
Abstract
Tannic acid (TA) is the primary bioactive component in the gallnut (Galla chinensis) and has exhibited the anticancer effects. However, the mechanism of its anti-cancer activity in nasopharyngeal carcinoma (NPC) remains unclear. This research aims to explore the underlying mechanism of TA in the treatment of nasopharyngeal cancer using network pharmacology, molecular docking and experimental validation. Firstly, the targets of TA and NPC were predicted and collected through databases, and the intersection targets were identified. Subsequently, protein-protein interaction (PPI) network analysis, Gene Ontology (GO) enrichment, Kyoto Encyclopedia of Genes Genomes (KEGG) pathway enrichment analysis, molecular docking and molecular dynamics (MD) simulation were conducted to uncover the potential mechanisms of TA in treatment of NPC. Finally, in vitro experiments were utilized to verify the mechanism of TA with anticancer activity in NPC. The results of network pharmacology revealed 42 intersection targets between NPC-related targets and TA-related targets. The phosphoinositide 3-kinase (PI3K)/protein kinase B (AKT) signaling was identified as the main target pathway of TA against NPC. Additionally, molecular docking and MD simulation confirmed the closely binding affinities of TA with AKT1. Furthermore, the results of in vitro experiments demonstrated that TA exerts anticancer activity against NPC by targeting the PI3K/AKT signaling pathway, leading to the suppression of cell proliferation. TA is a promising therapeutic candidate for NPC through PI3K/AKT signaling pathway. These results provide insights into the clinical application of TA, particularly when considered in combination with other therapeutic modalities.
Similar content being viewed by others

Introduction
Nasopharyngeal carcinoma (NPC) is a fatal epithelial tumor originating from the nasopharyngeal mucosal membrane and is associated with a distinct geographical distribution in North Africa and Southeast Asia1,2. Three histological subtypes of NPC are commonly found: basaloid carcinoma, keratinizing squamous cell carcinoma, and non-keratinizing carcinoma3,4. The etiology of NPC is associated with Epstein-Barr virus infection, genetics, and environmental factors. Additionally, NPC is characterized by a higher incidence in males, with a male to female ratio of about 2.5:15. With changes in lifestyle and environment, increased awareness of the risk pathogenesis, rapid advancement in imaging techniques, and the optimization of chemoradiotherapy strategies, the incidence and mortality of NPC have gradually declined1. However, some patients with NPC will experience recurrence and may die from distant metastasis or locoregional relapse. The ideal therapy for recurrent NPC is platinum-based combination chemotherapy. Despite advances in radiotherapy, some patients show unsatisfactory results due to radio-resistance and tumor recurrence after irradiation6,7,8. Therefore, it is urgent to explore potential anti-NPC approaches with minimal adverse effects on patients.
Natural products have demonstrated great potential as anti-cancer agents9,10. Due to their high biological activity and abundant resources, natural products have gradually become an attractive focus of drug research. Rhus chinensis Mill is an herbal medicine first recorded in the Chinese classic Shan Hai Jing thousands of years ago, and it shows substantial preventative and therapeutic value for various diseases, such as diarrhea, dysentery, cancer, diabetes mellitus, across several East and Southeast Asia countries11. In particular, the species of gallnut (Galla Chinensis) from Rhus chinensis Mill is widely used in medicine. Moreover, recent studies have reported that Galla Chinensis exhibits strong antiviral, anticancer, antibacterial, antioxidant, and hepatoprotective activities12. Tannic acid (TA) is the primary bioactive component in the Galla chinensis, has multiple pharmacological applications in the medical field13. In recent years, several studies have reported that TA exhibits potential anticancer activity against various types of malignant cancers, such as lung, liver, pancreatic, and colon cancer14. Research has also demonstrated that TA has potential as a therapeutic agent for NPC. In 2000, Lee et al. showed that two components of tannins, eugeniflorins D (1) and D (2) derived from the active fractions of Eugenia uniflora L., significantly inhibited EBV DNA polymerase more effectively than the positive control15. Additionally, other types of tannins such as pentagalloylglucose and grape seed proanthocyanidins, have also been shown to suppress cell proliferation, induce apoptosis and reduce the metastatic capacity of NPC cells16,17. Furthermore, TA, used as a delivering material in a nano-vaccine consisting of Epstein-Barr virus nuclear antigen 1 and a bi-adjuvant of Mn2+ and cytosine-phosphate-guanine formulated with TA, could significantly inhibit the development of mimicry NPC18. However, since TA consists of hydrolysable tannins, the precise mechanism of its therapeutic effects in NPC remains inadequately elucidated.
Network pharmacology is a novel methodological approach that elucidates drug treatment for various diseases by exploring the underlying mechanism involving multiple components, targets and pathways19,20,21,22. Furthermore, network pharmacology constructs a multi-level network to investigate the relationship between potential herbal components and diseases from a holistic perspective23. A study utilizing network pharmacology and experimental validation has revealed the potential mechanism of Hedyotis diffusa Willd against rheumatoid arthritis24. The molecular docking strategy clarifies the potential biochemical process which the drug interacts with target molecules based on high-throughput screening database25. A study has used molecular docking and chemical-protein interaction networks to identify potential inhibitors against Nipah Virus26. Network pharmacology and molecular docking have been widely applied to accurately elucidate the pharmacologic activity of potential therapeutic agents. Another study, utilizing network pharmacology, molecular docking and experimental validation, revealed that the potential mechanism of Lanxangia tsaoko in the treatment of type 2 diabetes mellitus is associated with PI3K/Akt, MAPK, RAGE/AGE, HIF-1 and FoxO signaling pathways27.
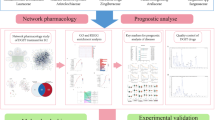
In the present study, we have employed a combination of network pharmacology, molecular docking and experiment approaches to demonstrate the active component of tannic acid in the treatment of NPC. This research provides experimental evidence for the development of gallnuts as a potential drug candidate for NPC. Figure 1 illustrates the workflow of this research.

The flow chart of this study.
Methods and materials
Reagents
Fetal bovine serum was procured from Sijiqing biotechnological industry (Hangzhou China). TA was provided by Sigma (Sigma, USA). High-glucose Dulbecco’s modified Eagle’s medium (DMEM) was obtained from Jinuo biotechnological industry (Hangzhou, China). Penicillin-streptomycin antibiotics were procured from Dalian Meilun biotechnological industry. The cell counting kit-8 (CCK-8) was purchased in APExBIO (APExBIO Inc, USA). The BCA protein concentration kit, RIPA lysis buffer, protease inhibitor and PI3K inhibitor-LY294002 were provided in Beyotime biotechnology (Shanghai, China). The primary antibodies for β-Actin, glyceraldehyde-3-phosphate dehydrogenase (GAPDH), phospho-phosphoinositide 3-kinase (PI3K), and phospho-protein kinase B (AKT) were obtained from Beyotime biotechnology (Shanghai, China). Antibodies for PI3K, and AKT were sourced from cell signaling technology (Massachusetts, USA). The protein ladders were purchased from Yamei biotechnology (Shanghai, China).
Prediction of NPC and TA targets
The NPC-related targets were identified using the keywords “Nasopharyngeal carcinoma” across four databases: TTD28, OMIM29, DisGeNET30, and GeneCards31.
The keyword “Tannic acid” was used to search for TA-related targets from the BATMAN-TCM32, HERB33, DrugCentral34, and HIT35 databases. Additionally, the SDF format of TA’s structure was used to predict targets via PharmMapper36. TA-related targets were mapped to gene names using UniProt. The intersection of TA and NPC targets represents the potential targets of TA against NPC.
Network construction and analysis
A compound-target-disease (C-T-D) network was constructed, where each node represents TA, potential targets, and NPC, respectively. Furthermore, the protein-protein interaction (PPI) network of potential targets was established by setting the minimum required interaction score to moderate confidence (0.400) in the STRING database37. Subsequently, CytoHubba, a plugin of Cytoscape software38, was used to identify hub nodes within the PPI network.
GO and KEGG enrichment analysis
Gene Ontology (GO) analysis was conducted using FunRich software39 equipped with the custom database (DAVID, version 6.8). Meanwhile, the Kyoto Encyclopedia of Genes and Genomes (KEGG) was performed using the Hiplot40,41,42.
Expression analysis of hub genes
Gene Expression Omnibus (GEO), a public functional genomics database, allows users to query and download high-throughput sequencing data, chips, and microarrays. The GSE53819 dataset was obtained from the GEO database and subsequently analyzed using the GEO2R tool (http://www.ncbi.nlm.nih.gov/geo/geo2r) to compare the expression levels of hub genes between NPC specimens and normal tissues.
Molecular docking
The intermolecular affinities between TA and key targets were validated by AutoDock Vina43. Firstly, the 3D structures of the receptor were retrieved from the PDB database44. Secondly, the structure of TA was acquired from the ChemSpider database. The ligand’s energy was then optimized using Chem3D software. Thirdly, the ligand and receptor were processed with AutoDock Tools. The charges of the ligand were calculated, and its position and number of rotational bonds were defined. After the bound ligand and water molecules of the receptor were exercising, polar hydrogens were added to determine the charges. Finally, the docking process was performed within a 3D-grid box that encapsulated the active site. Docking outcomes were visualized and analyzed using PyMOL and LigPlot+ software. Pathania et al.’s approach was adopted to validate the docking protocol. Specifically, the bound ligand extracted from the receptor was re-docked into the binding site, and the ligand conformations obtained from experiments and calculations were compared45.
Molecular dynamics simulations
The molecular dynamics (MD) technique was used to verify the stability of the protein-ligand complex, with the simulation performed using GROMACS (version 2020.6)46. The protein topology file was generated by AMBER99SB-ILDN force field, while the ligand topology file was created using the General AMBER Force Field (GAFF) with Sobtop47. Solvation was carried out using the SPC water cube model with a 1.0 nm distance. Then, Na+/Cl− ions were added to the system to neutralize its charge. For energy minimization, the steepest descent method was applied first, followed by the conjugate gradient method. The isothermal volume (NVT) and isothermal pressure (NPT) simulations were conducted separately for 2 ns at 300 K to equilibrate the simulated system to the desired temperature and pressure. Finally, the MD simulations were carried out for 100 ns.
Cell culture
Human NPC cell lines of 5–8 F and 6-10B were obtained from the American Type Culture Collection (USA). The cell culture conditions were described in our previous study48.
Cell viability assay
Cells were placed at a density of 1 × 104 cells per well in a 96-well plate containing DMEM medium. After 24 h incubation, the cells were treated to fresh DMEM medium with the different concentrations of TA (0, 10, 20, 40, 60, 80 µM) for 24–48 h. At the end of treatment, the medium was renewed with fresh DMEM containing 10 µL CCK-8 reagent and incubated for 2 h. Finally, the absorbance of the entire plate was measured using an enzyme-linked detector at 450 nm (Biotek Instruments, USA). Each treatment group was performed in six replicates, and the experiments were repeated three times.
Colony formation experiment
NPC cells were placed at a density with 800 cells per well in a 6-well plate. After cells adhesion for 24 h, TA was added at different doses (0, 10, 20, 40, 60 µM) for 48 h. Afterward, the culture medium was renewed with fresh medium, and the cells were incubated for an additional 14 days. Finally, the cells were fixed with 4% paraformaldehyde for 10 min at 4 °C, and dyed with 0.1% crystal violet reagent in 20 min. After removing the solution and allowing the cells to dry, images of each well were captured and analyzed using Image pro plus software.
Wound healing assay
Cells were seeded in 6-well cell plates and grown to 90% confluence. A 200 µL pipette tip was used to manually create the cell monolayer scratch. The cells were washed three times with PBS solution to clean excess detached cells. Different concentrations of TA (0, 10, 20, 40, 60, 80 µM) were added to the plates. Images of the scratch were captured using a microscopy (Biotek Instruments, USA) at 0, 24, and 48 h after TA treatment. Wound healing was quantified by Image Pro Plus software. The percentage of wound closure was calculated by subtracting the area at 0 h from the area at the specified point, and dividing by the 0 h width, and calculating the ratio of wound closure.
Western blotting
NPC cells were lysed in a protein extraction reagent containing a protease inhibitor. BCA concentration kit was utilized to standardize the protein concentration of the sample. Protein was separated by SDS-PAGE electrophoresis, transferred to PVDF membrane, and incubated with 3% skim milk powder for 1 h. The membrane was incubated with the primary antibody at 4 °C for 12 h, then, followed by incubation with the secondary antibody at 25 °C for 2 h. The PVDF membrane was exposure using a chemical imaging system (Azure, USA), and the grayscales of each gel were calculated by the software of Image pro plus.
Statistical analysis
All data were shown as mean ± SEM, and statistical analysis was performed using one-way ANOVA. All analyses were determined using SPSS software version 22 (SPSS Inc., Chicago, USA) and statistical graphs were generated by GraphPad Prism 8.0 Software (GraphPad Instruments, USA). Results were recognized as statistically significant at P values < 0.05.
Results
Network pharmacology-based analysis
Prediction of target proteins
The keyword “Nasopharyngeal carcinoma” was used to identify disease-related targets, resulting in the identification of 1095 NPC-related targets from GeneCards, DisGeNET, TTD, and OMIM. Subsequently, 160 TA-related targets were predicted from BATMAN-TCM, DrugCentral, HERB, HIT and Pharmmapper. After standardization, 42 intersection targets between NPC-related and TA-related targets were identified as potential targets for TA against NPC (Fig. 2A). Additionally, the C-T-D network was constructed, as shown in Fig. 2B.

Screening and network construction of potential targets. (A) Venn diagram of the intersection targets between tannic acid and nasopharyngeal carcinoma. (B) Compound-target-disease network. (C) PPI network of potential targets. (D) Hub genes screened from the PPI network by CytoHubba.
PPI network
A PPI network was constructed with 42 potential targets of TA against NPC. However, the ABO target was excluded due to a lack of interaction with other targets. As shown in Fig. 2C, the PPI network consisted of 41 genes and 287 interactions. Hub genes were filtered by CytoHubba, including AKT1, PTGS2, IGF1, HSP90AA1, HRAS, CCND1, MMP9, MAPK3, ALB, and SRC, which may be involved in the mechanism of TA against NPC. Hub nodes are depicted in Fig. 2D and detailed in Table S1.
GO and KEGG enrichment analysis
GO and KEGG analysis were performed on 42 potential targets using FunRich and Hiplot, respectively. The results of the GO analysis are displayed in Fig. 3A–C. Among them, cellular component-related items predominantly included cytosol, cytoplasm, mitochondrion, and perinuclear region of cytoplasm. These items are closely related to the metabolic shift and mitochondrial resetting of cancer stem cells in nasopharyngeal carcinoma49. The molecular function-related items were primarily associated with kinase activity, protein kinase activity, and ATP binding. TA may be a potential ATP inhibitor that binds to the active site of protein kinases (such as protein kinase B) for cancer treatment. Additionally, the biological process-related items were mainly linked to protein autophosphorylation, lipopolysaccharide-mediated signaling pathway, peptidyl-serine phosphorylation, and protein phosphorylation. Phosphorylation is a modification associated with carcinogenesis and cancer development that regulates cellular processes such as cell growth, proliferation, death, metabolism and signal transduction50. Previous report showed that lipopolysaccharide can regulate peroxide to affect the development of nasopharyngeal carcinoma51. The results of the KEGG are shown in Fig. 3D, where the PI3K-Akt signaling pathway is significantly enriched. PI3K-Akt signaling pathway plays a crucial role in cell migration of NPC52. As the significant pathway, the detailed signaling pathway diagram of the PI3K-Akt signaling pathway is displayed in Fig. S141,42.

GO and KEGG enrichment analysis of target intersections between tannic acid and nasopharyngeal carcinoma. (A) Cellular components. (B) Molecular functions. (C) Biological processes. (D) Bubble chart of KEGG pathways.
Expression levels of hub genes
The hub genes including AKT1, CCND1, IGF1, HSP90AA1 and HRAS were enriched in the PI3K-Akt signaling pathway. The expression levels of these genes were externally validated by the GEO database, and the expression boxplots were displayed in Fig. S2. The GEO2R analysis showed that the expression levels of AKT1, CCND1 and HRAS were significantly up-regulated, and IGF1 was significantly down-regulated in NPC compared with normal tissues. There were no significant differences in the HSP90AA1 expression level between NPC and normal tissues.
Molecular docking analysis
KEGG analysis implies that TA may be mediated by regulating the PI3K-Akt signaling pathway in NPC. AKT1, as a hub gene for TA against NPC, shows a significant difference in expression level between NPC and normal tissues. Therefore, we speculate that the underlying mechanism of TA in the treatment of NPC is related to AKT1. Subsequently, the AKT1 (PDB: 4GV1) was selected as a key target and validated by molecular docking. The experimental and docked conformation of the bound ligand indicates that the docking protocol has high reliability (Fig. S3). Theoretically, a ligand-receptor complex with a lower binding energy is considered to be a more stable configuration37. These docking results indicated that TA had a good affinity towards AKT1 with binding energies of − 9.9 kcal/mol. As is shown in Fig. 4A, B, in the interaction with AKT1, TA was bound to THR-160, GLY-162, GLU-228, ALA-230, ASP-274, ASP-292, THR-312 and ASP-439 by hydrogen bonds. These docking results indirectly corroborate the results of the KEGG analysis.

Analysis of molecular docking and molecular dynamics simulation. (A) The three-dimensional binding pattern between AKT1 protein and TA. (B) A two-dimensional interaction map between TA and AKT1. (C) RMSD of TA and AKT1 complex during a 100 ns molecular dynamics simulation.
Molecular dynamics assessment
MD simulations offer valuable insights into the dynamic stability of protein-ligand complexes. To validate the docking results, the MD simulations of the AKT1-TA complex were performed for 100 ns. The root means square deviation (RMSD) curves can dynamically reflect the fluctuations of protein and ligand conformations. As is shown in Fig. 4C, the AKT1-TA complex reached relative equilibrium after 60 ns. The RMSD results indicate that TA can stably bind to AKT1.
Experimental validation
TA suppresses NPC cells viability and cell morphology
Based on the results of network pharmacology and molecular docking, we further explored and verified that TA has potential therapeutic effects on NPC through in vitro experiments. To evaluate the influence of TA in NPC cells, the cell viability and morphology of 5–8 F and 6–10 B cells treated with various doses of TA were analyzed. The results exhibited that the cell viability of 5–8 F and 6–10 B cells was significantly inhibited by TA treatment. Specifically, the cell viability of 5–8 F cells decreased when treated with TA concentrations of 40 µM to 80 µM at 24 h, and the cell viability was obviously decreased at 48 h with the same concentration. The cell morphology of 5–8 F at 24 h and 48 h showed identical results with the cell viability, as shown in Fig. 5A–D. Besides, the cell viability of 6–10 B was inhibited by TA concentrations ranging from 40 µM to 80 µM at 24 h, and by 20 µM to 80 µM at 48 h. The cell morphology of 6–10 B at 24 h and 48 h is shown in Fig. 5E–H. These findings indicate that TA suppresses NPC cell viability with a time-dependent manner.

Tannic acid inhibits the cell viability and alters cell morphology of nasopharyngeal carcinoma cells. (A,C) Representative images of cell morphology in 5–8 F cells, scale bar = 1000 μm. (B,D) Cell viability after treatment with different doses of tannic acid for 24 h and 48 h in 5–8 F cells. (E,G) Representative images of cell morphology in 6-10B cells, scale bar = 1000 μm. (F,H) Cell viability after treatment with different doses of tannic acid for 24 h and 48 h in 6-10B cells. Data represent the mean ± SEM, ***P < 0.001.
TA decreases NPC cells colony formation
We further assessed colony-forming capacity of TA in NPC cells. The ability for colony formation in NPC cells was suppressed by TA in a dose-dependent status. Notably, the number of cell colonies gradually decreased in NPC cells treated with TA concentrations ranging from 20 to 60 µM (Fig. 6). This finding indicates that TA effectively inhibits the colony forming ability of NPC cells.

Tannic acid decreases the number of colony formations in nasopharyngeal carcinoma cells. (A) Representative images of colony formation in 5–8 F and 6-10B cells. (B) Quantification of the number of colony formations by tannic acid in 5–8 F and 6–10 B cells. Data represent the mean ± SEM, ***P < 0.001.
TA inhibits NPC cells wound closure
To explore whether the TA-induced growth inhibition of NPC cells was correlated with wound closure, a wound healing assay was completed to determine the growth inhibition ratio. The results revealed that TA obviously suppressed wound closure in NPC cells in a dose-dependent status. TA decreased wound healing in NPC cells at higher concentrations (40 to 60 µM) after 24 h of treatment. Notably, NPC cells inhibited wound closure even at lower doses of TA after 48 h treatment (Fig. 7).

Tannic acid inhibits wound closure in nasopharyngeal carcinoma cells. (A,C) Represented images of wound closure in 5–8 F and 6-10B cells, scale bar = 1000 μm. (B,D) Quantitative analysis of wound closure by tannic acid in 5–8 F and 6–10 B cells. Data represent the mean ± SEM, ***P < 0.001.
TA inhibits the PI3K/AKT signaling pathway in NPC cells
These in vitro experimental results suggest that TA has the inhibitory effect of cell viability, colony formation and wound closure in NPC cells. Additionally, the results of KEGG analysis, GEO analysis and molecular docking reveal that TA may modulate the PI3K/AKT signaling pathway in NPC. To further explore the mechanism of TA in NPC, we examined its effect on PI3K/AKT signaling pathway in NPC cells. The findings indicated that TA significantly reduced the protein expression ratio of p-AKT/AKT in NPC cells. However, TA only suppressed the protein expression level of p-P13K/PI3K at a concentration of 40 µM in NPC cells (Fig. 8). These results suggest that TA likely exerts its anti-cancer effect in NPC by inhibiting the PI3K/AKT signaling pathway. To further confirm the inhibitory role of TA on the PI3K/AKT signaling pathway in NPC cells, we pre-treated NPC cells with a PI3K inhibitor (LY294002, 10 μm) for 30 min before treating TA. Our results showed that the combination of TA and PI3K inhibitor significantly decreased cell viability and wound closure in NPC cells compared to the TA-only group at the same treatment time and dose (Fig. 9 and Fig. S4). In summary, our findings strongly support that TA inhibits PI3K/AKT signaling pathway, exerting an inhibitory effect of cell growth in NPC cells.

Tannic acid suppresses the PI3K/AKT signaling pathway in nasopharyngeal carcinoma cells. (A) Representative western blotting images of p-PI3K, PI3K, p-AKT, AKT, and β-Actin in 5–8 F and 6-10B cells. (B,C) Quantitative analysis of western blotting data in 5–8 F and 6-10B cells. Data represent the mean ± SEM, ***P < 0.001.

Tannic acid combined with PI3K inhibitor inhibits the growth of nasopharyngeal carcinoma cells. (A,B) Cell viability after treatment with different doses of tannic acid for 24 h and 48 h, combined with PI3K inhibitor. (C,E) Representative images of wound closure in 5–8 F and 6-10B cells. scale bar = 1000 μm. (D,F) Quantitative analysis of wound closure by tannic acid in 5–8 F and 6-10B cells. Data represent the mean ± SEM, ***P < 0.001.
Discussion
TA, the primary bioactive component in the gallnut (Galla chinensis), exhibits significant antioxidant and anti-cancer activities, making it a promising candidate for translational medicine. In this study, we employed a comprehensive approach combining network pharmacology, molecular docking, and experimental verification to demonstrate the potential of TA as a modulator for the treatment of NPC.
Natural products and their structural analogues have played a pivotal role in drug research, particularly in cancer treatment, attracting significant attention from researchers10,53. Gallnut (Galla chinensis), a natural product, has gained considerable recognition as a promising therapeutic agent for cancer. TA, the primary bioactive component of gallnut, has been extensively demonstrated for its anticancer activity of TA in both pharmaceutical and biological field14,54. New mechanisms of anticancer activity of TA have been reported, including its modulation of cancer cell metabolism, reduction of cancer cell production, and inhibition of epithelial-to-mesenchymal transition55. Nagesh K B Prashanth and colleagues have shown that TA distinctly indicated double roles by hindering lipogenic signaling and lipid metabolic pathways in prostate cancer cells56. Mhlanga Priscilla et al. have investigated that TA induces cell apoptosis through DNA fragmentation and oxidative stress in human hepatocellular carcinoma cells57. Furthermore, TA has been utilized in various applications, such as drug delivery systems, including nanoparticles and hydrogels for anticancer therapy58. Despite these findings, few studies have explored the anticancer effects of TA in NPC, highlighting the need for further investigation into its underlying anticancer mechanisms.
Based on network pharmacology analysis, we constructed the TA-NPC target network and identified 42 targeted intersections. Ten hub targets were recognized through network topology analysis in the PPI network, including AKT1, SRC, PTGS2, MMP9, MAPK3, IGF1, HSP90AA1, HRAS, CCND1, and ALB. Subsequently, GO and KEGG analysis revealed that the PI3K/AKT signaling pathway is a crucial target pathway of TA in treatment of NPC. The PI3K/AKT signaling pathway is related to a variety of cellular functions, which exerts vital roles in both normal physiological circumstances and pathological disorders, particularly contributing to the emergence and progression of tumors59. Numerous reports have verified that natural products target the PI3K/AKT signaling pathway in the cancer60. The PI3K/AKT pathway is closely linked to apoptosis, proliferation, invasion, metastasis, and resistance to radiotherapy and chemotherapy in NPC61. Aberrant activation of PI3K/AKT pathway is frequently elucidated in aggressive cancer, particularly the mutation of PIK3CA gene, which encodes the catalytic isoform of PI3K p110α. This mutation is commonly found in many malignant cancers and contributes to tumors oncogenesis, including in NPC62. In addition, the activation of PI3K/AKT pathway has been demonstrated to be involved in the etiology of NPC, especially in cases induced by Epstein-Barr virus63,64. These studies confirm that the aberrant activation of PI3K/AKT pathway promotes tumor-genesis in NPC. A range of anti-cancer drugs targeting the PI3K/AKT signaling pathway have been developed and shown to be effective in cancer that supported by the strong pre-clinical and clinical data65. Moreover, molecular docking results in our study exhibited that TA could closely dock with the target proteins of the PI3K/AKT pathways. These results suggested that TA may inhibit NPC by targeting multiple intersections and the PI3K/AKT signaling pathway.
Combined with these findings from network pharmacology and molecular docking, we performed in vitro experiments to confirm the anticancer activity of TA in NPC disease. Firstly, two NPC cell lines were treated with a range of TA doses, and the cell proliferation of NPC cells was considerably suppressed by TA. Chen et al. reported that TA demonstrated growth suppression in bladder cancer cells66. Furthermore, it has been indicated that TA inhibits cell proliferation through oxidative stress and apoptosis in human liver hepatocellular carcinoma cells57. Our results are consistent with these studies, indicating that TA has the potential to suppress the cell proliferation of NPC cells. Moreover, the data of network pharmacology predicted that TA may suppress cell proliferation of NPC cells by modulating the PI3K/AKT signaling pathway. Western blotting results indicated that TA strongly inhibited the PI3K/AKT pathway in NPC cells. The results from our study indicated that TA significantly decreased the protein level of PI3K/AKT signaling pathway in NPC cells. Besides, our data show that TA, when combined with a PI3K inhibitor, significantly suppresses cell proliferation than TA-only treatment in NPC cells. The PI3K/AKT signaling pathway is a crucial and stable signal transduction in eukaryotic cells that regulates cell growth, cell survival, and cell cycle progression. Dysregulation of the PI3K/AKT signaling transduction can promote cancer progression, and it is one of the most commonly modulated pathways in the disease of cancer, often linked to resistance to anticancer treatments67,68. Recently, Yang et al. reported that TA inhibited bisphenol A-induced cell pyroptosis by regulating the PTEN/PI3K/AKT pathway69. Furthermore, Chen et al. demonstrated that TA remarkably suppressed the phosphorylation level of AKT to inhibit cell proliferation in bladder cancer cells66. Taking together, these findings demonstrate that TA exerts its anticancer role in NPC cells by suppressing cell proliferation and modulating the PI3K/AKT signaling pathway.
Conclusion
In this study, we preliminarily predicted the intersection targets and the main target pathway of TA for the therapy of NPC through network pharmacology and molecular docking. Furthermore, our results demonstrated that TA inhibited cell proliferation and suppressed the PI3K/AKT signaling pathway through in vitro experiments, confirming the antitumor pharmacological effects of TA. This research has the potential to advance a promising pharmaceutical solution for treating NPC.
Data availability
The data and supportive information are available in the article.
References
Chen, Y. P. et al. Nasopharyngeal carcinoma. Lancet 394, 64–80. https://doi.org/10.1016/S0140-6736(19)30956-0 (2019).
Bossi, P. et al. Nasopharyngeal carcinoma: ESMO-EURACAN clinical practice guidelines for diagnosis, treatment and follow-up(dagger). Ann. Oncol. 32, 452–465. https://doi.org/10.1016/j.annonc.2020.12.007 (2021).
Lee, A. W. M. et al. Management of locally recurrent nasopharyngeal carcinoma. Cancer Treat. Rev. 79, 101890. https://doi.org/10.1016/j.ctrv.2019.101890 (2019).
Bossi, P., Gurizzan, C. & Chan, A. Immunotherapy for nasopharyngeal carcinoma: the earlier the better. JAMA 330, 1954–1955. https://doi.org/10.1001/jama.2023.22465 (2023).
Tang, L. L. et al. The Chinese Society of Clinical Oncology (CSCO) clinical guidelines for the diagnosis and treatment of nasopharyngeal carcinoma. Cancer Commun. (Lond). 41, 1195–1227. https://doi.org/10.1002/cac2.12218 (2021).
Poh, S. S. & Wee, J. T. Nasopharyngeal carcinoma treatment paradigm after HK0501—a potential way forward. Cancer Commun. (Lond) 42, 801–803. https://doi.org/10.1002/cac2.12339 (2022).
Juarez-Vignon Whaley, J. J. et al. Recurrent/metastatic nasopharyngeal carcinoma treatment from present to future: where are we and where are we heading? Curr. Treat. Options Oncol. 24, 1138–1166. https://doi.org/10.1007/s11864-023-01101-3 (2023).
Guan, S., Wei, J., Huang, L. & Wu, L. Chemotherapy and chemo-resistance in nasopharyngeal carcinoma. Eur. J. Med. Chem. 207, 112758. https://doi.org/10.1016/j.ejmech.2020.112758 (2020).
Zhang, L. et al. The strategies and techniques of drug discovery from natural products. Pharmacol. Ther. 216, 107686. https://doi.org/10.1016/j.pharmthera.2020.107686 (2020).
Atanasov, A. G., Zotchev, S. B., Dirsch, V. M. & Supuran, C. T. International Natural Product Sciences Taskforce. Natural products in drug discovery: advances and opportunities. Nat. Rev. Drug Discov. 20, 200–216 (2021). https://doi.org/10.1038/s41573-020-00114-z
Li, M. et al. A comprehensive review on ethnobotanical, phytochemical and pharmacological aspects of Rhus chinensis Mill. J. Ethnopharmacol. 293, 115288. https://doi.org/10.1016/j.jep.2022.115288 (2022).
Djakpo, O. & Yao, W. Rhus chinensis and Galla Chinensis–folklore to modern evidence: review. Phytother. Res. 24, 1739–1747. https://doi.org/10.1002/ptr.3215 (2010).
Cai, Y. et al. Recent advances in anticancer activities and drug delivery systems of tannins. Med. Res. Rev. 37, 665–701. https://doi.org/10.1002/med.21422 (2017).
Youness, R. A. et al. Recent advances in tannic acid (gallotannin) anticancer activities and drug delivery systems for efficacy improvement; a comprehensive review. Molecules https://doi.org/10.3390/molecules26051486 (2021).
Lee, M. H., Chiou, J. F., Yen, K. Y. & Yang, L. L. EBV DNA polymerase inhibition of tannins from Eugenia uniflora. Cancer Lett. 154, 131–136. https://doi.org/10.1016/s0304-3835(00)00353-0 (2000).
Yao, K. et al. Grape seed proanthocyanidins induce apoptosis through the mitochondrial pathway in nasopharyngeal carcinoma CNE-2 cells. Oncol. Rep. 36, 771–778. https://doi.org/10.3892/or.2016.4855 (2016).
Fan, C. W. et al. Pentagalloylglucose suppresses the growth and migration of human nasopharyngeal cancer cells via the GSK3beta/beta-catenin pathway in vitro and in vivo. Phytomedicine 102, 154192. https://doi.org/10.1016/j.phymed.2022.154192 (2022).
Wen, Z. et al. Nanovaccines fostering tertiary lymphoid structure to attack mimicry nasopharyngeal carcinoma. ACS Nano 17, 7194–7206. https://doi.org/10.1021/acsnano.2c09619 (2023).
Nogales, C. et al. Network pharmacology: curing causal mechanisms instead of treating symptoms. Trends Pharmacol. Sci. 43, 136–150. https://doi.org/10.1016/j.tips.2021.11.004 (2022).
Yang, J. et al. Network pharmacology, molecular docking and experimental study of CEP in nasopharyngeal carcinoma. J. Ethnopharmacol. 323, 117667. https://doi.org/10.1016/j.jep.2023.117667 (2024).
Li, S. Network pharmacology evaluation method guidance-draft. World J. Tradit Chin. Med. 7, 146–154 (2021).
Wang, Z. Y., Wang, X., Zhang, D. Y., Hu, Y. J. & Li, S. Traditional Chinese medicine network pharmacology: development in new era under guidance of network pharmacology evaluation method guidance. China J. Chin. Mater. Med. 47, 7–17. https://doi.org/10.19540/j.cnki.cjcmm.20210914.702 (2022).
Zhang, P. et al. Network pharmacology: towards the artificial intelligence-based precision traditional Chinese medicine. Brief. Bioinform. https://doi.org/10.1093/bib/bbad518 (2023).
Deng, H. et al. Network pharmacology and experimental validation to identify the potential mechanism of Hedyotis diffusa Willd against rheumatoid arthritis. Sci. Rep. 13, 1425. https://doi.org/10.1038/s41598-022-25579-3 (2023).
Li, T., Guo, R., Zong, Q. & Ling, G. Application of molecular docking in elaborating molecular mechanisms and interactions of supramolecular cyclodextrin. Carbohydr. Polym. 276, 118644. https://doi.org/10.1016/j.carbpol.2021.118644 (2022).
Pathania, S., Randhawa, V. & Kumar, M. Identifying potential entry inhibitors for emerging Nipah virus by molecular docking and chemical-protein interaction network. J. Biomol. Struct. Dyn. 38, 5108–5125. https://doi.org/10.1080/07391102.2019.1696705 (2020).
Wang, Z. et al. Network pharmacology, molecular docking and experimental validation to elucidate the anti-T2DM mechanism of Lanxangia Tsaoko. Fitoterapia 178, 106117. https://doi.org/10.1016/j.fitote.2024.106117 (2024).
Zhou, Y. et al. Therapeutic target database describing target druggability information. Nucleic Acids Res. 52, D1465–D1477. https://doi.org/10.1093/nar/gkad751 (2024).
Hamosh, A., Scott, A. F., Amberger, J. S., Bocchini, C. A. & McKusick, V. A. Online mendelian inheritance in man (OMIM), a knowledgebase of human genes and genetic disorders. Nucleic Acids Res. 33, D514–D517. https://doi.org/10.1093/nar/gki033 (2005).
Pinero, J., Sauch, J., Sanz, F. & Furlong, L. I. The DisGeNET cytoscape app: exploring and visualizing disease genomics data. Comput. Struct. Biotechnol. J. 19, 2960–2967. https://doi.org/10.1016/j.csbj.2021.05.015 (2021).
Fishilevich, S. et al. Genic insights from integrated human proteomics in GeneCards. Database (Oxford). https://doi.org/10.1093/database/baw030 (2016).
Kong, X. et al. BATMAN-TCM 2.0: an enhanced integrative database for known and predicted interactions between traditional Chinese medicine ingredients and target proteins. Nucleic Acids Res. 52, D1110–D1120. https://doi.org/10.1093/nar/gkad926 (2024).
Fang, S. et al. HERB: a high-throughput experiment- and reference-guided database of traditional Chinese medicine. Nucleic Acids Res. 49, D1197–D1206. https://doi.org/10.1093/nar/gkaa1063 (2021).
Ursu, O. et al. DrugCentral: online drug compendium. Nucleic Acids Res. 45, D932–D939. https://doi.org/10.1093/nar/gkw993 (2017).
Yan, D. et al. HIT 2.0: an enhanced platform for herbal ingredients’ targets. Nucleic Acids Res. 50, D1238–D1243. https://doi.org/10.1093/nar/gkab1011 (2022).
Liu, X. et al. PharmMapper server: a web server for potential drug target identification using pharmacophore mapping approach. Nucleic Acids Res. 38, W609–W614. https://doi.org/10.1093/nar/gkq300 (2010).
Yan, P. et al. Network pharmacology combined with metabolomics and lipidomics to reveal the hypolipidemic mechanism of Alismatis rhizoma in hyperlipidemic mice. Food Funct. 13, 4714–4733. https://doi.org/10.1039/d1fo04386b (2022).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. https://doi.org/10.1101/gr.1239303 (2003).
Fonseka, P., Pathan, M., Chitti, S. V., Kang, T. & Mathivanan, S. FunRich enables enrichment analysis of OMICs datasets. J. Mol. Biol. 433, 166747. https://doi.org/10.1016/j.jmb.2020.166747 (2021).
Li, J. et al. Hiplot: a comprehensive and easy-to-use web service for boosting publication-ready biomedical data visualization. Brief. Bioinform. https://doi.org/10.1093/bib/bbac261 (2022).
Kanehisa, M. & Goto, S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 28, 27–30. https://doi.org/10.1093/nar/28.1.27 (2000).
Kanehisa, M., Sato, Y., Kawashima, M., Furumichi, M. & Tanabe, M. KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 44, D457–D462. https://doi.org/10.1093/nar/gkv1070 (2016).
Eberhardt, J., Santos-Martins, D., Tillack, A. F. & Forli, S. AutoDock Vina 1.2.0: new docking methods, expanded force field, and Python Bindings. J. Chem. Inf. Model. 61, 3891–3898. https://doi.org/10.1021/acs.jcim.1c00203 (2021).
Rose, P. W. et al. The RCSB protein data bank: integrative view of protein, gene and 3D structural information. Nucleic Acids Res. 45, D271–D281. https://doi.org/10.1093/nar/gkw1000 (2017).
Pathania, S., Randhawa, V. & Bagler, G. Prospecting for novel plant-derived molecules of Rauvolfia serpentina as inhibitors of Aldose Reductase, a potent drug target for diabetes and its complications. Plos One 8, e61327. https://doi.org/10.1371/journal.pone.0061327 (2013).
Abraham, M. J. et al. High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 1. GROMACS, 19–25 (2015).
Lu, T. Sobtop, Version 1.0(dev5) (2024). http://sobereva.com/soft/Sobtop
Wang, M. et al. Tannic acid attenuates intestinal oxidative damage by improving antioxidant capacity and intestinal barrier in weaned piglets and IPEC-J2 cells. Front. Nutr. 9, 1012207. https://doi.org/10.3389/fnut.2022.1012207 (2022).
Shen, Y. A., Wang, C. Y., Hsieh, Y. T., Chen, Y. J. & Wei, Y. H. Metabolic reprogramming orchestrates cancer stem cell properties in nasopharyngeal carcinoma. Cell. Cycle 14, 86–98. https://doi.org/10.4161/15384101.2014.974419 (2015).
Capaci, V. et al. Phospho-DIGE identified phosphoproteins involved in pathways related to tumour growth in endometrial cancer. Int. J. Mol. Sci. https://doi.org/10.3390/ijms241511987 (2023).
Hou, T. et al. Lipopolysaccharide (LPS) regulates peroxide to affect the development of nasopharyngeal carcinoma by suppressing Act1 expression. Int. J. Clin. Exp. Pathol. 10, 6139–6145 (2017).
Fan, X. et al. YBX3 mediates the metastasis of nasopharyngeal carcinoma via PI3K/AKT signaling. Front. Oncol. 11, 617621. https://doi.org/10.3389/fonc.2021.617621 (2021).
Kashyap, D. et al. Natural product-based nanoformulations for cancer therapy: opportunities and challenges. Semin. Cancer Biol. 69, 5–23. https://doi.org/10.1016/j.semcancer.2019.08.014 (2021).
Ghasemian, M. et al. Recent progress in tannic acid based approaches as a natural polyphenolic biomaterial for cancer therapy: a review. Biomed. Pharmacother. 166, 115328. https://doi.org/10.1016/j.biopha.2023.115328 (2023).
Kleszcz, R., Majchrzak-Celinska, A. & Baer-Dubowska, W. Tannins in cancer prevention and therapy. Br. J. Pharmacol. https://doi.org/10.1111/bph.16224 (2023).
Nagesh, P. K. B. et al. Tannic acid inhibits lipid metabolism and induce ROS in prostate cancer cells. Sci. Rep. 10, 980. https://doi.org/10.1038/s41598-020-57932-9 (2020).
Mhlanga, P. et al. Mechanistic insights into oxidative stress and apoptosis mediated by tannic acid in human liver hepatocellular carcinoma cells. Int. J. Mol. Sci. https://doi.org/10.3390/ijms20246145 (2019).
Chowdhury, P. et al. Tannic acid-inspired paclitaxel nanoparticles for enhanced anticancer effects in breast cancer cells. J. Colloid Interface Sci. 535, 133–148. https://doi.org/10.1016/j.jcis.2018.09.072 (2019).
He, Y. et al. Targeting PI3K/Akt signal transduction for cancer therapy. Signal. Transduct. Target. Ther. 6, 425. https://doi.org/10.1038/s41392-021-00828-5 (2021).
Tewari, D., Patni, P., Bishayee, A., Sah, A. N. & Bishayee, A. Natural products targeting the PI3K-Akt-mTOR signaling pathway in cancer: a novel therapeutic strategy. Semin. Cancer Biol. 80, 1–17. https://doi.org/10.1016/j.semcancer.2019.12.008 (2022).
Liu, P., Cheng, H., Roberts, T. M. & Zhao, J. J. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat. Rev. Drug Discov. 8, 627–644. https://doi.org/10.1038/nrd2926 (2009).
Wen, L. et al. Influences of S100A8 and S100A9 on proliferation of nasopharyngeal carcinoma cells through PI3K/Akt signaling pathway. Biomed. Res. Int. 2021, 9917365. https://doi.org/10.1155/2021/9917365 (2021).
Li, H. L., Deng, N. H., He, X. S. & Li, Y. H. Small biomarkers with massive impacts: PI3K/AKT/mTOR signalling and microRNA crosstalk regulate nasopharyngeal carcinoma. Biomark. Res. 10, 52. https://doi.org/10.1186/s40364-022-00397-x (2022).
Fruman, D. A. & Rommel, C. PI3K and cancer: lessons, challenges and opportunities. Nat. Rev. Drug Discov. 13, 140–156. https://doi.org/10.1038/nrd4204 (2014).
Ediriweera, M. K., Tennekoon, K. H. & Samarakoon, S. R. Role of the PI3K/AKT/mTOR signaling pathway in ovarian cancer: Biological and therapeutic significance. Semin. Cancer Biol. 59, 147–160. https://doi.org/10.1016/j.semcancer.2019.05.012 (2019).
Chen, M. C. et al. Tannic acid attenuate AKT phosphorylation to inhibit UMUC3 bladder cancer cell proliferation. Mol. Cell. Biochem. 477, 2863–2869. https://doi.org/10.1007/s11010-022-04454-9 (2022).
Glaviano, A. et al. PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Mol. Cancer 22, 138. https://doi.org/10.1186/s12943-023-01827-6 (2023).
Alzahrani, A. S. PI3K/Akt/mTOR inhibitors in cancer: at the bench and bedside. Semin. Cancer Biol. 59, 125–132. https://doi.org/10.1016/j.semcancer.2019.07.009 (2019).
Yang, X., Zhou, Y., Yu, T., Li, K. & Xu, S. TAN (tannic acid) inhibits BPA-induced pyroptosis of L8824 (grass carp hepatocytes) by regulating PTEN/PI3K/AKT pathway. Fish. Shellfish Immunol. 146, 109384. https://doi.org/10.1016/j.fsi.2024.109384 (2024).
Acknowledgements
We would like to acknowledge the help of Professor Jinyue Hu, pharmacist-in-charge Ying Zeng, and research assistant Manli Jiang, Xueting Liu in this manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (82203349), the Hunan Provincial Department of Education funded research projects (22C0225 and 23B0438), the Changsha Natural Science Foundation (kq2403173), the Project of Changsha Central Hospital (YNKY202202, YNKY202301 and YNKY202305).
Author information
Authors and Affiliations
Contributions
M.W. W and P. Y performed the experiments and wrote the main manuscript text. L. M. H designed the experiments and revised the manuscript. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, M., He, L. & Yan, P. Integrated network pharmacology, molecular docking and experimental validation to investigate the mechanism of tannic acid in nasopharyngeal cancer. Sci Rep 15, 5645 (2025). https://doi.org/10.1038/s41598-025-90211-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-90211-z
