency. PloS One. 2, e331. https://doi.org/10.1371/journal.pone.0000331 (2007).
Lambrechts, L. et al. Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. U.S.A. 108, 7460–7465. https://doi.org/10.1073/pnas.1101377108 (2011).
Roiz, D., Ruiz, S., Soriguer, R. & Figuerola, J. Climatic effects on mosquito abundance in mediterranean wetlands. Parasites Vectors. 7, 333. https://doi.org/10.1186/1756-3305-7-333 (2014).
da Ferreira, C. Meteorological variables and mosquito monitoring are good predictors for infestation trends of Aedes aegypti, the vector of dengue, Chikungunya and Zika. Parasites Vectors. 10, 1–11 (2017).
Weaver, S. C. & Lecuit, M. Chikungunya virus and the global spread of a mosquito-borne disease. N. Engl. J. Med. 372, 1231–1239. https://doi.org/10.1056/NEJMra1406035 (2015).
Kim, H., Lee, S., Shin, N. R. & Hwang, K. Status of malaria and diagnosis rate in the Republic of Korea, 2018–2022. Public. Health Wkly. Rep. 16, 852–866. https://doi.org/10.56786/PHWR.2023.16.26.3 (2023).
Korea Disease Control and Prevention Agency. 2023 Infectious Disease Report Annual Bulletin (2023 감염병 신고 현황 연보) (Korea Disease Control and Prevention Agency, 2024).
Lee, Y. J., Wang, J. S., Kang, H. J., Lee, D. Y. & Han, M. G. Laboratory-based diagnosis results of Japanese encephalitis in the Republic of Korea. Public. Health Wkly. Rep. 15, 2295–2303 (2022).
Lee, J. S. & Hong, H. K. Seasonal prevalence and behaviour of Aedes Togoi. Korean J. Parasitol. 33, 19–26 (1995).
Jeong, Y. S. & Lee, D. K. Prevalence and seasonal abundance of the dominant mosquito species in a large marsh near Coast of Ulsan. Korean J. Appl. Entomol. 42 (2), 125–132 (2003).
Yeom, J. S. & Park, J. W. Status of Vivax malaria after re-emergence in South Korea. Infect. Chemother. 40, 191–198 (2008).
Chae, S. M., Kim, D. J., Yoon, S. J. & Shin, H. S. The impact of temperature rise and regional factors on malaria risk. Health Social Welf. Rev. 34 (1), 436–455 (2014).
Kim, S. Y. & Kim, Y. H. Analysis of spatial pattern changes and its human and environmental factors of the malaria risks in Korea. J. Korean Cartographic Assoc. 17, 17–85 (2017).
Ligsay, A., Telle, O. & Paul, R. Challenges to mitigating the urban health burden of mosquito-borne diseases in the face of climate change. Int. J. Environ. Res. Public Health. 18, 5035 (2021).
Yi, H. et al. Effects of global warming on mosquitoes & mosquito-borne diseases and the new strategies for mosquito control. Entomol. Res. 44, 215–235. https://doi.org/10.1111/1748-5967.12084 (2014).
Seoul Metropolitan Government. Seoul Registered Population Statistics. https://data.seoul.go.kr/dataList/419/S/2/datasetView.do (2025).
Mercer, D. R., Brown, E. J. & Sheeley, S. L. Mosquito (Diptera: Culicidae) development within microhabitats of an Iowa wetland. J. Med. Entomol. 42, 685–693. https://doi.org/10.1093/jmedent/42.4.685 (2005).
Buckner, E. A., Blackmore, M. S., Golladay, S. W. & Covich, A. P. Weather and landscape factors associated with adult mosquito abundance in Southwestern georgia, U.S.A. J. Vector Ecol. 36, 269–278. https://doi.org/10.1111/j.1948-7134.2011.00167.x (2011).
Landau, K. I. & van Leeuwen, W. J. D. Fine scale Spatial urban land cover factors associated with adult mosquito abundance and risk in Tucson, Arizona. J. Vector Ecol. 37, 407–418. https://doi.org/10.1111/j.1948-7134.2012.00245.x (2012).
Gubler, D. J. et al. Climate variability and change in the united states: potential impacts on vector-and rodent-borne diseases. Environ. Health Perspect. 109 (Suppl 2), 223–233 (2001).
Nyamukondiwa, C., Weldon, C. W., Chown, S. L., le Roux, P. C. & Terblanche, J. S. Thermal biology, population fluctuations and implications of temperature extremes for the management of two globally significant insect pests. J. Insect. Physiol. 59, 1199–1211 (2013).
Carrington, L. B., Armijos, M. V., Lambrechts, L., Barker, C. M. & Scott, T. W. Effects of fluctuating daily temperatures at critical thermal extremes on Aedes aegypti life-history traits. PloS One. 8, e58824 (2013).
Chaves, L. & Kitron, U. Weather variability impacts on oviposition dynamics of the Southern house mosquito at intermediate time scales. Bull. Entomol. Res. 101, 633–641 (2011).
Valdez, L. D., Sibona, G. J., Diaz, L. A., Contigiani, M. & Condat, C. Effects of rainfall on culex mosquito population dynamics. J. Theor. Biol. 421, 28–38 (2017).
Chuang, T. W., Hildreth, M. B., Vanroekel, D. L. & Wimberly, M. C. Weather and land cover influences on mosquito populations in Sioux falls, South Dakota. J. Med. Entomol. 48, 669–679 (2011).
Yang, G. J., Brook, B. W., Whelan, P. I., Cleland, S. & Bradshaw, C. J. A. Endogenous and exogenous factors controlling temporal abundance patterns of tropical mosquitoes. Ecol. Appl. 18, 2028–2040. https://doi.org/10.1890/07-1209.1 (2008).
Capelli, G. et al. First report in Italy of the exotic mosquito species Aedes (Finlaya) koreicus, a potential vector of arboviruses and filariae. Parasites Vectors. 4, 188 (2011).
Sugihara, G. et al. Detecting causality in complex ecosystems. Science 338, 496–500. https://doi.org/10.1126/science.1227079 (2012).
Choi, W. I., Jung, J. B., Kim, M. J., Kwon, T. S. & Park, Y. S. Human-mediated forest transition shapes long-term occurrence patterns of pine caterpillar (Dendrolimus spectabilis) in the Korean Peninsula. J. Environ. Manage. 392, 126710. https://doi.org/10.1016/j.jenvman.2025.126710 (2025).
R Core Team. R. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing (Foundation for Statistical Computing, Vienna, Austria, 2024). https://www.R-project.org/.
Soetaert, K. plot3D: Plotting Multi-Dimensional Data. R package version 1.4.1. https://CRAN.R-project.org/package=plot3D (2024).
Wood, S. N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. Royal Stat. Society: Ser. B (Statistical Methodology). 73, 3–36 (2011).
Ye, H., Clark, A., Deyle, E. & Munch, S. rEDM: Applications of Empirical Dynamic Modeling from Time Series. R package version 0.7.5. https://github.com/ha0ye/rEDM (2024).
Acknowledgements
This work was supported by the National Research Foundation of Korea(NRF) grant funded by the Korea government (MSIT) (RS-2024-00456138) and by Korea Environment Industry & Technology Institute (KEITI) through Aquatic Ecosystem Conservation Research Program funded by Korea Ministry of Environment (MOE) (KEITI-2020003050003).
Author information
Authors and Affiliations
Contributions
D.-S., L.: Data curation, methodology, formal analysis, writing – original draft, writing – review and editing. Y.-S., P.: Conceptualization, methodology, funding acquisition, supervision, writing – original draft, writing – review and editing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
This article does not contain any studies with human participants or animals (vertebrates) performed by any of the authors.
Consent to participate
Informed consent was obtained from all individual participants included in the study.
Consent to publish
Consent was obtained from all individual participants included in the study.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Lee, DS., Park, YS. Effects of cumulative temperature and precipitation patterns on mosquito abundance in urban Seoul, South Korea. Sci Rep 15, 32790 (2025). https://doi.org/10.1038/s41598-025-17975-2
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-17975-2
