
Abstract
Animal cognitive and behavioral testing often requires a reward to motivate task acquisition, yet reward perception may depend on reward type and previous experience. We first assessed the effects of reward types on initial discrimination and reversal learning ability in weaned dairy calves, with calves tested in a T-maze containing either a food, social, or exit reward. During the initial stage, success rate was greater for calves provided the social vs. food reward, whereas success in the reversal stage was greater for calves provided the exit vs. social reward. We also assessed effects of social experience in calves previously housed individually or in pairs tested with either a social or exit reward. Interactive effects of previous housing and reward type were evident during the initial discrimination stage of testing, with previously pair housed calves relatively more successful if given the exit reward and individually housed calves relatively more successful if given the social reward. During the reversal stage, success rate was greater for calves given the exit reward, particularly if previously housed individually. These results suggest that previous social experience can affect the subjective perception of different reward types, and that the subjective valuation of an exit reward may uniquely increase with repeated testing.
Similar content being viewed by others

Introduction
The evaluation of animal cognition has interdisciplinary applications, with uses in a variety of fields ranging from wildlife conservation1to human neuroscience2. In recent years the assessment of cognition in agricultural animals has gained interest due to potential implications for animal performance and welfare. For instance, dairy cattle routinely encounter potential stressors such as housing and management transitions, which can cause undesirable physiological responses such as decreased immune competence, health status, and milk yield3,4, and negatively impact affective state5,6. Individuals that are more cognitively flexible, or better able to mentally and behaviorally adapt to environmental change7,8, may be less affected by potential stressors and thus may see a comparative improvement in welfare and performance9. For example, dairy calves that were seen to be more cognitively flexible in a behavioral test subsequently adjusted their behavior more rapidly when turned out to pasture and simultaneously regrouped, demonstrating greater exploration of the environmental space and a relatively lower latency to feed10. As such, providing an environment that enables adequate cognitive development may be important, and research has suggested that aspects of the early life environment such as social housing11, feeding12, and environmental enrichment13 can affect dairy calf cognitive flexibility.
Cognitive flexibility is often assessed using reversal learning tasks in which an animal learns a rule to obtain a reward and then must adapt to obtain the reward once the rule has changed14. Importantly, the reward provided within these assessments must be sufficiently hedonic to motivate a desire to obtain it15. Food is often used as a reward within such tasks across species. For example, cattle may be offered grain-based concentrates16,17, sweetened liquid solutions18,19,20, or milk if at the pre-weaning stage of life11,12,21,22. Though frequently used, the motivational properties of food may be inconsistent, altered by factors such as hunger levels and the perceived value of the food relative to availability of food sources in the environment (e.g. in chickens23; pigs24; and cattle25). Consequently, tests using food may require prior food restriction to initiate hunger to be sufficiently motivating, which can negatively affect learning ability (e.g. in chickens26; and calves27), and can be detrimental to animal welfare28.
Alternatives to food rewards for behavioral testing have received less attention. For example, social contact may be considered rewarding (e.g. in calves29; and sheep30,31) yet little appears to be known about the potential confounding effects of prior experience when using this type of reward. An important management consideration affecting dairy calves is the provision of social contact during early life, which remains variable on commercial dairy farms. Though early life social housing is being increasingly utilized, it is still common practice for dairy calves to be housed individually from birth until they are weaned32. Provision of social companions may affect general success during maze testing (e.g. in rats33; and calves11), but the effects of previous social experience on the perception of social rewards is unclear. Considering that social housing has been seen to affect calf social behavior (e.g. see34,35,36,37), it may also affect individual motivation to gain access to social contact in a behavioral testing scenario.
Another potential form of reward is the ability to exit the behavioral test. This reward type may be effective if animals are motivated to regain access to a more familiar environment or to escape a testing scenario that is perceived as aversive. This type of reward has been utilized for some species (e.g. pigs38; squirrels39; and guppies40), though it has not to our knowledge been previously used as a form of reward for cattle. This type of reward may avoid biases related to the perception of food and social contact due to prior experience and thus may offer a promising alternative to food and social rewards.
The first aim of this study was to assess the effects of different reward types on the learning ability of dairy calves during a test of cognitive flexibility comprised of an initial discrimination stage and a reversal learning stage. We investigated the efficacy of three reward types, including the commonly used food reward, as well as a social reward (access to a familiar companion) and an exit reward (ability to leave the testing facility) as alternatives. We predicted that all three reward types may enable successful cue discrimination but had no specific predictions as to the relative effects of different reward types. The second aim of this study was to assess the effects of previous social experience on the efficacy of social rewards within the same test. Considering previous evidence of reduced social behavior in individually reared calves36,37,41, we hypothesized that previously individually reared calves may be less successful when provided the social reward than previously socially housed calves.
Results
Aim 1: comparison of food, social contact, and exit rewards
During the initial discrimination stage of testing, calves that received the social reward required the fewest sessions to pass and had the highest overall success rate, while calves that received the food reward had the lowest rate of success (Kaplan-Meier estimates; Table 1). Reward type was seen to affect the overall learning ability of calves (P = 0.02; Cox proportional hazard estimates; Table 2), where the likelihood of success significantly differed between calves receiving either the social or food reward (P = 0.007), with the exit reward yielding intermediate outcomes. Hazard ratio estimates indicated that, for any given session, calves that received the social reward were 6.5 times more likely to pass than those that received the food reward (Table 2; Fig. 1a). Birthweight also predicted success at this stage (P < 0.001), with hazard ratios indicating that for every 1 kg increase in birthweight calves were 1.3 times more likely to succeed during any given session (Table 2).
During the reversal learning stage of testing, calves that received the exit reward required the fewest sessions to pass and had the highest overall success rate (Kaplan-Meier estimates; Table 1). Calves that received the exit reward had a significantly greater chance of success than those that received the social reward (P = 0.04; Cox proportional hazard estimates; Table 2), with the food reward yielding intermediate outcomes. Hazard ratio estimates indicated that, for any given session, calves that received the exit reward were 4.1 times more likely to succeed than those that received the social reward (Table 2; Fig. 1b). Birthweight also predicted success at this stage (P = 0.03), with hazard ratios indicating that for every 1 kg increase in birthweight calves were 0.7 times as likely to succeed during any given session (Table 2).

Cox proportional hazards probability estimates for number of sessions required to pass A) the initial discrimination stage of testing, and B) the reversal stage of testing, for calves provided different reward types (social, exit, or food; Aim 1).
Aim 2: interactive effects of previous social housing and reward type
During the initial discrimination stage of testing, previously pair-housed calves that received the exit reward required the fewest sessions to pass and had the highest overall success rate, while previously individually housed calves that received the exit reward required the greatest number of sessions to pass and had the lowest overall success rate (Kaplan-Meier estimates; Table 1). Correspondingly, there was an interactive effect of housing treatment and reward type on learning ability (P = 0.03; Cox proportional hazards estimates; Table 2; Fig. 2a), with no main effects of housing treatment (P = 0.12) or reward type (P = 0.13). Birthweight also predicted success at this stage (P = 0.005), with hazard ratios indicating that for every 1 kg increase in birthweight calves were 1.2 times more likely to succeed during any given session (Table 2).
During the reversal learning stage of testing, calves provided the exit reward required the fewest sessions to pass and had the highest overall success rate, while individually reared calves that received the social reward required the greatest number of sessions to pass and had the lowest overall rate of success (Kaplan-Meier estimates; Table 1). There was a main effect of reward type on learning ability (P = 0.02; Cox proportional hazard estimates; Table 2), particularly within previously individually housed calves, with hazard ratio estimates indicating that, for any given session, individually reared calves that received the exit reward were 5.8 times more likely to succeed than those that received the social reward (P = 0.02; Figure 2b). In contrast, the success of previously pair-housed calves was not affected by reward type (P= 0.72). There was no main effect of previous housing treatment on overall success (P = 0.43). Birthweight again predicted success at this stage (P= 0.02), with hazard ratios indicating that for every 1 kg increase in birthweight calves were 0.8 times as likely to succeed during any given session (Table 2)

Cox proportional hazards probability estimates for number of sessions required to pass A) the initial discrimination stage of testing, and B) the reversal stage of testing, for calves previously housed individually or in pairs and assigned to receive either the social or exit reward.
Discussion
The evaluation of livestock cognition can provide information relevant for the assessment and improvement of agricultural animal welfare and performance, yet established methodology is lacking across species and age classes. This research aimed to compare the suitability of different reward types for use in dairy calf cognitive and behavioral testing, and to determine the effects of previous social experience on the perception of social rewards.
In comparing reward types, we found that initial discriminative learning was generally improved through provision of a social or exit reward rather than a food reward. This result aligns with previous research suggesting that animals may show preferences for social contact over food (e.g. in calves42, chimpanzees43and rabbits44), though the extent to which these findings may be generalized may vary with study conditions. Because feed restriction can be detrimental to welfare and negatively affect learning, we did not restrict intake prior to testing26,27and thus our results may reflect a lack of hunger25. Alternatively, it is possible that the perceived value of the food reward was insufficient to motivate task completion, as the desire to obtain food, though often driven by metabolic need, can also arise from the hedonic pleasure derived from eating45. As such, providing a highly preferred feed type may be sufficiently rewarding even in the absence of feed restriction (e.g., in chickens46), although a suitable option in this age class of cattle would require further study.
Considering established effects of early social contact on behavioral development, we additionally investigated the effects of early social housing on response to different reward types. During the initial learning stage, we found evidence of an interactive effect of previous social housing on response to reward, where calves reared with social contact from birth were relatively more successful when provided the exit reward rather than the social reward, while calves reared individually were relatively more successful when given the social reward rather than the exit reward. These findings may suggest effects of previous social experience on reward perception that may require consideration when assessing cognition. Although some previous studies have suggested that individually reared calves may approach other calves less readily due to increased fearfulness (e.g. see35,41), our findings align with observations of a rebound effect in socialization, where motivation to access social contact may increase after periods of social deprivation47. For example, rats subjected to social isolation were seen to choose social interaction over food more than rats that were socially raised, even after food deprivation48, while stallions housed in isolation for 9 months engaged in more frequent positive social interactions and maintained closer proximity with conspecifics when grouped at pasture than stallions that were previously socially housed49.
Effects of reward types differed during the reversal learning stage compared to the initial discrimination stage of testing, with success rates greatest for calves that received the exit reward and lowest for calves that received the social reward. We suggest that a possible explanation for the different effects of reward type between stages could be altered motivation to access the reward. The stimulus-response link connecting a reward with an individual’s desire to obtain it can change as a function of experience50, and typically repeated consumption of a reward reduces its subjective valuation51. While the desire to obtain rewards such as food or social contact may decrease with repeated testing as hunger or social motivation diminish, reward satiation depends on the individual properties of the reward51. If calves become increasingly disinterested or even frustrated with repeated testing, the subjective valuation of the exit reward may increase, thus offering potentially unique and advantageous motivational properties. However, the generalizability of this finding will depend on how animals perceive the testing area relative to the exit area, and how this perception changes over time. In the present study, heifers may have been particularly motivated to exit to regain visual access to their home pen and social companions, but it is possible that different testing scenarios may yield different motivations. An exit alone, without simultaneous access to a positively perceived stimulus upon exiting, may lower motivation to exit. Further, habituation through repeated testing may decrease the aversiveness of the test environment, resulting in the test being perceived as progressively less aversive, and indeed animals may even become increasingly motivated to participate if the test itself is perceived as rewarding (e.g. see17).
Alternatively, it is also possible that the characteristics of each reward type may differentially affect success across stages, as the reversal learning stage seeks to assess cognitive flexibility while the initial learning stage focuses on the ability to correctly discriminate cues. Cognitive flexibility can be impaired by physiological arousal or stress52,53, which could be elicited by features of the reward itself. We found that the success of previously individually housed calves was sensitive to reward type during the reversal learning stage, with lower success rates seen when calves were provided the social reward. In contrast, we saw no significant effect of reward type on reversal learning outcomes in previously pair-housed calves. It is possible that the presence of the stimulus calf provided as the social reward could evoke stress or fear for calves with less previous social experience35, consequently diminishing cognitive flexibility. These effects are also subject to potential nuisance variables such as the unique relationships between focal calves and calves used as a social reward or calves situated in the home pen, as well as the behavior of these other calves during testing. Individual difference in personality must also be considered, as calves have been seen to show individual differences in traits such as exploratory behavior, fearfulness, and sociability54,55, which may also affect testing outcomes and responses to different reward types.
An incidental finding of this study was that birthweight (but not body weight concurrent with testing) was a significant predictor of testing success. We found that birthweight positively correlated with success during the initial stage of testing yet was negatively correlated during the reversal stage, as has previously been seen in piglets (initial stage56,57,58,59; reversal stage60,61). Taken together, these results suggest that lower birthweight animals may experience poorer spatial learning and memory yet demonstrate improved cognitive flexibility when compared with their higher birthweight counterparts. Possible mechanisms underlying cognitive deficiencies in lower birthweight individuals, as seen within our initial stage of testing, could include neurodevelopmental impairment during late natal and early post-natal periods57,62,63,64, a comparative reduction in overall brain volume60,65,66,67, a compensatory growth-performance trade-off68, or an increase in levels of chronic stress due to lower social standing or due to a reduced capacity to compete for resources58,69. Potential explanations for improvements in cognitive abilities due to lower birthweights, as seen within our reversal stage of testing, may include a stronger motivation to obtain rewards60, a decrease in chronic stress due to a blunted stress response60,70, or brain pre-conditioning, in which sublethal insults to the brain (such as an insufficient supply of blood or oxygen in-utero) may result in increased protection against subsequent stressors60,71. While previous literature offers some potential mechanisms that may explain our results, this incidental finding requires further targeted research to confirm and explore the relationship between birthweight and cognitive abilities in dairy calves.
In conclusion, we investigated the effects of different reward types on cognitive testing outcomes in weaned dairy calves and explored potential interactive effects of previous social experience on the perception of rewards. Our results suggest that a novel exit reward was effective and may be valuable for use in future dairy cattle cognitive and behavioral testing. Further, our results suggest that social experience can affect the subjective perception of rewards, possibly increasing sensitivity to social contact provided as a reward.
Materials and methods
Animal management and experimental design
All study procedures were reviewed and approved by the University of Florida Institutional Animal Care and Use Committee (protocol #201910617) and all experiments were conducted in accordance with institutional guidelines and regulations. A total of 50 female Holstein calves were enrolled into the study at approximately 24 h of age at the University of Florida Dairy Research Unit (Hague, Florida, USA). Calves were provided ad-libitum access to water and pelleted calf starter (22% CP and 2% fat, Ampli-Calf Starter Warm Weather 20, Purina Animal Nutrition LLC, Shoreview, MN) from birth. Calves were given 4–8 L of quality-controlled colostrum within 24 h of birth and then received milk replacer (28% CP and 20% fat; Suwanee Valley Feeds, Trenton, FL) from a teat bucket (Peach Teat Limited, Christchurch, New Zealand) at a rate of 8 L/d in 2 feedings. At 6 weeks of age, calves were weaned according to a 10-d step-down protocol (6 L/d in 2 meals on days 1–4, 4 L/d in 2 meals on days 5–8, and 2 L/d in 2 meals on days 9–10). Calves received weekly clinical examinations by a veterinarian, and daily health checks by farm and research personnel.
Calves were randomly assigned to one of two housing treatments within 72 h of birth: individual housing (n = 30 calves) or pair housing (n = 20 calves; 1 focal calf/pen, paired with another calf born within 48 h). Calves were housed in wire mesh pens (dimensions 0.9 × 1.8 m for individual pens; 1.8 × 1.8 m for pair pens), located under an open-sided barn protecting from downwards rain and wind, bedded with sand that was replaced weekly. Pens were interspersed between individual and pair pens, spaced to provide visual but not physical contact between adjacent pens. At 2 d postweaning (56 ± 6 d of age), calves were mingled between previous housing treatments and group-housed in the same open-sided barn (3–6 calves/pen; pen dimension 3.7 × 8.0 m). At 9 d postweaning (63 ± 6 d of age), calves were moved into a group pen on pasture (10 calves/group; dimension 15.2 × 45.7 m). The pen provided a sheltered feeding area with ad-libitum access to calf starter and water. Calves were weighed using a floor scale (PS-2000 scale, Brecknell Scales, Fairmont, MN, USA) at birth and again when turned out to pasture (63 ± 6 d of age).
After 10 d habituation to the pasture pen (73 ± 6 d of age) calves were subject to a test designed to assess cognitive flexibility, described in more detail below. For Aim 1, previously individually housed calves were tested with either a food reward (n = 10 calves), a social reward (n = 10 calves), or an exit reward (n = 10 calves). For Aim 2, previously pair housed calves were tested with either a social reward (n = 10 calves) or an exit reward (n = 10 calves) and compared with a subset of previously individually housed calves assigned to the corresponding reward types for Aim 1.
Data collection
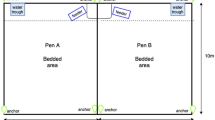
To assess cognitive flexibility, we used a T-maze test, a two-armed maze in which a reward was spatially located in one of the arms during an initial discrimination learning phase, and subsequently relocated to the other arm during a reversal learning phase8. The test design was similar to a previous study in pigs72, subsequently adapted for use in pre-weaned calves12. The maze, which bordered the subjects’ home pen (Fig. 3), was constructed using cow gates covered with opaque black corrugated plastic to create a visual barrier to both the outside of the maze and to the rewards from the base of the T (height 1.83 m). The reward location was evenly counterbalanced across subjects and reward types to prevent effects of side preferences due to laterality73.

Diagram of T-Maze location and dimensions.
Prior to their first testing session, calves were escorted through the maze, firstly to the unrewarded arm, and then to the rewarded arm, where they were allowed to interact with their designated reward, and then to exit via the rewarded arm. Calves were then given their first testing session, in which they were released into the base of the T-maze through an opaque door that was immediately closed behind them. Each session lasted for a maximum of 3 min, or until any part of the calf surpassed the maze exit (only applicable for exit reward). To successfully ‘pass’ each stage of the maze, calves had to move directly to the arm of the maze containing the reward (without first going to the unrewarded arm) for 3 consecutive sessions, a criterion based on a previous study which found that after successfully completing these types of sessions 3 times calves no longer made regressive errors12. Once calves successfully reached criterion for passing the initial learning stage the reward side was switched, and the same criterion for success applied to the reversal learning stage. Calves were allowed a maximum of 18 sessions to meet criterion during each stage. Calves received 5 sessions/d for a maximum of 6 days and were given an additional sixth session on any given day if they correctly navigated the fourth and fifth sessions to enable them the opportunity to meet criterion that day.
For the food reward, an opaque bucket containing 1 cup of pelleted starter (familiar, provided in regular diet) mixed with 1 handful of chopped hay (unfamiliar, not provided prior to testing) was located on the ground in the rewarded arm of the maze (Fig. 4a). Hay was selected as a form of food reward given evidence that heifers are motivated to obtain forage74, and may consume hay preferentially at the time of feed delivery75, while the starter pellet was offered simultaneously as a more familiar food option to minimize the potential effects of food neophobia. Buckets containing the same food reward were also placed just outside of the non-rewarded arm to balance olfactory cues. For the social reward, a non-study stimulus calf from the same home pasture (first introduced to all test calves during post-weaning grouping) was placed in a holding area in the rewarded arm of the maze (Fig. 4b). The stimulus calf was enclosed by a gate that enabled the test calf visual and tactile contact, while preventing movement of the stimulus calf around the maze. Another non-study calf was placed in the holding pen in the unrewarded side of the maze to balance auditory and olfactory cues, this time with an opaque plastic barrier preventing visual and tactile contact. For the exit reward, a gate half the width of the arm of the maze was left open, allowing calves to exit the maze into a holding area located adjacent to the home pasture, enabling the calf visual contact with the home pen and pen mates (Fig. 4c).

Diagram of T-Maze set-up for (A) food reward, (B) social reward, and (C) exit reward (including indication of the location of home pen relative to the holding area the calves entered after exiting the maze). Solid lines represent opaque fencing. Dashed lines represent non-opaque fencing enabling visual and tactile contact between areas. Reward side was evenly counterbalanced for each reward type.
Statistical analysis
Statistical analyses were performed using SAS (Version: OnDemand for Academics; SAS Institute Inc., Cary, NC, USA), with calf as the experimental unit. Survival analyses were performed separately by stage of testing, using time-to-event models fitted to the data to quantify the effect of explanatory variables on the number of sessions required (latency) to reach the passing criterion (event) for each stage of the behavioral test76. Kaplan-Meier survival estimates were produced to enable initial data screening within the confounds of a survival analysis model (PROC LIFETEST). Cox proportional hazards models were subsequently used to implement a more complete model enabling inclusion of multiple predictor variables and interaction terms where appropriate (PROC PHREG). Calves that did not meet the criterion within the allotted timeframe were included as censored values within both types of analysis.
The Cox proportional hazards model parameterizes the hazard function \(\lambda (t,x)\) as
where \(\lambda _{0} (t)\) is the baseline hazard function (hazard for individuals with all covariates equal to zero) and the exponential function captures the proportional association between covariates and hazard. Thus, \(\lambda (t,x)\) represents the hazard of the event at time (i.e., likelihood of passing the task if they haven’t already) for heifers with covariate values \(x\)1,…,\(x\). The set of covariates \(x\) used in the model for Aim 1 were reward type (food, social, exit), and birthweight. The set of covariates \(\:x\) used in the model for Aim 2 were reward type (social, exit), previous housing treatment (individually housed, pair housed), the interaction between reward type and previous housing treatment, and birthweight, which was considered as a predictor variable given previous evidence of effects of birthweight on later cognition in other species (e.g. humans77, and pigs56,57,58,59,60). ANOVA tests were used to confirm that birthweight (mean ± SD = 38 ± 4 kg) did not differ between reward types or housing treatments prior to model inclusion. Additional variables that were also initially considered as covariates included reward side (given potential effects of side preferences due to laterality78), season of testing (given possible effects of heat stress on activity levels79), and bodyweight at the time of testing (temporarily replacing birthweight within models to determine that the effects of birthweight were not due to body size at the time of testing). These variables were not seen to be significant or to improve model fit and so were not retained in the final models. Week of testing (corresponding to concurrent turnout group; 1–4 heifers tested per week) was also initially included as a frailty in the models. However, there was no significant random effect of this variable (as indicated by insignificant estimates for the associated variance, and resulting in only minor changes to standard errors upon the use of robust variance) and thus it was also not included within the final models. The assumption of proportional hazards was checked using a Kolmogorov-Smirnov test, and by plotting the time component of the model against the Schoenfeld residuals to visually check that the correlation produced an approximately zero slope. A plot of the deviance residuals set against the linear predictors was used to assess model fit and presence of outliers. Significance was declared at P < 0.05.
Data availability
Data are available upon request to the corresponding author (emillerc@ufl.edu).
References
Greggor, A. L., Clayton, N. S., Phalan, B. & Thornton, A. Comparative cognition for conservationists. Trends Ecol. Evol. 29, 489–495 (2014).
Carlson, B. A. Diversity matters: the importance of comparative studies and the potential for synergy between neuroscience and evolutionary biology. Arch. Neurol. 69, 987–993 (2012).
Keeling, L. & Jensen, P. Abnormal behaviour, stress and welfare. in The ethology of domestic animals: an introductory text, modular texts (ed. Jensen, P.) 85–101CABI, https://doi.org/10.1079/9781845935368.0085 (2009).
Moberg, G. P. CABI,. Biological response to stress: implications for animal welfare. in The biology of animal stress: basic principles and implications for animal welfare. (eds. Moberg, G. P. & Mench, J. A.) 1–21 . https://doi.org/10.1079/9780851993591.0001 (2000).
Broom, D. M. & Johnson, K. G. Stress and animal welfareSpringer Netherlands,. https://doi.org/10.1007/978-94-024-0980-2 (1993).
Clark, J. D., Rager, D. R. & Calpin, J. P. Animal well-being II. Stress and distress. Comp. Med. 47, 571-579 (1997).
Scott, W. A. Cognitive complexity and cognitive flexibility. Sociometry 25, 405 (1962).
Brown, V. J. & Tait, D. S. Encyclopedia of Psychopharmacology (Springer Science & Business Media, 2010).
Koolhaas, J. M. et al. Coping styles in animals: current status in behavior and stress-physiology. Neurosci. Biobehav Rev. 23, 925–935 (1999).
Horvath, K. C. & Miller-Cushon, E. K. Characterizing social behavior, activity, and associations between cognition and behavior upon social grouping of weaned dairy calves. J. Dairy. Sci. 101, 7287–7296 (2018).
Gaillard, C., Meagher, R. K., von Keyserlingk, M. A. G. & Weary, D. M. Social housing improves dairy calves’ performance in two cognitive tests. PLoS ONE. 9, e90205 (2014).
Horvath, K., Fernandez, M. & Miller-Cushon, E. K. The effect of feeding enrichment in the milk-feeding stage on the cognition of dairy calves in a T-maze. Appl. Anim. Behav. Sci. 187, 8–14 (2017).
Zhang, C., Juniper, D. T. & Meagher, R. K. Effects of physical enrichment and pair housing before weaning on growth, behaviour and cognitive ability of calves after weaning and regrouping. Appl. Anim. Behav. Sci. 249, 105606 (2022).
Gelfo, F. Does experience enhance cognitive flexibility? An overview of the evidence provided by the environmental enrichment studies. Front. Behav. Neurosci. 13, 150 (2019).
Berridge, K. C. From prediction error to incentive salience: mesolimbic computation of reward motivation. Eur. J. Neurosci. 35, 1124–1143 (2012).
Hirata, M., Tomita, C. & Yamada, K. Use of a maze test to assess Spatial learning and memory in cattle: can cattle traverse a complex maze? Appl. Anim. Behav. Sci. 180, 18–25 (2016).
Meagher, R. K., Strazhnik, E., von Keyserlingk, M. A. G. & Weary, D. M. Assessing the motivation to learn in cattle. Sci. Rep. 10, 6847 (2020).
Hellekant, G., Hård af Segerstad, C. & Roberts, T. W. Sweet taste in the calf: III. Behavioral responses to sweeteners. Physiol. Behav. 56, 555–562 (1994).
Ginane, C., Baumont, R. & Favreau-Peigné, A. Perception and hedonic value of basic tastes in domestic ruminants. Physiol. Behav. 104, 666–674 (2011).
Lecorps, B., Nogues, E., von Keyserlingk, M. A. G. & Weary, D. M. Pessimistic dairy calves are more vulnerable to pain-induced anhedonia. PLoS ONE. 15, e0242100 (2020).
Lecorps, B., Woodroffe, R. E., von Keyserlingk, M. A. G. & Weary, D. M. Assessing cognitive performance in dairy calves using a modified hole-board test. Anim. Cogn. https://doi.org/10.1007/s10071-022-01617-5 (2022).
Meagher, R. K. et al. Effects of degree and timing of social housing on reversal learning and response to novel objects in dairy calves. PLoS ONE. 10, e0132828 (2015).
Bokkers, E. A. M., Koene, P., Rodenburg, T. B., Zimmerman, P. H. & Spruijt, B. M. Working for food under conditions of varying motivation in broilers. Anim. Behav. 68, 105–113 (2004).
Lawrence, A. B. & Illius, A. W. Methodology for measuring hunger and food needs using operant conditioning in the pig. Appl. Anim. Behav. Sci. 24, 273–285 (1989).
Schütz, K., Davison, D. & Matthews, L. Do different levels of moderate feed deprivation in dairy cows affect feeding motivation? Appl. Anim. Behav. Sci. 101, 253–263 (2006).
Buckley, L. A. et al. Too hungry to learn? Hungry broiler breeders fail to learn a Y-maze food quantity discrimination task. Anim Welf. 20, 469–481 (2011).
Lecorps, B., Woodroffe, R. E., von Keyserlingk, M. A. G. & Weary, D. M. Hunger affects cognitive performance of dairy calves. Biol. Lett. 19, 20220475 (2023).
D’Eath, R. B., Tolkamp, B. J., Kyriazakis, I. & Lawrence, A. B. Freedom from hunger’ and preventing obesity: the animal welfare implications of reducing food quantity or quality. Anim. Behav. 77, 275–288 (2009).
Holm, L., Jensen, M. B. & Jeppesen, L. L. Calves’ motivation for access to two different types of social contact measured by operant conditioning. Appl. Anim. Behav. Sci. 79, 175–194 (2002).
Hunter, D. S. et al. Do I turn left or right? Effects of sex, age, experience and exit route on maze test performance in sheep. Physiol. Behav. 139, 244–253 (2015).
Lee, C., Colegate, S. & Fisher, A. D. Development of a maze test and its application to assess Spatial learning and memory in Merino sheep. Appl. Anim. Behav. Sci. 96, 43–51 (2006).
Costa, J. H. C., von Keyserlingk, M. A. G. & Weary, D. M. Invited review: effects of group housing of dairy calves on behavior, cognition, performance, and health. J. Dairy. Sci. 99, 2453–2467 (2016).
Einon, D. Spatial memory and response strategies in rats: age, sex and rearing differences in performance. Q. J. Exp. Psychol. 32, 473–489 (1980).
Chua, B., van Coenen, E., Weary, D. M. & D. J. & Effects of pair versus individual housing on the behavior and performance of dairy calves. J. Dairy. Sci. 85, 360–364 (2002).
De Paula Vieira, A., de Passillé, A. M. & Weary, D. M. Effects of the early social environment on behavioral responses of dairy calves to novel events. J. Dairy. Sci. 95, 5149–5155 (2012).
Duve, L. R. & Jensen, M. B. The level of social contact affects social behaviour in pre-weaned dairy calves. Appl. Anim. Behav. Sci. 135, 34–43 (2011).
Duve, L. R. & Jensen, M. B. Social behavior of young dairy calves housed with limited or full social contact with a peer. J. Dairy. Sci. 95, 5936–5945 (2012).
Jansen, J., Bolhuis, J. E., Schouten, W. G. P., Spruijt, B. M. & Wiegant, V. M. Spatial learning in pigs: effects of environmental enrichment and individual characteristics on behaviour and performance. Anim. Cogn. 12, 303–315 (2009).
Marks, K. A., Vizconde, D. L., Gibson, E. S., Rodriguez, J. R. & Nunes, S. Play behavior and responses to novel situations in juvenile ground squirrels. J. Mammal. 98, 1202–1210 (2017).
Lucon-Xiccato, T. & Bisazza, A. Complex maze learning by fish. Anim. Behav. 125, 69–75 (2017).
Lindner, E. E., Gingerich, K. N. & Miller-Cushon, E. K. Effects of early social contact on dairy calf response to initial social grouping and regrouping. J. Dairy. Sci. 104, 10090–10099 (2021).
Lensink, J., Veissier, I. & Boissy, A. Enhancement of performances in a learning task in suckler calves after weaning and relocation: motivational versus cognitive control? Appl. Anim. Behav. Sci. 100, 171–181 (2006).
Mason, W. A., Saxon, S. V. & Sharpe, L. G. Preferential responses of young chimpanzees to food and social rewards. Psychol. Rec. 13, 341–345 (1963).
Seaman, S. C., Waran, N. K., Mason, G. & D’Eath, R. B. Animal economics: assessing the motivation of female laboratory rabbits to reach a platform, social contact and food. Anim. Behav. 75, 31–42 (2008).
Recio-Román, A., Recio-Menéndez, M. & Román-González, M. V. Food reward and food choice. an inquiry through the liking and wanting model. Nutrients 12, (2020).
Angevaare, M. J., Prins, S., van der Staay, F. J. & Nordquist, R. E. The effect of maternal care and infrared beak trimming on development, performance and behavior of silver Nick hens. Appl. Anim. Behav. Sci. 140, 70–84 (2012).
Dellmeier, G. R., Friend, T. H. & Gbur, E. E. Comparison of four methods of calf confinement. II. Behavior. J. Anim. Sci. 60, 1102–1109 (1985).
Ikemoto, S. & Panksepp, J. The effects of early social isolation on the motivation for social play in juvenile rats. Dev. Psychobiol. 25, 261–274 (1992).
Christensen, J. W., Ladewig, J., Søndergaard, E. & Malmkvist, J. Effects of individual versus group stabling on social behaviour in domestic stallions. Appl. Anim. Behav. Sci. 75, 233–248 (2002).
Toates, F. The interaction of cognitive and stimulus-response processes in the control of behaviour. Neurosci. Biobehav Rev. 22, 59–83 (1998).
Pastor-Bernier, A., Stasiak, A. & Schultz, W. Reward-specific satiety affects subjective value signals in orbitofrontal cortex during multicomponent economic choice. Proc. Natl. Acad. Sci. USA 118, e2022650118 (2021).
Park, J. & Moghaddam, B. Impact of anxiety on prefrontal cortex encoding of cognitive flexibility. Neuroscience 345, 193–202 (2017).
George, S. A. et al. Alterations in cognitive flexibility in a rat model of post-traumatic stress disorder. Behav. Brain Res. 286, 256–264 (2015).
Neave, H. W., Costa, J. H. C., Weary, D. M. & von Keyserlingk, M. A. G. Personality is associated with feeding behavior and performance in dairy calves. J. Dairy. Sci. 101, 7437–7449 (2018).
Lecorps, B., Kappel, S., Weary, D. M. & von Keyserlingk, M. A. G. Dairy calves’ personality traits predict social proximity and response to an emotional challenge. Sci. Rep. 8, 16350 (2018).
Gieling, E. T., Park, S. Y., Nordquist, R. E. & van der Staay, F. J. Cognitive performance of low- and normal-birth-weight piglets in a Spatial hole-board discrimination task. Pediatr. Res. 71, 71–76 (2012).
Radlowski, E. C. et al. A neonatal piglet model for investigating brain and cognitive development in small for gestational age human infants. PLoS ONE. 9, e91951 (2014).
Roelofs, S., van Bommel, I., Melis, S., van der Staay, F. J. & Nordquist, R. E. Low birth weight impairs acquisition of Spatial memory task in pigs. Front. Vet. Sci. 5, 142 (2018).
Roelofs, S. et al. Discrimination learning and judgment bias in low birth weight pigs. Anim. Cogn. 22, 657–671 (2019).
Antonides, A., Schoonderwoerd, A. C., Nordquist, R. E. & van der Staay, F. J. Very low birth weight piglets show improved cognitive performance in the Spatial cognitive holeboard task. Front. Behav. Neurosci. 9, 43 (2015).
Murphy, E., Kraak, L., Nordquist, R. E. & van der Staay, F. J. Successive and conditional discrimination learning in pigs. Anim. Cogn. 16, 883–893 (2013).
Arcangeli, T., Thilaganathan, B., Hooper, R., Khan, K. S. & Bhide, A. Neurodevelopmental delay in small babies at term: a systematic review. Ultrasound Obstet. Gynecol. 40, 267–275 (2012).
Camprubí Camprubí, M. et al. Learning and memory disabilities in IUGR babies: functional and molecular analysis in a rat model. Brain Behav. 7, e00631 (2017).
Vallet, J. L. & Miles, J. R. Comparison of myelination between large and small pig fetuses during late gestation. Anim. Reprod. Sci. 132, 50–57 (2012).
Buechel, S. D., Boussard, A., Kotrschal, A. & van der Bijl, W. & Kolm, N. Brain size affects performance in a reversal-learning test. Proc. Biol. Sci. 285, (2018).
Martinussen, M. et al. Segmental brain volumes and cognitive and perceptual correlates in 15-year-old adolescents with low birth weight. J. Pediatr. 155, 848–853e1 (2009).
Taylor, H. G. et al. Brain volumes in adolescents with very low birth weight: effects on brain structure and associations with neuropsychological outcomes. Dev. Neuropsychol. 36, 96–117 (2011).
Fisher, M. O., Nager, R. G. & Monaghan, P. Compensatory growth impairs adult cognitive performance. PLoS Biol. 4, e251 (2006).
Sandi, C. Stress and cognition. Wiley Interdiscip Rev. Cogn. Sci. 4, 245–261 (2013).
Schäffer, L. et al. Blunted stress response in small for gestational age neonates. Pediatr. Res. 65, 231–235 (2009).
Stetler, R. A. et al. Preconditioning provides neuroprotection in models of CNS disease: paradigms and clinical significance. Prog Neurobiol. 114, 58–83 (2014).
Bolhuis, J. E., Schouten, W. G. P., de Leeuw, J. A., Schrama, J. W. & Wiegant, V. M. Individual coping characteristics, rearing conditions and behavioural flexibility in pigs. Behav. Brain Res. 152, 351–360 (2004).
Hosoi, E., Rittenhouse, L. R., Swift, D. M. & Richards, R. W. Foraging strategies of cattle in a Y-maze: influence of food availability. Appl. Anim. Behav. Sci. 43, 189–196 (1995).
Van Os, J. M. C., Mintline, E. M., DeVries, T. J. & Tucker, C. B. Domestic cattle (Bos Taurus Taurus) are motivated to obtain forage and demonstrate contrafreeloading. PLoS ONE. 13, e0193109 (2018).
Miller-Cushon, E. K., Bergeron, R., Leslie, K. E., Mason, G. J. & DeVries, T. J. Effect of feed presentation on feeding patterns of dairy calves. J. Dairy. Sci. 96, 7260–7268 (2013).
Cox, D. R. Regression models and Life-Tables. J. Roy. Stat. Soc.: Ser. B (Methodol.). 34, 187–202 (1972).
Farajdokht, F. et al. Very low birth weight is associated with brain structure abnormalities and cognitive function impairments: A systematic review. Brain Cogn. 118, 80–89 (2017).
Robins, A. & Phillips, C. Lateralised visual processing in domestic cattle herds responding to novel and familiar stimuli. Laterality 15, 514–534 (2010).
Herbut, P. et al. The effects of heat stress on the behaviour of dairy cows – a review. Annals Anim. Sci. 21, 385–402 (2021).
Acknowledgements
We thank the staff at the University of Florida Dairy Research Unit, in particular Jesse Hooten and Sarah Norris for technical assistance. We acknowledge assistance of the many student research assistants that helped collect data, in particular Daniel Clein, Maria Belen Ugarte Marin, Taylor Jackson, and Aurora Amesbury.
Funding
This research was funded by the USDA National Institute of Food and Agriculture, grant number 2019-67015-29571.
Author information
Authors and Affiliations
Contributions
J.B.K contributed to study design, data collection, data analyses, and manuscript preparation. J.F. contributed to data analyses. E.M.C. contributed to funding acquisition, study design, and manuscript preparation.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Bonney-King, J., Fischer, J. & Miller-Cushon, E. Effects of reward type and previous social experience on cognitive testing outcomes of weaned dairy calves. Sci Rep 15, 7571 (2025). https://doi.org/10.1038/s41598-025-91843-x
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41598-025-91843-x
