
Abstract
Pomegranate (Punica granatum L.) is a highly diverse species with significant variation in morphological and pomological traits. Characterizing this diversity is essential for improving breeding strategies, particularly to enhance fruit quality, yield, and consumer acceptance. This study used statistical tools, including correlation matrix analysis (CMA), principal component analysis (PCA), multiple regression analysis (MRA), and heat map analysis (HMA) to explore trait clustering and genotype differentiation. PCA revealed that the first three components accounted for 31.54% of the total variation. PC1 (11.66%) was driven by traits such as fruit length (0.94), fruit diameter (0.93), fruit weight (0.93), and fruit peel weight (0.93). Heat map analysis grouped variables into four clusters, with traits like total soluble solids (7–22%, mean: 14.72% ± 2.99) and 100-aril fresh weight (19.34–49.46 g, mean: 32.51 g ± 7.93) being critical for marketability. Genotypes were divided into four groups, with subgroup D2 comprising ‘Karimabad-6’, ‘Karimabad-5’, ‘Karimabad-3’, ‘Karimabad-2’, ‘Karimabad-4’, ‘Sangan-2’, ‘Padik-4’, ‘Karimabad-8’, ‘Karimabad-7’, ‘Sangan-1’, ‘Padik-11’, ‘Padik-3’, ‘Padik-2’, ‘Sangan-4’, ‘Sangan-3’, ‘Padik-12’, ‘Karimabad-1’, ‘Sangan-9’, ‘Sangan-8’, ‘Padik-1’, and ‘Daman-9’. MRA identified significant correlations for key traits: fruit weight showed a positive correlation with fruit diameter (β = 0.66, p < 0.00) and fruit length (β = 0.32, p < 0.01), while fruit peel weight showed positive correlations with fruit weight (β = 0.92, p < 0.00) and fruit peel thickness (β = 0.12, p < 0.00). Total soluble solids showed a positive correlation with 100-aril fresh weight (β = 0.80, p < 0.00). These correlations, which were determined to be statistically significant by MRA, are supported by CMA. In the PCA biplot analysis, ‘Daman-2’, ‘Daman-5’, ‘Daman-7’, ‘Karimabad-4’, ‘Padik-9’, ‘Sangan-2’, and ‘Sangan-4’ genotypes were identified as outliers with extreme combinations of fruit traits, falling outside the 95% confidence ellipse, suggesting their potential for breeding programs targeting unique features. This study emphasizes the importance of fruit-related traits, including size, weight, and soluble solids, in genotype differentiation and marketability. Heat map and PCA analyses provided a comprehensive framework for clustering variables and genotypes, identifying actionable targets for breeding. Notably, ‘Sangan-1’, ‘Sangan-2’, ‘Karimabad-1’, ‘Karimabad-2’, ‘Karimabad-3’, and ‘Karimabad-4’ were identified as very soft-seeded genotypes, highlighting their potential for consumer-preferred traits. These findings contribute to improving pomegranate breeding strategies by integrating genetic diversity with agronomic and market demands.
Similar content being viewed by others


Introduction
The pomegranate (Punica granatum L.), which has an important historical and cultural heritage, is one of the oldest cultivated crops known due to its various nutritional, medicinal, and ornamental qualities1. Believed to have originated in the regions of Iran and Central Asia, the fruit has gradually spread to various parts of the world, including the Mediterranean basin, South Asia, and the Americas, driven by ancient trade networks and human migration2. Its remarkable adaptability to different environmental conditions, especially arid and semi-arid climates, underscores its capacity to withstand abiotic challenges such as drought and soil salinity3,4.
According to the latest FAOSTAT data for 2023, the global production of pomegranates is led by Iran, with over 1 million tons produced annually, accounting for roughly a third of total global production. India follows with approximately 850,000 tons, and Türkiye comes in third with more than 500,000 tons. The United States (mainly from California) produces around 144,000 tons, while China rounds out the top five with about 130,000 tons5.
In recent years, pomegranate has gained widespread global recognition, largely due to its rich composition of bioactive compounds, such as polyphenols, flavonoids, and tannins6. These compounds are associated with numerous health-promoting effects, including powerful antioxidative and anti-inflammatory activities, as well as potential roles in preventing chronic diseases like cardiovascular disorders and certain types of cancer7,8. This growing popularity reflects not only its functional health benefits but also its cultural and economic significance in both traditional and modern contexts.
The diversity of pomegranates is remarkable, exhibiting variations in traits such as fruit size, aril color, juice yield, and seed texture9. Among these, seed hardness plays a critical role in consumer preference and marketability. Plant trait determination and genetic characterization studies are important in establishing plant breeding programs10,11. Current breeding efforts are focused on overcoming the challenges posed by hard-seeded cultivars, which currently dominate the commercial market, by developing softer-seeded varieties that enhance both palatability and nutritional benefits12. Additionally, the categorization of pomegranate genotypes into sweet, sweet-sour, and sour, as well as their classification based on ripening seasons and usage purposes (e.g., table or juice varieties), highlights the phenotypic and genetic diversity within the species13,14.
Iran is a center for pomegranate diversity, providing a natural research area to study the genetic variations of this plant15. The country’s distinctive climatic conditions, notably its vast arid and semi-arid regions, facilitate the cultivation of a wide array of genotypes, from wild variants to highly domesticated landraces16. These populations display notable phenotypic variation, which can be observed in traits such as ripening times, aril sweetness, and resistance to both biotic and abiotic stressors. Conducting morphological and pomological analyses of these genotypes is essential for comprehending and preserving this genetic diversity, which is increasingly threatened by habitat degradation and genetic erosion due to the dominance of commercial monocropping systems1,17.
The characterization of pomegranate genetic resources through morphological traits such as fruit shape, aril color, and seed texture, in combination with statistical methods like correlation matrix analysis, principal component analysis, and heat map analysis, enables accurate classification and selection of parental lines for breeding programs18. This information is crucial for meeting market demands that increasingly emphasize qualities such as soft seeds, high juice content, and balanced flavor profiles. For example, globally renowned cultivars such as ‘Hicaznar’ (Türkiye), ‘Mollar de Elche’ (Spain), and ‘Wonderful’ (USA) have achieved their prominence through breeding strategies that effectively employed these analytical approaches19,20.
Studies on the genetic diversity of wild pomegranates can reveal various aspects of the domestication process and the reasons behind their diversification21. Despite its potential, research on the genetic resources of this wild plant species is relatively scarce1,18,22. Therefore, the objective of this study is to identify the phenotypic variation among native pomegranate genotypes collected from four different regions in the Sistan-va-Baluchestan province of Iran. Additionally, this study primarily aims to identify soft-seeded genotypes, with a subsequent focus on detecting promising genotypes characterized by superior fruit quality.
Materials and methods
Plant material
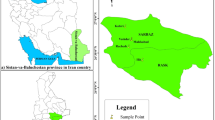
Morphological and pomological diversity of 47 native pomegranate genotypes was evaluated to introduce superior selections in four areas of Sistan-va-Baluchestan province, Iran. Geographic locations of collection sites of the studied genotypes are shown in Fig. 1. For accurate sampling, a minimum distance of 200 m was maintained between genotypes of each area so that clone samples were not collected. The formal identification of the specimens was performed by Prof. Dr. Ali Khadivi. A herbarium voucher specimen with sediment number PG-2213 has been donated to the public available herbarium of the Faculty of Agriculture and Natural Resources of Arak University, Iran.

Geographic locations of collection sites of the studied genotypes. The symbols represent the genotypes of each area, including Daman (D), Karimabad (K), Padik (P), and Sangan (S).
Morphological and pomological evaluations
64 morphological and pomological traits were used to evaluate phenotypic diversity and to select superior genotypes (Table 1). A total of 20 adult leaves and 20 mature fruits per genotype were randomly selected and harvested. The traits related to the dimensions of leaf, fruit, aril, and seed, were measured using a digital caliper (Loyka B5110-150-IP54 protected, China). A digital scale with an accuracy of 0.01 g was used to measure the weight of fruit, peel, and aril (JNB5002, JOANLAB, Zhejiang, China). The qualitative traits (Table 2) were visually examined and coded using the method introduced by Mars and Marrakchi23. The total soluble solids (TSS) were determined using a refractometer (pocket PAL-1 ATAGO Corporation, Tokyo, Japan) and expressed in ˚Brix.
Statistical analysis
Analysis of variance (One-way ANOVA, p < 0.05) was performed to evaluate the variation among genotypes based on the traits measured using JMP® Pro 17 software24. Pearson (r) correlation coefficients were used to determine the relationship between the recorded traits using Origin Pro® 2024b software25. Principal component analysis (PCA) was applied using Pro® 2024b software to identify the key traits influencing genotype grouping. To enhance the interpretability of the components, the Varimax rotation with Kaiser Normalization method was employed. This approach clarified the relationships between components, ensuring a more comprehensible and meaningful analysis. Heat map analysis based on Ward’s method and Euclidean distance coefficients using Origin Pro® 2024b software was used to classify genotypes and variables. The first and second principal components (PC1/PC2) were used to draw a two-dimensional biplot by determining the distribution of genotypes and quantitative variables using Origin Pro® 2024b software. Moreover, fruit-related traits were considered as dependent variables, and the characteristics affecting these characters were determined using multiple regression analysis (MRA). The MRA was conducted using ‘‘stepwise’’ method of ‘‘linear regression analysis’’ option of SPSS® (SPSS Inc., Chicago, IL, USA) statistics26.
Results and discussion
Descriptive statistics among genotypes
Descriptive statistics for the pomological and morphological traits utilized in the studied pomegranate genotypes are presented in detail in Table 1. Goodarzi et al.27, Mostafa et al.28, Elwakil et al.29, and Khadivi et al.30 reported that certain variables did not show significant differences among the genotypes (CV = 0.00%) in their studies, leading them to exclude these variables from their analyses. As a result, since two variables (leaf serration presence and fruit peel cracking presence) did not show a significant difference (CV = 0.00%) among the genotypes, the evaluations were made based on 62 variables. In this context, one-way ANOVA (p < 0.05) revealed significant variations among the assessed genotypes. The highest variation was observed in thorn length on annual shoot (175.59%), pacifier presence on fruit (164.08%), fruit peel necrotic presence (136.02%), thorn thickness on annual shoot (124.50%), and thorn presence on shoot (124.00%). In contrast, the lowest variation was recorded in sepal length (14.46%), seed length (14.06%), seed width (13.44%), sepal number (13.11%), and aril length (10.12%). Notably, 52 out of 62 variables (representing 83.87% in total) had coefficients of variation (CVs) greater than 20.00%. Our findings are consistent with a similar study conducted on pomegranate in Iran, which reported a CV of 84.50%31. This value indicates a high degree of variation among the examined genotypes32.
Leaf length ranged from 35.98 mm to 95.99 mm. Leaf width varied between 11.68 mm and 29.59 mm. Petiole length changed from 3.89 mm to 10.65 mm, while petiole diameter ranged from 0.71 mm to 1.63 mm. The thorn length on annual shoot ranged from 0 mm to 24 mm, while thorn thickness on annual shoot varied between 0 mm and 1.28 mm. Fruit length changed from 40.94 mm to 94.45 mm, whereas fruit diameter ranged from 44.44 mm to 102.7 mm. The sepal number varied between 4 and 8, and sepal length ranged from 9.88 mm to 18.35 mm. The sepal base width changed from 5.34 mm to 12.85 mm. Additionally, fruit calyx length ranged from 11.55 mm to 27.41 mm, and the fruit calyx diameter varied between 11.2 mm and 32.69 mm. Fruit weight changed from 41.95 g to 452.61 g, with fruit stalk length ranging from 5.38 mm to 61.69 mm. Similarly, fruit peel thickness varied between 1.82 mm and 6.35 mm, and fruit peel weight ranged from 22.49 g to 201.67 g. For 100-aril fresh weight, the values ranged from 19.34 g to 49.46 g, while aril length varied between 9.42 mm and 14.54 mm. aril width changed from 4.32 mm to 9.95 mm, and seed length ranged from 4.47 mm to 9.94 mm. Finally, seed width varied between 2.16 mm and 3.85 mm. Lastly, the total soluble solids content ranged from 7 to 22%.
A study conducted in Türkiye reported that fruit weight ranged from 69.9 to 795.3 g19. Khadivi et al.18 reported that leaf length ranged from 29.21 to 84.90 mm, leaf width from 8.69 to 25.78 mm, petiole length from 2.24 to 9.51 mm, fruit length from 28.07 to 98.89 mm, fruit diameter from 53.98 to 92.44 mm, fruit peel thickness from 0.53 to 5.30 mm, 100-aril fresh weight from 24.49 to 51.32 g, and aril width from 1.81 to 7.99 mm. Khadivi et al.31 detected that petiole length ranged from 1.62 to 11.55 mm, petiole diameter from 0.42 to 1.60 mm, thorn length on annual shoots from 0.00 to 41.02 mm, thorn thickness on annual shoots from 0.00 to 2.98 mm, sepal number from 3 to 8, sepal length from 6.36 to 13.91 mm, sepal base width from 4.08 to 9.11 mm, aril width from 3.08 to 8.73 mm, seed length from 4.82 to 8.00 mm, and total soluble solids from 9.30 to 27.00%. Khadivi et al.31 determined that fruit calyx length ranged from 11.72 to 30.22 mm, aril length from 8.98 to 12.35 mm, aril width from 4.81 to 7.57 mm, seed length from 4.81 to 7.57 mm, and seed width from 1.85 to 3.18 mm. Khadivi et al.22 found that in their study conducted in Iran, fruit length ranged from 46.22 to 93.04 mm, fruit diameter from 48.42 to 108.69 mm, 100-aril fresh weight from 15.65 to 48.23 g, aril length from 8.24 to 12.75 mm, aril width from 3.45 to 9.45 mm, and seed width from 2.55 to 4.01 mm.
Overall, our findings are parallel to those of previous researchers. This parallelism emphasizes the reliability and validity of our results.
The frequency distribution of the measured qualitative pomological and morphological characteristics in the pomegranate genotypes studied is presented in detail in Table 2. The morphological and pomological traits of the studied pomegranate genotypes displayed considerable diversity. The most common tree growth habit in the study was spreading, observed in 19 genotypes, suggesting that this form may be more adaptable or beneficial in the given conditions. Open growth, seen in 13 genotypes, indicates a structure with more spacing between branches, potentially improving light absorption and airflow. The weeping habit, found in 8 genotypes, is less common and often ornamental but may face challenges like branch damage. Semi-erect (5 genotypes) and erect (2 genotypes) forms are less frequent, possibly due to limited ecological or structural advantages compared to spreading and open types. Overall, the prevalence of spreading and open forms highlights their better adaptation to the environment. The finding reveals that the majority of the genotypes (26 out of 47) exhibited intermediate growth vigor, suggesting that this level of vigor might be the most common or stable trait in the studied population under the given conditions. High vigor was observed in 16 genotypes, indicating that these genotypes are particularly robust and may have advantages in terms of growth speed, adaptability, and resource utilization. However, the low vigor group, comprising only 5 genotypes, represents a minority, which could be indicative of limited growth potential, possibly due to genetic factors, environmental stresses, or suboptimal growth conditions. The predominance of intermediate vigor suggests a balance between the extremes of high and low vigor, which could be a result of natural selection or a strategy for survival in variable environmental conditions17.
Branching was predominantly high (21 genotypes), intermediate (19 genotypes), and low (7 genotypes). The pattern of canopy density closely mirrors that of growth vigor, with the majority of genotypes (22 out of 44) exhibiting intermediate density. This suggests that a moderate canopy density may be a common or favorable trait under the studied conditions, potentially optimizing light capture while avoiding the drawbacks of both overly dense and sparse canopies. As a result, the dominance of intermediate density indicates a tendency toward more efficient canopy structures for optimal growth and resource acquisition. For trunk type, multi-trunk trees with intermediate structure were the most frequent (19 genotypes), followed by multi-trunk/low (16 genotypes) and multi-trunk/high (10 genotypes). Only 2 genotypes had a single trunk.
The tendency to suckering was intermediate in 23 genotypes, suggesting that moderate sucker production might be a common or stable trait under the studied conditions. High suckering, observed in 16 genotypes, could indicate a strong capacity for vegetative reproduction, which may be advantageous for rapid colonization or recovery after damage. However, excessive suckering might also lead to resource competition among shoots, potentially limiting overall growth. On the other hand, low suckering, found in 8 genotypes, suggests limited vegetative reproduction, which might be beneficial in environments where resource allocation to the main plant is prioritized, but it could also reduce the plant’s ability to regenerate quickly after disturbance. The prevalence of intermediate suckering suggests a balance between these trade-offs, where moderate suckering could be advantageous in optimizing both growth and survival. Leaf density was predominantly intermediate, observed in 23 genotypes, indicating a balanced leaf arrangement that may be optimal for maximizing light capture while minimizing shading effects. High leaf density, found in 19 genotypes, suggests an adaptation for maximizing photosynthetic capacity, which could be beneficial in environments with ample sunlight and nutrient availability. However, it could also lead to overcrowding and reduced airflow. Low leaf density, seen in 5 genotypes, might indicate a strategy to reduce competition for resources or to adapt to environments where light penetration or air circulation is more critical. When considering leaf shape, lanceolate was the dominant form, observed in 23 genotypes, while 16 had broad elliptic leaves and 8 were elliptic-lanceolate.
Rounded apexes were the most common leaf apex shape, present in 23 genotypes, followed by obtuse (22 genotypes) and acute (2 genotypes). The upper surface color of leaves was predominantly green (24 genotypes), while light green was observed in 20 genotypes and dark green in 3. On the lower surface, nearly all genotypes (42) had light green leaves, with only 5 displaying a green lower surface. Leaf margins were curly in 32 genotypes, while 15 had smooth margins. The color of petioles was mainly red-cream (32 genotypes), followed by cream-green (10 genotypes) and cream (5 genotypes).
Thorn presence on shoots was detected in 19 genotypes, whereas 28 were thornless. The diversity in ripening times reflects the adaptation strategies of pomegranate to the environmental conditions in which it is grown. The 6 genotypes ripening in mid-September have an early ripening period, which may offer the advantage of an earlier harvest. Early ripening pomegranates can be advantageous in terms of marketing, as early-season products may be in demand, and early ripening can also provide protection against some pests. The 14 genotypes ripening at the end of September allow for a slightly later harvest, potentially resulting in improved fruit quality and a more stable ripening process. Meanwhile, the 27 genotypes ripening in early October represent the most common ripening period for pomegranates. This period allows the fruit to reach optimal quality and enhances fruit yield. Overall, the prevalence of genotypes ripening in early October suggests that this period is the most suitable ripening time for the regional climate conditions. It implies that pomegranate producers may prefer these genotypes to obtain more stable and high-quality products. This diversity demonstrates the ability to adapt to various climatic conditions and offers flexibility to producers by providing market diversity. In this study, yield varied among the pomegranate genotypes, with 27 genotypes exhibiting high yield, indicating strong productivity potential under the given conditions. These high-yielding genotypes may be more resilient or better adapted to the environmental factors, allowing them to produce larger quantities of fruit. 13 genotypes had intermediate yield, suggesting moderate productivity, which could indicate a balance between environmental adaptability and growth constraints. Finally, 7 genotypes displayed low yield, which could be due to factors such as poor environmental adaptation, resource limitations, or genetic factors that limit fruit production. The variation in yield highlights the genetic diversity of pomegranates and the potential for selecting genotypes that maximize productivity while considering environmental factors. In terms of fruit shape, round fruits were the most common, observed in 22 genotypes, which suggests this shape may be preferred for its uniformity and potential market appeal. Oblate fruits, found in 13 genotypes, also appear frequently and may offer advantages in terms of handling and packing. Oval, elliptical, and ovate shapes, seen in fewer genotypes (6, 4, and 2 respectively), might indicate more specialized adaptations, possibly reflecting specific environmental or genetic factors that influence fruit development. The predominance of round and oblate shapes could suggest their suitability for commercial production, where consistent shape is often desired for easier harvesting and better presentation in the market.
Convex fruit bases were common in 23 genotypes, while truncate bases were observed in 16, and angular bases in 8. Most fruits (43 genotypes) were symmetrical, but asymmetry was noted in 4 genotypes. Pacifiers were absent in 30 genotypes, present but short in 14, and medium-sized in 3. Stamen density within the fruit calyx was categorized as high (17 genotypes), intermediate (12 genotypes), and low (18 genotypes).
Open calyx forms were predominant, present in 19 genotypes, while 13 were semi-open and 15 had closed calyces. Fruit necks were present in 32 genotypes, while 5 lacked a neck. Fruit peel color exhibited considerable variation among the genotypes. 20 genotypes displayed a yellow-green peel, which is often associated with early ripening stages and may be preferred for certain markets due to its visual appeal. 13 genotypes had a red-yellow peel, suggesting a more mature, ripened fruit, which could indicate better taste or higher sugar content, often desired by consumers. Nine genotypes showed a yellow-red peel, likely reflecting a different stage of ripening or an adaptation to specific growing conditions. 3 genotypes with a red peel may represent more specialized genotypes with distinct flavor profiles or environmental adaptations, while the 2 green-red genotypes might indicate a unique developmental characteristic. The variation in peel color highlights the genetic diversity of pomegranates and offers opportunities for selecting varieties based on consumer preferences and market demand.
The presence of fruit peel necrosis was mostly absent, noted in 23 genotypes, while low levels were observed in 12, intermediate in 6, and high in another 6 genotypes. Fruit peel sunburn was low in 26 genotypes, absent in 13, intermediate in 6, and high in just 2 genotypes. Internal peel color was predominantly cream in 26 genotypes and yellow in 21 genotypes. Similarly, internal layer color was cream in 40 genotypes, with only 7 displaying yellow layers.
Internal layer thickness was low in 27 genotypes, intermediate in 12, and high in 8. Septum color showed variation, with milky being the most common (27 genotypes), followed by cream (14 genotypes), glassy (4 genotypes), and yellow (2 genotypes). Septum thickness was classified as intermediate (27 genotypes), low (10 genotypes), and high (10 genotypes). Septum transparency was predominantly intermediate, found in 26 genotypes, while 12 were classified as low and 9 as high.
Aril adherence to the internal peel was intermediate in 28 genotypes, low in 13, and high in 6. Triangular aril shapes were the most frequent, observed in 34 genotypes, while prismatic shapes were found in 10 genotypes, and oval shapes in 3. Aril color varied significantly: glassy arils were the most common, found in 24 genotypes, followed by pale red (12 genotypes), pale milk (8 genotypes), and red (3 genotypes).
Seed hardness exhibited variation among the pomegranate genotypes; 29 genotypes had hard seeds, which may contribute to better seed protection and longer shelf life. Hard seeds can also enhance seed dispersal and germination success under specific environmental conditions. The 12 genotypes exhibited semi-soft seeds, striking a balance between durability and ease of consumption, which may appeal to different market segments. Finally, 6 genotypes (‘Sangan-1’, ‘Sangan-2’, ‘Karimabad-1’, ‘Karimabad-2’, ‘Karimabad-3’, and ‘Karimabad-4’) contained soft seeds; these seeds are generally preferred for fresh consumption due to their more pleasant taste, making them directly consumable. Soft seeds provide a significant advantage for fresh fruit consumption, as, unlike hard seeds, they do not pose a chewing difficulty to the consumer. This characteristic stands out, particularly in markets where fresh pomegranate consumption is increasing. The diversity in seed hardness reflects the genetic diversity of pomegranates and highlights the importance of varying seed textures concerning consumer preferences, as well as adaptation strategies to specific growing conditions. Fruit juice was colorless in 31 genotypes, pink in 9, and red in 7 genotypes. Fruit taste varied significantly across the pomegranate genotypes. The majority of the genotypes, 22 in total, exhibited a sour-sweet taste, which is typically considered the most desirable and commercially viable flavor profile due to its balance of tartness and sweetness. 19 genotypes (‘Daman-7’, ‘Sangan-1’, ‘Sangan-2’, ‘Sangan-9’, ‘Sangan-11’, ‘Sangan-12’, ‘Sangan-13’, ‘Sangan-15’, ‘Sangan-17’, ‘Karimabad-1’, ‘Karimabad-2’, ‘Karimabad-3’, ‘Karimabad-4’, ‘Padik-1’, ‘Padik-5’, ‘Padik-6’, ‘Padik-7’, ‘Padik-9’, and ‘Padik-10’) were sweet, which may appeal to consumers preferring a milder, more sugar-dominant taste, commonly sought for fresh consumption or juicing. On the other hand, 6 genotypes were sour, which could be more suitable for specific culinary uses or regional preferences. Fruit quality varied among the pomegranate genotypes, with 28 genotypes exhibiting intermediate quality, suggesting a balanced combination of desirable traits such as taste, appearance, and texture, suitable for a broad market. 10 genotypes (‘Sangan-1’, ‘Sangan-8’, ‘Sangan-10’, ‘Sangan-13’, ‘Sangan-14’, ‘Sangan-16’, ‘Padik-4’, ‘Padik-8’, ‘Padik-9’, and ‘Padik-10’) demonstrated high quality, which may indicate superior characteristics such as better taste, texture, and visual appeal, making them ideal for premium markets. On the other hand, 9 genotypes showed low quality, potentially due to factors like lower taste intensity, defects in appearance, or poor texture. Our findings show similarities and differences compared to the results of similar studies on pomegranate1,18,22,31,33. These differences are believed to stem from the diversity of the genotypes used and environmental factors.
Due to the limited number of similar studies on the subject, our findings have been thoroughly and independently discussed among themselves, as well as concerning the few existing studies in the literature. In doing so, we have provided a deeper understanding and a fresh perspective on the field, bridging critical gaps and enriching the existing body of knowledge. This approach not only highlights the novelty and comprehensiveness of our research but also underscores its contribution to advancing the academic discourse on pomegranate studies.
The fruits and leaves of the pomegranate genotypes studied, as well as the fruits and leaves of the selected soft-seeded pomegranate genotypes, are provided in Figs. 2 and 3, respectively.

The fruits and leaves of the pomegranate genotypes studied. The symbols represent the genotypes of each area, including Daman (D), Karimabad (K), Padik (P), and Sangan (S).

The fruits and leaves of the very soft-seeded pomegranate genotypes selected.
Correlation matrix analysis (CMA)
Correlation matrix analysis is a method used to assess the linear relationships between multiple variables. One common approach for calculating these relationships is Pearson’s correlation (r)34, which is widely used to measure the relationship between quantitative data35. By calculating the correlation coefficients for each pair of variables, the matrix reveals the strength and direction of these relationships, with correlation values ranging from − 1 (strongly negative) to + 1 (strongly positive), and values close to 0 indicating weak or no relationship. This analysis helps identify patterns, dependencies, or redundancies among variables, aiding decisions on dimensionality reduction or further statistical modeling. The results are often visualized in heat maps for easier interpretation, and statistical significance is assessed using p-values, with values less than 0.05 typically indicating significant correlations36.
Accordingly, the simple correlations among the quantitative pomological and morphological variables used in the studied pomegranate genotypes are presented in Fig. 4. The correlations observed among leaf characteristics provide insight into the relationships between different aspects of leaf morphology in pomegranate genotypes. The positive correlation between leaf length and leaf width (r = 0.61, p < 0.01) indicates that as leaves become longer, they also tend to become wider, suggesting proportional growth in leaf dimensions. This proportionality may reflect genetic factors that regulate balanced leaf expansion, which could optimize photosynthesis by increasing the surface area exposed to sunlight. The strong positive correlation between leaf length and petiole length (r = 0.69, p < 0.01) suggests that larger leaves require longer petioles for adequate structural support. This relationship may also enhance light capture efficiency by positioning the leaf blade more effectively within the canopy. Similarly, the correlations involving petiole diameter with both leaf length (r = 0.68, p < 0.01) and petiole length (r = 0.68, p < 0.01) highlight the structural adaptations necessary to support larger leaves. Thicker petioles are likely needed to provide mechanical strength and facilitate efficient nutrient and water transport to larger leaf blades. These relationships collectively emphasize the coordinated development of leaf dimensions and their supporting structures, which are critical for maintaining functional efficiency and ensuring optimal plant performance.
The observed significant and positive correlation between thorn thickness on annual shoots and thorn length on annual shoot (r = 0.68, p < 0.01) suggests a proportional relationship between these two traits, indicating that as thorns become thicker, they also tend to become longer. From an ecological perspective, larger thorns may provide enhanced defense mechanisms against herbivores or environmental stressors.
For fruit-related traits, fruit diameter and fruit length exhibited a positive correlation (r = 0.95, p < 0.01), indicating their linear dependence. Additionally, fruit weight was correlated with both fruit diameter (r = 0.97, p < 0.01) and fruit length (r = 0.95, p < 0.01), highlighting the role of size in determining fruit weight. These relationships demonstrate the interconnectedness of fruit morphology, where attributes such as diameter, length, and weight are proportionally linked. Understanding these correlations can assist in selecting genotypes with optimal fruit size, which is a key trait for commercial production, contributing to higher yields and improved marketability.
Sepal number showed a correlation with fruit diameter (r = 0.35, p < 0.05), indicating a potential link between sepal number and fruit size. The correlations of sepal base width with sepal length (r = 0.56, p < 0.01) and seed length (r = 0.33, p < 0.05) suggest that sepal characteristics influence both fruit structure and seed development, highlighting the interconnection between external and internal fruit traits.
Fruit calyx traits also demonstrated notable correlations. Fruit calyx length was positively associated with thorn length on annual shoot (r = 0.32, p < 0.05), thorn thickness on annual shoot (r = 0.31, p < 0.05), fruit length (r = 0.30, p < 0.05), and sepal number (r = 0.34, p < 0.05) whereas fruit calyx diameter was linked to sepal length (r = 0.30, p < 0.05).
The strong correlations observed between 100-aril fresh weight and both aril length (r = 0.69, p < 0.01) and aril width (r = 0.67, p < 0.01) emphasize the critical role of aril size in determining the overall weight of the fruit. Larger arils directly contribute to a heavier fruit, highlighting their significance in fruit mass. Additionally, the positive correlation between aril width and fruit diameter (r = 0.33, p < 0.05) suggests that as the fruit diameter increases, the aril size tends to increase as well.
The correlations involving fruit peel weight highlight its relationships with various pomological and morphological traits, reflecting its integral role in fruit development. The positive correlation with thorn length on annual shoots (r = 0.35, p < 0.05) suggests that larger or more robust plants, which typically have longer thorns, may contribute to heavier peels, potentially indicating a link between vegetative growth and fruit characteristics. The correlations with fruit length (r = 0.91, p < 0.01) and fruit diameter (r = 0.91, p < 0.01) indicate that as fruit size increases, peel weight rises proportionally. This is expected since larger fruits require thicker and heavier peels for structural integrity and protection of the inner components, such as arils. The positive relationship with sepal number (r = 0.38, p < 0.01) may reflect a developmental link where fruits with more sepal tend to produce heavier peels. This suggests that genotypes with more pronounced sepal structures may develop thicker or denser peels. Similarly, the correlation with fruit calyx length (r = 0.42, p < 0.01) highlights the role of calyx development in peel characteristics. Longer calyces may indicate a more developed outer structure, contributing to an increase in peel mass. This relationship underscores the importance of structural traits in influencing peel weight. The correlation with fruit weight (r = 0.92, p < 0.01) demonstrates that peel weight is a key contributor to overall fruit mass. This finding highlights the importance of peel traits in determining marketable yield and supports the idea that heavier peels are associated with larger, more commercially desirable fruits. Lastly, the correlation with fruit peel thickness (r = 0.30, p < 0.05) confirms that thicker peels naturally weigh more, further emphasizing the functional relationship between fruit peel thickness and peel mass. This trait is particularly relevant for fruit durability during transportation and storage, as thicker peels provide better protection for the arils. Overall, these findings suggest that peel characteristics are closely linked to fruit size, structure, and marketability. The relationships observed indicate that selecting genotypes with optimal peel traits could improve both yield and post-harvest quality in pomegranate production.
The positive correlations between seed width and aril width (r = 0.36, p < 0.05), and between seed width and seed length (r = 0.50, p < 0.01), indicate that seed dimensions grow proportionally and are also related to aril width. This suggests that larger seeds may be associated with larger arils, and seed size could potentially influence fruit quality.
Lastly, total soluble solids, a critical pomological trait, showed positive correlations with both leaf length (r = 0.29, p < 0.05) and 100-aril fresh weight (r = 0.35, p < 0.05). This indicates a relationship between larger leaves and arils with higher sugar content. Our findings align with the results of similar studies conducted on pomegranate1,18,31,33,37.
This information provides valuable insights for breeding programs focused on improving fruit and seed quality, offering a solid foundation for strategies aimed at enhancing both productivity and marketable traits. By leveraging these findings, breeders can prioritize genotypes that combine optimal size, weight, and internal characteristics, ultimately contributing to superior fruit quality and higher agricultural efficiency. These results underscore the importance of an integrated approach to pomegranate improvement, addressing key morphological and pomological traits to meet both commercial and consumer demands38,39.
Given the limited number of similar studies on the subject, our findings have been thoroughly and independently discussed, both concerning each other and in the context of the few existing studies in the literature. By doing so, we have contributed a deeper understanding and a fresh perspective to the field, bridging key gaps and enriching the current body of knowledge. This approach not only emphasizes the originality and comprehensiveness of our research but also underscores its role in advancing the academic discourse on pomegranate studies.

Simple correlations among the quantitative pomological and morphological variables utilized in the studied pomegranate genotypes. For abbreviations, please see Table 1.
Multiple regression analysis (MRA)
In multiple regression analysis (MRA), a statistical method used to explore the relationship between a dependent variable and multiple independent variables, several key metrics are critical for evaluating the model’s performance. The correlation coefficient (r) indicates the strength and direction of the relationship between the variables. Coefficient of determination (r²) shows how well the independent variables explain the variance in the dependent variable, with values closer to 1 indicating better explanatory power. The standardized beta coefficients (β) allow comparison of the relative influence of predictors, as they standardize the variables to the same scale. t values are used to assess whether the coefficients are significantly different from zero, and the p values indicate statistical significance. A p-value less than 0.05 typically suggests that the predictor variable is significantly contributing to the model. Additionally, Variance Inflation Factors (VIF) are calculated to check for multicollinearity, ensuring that the independent variables are not highly correlated with one another, which could distort the results. These metrics, when interpreted together, provide a comprehensive view of the relationships in the data and the strength of the model’s predictions40,41,42.
Initially, after calculating the simple correlation coefficients, fruit weight, fruit peel weight, 100-aril fresh weight, seed length, seed width, and total soluble solids were considered as dependent variables, and the direct and indirect effects of the independent variables on these key traits were examined. The results of the MRA revealed that fruit weight was associated with two traits, fruit peel weight with seven traits, 100-aril fresh weight with four traits, seed length with two traits, seed width with two traits, and total soluble solids with two traits (Table 3).
Fruit weight was strongly influenced by fruit diameter (β = 0.66, p < 0.00) and fruit length (β = 0.32, p < 0.01). This demonstrates that fruit size, encompassing both diameter and length, plays a dominant role in determining fruit weight. The significance of these traits underscores the need to prioritize genotypes with larger dimensions to achieve higher yields, which is crucial for commercial production and marketability.
The MRA indicates that fruit peel weight is primarily influenced by fruit weight (β = 0.92, p < 0.00), underscoring the close relationship between overall fruit mass and peel weight. Additionally, fruit calyx length (β = 0.20, p < 0.00) and fruit peel thickness (β = 0.12, p < 0.00) are positively associated with peel weight, highlighting the importance of structural and protective traits. Interestingly, the negative correlation with aril width (β = − 0.20, p < 0.00) suggests a potential trade-off between peel robustness and internal aril dimensions, possibly due to resource allocation during fruit development. Similarly, total soluble solids (β = − 0.11, p < 0.01) showed a negative association, indicating that fruits with thicker peels might exhibit lower sugar concentrations, which could impact fruit sweetness. Additional variables, such as seed width (β = 0.16, p < 0.00) and fruit calyx diameter (β = − 0.09, p < 0.03), also contribute to variations in peel weight. These findings reveal a multifaceted relationship where both internal and external traits influence peel characteristics, emphasizing the need for balanced selection criteria in breeding programs.
The primary factors influencing 100-aril fresh weight were aril length (β = 0.53, p < 0.00), aril width (β = 0.40, p < 0.00), and total soluble solids (β = 0.33, p < 0.00). These traits are critical for improving fruit sweetness and overall aril size, which directly affect consumer preferences and market demand. Notably, the negative correlation with thorn length on annual shoot (β = − 0.20, p < 0.01) highlights a potential inverse relationship between vegetative growth and fruit quality.
Seed length was primarily influenced by seed width (β = 0.36, p < 0.01) and fruit calyx diameter (β = 0.32, p < 0.02). This suggests that external fruit structures, such as the calyx, may play a developmental role in shaping seed characteristics. Similarly, seed width was influenced by aril width (β = 0.35, p < 0.01) and sepal length (β = 0.27, p < 0.03), indicating an interconnection between internal and external fruit traits.
The total soluble solids, a critical determinant of fruit sweetness and quality, was positively associated with 100-aril fresh weight (β = 0.80, p < 0.00), demonstrating the importance of aril size in determining sugar content. Interestingly, a negative association with aril length (β = − 0.65, p < 0.00) indicates a potential trade-off between aril elongation and sugar concentration, possibly due to resource allocation during development. The bold values are supported by the correlation matrix analysis.
As a result, the findings of the MRA highlight the significant relationships between key morphological and pomological traits in pomegranate genotypes. Fruit weight is primarily determined by fruit diameter and length, while fruit peel weight is strongly influenced by fruit size, peel thickness, and calyx length. The findings also reveal trade-offs between peel robustness and internal fruit quality, such as aril size and sugar content. The analysis of 100-aril fresh weight underscores the importance of aril size for fruit sweetness, with a potential inverse relationship between vegetative growth and fruit quality. Additionally, seed characteristics are influenced by external fruit structures, such as the calyx and sepal length, indicating the interconnectedness of internal and external traits. These insights provide valuable information for breeding programs aimed at improving fruit quality, yield, and consumer appeal.
In previous studies on pomegranates, we did not encounter the use of MRA. Therefore, our results have been independently and thoroughly discussed among themselves. This contributes a novel and comprehensive approach to the existing literature, filling a significant gap and providing new insights into the relationships between various morphological and pomological traits. By incorporating MRA, our study adds depth and precision to understanding the complex dynamics within pomegranate traits, offering valuable data for future breeding programs and improving fruit quality and yield. This work enhances the academic discourse by offering a fresh perspective, which is essential for advancing the field of pomegranate research.
Principal component analysis (PCA)
In this study, principal component analysis (PCA) was applied using both quantitative and qualitative data. PCA is a technique that works exclusively with continuous, numerical data, so qualitative data were converted into numerical form to ensure compatibility with PCA43. The qualitative variables were numerically encoded based on the nature of the categorical data. Label encoding was used for ordinal categories, as this method is more suitable for ordered data44. In cases where there was no inherent ordering between categories, one-hot encoding could be considered, but this was not necessary in this study.
Once the data were appropriately encoded, PCA was performed to reduce the dimensionality of the dataset and identify the principal components that explained the most variance. The PCA procedure involved calculating the covariance matrix, extracting eigenvectors and eigenvalues, and transforming the data into new, orthogonal components45. The eigenvalues were extended to 1, and the variance explained by the principal components was examined46. To enhance the interpretability of the components, the Varimax rotation with Kaiser Normalization method was applied. This approach clarified the relationships between components, maximizing the variance of squared loadings of each variable on the components47. This process enabled a deeper understanding of the data structure and helped in identifying the key traits influencing genotype grouping. Thus, it allows for a simpler analysis by reducing the data size and increases the power of the analysis results by minimizing the loss of information48. The results are presented in Table 4, along with the variance explained by each principal component.
Accordingly, the first 17 components explain a cumulative variance of 84.00%. The first three components, PC1 (11.66%), PC2 (10.42%), and PC3 (9.46%), together account for 31.54% of the total variance, capturing a substantial portion of the dataset’s variability. The significance of these components was confirmed, with PC1, PC2, and PC3 being statistically significant (**p < 0.05).
PC1 explains 11.66% of the total variance and is heavily influenced by fruit-related traits. The bolded eigenvector coefficients reveal that fruit length (0.94), fruit diameter (0.93), fruit weight (0.93), and fruit peel weight (0.93) are the strongest contributors. These traits are critical indicators of fruit size and overall marketability, making PC1 a reflection of fruit dimensions and weight characteristics. The dominance of fruit traits in PC1 aligns with the importance of these traits in both commercial and breeding contexts. Larger fruit size and higher weight are desirable for their economic value in fresh fruit markets and processing industries. Furthermore, these traits are often used as selection criteria for superior genotypes23,49. The clustering of these variables within PC1 suggests a strong correlation among them, emphasizing their collective importance in genotype differentiation1,19,50. The findings we obtained for PC1 are in agreement with the researchers’ findings.
PC2 accounts for 10.42% of the total variance and is primarily supported by the following traits in order of contribution: thorn thickness on the annual shoot (0.95), thorn presence on the shoot (0.95), thorn length on the annual shoot (0.76), fruit neck presence (0.54), and ripening time (− 0.57), which exhibits a strong negative association with this component. The high contribution of thorn-related traits underscores their significance in managing pomegranate orchards. Thorns can pose challenges during harvesting and pruning, making their reduction a priority for commercial cultivation. The inclusion of fruit neck presence further highlights its relevance, as it can influence fruit structure and ease of handling, indirectly impacting market preference and harvest efficiency. Simultaneously, ripening time remains critical for market availability and regional adaptation, with early ripening genotypes being particularly valuable for extending the harvest season. The inverse relationship between ripening time and thorn traits, along with the moderate contribution of fruit neck presence, suggests potential genetic or developmental links worth further investigation in breeding programs. In similar studies, Khadivi-Khub et al.1 reported that PC2 was primarily supported by 100-aril dry weight, aril length, 100-aril fresh weight, and aril diameter traits. Similar findings were also detected by Khadivi et al.22.
PC3 explains 9.46% of the total variance and is influenced primarily by vegetative growth traits, including canopy density (0.86), leaf density (0.84), branching (0.81), tree growth vigor (0.78), and tendency to suckering (0.77). These characteristics define the structural architecture of the tree, which has significant implications for orchard management and productivity. The strong contribution of canopy density and leaf density to PC3 highlights the importance of vegetative growth in shaping tree performance. Genotypes with denser canopies, higher leaf density, and vigorous branching may achieve higher photosynthetic efficiency, leading to improved fruit yield. However, excessive vegetative growth can also increase maintenance costs due to the need for frequent pruning. PC3, therefore, reflects a balance between growth vigor, leaf density, and structural management. Khadivi-Khub et al.1 determined that PC3 is primarily supported by traits such as fruit length/diameter, fruit shape, and fruit crown length, whereas Khadivi and Arab33 stated that it is supported by traits such as aril color, fruit juice color, and fruit quality.
The findings we obtained for PC2 and PC3 do not show similarity with those of previous researchers. This discrepancy is likely attributed to the differences in the genotypes used as materials and various ecological factors.
In conclusion, the PCA findings highlight the multifaceted nature of pomegranate genotype evaluation. PC1 emphasizes fruit size and weight, which are critical for market preferences, while PC2 sheds light on thorniness and ripening time, factors essential for orchard operations and market timing. PC3, on the other hand, focuses on canopy and branching, providing insights into tree architecture and maintenance requirements. Together, these components offer a comprehensive framework for selecting genotypes that balance commercial value, ease of management, and adaptability. For breeding programs, the significant contribution of fruit traits (PC1) suggests that genotypes with larger and heavier fruits should be prioritized for commercial production. Conversely, PC2 highlights the need to reduce thorniness while optimizing ripening time. Lastly, PC3 emphasizes the importance of balancing vegetative growth traits to ensure manageable tree architecture without compromising yield potential. The first three components collectively guide breeding and selection strategies, focusing on market-driven traits, orchard management efficiency, and adaptability to different growing conditions.
PCA is based on components derived from quantitative data, and the biplot is used to visualize these components and loading vectors. Although categorical variables have been converted into numerical form, their non-linear relationships may make their visualization in a biplot misleading51. Therefore, only the results from quantitative data have been visualized in the biplot, ensuring an accurate and meaningful representation52. The biplot represents both observations and variables in the same space, offering an effective method for dimensionality reduction. It particularly helps in extracting meaningful results from large datasets. Arrows indicate the magnitude and direction of the variables, while the distance between data points reflects their similarity53.
Biplot for the studied pomegranate genotypes and quantitative variables based on PC1/PC2 of pomological and morphological traits is shown in Fig. 5. Thus, the first two components (PC1 = 20.93% and PC2 = 16.03%) account for 36.96% of the total variation.
The genotypes were distributed across the four clusters of the biplot, while the variables were grouped into clusters 1, 2, and 4. In cluster 1, genotypes ‘Karimabad-5’, ‘Karimabad-6’, ‘Padik-2’, ‘Padik-4’, ‘Padik-5’, ‘Padik-6’, ‘Padik-7’, ‘Padik-10’, ‘Padik-11’, ‘Padik-13’, ‘Sangan-5’, ‘Sangan-6’, ‘Sangan-7’, and ‘Sangan-9’ were associated with the variables leaf width, sepal base width, and fruit calyx diameter. This suggests that these genotypes share common morphological characteristics related to leaf and calyx dimensions, making them suitable for breeding programs that prioritize vegetative growth traits.
In cluster 2, genotypes ‘Padik-8’, ‘Padik-9’, ‘Padik-12’, ‘Sangan-8’, ‘Sangan-12’, ‘Sangan-13’, ‘Sangan-16’, and ‘Sangan-17’ were associated with the variables leaf length, petiole length, petiole diameter, sepal length, 100-aril fresh weight, aril length, aril width, seed length, seed width, and total soluble solids. This cluster primarily represents genotypes with superior reproductive and fruit quality traits, particularly those affecting sweetness, seed dimensions, and fruit size, which are significant for consumer acceptance and market value.
In cluster 3, genotypes ‘Daman-9’, ‘Karimabad-1’, ‘Karimabad-2’, ‘Karimabad-3’, ‘Karimabad-4’, ‘Karimabad-7’, ‘Karimabad-8’, ‘Padik-3’, ‘Sangan-1’, ‘Sangan-2’, ‘Sangan-3’, and ‘Sangan-4’ were positioned without a strong association with specific variables. These genotypes likely exhibit intermediate or undefined characteristics, which could be indicative of genetic variability and potential adaptability under different cultivation conditions.
In cluster 4, genotypes ‘Daman-1’, ‘Daman-2’, ‘Daman-3’, ‘Daman-4’, ‘Daman-5’, ‘Daman-6’, ‘Daman-7’, ‘Daman-8’, ‘Padik-1’, ‘Sangan-10’, ‘Sangan-11’, ‘Sangan-14’, and ‘Sangan-15’ were associated with variables such as thorn length on annual shoot, thorn thickness on annual shoot, fruit length, fruit diameter, sepal number, fruit calyx length, fruit weight, fruit stalk length, fruit peel thickness, and fruit peel weight. This cluster highlights genotypes with robust fruit traits and thorn morphology, indicating suitability for both fruit production and ornamental purposes.
Additionally, the genotypes ‘Daman-2’, ‘Daman-5’, ‘Daman-7’, ‘Karimabad-4’, ‘Padik-9’, ‘Sangan-2’, and ‘Sangan-4’ were located outside the 95% confidence ellipse along with the variables fruit length, fruit diameter, fruit weight, and fruit peel weight. This positioning suggests that these genotypes exhibit unique or extreme trait combinations, making them valuable for introducing novel traits into breeding programs.
The 95% confidence ellipse represents an area where 95% of the data points fall. Points outside this ellipse are considered outliers, which deviate from the normal distribution and typically require further investigation. These outliers may indicate interesting findings related to genetic diversity or environmental factors, but their validity should be confirmed through additional testing54,55.
Overall, the distribution of genotypes and traits across the biplot underscores the diversity within the studied population. The clustering provides actionable insights for breeding programs, allowing the selection of genotypes tailored to specific traits, such as fruit quality, vegetative growth, or thorn morphology. The identification of outliers offers opportunities to incorporate genetic novelty and enhance variability, which could improve adaptability and productivity in varying environmental conditions. This analysis highlights the potential for targeted breeding strategies to optimize both agronomic and commercial traits in pomegranate cultivation.

Biplot for the studied pomegranate genotypes and quantitative variables based on PC1/PC2 of pomological and morphological traits. For abbreviations, please see Table 1.
Heat map analysis (HMA)
A heat map analysis (HMA) was performed using Ward’s hierarchical clustering method combined with Euclidean distance as a metric to group and visualize relationships among the data56. This method is particularly effective for identifying patterns and relationships within datasets characterized by quantitative variables. The analysis relied on numerical data derived from morphological and pomological measurements, ensuring precise and accurate calculations for both distance and clustering. The Euclidean distance metric was chosen because it is well-suited for measuring direct, linear distances between quantitative observations, allowing for an intuitive representation of similarity or dissimilarity among data points57. Ward’s method was applied to minimize variance within clusters, creating a hierarchical grouping that optimally represents relationships among the variables. The resulting heat map effectively displays these relationships, with a color gradient representing the magnitude of the quantitative data values across different clusters or observations. This approach ensures that the analysis remains robust and interpretable, relying exclusively on numerical data to enhance the clarity and reliability of the visual output58. Visualization of clustering patterns of pomegranate genotypes and variables based on morphological and pomological characterizations using a heat map is presented in detail in Fig. 6.
Based on the heat map analysis, the variables were initially divided into two main groups, A and B. Subsequently, each group was further subdivided into two subgroups (A1, A2, and B1, B2). The A1 subgroup contained the variables leaf length, fruit length, fruit diameter, and fruit peel weight, while the A2 subgroup included only the variable fruit weight. In the B1 subgroup, the variables were leaf width, fruit calyx length, fruit calyx diameter, 100-aril fresh weight, sepal length, aril length, total soluble solids, fruit stalk length, petiole length, sepal number, aril width, sepal base width, and seed length. Finally, the B2 subgroup included petiole diameter, thorn thickness on annual shoot, thorn length on annual shoot, fruit peel thickness, and seed width.
The genotypes were similarly divided into groups. Initially, the genotypes were split into two primary groups, C and D, which were then further divided into two subgroups each (C1, C2, and D1, D2). The C1 subgroup included the genotypes ‘Padik-10’, ‘Padik-9’, ‘Padik-8’, ‘Padik-7’, ‘Padik-6’, ‘Padik-13’, ‘Padik-5’, ‘Sangan-7’, ‘Sangan-6’, and ‘Sangan-5’, while the C2 subgroup contained ‘Sangan-12’, ‘Sangan-15’, ‘Sangan-10’, ‘Sangan-14’, ‘Sangan-16’, ‘Sangan-17’, ‘Sangan-11’, ‘Sangan-13’, and ‘Daman-8’. In the D1 subgroup, the genotypes included ‘Daman-6’, ‘Daman-5’, ‘Daman-4’, ‘Daman-3’, ‘Daman-7’, ‘Daman-2’, and ‘Daman-1’, whereas the D2 subgroup consisted of ‘Karimabad-6’, ‘Karimabad-5’, ‘Karimabad-3’, ‘Karimabad-2’, ‘Karimabad-4’, ‘Sangan-2’, ‘Padik-4’, ‘Karimabad-8’, ‘Karimabad-7’, ‘Sangan-1’, ‘Padik-11’, ‘Padik-3’, ‘Padik-2’, ‘Sangan-4’, ‘Sangan-3’, ‘Padik-12’, ‘Karimabad-1’, ‘Sangan-9’, ‘Sangan-8’, ‘Padik-1’, and ‘Daman-9’.
The results of this heat map analysis reveal key clustering patterns and provide meaningful insights into the relationships between the variables and genotypes. Subgroup A1 primarily contains variables associated with size-related traits of leaves and fruits. These traits are critical indicators of both vegetative growth and fruit quality, making this subgroup highly relevant for breeding programs targeting overall yield and commercial appeal. The inclusion of fruit weight as the sole variable in subgroup A2 highlights its distinct contribution to phenotypic diversity, suggesting that this variable may have unique genetic and environmental determinants compared to other fruit-related traits.
The variables in subgroup B1 encompass a diverse range of traits, including reproductive, structural, and compositional characteristics. This subgroup indicates the genotypes’ multifaceted contributions to fruit quality, particularly traits like total soluble solids and 100-aril fresh weight, which directly influence consumer acceptance and marketability. On the other hand, subgroup B2 comprises structural traits such as thorn and petiole dimensions, emphasizing morphological adaptations that may be crucial for environmental resilience and pest resistance.
The clustering of genotypes also provides actionable insights. Genotypes in subgroup C1 appear to share similar characteristics, particularly concerning traits in subgroup A1, suggesting that these genotypes are characterized by larger leaves and fruits. This subgroup likely represents genotypes with superior vegetative vigor and fruit size, which are desirable traits for high-yield production systems. Subgroup C2, meanwhile, includes genotypes associated with diverse traits in subgroup B1, indicating their suitability for breeding programs focused on improving both fruit quality and compositional traits, such as aril and seed dimensions.
Subgroup D1 contains genotypes that are more closely associated with subgroup B2 traits, which include thorn morphology and structural attributes. These genotypes may be well-suited for environments where thorn traits and structural robustness are advantageous, such as arid or pest-prone regions. Finally, subgroup D2 represents the most diverse collection of genotypes, linked to multiple traits spanning both A and B groups. This diversity makes these genotypes particularly valuable for introducing broad-spectrum traits into breeding programs, enabling adaptability and resilience in varying environmental conditions.
In summary, the heat map analysis underscores the complexity and diversity within the pomegranate genotypes and variables. The clustering patterns provide a systematic framework for selecting genotypes with specific desirable traits. For instance, traits in subgroups A1 and B1 are pivotal for improving fruit size and quality, while those in subgroup B2 contribute to structural resilience. The genotype clusters reflect these trait groupings, allowing for targeted breeding strategies that align with both agronomic and market demands. The inclusion of diverse and outlier genotypes in subgroup D2 further enriches the genetic pool, offering the potential for novel trait combinations and enhanced adaptability.

Visualization of clustering patterns of pomegranate genotypes and variables based on morphological and pomological characterizations using heat map.
Conclusion
This study provides a comprehensive assessment of morphological and pomological traits in pomegranate genotypes, revealing significant genetic and phenotypic diversity within the species. The findings underscore the importance of selecting genotypes based on traits that meet market demands, improve agricultural productivity, and ensure adaptability to diverse environmental conditions. Importantly, the genotypes identified in this study not only possess good quality characteristics, but also exhibit strong agronomic traits. This combination of quality and agronomic performance makes them promising candidates for breeding programs aimed at addressing both market needs and environmental challenges. Through advanced statistical techniques such as PCA and MRA, key traits influencing fruit quality, yield, and adaptability were identified.
The results highlight the crucial role of specific traits in genotype differentiation. Traits related to fruit size and weight (PC1), thorn presence, ripening time (PC2), and vegetative growth (PC3) were identified as key contributors to genetic variation. Genotypes with desirable characteristics, such as larger fruits, balanced vegetative growth, and reduced thorniness, were emphasized for their commercial potential, given their marketability and ease of management. The heat map analysis reinforced these findings by clustering variables and genotypes into distinct groups, offering valuable insights for targeted breeding strategies.
Significantly, ‘Sangan-1’, ‘Sangan-2’, ‘Karimabad-1’, ‘Karimabad-2’, ‘Karimabad-3’, and ‘Karimabad-4’ were identified as very soft-seeded genotypes, a highly sought-after trait for fresh consumption. These genotypes align with consumer preferences, enhancing palatability and market demand. Additionally, ‘Daman-7’, ‘Sangan-1’, ‘Sangan-2’, ‘Sangan-9’, ‘Sangan-11’, ‘Sangan-12’, ‘Sangan-13’, ‘Sangan-15’, ‘Sangan-17’, ‘Karimabad-1’, ‘Karimabad-2’, ‘Karimabad-3’, ‘Karimabad-4’, ‘Padik-1’, ‘Padik-5’, ‘Padik-6’, ‘Padik-7’, ‘Padik-9’, and ‘Padik-10’ were classified as sweet, with sweetness being a key factor in consumer preference and boosting the commercial potential of these genotypes.
Moreover, ‘Sangan-1’, ‘Sangan-8’, ‘Sangan-10’, ‘Sangan-13’, ‘Sangan-14’, ‘Sangan-16’, ‘Padik-4’, ‘Padik-8’, ‘Padik-9’, and ‘Padik-10’ were identified as high-quality genotypes, offering a valuable resource for breeding premium cultivars. Notably, ‘Sangan-1’ appeared in the very soft-seeded, sweet, and high-quality categories, positioning it as an especially promising genotype for breeding programs targeting multiple desirable traits.
Interestingly, outlier genotypes such as ‘Sangan-2’ and ‘Karimabad-4’, which belong to the soft-seeded group, were located outside the 95% confidence ellipse. This positioning suggests that these genotypes exhibit unique or extreme trait combinations, presenting opportunities to introduce novel characteristics into breeding programs. Their distinctiveness warrants further investigation to better understand their potential to enhance genetic diversity and improve adaptability.
In conclusion, this study addresses critical gaps in understanding pomegranate genotypic diversity by providing actionable insights into trait selection and breeding. The identification of key genotypes, such as ‘Sangan-1’, ‘Sangan-2’, and ‘Karimabad-4’, underscores the potential to develop superior cultivars that balance high yield, consumer appeal, and adaptability. By integrating advanced statistical analyses with detailed phenotypic evaluations, this research provides a solid foundation for future breeding strategies aimed at optimizing both agronomic and commercial traits in pomegranate cultivation.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
Khadivi-Khub, A., Kameli, M., Moshfeghi, N. & Ebrahimi, A. Phenotypic characterization and relatedness among some Iranian pomegranate (Punica granatum L.) accessions. Trees 29, 893–901. https://doi.org/10.1007/s00468-015-1172-9 (2015).
Radunić, M., Jukić Špika, M. & Gadže, J. Phenotypic diversity of pomegranate cultivars: discriminating power of some morphological and fruit chemical characteristics. Horticulturae 10 (6), 563. https://doi.org/10.3390/horticulturae10060563 (2024).
Levin, G. M. Pomegranate Roads: A Soviet Botanist’s Exile from Eden, 1st edn, 15–183 (Floreant, 2006).
Simmonds, N. W. Evolution of Crop Plants (Longmans, 1976).
NATIONSLAKES. Which Country Produces the Most Pomegranates?. https://globaltopstats.com/which-country-produces-the-most-pomegranates/ (Accessed 4 December 2024) (2024).
Khadivi, A., Rezagholi, M. & Shams, M. Phytochemical properties and bioactive compounds of pomegranate (Punica granatum L.). J. Hortic. Sci. Biotechnol. 99 (6), 639–652. https://doi.org/10.1080/14620316.2024.2371597 (2024a).
Calani, L. et al. Ultra-HPLC–MS N (poly) phenolic profiling and chemometric analysis of juices from ancient Punica granatum L. cultivars: A nontargeted approach. J. Agric. Food Chem. 61 (23), 5600–5609. https://doi.org/10.1021/jf400387c (2013).
Thakur, N. S., Dhaygude, G. S. & Gupta, A. Physico-chemical characteristics of wild pomegranate fruits in different locations of Himachal Pradesh. Int. J. Farm. Sci. 1 (2), 37–44 (2011).
Shilpa, P. et al. Pomegranate genetic resources: Conservation and utilization. In Fruit and Nut Crops 529–570. https://doi.org/10.1007/978-981-99-5348-6_18 (Springer, 2024).
Coşkun, O. F. & Gülşen, O. Determination of markers associated with important agronomic traits of watermelon (Citrullus lanatus L.). J. Agric. Sci. Technol. 26 (6), 1359–1371. https://doi.org/10.22034/JAST.26.6.1359 (2024).
Coşkun, Ö. F., Toprak, S. & Mavi, K. Some seed properties and molecular analysis with inter-primary binding site (iPBS) retrotranposons markers of edible-seeded watermelon genotypes. Genet. Resour. Crop Evol. 71, 3151–3162. https://doi.org/10.1007/s10722-023-01845-9 (2024).
Holland, D., Hatib, K. & Bar-Ya’akov, I. Pomegranate: botany, horticulture, breeding. Hortic. Rev. 35, 127–191 (2009).
Mars, M. Pomegranate plant material: genetic resources and breeding, a review. Opt. Mediterr. Ser. A 42, 55–62 (2000).
Stover, E. D. & Mercure, E. W. The pomegranate: a new look at the fruit of paradise. HortScience 42 (5), 1088–1092. https://doi.org/10.21273/HORTSCI.42.5.1088 (2007).
Shahsavari, S., Noormohammadi, Z., Sheidai, M., Farahani, F. & Vazifeshenas, M. R. A bioinformatic insight into the genetic diversity within pomegranate cultivars: from nuclear to Chloroplast genes. Genet. Resour. Crop Evol. 1, 1–11. https://doi.org/10.1007/s10722-021-01297-z (2022).
Venkitasamy, C., Zhao, L., Zhang, R. & Pan, Z. Pomegranate. In Integrated Processing Technologies for Food and Agricultural By-Products 181–216. https://doi.org/10.1016/B978-0-12-814138-0.00008-3 (Academic Press, 2019).
Levin, G. M. Pomegranate (Punica granatum) plant genetic resources in Turkmenistan. Plant Genet. Resour. Newsl. 97, 31–36 (1994).
Khadivi, A., Ayenehkar, D., Kazemi, M. & Khaleghi, A. Phenotypic and pomological characterization of a pomegranate (Punica granatum L.) germplasm collection and identification of the promising selections. Sci. Hort. 238, 234–245. https://doi.org/10.1016/j.scienta.2018.04.062 (2018).
Caliskan, O. & Bayazit, S. Morpho-pomological and chemical diversity of pomegranate genotypes grown in Eastern Mediterranean region of Turkey. J. Agric. Sci. Technol. 15 (7), 1449–1460 (2013).
Costa, Y. & Melgarejo, P. A study of the production costs of two pomegranate varieties grown in poor quality soils. Opt. Méditerr. Série Sémin. Méditerr. 42, 49–53 (2000).
Langlie, B. S., Mueller, N. G., Spengler, R. N. & Fritz, G. J. Agricultural origins from the ground up: archaeological approaches to plant domestication. Am. J. Bot. 101 (10), 1601–1617. https://doi.org/10.3732/ajb.1400145 (2014).
Khadivi, A., Hosseini, A. S. & Naderi, A. Morphological and pomological variation of pomegranate (Punica granatum L.) genotypes. Genet. Resour. Crop Evol. 71 (4), 1521–1528. https://doi.org/10.1007/s10722-023-01715-4 (2024b).
Mars, M. & Marrakchi, M. Diversity of pomegranate (Punica granatum L.) germplasm in Tunisia. Genet. Resour. Crop Evol. 46, 461–467 (1999).
JMP®. https://www.jmp.com/en_us/home.html (Accessed 4 December 2024) (2024).
OriginLab®. https://www.originlab.com (Accessed 4 December 2024) (2024).
Norusis, M. J. SPSS/PC Advanced Statistics (SPSS Inc., 1998).
Goodarzi, S., Khadivi, A., Abbasifar, A. & Akramian, M. Phenotypic, pomological and chemical variations of the seedless barberry (Berberis vulgaris L. var. asperma). Sci. Hort. 238, 38–50. https://doi.org/10.1016/j.scienta.2018.04.040 (2018).
Mostafa, S., Hussein, B. A., Sayed, H. A., ElItriby, H. A. & Hussein, E. H. Genetic diversity assessment among some Ficus species using morphological characters and AFLPS. Plant Arch. 20 (1), 1395–1404 (2020).
Elwakil, H. E. et al. Genetical and morphological studies on Ficus trees. J. Adv. Agric. Res. 26 (2), 60–73. https://doi.org/10.21608/jalexu.2021.171573 (2021).
Khadivi, A., Mirheidari, F. & Moradi, Y. Multivariate analysis of Ficus benghalensis L. based on morphological characterizations. S. Afr. J. Bot. 150, 1080–1086. https://doi.org/10.1016/j.sajb.2022.09.025 (2022).
Khadivi, A., Mirheidari, F., Moradi, Y. & Paryan, S. Morphological variability of wild pomegranate (Punica granatum L.) accessions from natural habitats in the Northern parts of Iran. Sci. Hort. 264, 109165. https://doi.org/10.1016/j.scienta.2019.109165 (2020).
Khadivi, A., Mirheidari, F. & Moradi, Y. Morphological variation of Persian oak (Quercus brantii Lindl.) in Kohgiluyeh-va-Boyerahmad Province, Iran. Trees 1, 1–16. https://doi.org/10.1007/s00468-024-02528-3 (2024).
Khadivi, A. & Arab, M. Identification of the superior genotypes of pomegranate (Punica granatum L.) using morphological and fruit characters. Food Sci. Nutr. 9 (8), 4578–4588. https://doi.org/10.1002/fsn3.2450 (2021).
Pearson, K. I. Mathematical contributions to the theory of evolution—VII. On the correlation of characters not quantitatively measurable. Philos. Trans. R. Soc. Lond. 195 (262–273), 1–47. https://doi.org/10.1098/rsta.1900.0022 (1900).
Creswell, J. W. & Creswell, J. D. Research Design: Qualitative, Quantitative, and Mixed Methods Approaches (Sage, 2017).
Tabachnick, B. G., Fidell, L. S. & Ullman, J. B. Using Multivariate Statistics, vol. 6, 497–516 (Pearson, 2013).
Karimi, H. R. & Mirdehghan, S. H. Correlation between the morphological characters of pomegranate (Punica granatum) traits and their implications for breeding. Turk. J. Bot. 37 (2), 355–362. https://doi.org/10.3906/bot-1111-14 (2013).
Falconer, D. S. & Mackay, T. F. C. Introduction to Quantitative Genetics, 4 Edn (Longmans Green, 1996).
Tancred, S. J., Zeppa, A. G., Cooper, M. & Stringer, J. K. Heritability and patterns of inheritance of the ripening date of apples. HortScience 30 (2), 325–328. https://doi.org/10.21273/HORTSCI.30.2.325 (1995).
Cohen, J., Cohen, P., West, S. G. & Aiken, L. S. Applied Multiple Regression/Correlation Analysis for the Behavioral Sciences. https://doi.org/10.4324/9780203774441 (Routledge, 2013).
Hair, J. F., Black, W. C., Babin, B. J. & Anderson, R. E. Multivariate Data Analysis, 7th Edn (Pearson, 2010).
Montgomery, D. C., Peck, E. A. & Vining, G. G. Introduction to Linear Regression Analysis (Wiley, 2021).
Jolliffe, I. T. Principal Component Analysis for Special Types of Data 338–372. https://doi.org/10.1007/0-387-22440-8_13 (Springer, 2022).
Hastie, T., Tibshirani, R. & Friedman, J. The Elements of Statistical Learning: Data Mining, Inference, and Prediction. https://doi.org/10.1007/978-0-387-21606-5 (2009).
Jackson, J. E. A User’s Guide to Principal Components (Wiley, 2005).
Abdi, H. & Williams, L. J. Principal component analysis. Wiley Interdiscipl. Rev. Comput. Stat. 2 (4), 433–459. https://doi.org/10.1002/wics.101 (2010).
Kaiser, H. F. The varimax criterion for analytic rotation in factor analysis. Psychometrika 23 (3), 187–200. https://doi.org/10.1007/BF02289233 (1958).
Kaiser, H. F. The application of electronic computers to factor analysis. Educ. Psychol. Meas. 20 (1), 141–151. https://doi.org/10.1177/001316446002000116 (1960).
Shahkoomahally, S., Khadivi, A., Brecht, J. K. & Sarkhosh, A. Chemical and physical attributes of fruit juice and peel of pomegranate genotypes grown in Florida, USA. Food Chem. 342, 128302. https://doi.org/10.1016/j.foodchem.2020.128302 (2021).
Durgac, C. et al. Molecular and pomological diversity among pomegranate (Punica granatum L.) cultivars in Eastern mediterranean region of Turkey. Afr. J. Biotechnol. 7 (9), 1294–1301 (2008).
Wold, S., Esbensen, K. & Geladi, P. Principal component analysis. Chemometr. Intell. Lab. Syst. 2 (1–3), 37–52. https://doi.org/10.1016/0169-7439(87)80084-9 (1987).
Gabriel, K. R. The biplot graphic display of matrices with application to principal component analysis. Biometrika 58 (3), 453–467. https://doi.org/10.1093/biomet/58.3.453 (1971).
Johnson, R. A. & Wichern, D. W. Applied Multivariate Statistical Analysis, 6th edn (Pearson, 2014).
Hoover, W. E. Algorithms for confidence circles and ellipses. National Oceanic and Atlloapheric Adminlatntion (NOAA) Technical Report NOS 107 (1984).
Mardia, K. V., Kent, J. T. & Taylor, C. C. Multivariate Analysis, vol. 88 (Wiley, 2024).
Ward, J. H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 58 (301), 236–244. https://doi.org/10.1080/01621459.1963.10500845 (1963).
Jain, A. K. & Dubes, R. C. Algorithms for Clustering Data (Prentice-Hall, 1988).
Wilkinson, L. & Friendly, M. The history of the cluster heat map. Am. Stat. 63 (2), 179–184. https://doi.org/10.1198/tas.2009.0033 (2009).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study’s conception and design. FM, AS, YM, and AK experimented and collected data. YT analyzed data and wrote and edited the manuscript. AK edited the manuscript. All authors approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
For this study, we acquired permission to study pomegranate issued by the Agricultural and Natural Resources Ministry of Iran. The either cultivated or wild-growing plants sampled comply with relevant institutional, national, and international guidelines and domestic legislation of Iran.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Khadivi, A., Mirheidari, F., Saeidifar, A. et al. Morphological and pomological characterizations of pomegranate (Punica granatum L.) to select superior genotypes. Sci Rep 15, 7038 (2025). https://doi.org/10.1038/s41598-025-91595-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-91595-8
Keywords
This article is cited by
-
Multivariate assessment of morpho-biochemical and bioactive diversity in Syzygium cumini (L.) Skeels for the selection of superior genotype and breeding applications
BMC Plant Biology (2025)
-
Taxonomic significance of morphological and molecular variation in Egyptian Malvaceae species
BMC Plant Biology (2025)
-
Determination of gene association analysis of some leaf traits in mulberries using simple sequence repeat (SSR) markers
Genetic Resources and Crop Evolution (2025)
