
Abstract
Currently, the use of anti-stress elicitors such as melatonin, N-acetyl-5-methoxytryptamine, is known as an effective method to reduce the negative impacts of stressful environments. Therefore, a two factor- five level (irrigation levels: 25–75% field capacity (FC) and melatonin: 0-300 mM) dry yield optimization and modeling were done using central composite design, essential oil quantity and quality, total phenol, flavonoid and chlorophyll content and also some physiological properties of grapefruit mint (Mentha suaveolens × Mentha piperita). The results showed that the grapefruit mint biomass in severe drought stress was reduced by 73% in comparison with normal irrigation. However, the application of 155 mM melatonin enhanced biomass yield, phenol, flavonoid, essential oil content and essential oil yield by 92.2, 33.4, 55.1, 45.3 and 143%, in comparison with control. Optimization analysis predicted that under FC 65% irrigation level and application of 240 mM melatonin would positively assist in achieving the maximum chlorophyll, phenol, flavonoid content and essential oil yield of grapefruit mint. In addition, the maximum content of linalool and linalool acetate was observed in FC 50% treated with 155 mM melatonin. Generally, it can be concluded that the application of melatonin, especially in 155 mM level, improved plant growth, essential oil quantity and quality and also plant physiological characteristics under drought stress.
Similar content being viewed by others

Introduction
Nowadays, growing consumer interest to using herbal-based and organic products such as medicinal and aromatic plants has led to great demand in the botanicals industry. The extensive use of MAPs in various industries, including pharmaceuticals, cosmetics, and food, highlights the importance of re-evaluating research, development, production, and application1. The Mentha genus, which comprises over 60 species, is well-known for its medicinal and aromatic value2,3. Grapefruit mint, a sterile hybrid resulting from the cross between Mentha piperita and Mentha suaveolens, is one of the perennial herbs belonging to Lamiaceae family4. The plant is used in various industries including pharmaceutical, cosmetic, food, and nutraceutical industries5. It has been reported that linalool, linalool acetate, and linalyl anthranilate are the main essential oil constituents of grapefruit mint which showed promise for clinical applications in pain relief such anti-tumor, antibacterial, etc. uses5,6.
Due to the low amount of average rainfall in Iran (250 mm), which is about one third of the average rainfall in the world, this country is located in a dry climatic zone7,8. In agriculture section, drought stress affects negatively different aspect of the plant growth characteristics and productivity8,9. It has been reported that drought stress decrease photosynthesis rate as a result of reduction of CO2absorption, increase of the production of reactive oxygen species (ROS) species which leads to decreasing the quantity and quality of plant’s productivity7,10. In recent years, developing innovative strategies to enhance plant efficiency under challenging environmental conditions has become a critical focus in agriculture11. Melatonin, scientifically known as N-acetyl-5-methoxytryptamine, serves as a versatile signaling molecule and vital bioactive compound in vascular plants. It mitigates the detrimental impacts of various abiotic stresses and promotes growth and physiological functions across numerous plant species11. Previous studies reported that melatonin application had different beneficial roles in plants such as improving germination, root elongation, photosynthesis, sucrose metabolism, abiotic stress resistance and redox reactions12. One key role of melatonin is its ability to reduce chlorophyll breakdown by downregulating genes associated with chlorophyll degradation. Additionally, melatonin’s endogenous application lowers reactive oxygen species (ROS) levels, thereby enhancing plant tolerance to drought stress12,13. Mazrou et al.14 reported that drought stress reduced the herbal yield of Pelargonium graveolens L. (Herit), while this reduction was only 14.38% when melatonin was applied. Also, these authors concluded that melatonin application reduced hydrogen peroxide and malondialdehyde accumulation by 71.11 and 48.30% under drought conditions.
The diversity of Iran’s climate and its different ecological conditions are among the factors that have made cultivating medicinal and aromatic plants in this country with relatively high-income generation. In contrast, different stressful conditions such as drought stress affect negatively the productivity of above-mentioned plants. Currently, the use of anti-stress elicitors such as melatonin is known as an effective method to reduce the negative impacts of stressful conditions. Noticeably, the available information concerning melatonin functions in enhancing drought stress resistance is mainly from field crops or other horticultural plants. However, there is a paucity of information about the effectiveness of melatonin application on the physiological and biochemical properties of medicinal and aromatic plants under various biotic and abiotic stress. Therefore, the study was aimed to investigate the effect of melatonin application on drought stressed grapefruit mint (based on the productivity, phytochemical and physiological characteristics) using response surface methodology.
Results
Based on the analysis of variance results both developed models were significant for all studied traits (dry weight, chlorophyll content, malondialdehyde concentration, proline content, H2O2 concentration, phenol and flavonoid content, essential oil content and yield) (Tables 1 and 2). At the 1% level, the effects of melatonin application and drought stress alone were significant for dry weight, chlorophyll content, and malondialdehyde concentration, proline content, H2O2 concentration, flavonoid content, essential oil content and essential oil yield of grapefruit mint. Also, the phenol content of grapefruit mint was significantly affected by interaction of irrigation levels and melatonin application.
Dry weight
Table 3 shows the means of studied traits form different treatments following CCD configuration. The highest dry weight of grapefruit mint (94.25 g) was observed in FC 75% treated with 155 mM melatonin, whereas the lowest dry weight of the plant (25.45 g) was achieved in severe drought stress (FC 25%) treated with 155 mM melatonin (Table 3; Fig. 1a, b). The dry yield of grapefruit mint in severe drought stress conditions (FC 25%) was reduced by 73% in comparison with normal irrigation conditions (FC 75%). Additionally, the application of 70, 155, 240 and 300 mM melatonin enhanced the dry yield of grapefruit mint by 29.3, 69.2, 92.2 and 12.2% in comparison with control (non-application of melatonin). Optimization analysis predicted that under FC 75% irrigation level and application of 155 mM melatonin would positively assist in achieving the highest dry yield of grapefruit mint (95 g) (Table 4).

Response surface plots [3D (a) and contour plots (b)] of grapefruit mint dry weight under different irrigation levels and melatonin application.
Chlorophyll content
Chlorophyll a (2.89 mg g−1 FW), b (0.92 mg g−1 FW), and total (3.81 mg g−1 FW) were at their highest levels in FC 75% treated with 155 mM melatonin. The lowest levels of chlorophyll a (1.39 mg g−1 FW), b (0.42 mg g−1 FW), and total (1.82 mg g−1 FW) were noted in severe drought stress (FC 25%) when 155 mM melatonin was administered (Table 3; Fig. 2a, b, c). The amount of chlorophyll a, b, and total was 52, 54, and 52% reduced under severe drought stress conditions (FC 25%) than under normal irrigation conditions (FC 75%). Applying 240 mM of melatonin had the biggest impact on chlorophyll a, b, and total among the various doses; compared to control, the aforementioned traits increased by 84, 98, and 86%. According to optimization analysis, the highest levels of chlorophyll a (2.92 mg g−1 FW), b (0.94 mg g−1 FW), and total (3.86 mg g−1 FW) would be achieved under FC 65% irrigation level and 240 mM melatonin application (Table 4).

Response surface plots [3D (a) and contour plots (b)] of grapefruit mint chlorophylls content under different irrigation levels and melatonin application.
Malondialdehyde (MDA)
The highest MDA content (6.42 nmol g−1 FW) was achieved in severe drought stress (FC 25%) treated with 155 mM melatonin. In contrast, the lowest MDA (3.34 nmol g−1 FW) was belonged to FC 75% treated with 155 mM melatonin. In comparison with normal irrigation conditions (FC 75%), the MDA content of grapefruit mint was enhanced by 92.2% in severe drought stress conditions (FC 25%). Additionally, the application of 70, 155, 240 and 300 mM melatonin decreased the MDA content of grapefruit mint by 11.1, 17.8, 23.2 and 17% in comparison with control (Table 3; Fig. 3a1, a2). Optimization analysis predicted that under FC 75% irrigation level and application of 240 mM melatonin would positively assist in achieving the minimum MDA content of grapefruit mint (Table 4).

Response surface plots [3D (a1) and contour plots (a2)] of grapefruit mint malondialdehyde (MDA) under different irrigation levels and melatonin application.
Proline content
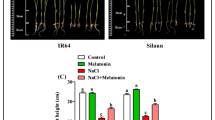
The maximum proline content (3.43 µmole g−1 FW) was achieved in severe drought stress (FC 25%) treated with 155 mM melatonin. In contrast, the lowest proline content (1.75 µmole g−1 FW) was belonged to FC 75% treated with 155 mM melatonin. In comparison with normal irrigation conditions (FC 75%), the proline concentration was enhanced by 96% in severe drought stress conditions (FC 25%). Additionally, the application of 70, 155, 240 and 300 mM melatonin decreased the proline content of grapefruit mint by 4.2, 14.1, 16 and 12.8% in comparison with control (Table 3; Fig. 4 b1, b2). Optimization analysis predicted that under FC 75% irrigation level and application of 240 mM melatonin would positively assist in achieving the minimum proline content of grapefruit mint (1.62 µmole g−1 FW) (Table 4).

Response surface plots [3D (b1) and contour plots (b2)] of grapefruit mint proline under different irrigation levels and melatonin application.
H2O2 concentration
The highest H2O2 concentration (4.77 µmole g−1 FW) was achieved in severe drought stress (FC 25%) treated with 155 mM melatonin. In contrast, the lowest H2O2 concentration (1.53 µmole g−1 FW) was belonged to FC 75% treated with 155 mM melatonin. In comparison with normal irrigation conditions (FC 75%), the H2O2 concentration was enhanced by 211.8% in severe drought stress conditions (FC 25%). Additionally, the application of 70, 155 and 240 mM melatonin decreased the H2O2 concentration of grapefruit mint by 8.9, 8.5 and 26.9% in comparison with control (Table 3; Fig. 5 c1, c2). Optimization analysis predicted that under FC 75% irrigation level and application of 155 mM melatonin would positively assist in achieving the minimum H2O2 content of grapefruit mint (1.51 µmole g−1 FW) (Table 4).

Response surface plots [contour plots (c1) and 3D (c2)] of grapefruit mint H2O2 concentration under different irrigation levels and melatonin application.
Phenol content
The highest phenol content (31.93 mg GAE g−1 DW) was achieved in moderate drought stress (FC 50%) treated with 155 mM melatonin. In contrast, the lowest phenol content (18.01 mg GAE g−1 DW) was belonged to FC 75% treated with 155 mM melatonin. Additionally, the application of 70, 155, 240 and 300 mM melatonin enhanced the phenol content of grapefruit mint by 7.1, 33.4, 31.5 and 5.1% in comparison with control (Table 3; Fig. 6 a1, a2). Optimization analysis predicted that under FC 65% irrigation level and application of 240 mM melatonin would positively assist in achieving the maximum phenol content of grapefruit mint (32.41 mg GAE g−1 DW) (Table 4).

Response surface plots [3D (a1) and contour plots (a2)] of grapefruit mint phenol content under different irrigation levels and melatonin application.
Flavonoid content
The maximum flavonoid content (16.33 mg QE g−1 DW) was achieved in moderate drought stress (FC 50%) treated with 155 mM melatonin. In contrast, the minimum flavonoid content (6.48 mg QE g−1 DW) was belonged to FC 75% treated with 155 mM melatonin. Additionally, the application of 70, 155, 240 and 300 mM melatonin enhanced the flavonoid content of grapefruit mint by 28.8, 55.1, 45.6 and 31.4% in comparison with control (Table 3; Fig. 7 b1, b2). Optimization analysis predicted that under FC 65% irrigation level and application of 155 mM melatonin would positively assist in achieving the maximum flavonoid content of grapefruit mint (16.64 mg QE g−1 DW) (Table 4).

Response surface plots [3D (b1) and contour plots (b2)] of grapefruit mint flavonoid content under different irrigation levels and melatonin application.
Essential oil content
The maximum essential oil content (2.38%) was achieved in moderate drought stress (FC 50%) treated with 155 mM melatonin. In contrast, the minimum essential oil content (1.01%) was belonged to FC 75% treated with 155 mM melatonin. In addition, application of 70, 155, 240 and 300 mM melatonin enhanced the essential oil content of grapefruit mint by 11.5, 45.3, 54 and 23.1% in comparison with control (Table 3; Fig. 8 a1, a2). Optimization analysis predicted that under FC 50% irrigation level and application of 240 mM melatonin would positively assist in achieving the maximum essential oil content (2.39%) (Table 4).

Response surface plots [3D (a) and contour plots (b)] of grapefruit mint essential oil content and essential oil yield under different irrigation levels and melatonin application.
Essential oil yield
The maximum essential oil yield (1.75 g plant−1) was achieved in FC 65% treated with 155 mM melatonin. In contrast, the minimum essential oil yield (0.3 g plant−1) was belonged to FC 25% treated with 155 mM melatonin. Application of 70, 155, 240 and 300 mM melatonin increased the essential oil yield of grapefruit mint by 44, 143, 196 and 38% in comparison with control (Table 3; Fig. 8 b1, b2). Optimization analysis predicted that under FC 65% irrigation level and application of 240 mM melatonin would positively assist in achieving the maximum essential oil yield (1.83 g plant−1) (Table 4).
Essential oil constituents
The main grapefruit mint essential oil constituents were linalool (44.6–51.46%), linalool acetate (29.46–33.16%) and α-terpineol (3.31–4.86%), respectively. The highest content of linalool and linalool acetate was observed in FC 50% irrigation level treated with 155 mM melatonin application that was 15.4 and 11.3% greater than compared with normal irrigation conditions (FC 75%). Averagely, application of 70, 155, 240 and 300 mM melatonin increased the content of linalool by 1.3, 2.4, 7.3 and 4.1% and the content of linalool acetate by 1.4, 5.1, 9.5 and 4.3% in comparison with control. Also, the maximum content of α-terpineol was achieved in FC 65% irrigation level treated with 240 mM melatonin application (Table 5).
Discussion
The obtained results from this study showed that the dry yield of grapefruit mint declined in drought stress conditions. In this condition, the nutrient absorption efficiency decreased around the root zone as a result of lower mobility and rate of mineral diffusion which eventually lead to reduction of plant performance and productivity15. Also, the decreasing of plants productivity under drought stress conditions is related to the negative effects of water shortage on the cell turgor pressure that affect cell growth characteristics (elongation and volume), and eventually reducing stomata conductivity, CO2uptake and photosynthesis rate16,17. Alike, Amani Machiani et al.18 noted that the dry yield of thyme (Thymus vulgarisL.) was reduced by 13% and 40.3% in mild and severe drought stress conditions. On the other hand, application of melatonin enhanced the grapefruit mint dry yield under non-stress and drought stress conditions. Melatonin also interacts with the auxin receptor, acting as an auxin agonist, which promotes cell division and expansion, ultimately boosting plant productivity15. Additionally, the melatonin application enhanced the photosynthesis pigments under drought stress conditions which improve translocation from source to sink and finally improve plant performance under stressful conditions19. Also, Ahmad et al.20noted that melatonin increases the expression of genes involved in plant growth and development, abiotic stress resistance by removing reactive oxygen species (ROS) compounds which lead to improving plant performance under stressful conditions21. It has been reported that the melatonin application significantly enhanced the leaf area, total carbohydrates, fresh and dry weights of peppermint (Mentha piperita L.) seedlings.
The chlorophyll content plays an important role in plant performance through increasing the photosynthesis capacity and rate. The results demonstrated that drought stress decrease the content of chlorophyll a, b and total. Under stressful conditions, increasing the content of ROS compounds such as H2O2which lead to enhancement of lipid peroxidation on the membranes8. These conditions enhance the rate of chloroplast breakdown which is directly impacted the amount of plant photosynthesis22. Research indicates that reduced chlorophyll content under water-limiting conditions may result from a decreased accumulation of chlorophyll biosynthetic intermediates, such as glutamate-1-semialdehyde (GSA) and 5-aminolevulinic acid23. However, under drought stress conditions, application of melatonin enhanced chlorophyll content through decreasing the ROS compounds activity (such as H2O2) and decreasing chloroplast breakdown24. Additionally, Zoufan et al.25 noted that melatonin application could enhance the chlorophyll content through increasing the synthesis of chlorophyll precursor compounds such as ALA and porphyrin. Similarly, Khosravi et al.26 reported that the melatonin application enhanced the chlorophyll content of Hot peppers (Capsicum spp.) by 17 and 18% in non-stress and drought stress conditions, respectively. Also, Imran et al.27 concluded that application of melatonin under drought stress conditions enhanced the chlorophyll content of soybean (Glycine max L.) plant.
Findings also show that drought stress significantly elevates proline levels in thyme seedlings. In such conditions, plants adjust their water relations to preserve cellular functions by synthesizing and accumulating osmotolerant metabolites, such as proline and other compatible solutes. Increased proline content under stress helps plants maintain osmotic balance and cell turgor, stabilize membranes, prevent electrolyte leakage, and keep ROS levels within normal ranges, which collectively reduce the adverse effects of drought stress28,29. Similarly, The MDA content and H2O2 activity was enhanced under drought stress conditions. The increased productivity of ROS compounds like H2O2may account for the increase in MDA content under drought stress conditions by increasing lipid peroxidation and the production of different aldehydes like MDA8,30. In contrast, application of melatonin decreased the MDA content and H2O2activity in stressful conditions. Melatonin as antioxidant compounds activate the expression of specific resistance genes under drought and other stress conditions which lead to increasing the antioxidant activity and keep the ROS compounds in normal ranges31. Therefore, the decreasing MDA concentration by melatonin application could be attributed to the role of melatonin in decreasing H2O2 activity and reduction of lipid peroxidation membranes.
The essential oils are natural substances occurring volatile aroma compounds synthesized by medicinal and aromatic plants and used in various industries such as medicine, cosmetics, food and etc. In this study, the essential oil content and main essential oil constituents of grapefruit mint including linalool and linalool acetate as well as phenol and flavonoid content enhanced under drought stress conditions. One of the important and influential factors in the process of photosynthesis rate is the ratio of NADP+/NADPH + H+15. In photosynthesis process, the decreasing regeneration of NADP+ compounds (caused in drought and other stressful conditions) lead to decrease the ratio of NADP+/NADPH + H+ and ultimately photosynthesis rate. Therefore, the productivity of essential oils compounds through using of NADPH + H+ stored in plant cells, balance the ratio of NADP+/NADPH + H+and improve the efficiency of the photosynthesis cycle18. It can be inferred that secondary metabolites, including essential oils, phenols, and flavonoids, enhance the resilience of medicinal and aromatic plants under stress. For instance, the essential oil content of Balangu (Lallemantia iberica) increased by 283% under drought conditions32. Similarly, Amani Machiani et al.18 found that the essential oil content in thyme (Thymus vulgaris L.) increased by 34% under moderate and 25% under severe drought stress. They noted that key essential oil components, such as thymol and γ-terpinene, showed enhanced levels under moderate water stress.
In addition, application of melatonin enhanced phenol, flavonoid and essential oil quantity and quality of grapefruit mint plants. In this study, application of melatonin enhanced the chlorophyll content which led to an improved photosynthesis rate. Increasing the availability of photosynthetic products have positive roles on the plants essential oil productivity through enhancement of secondary metabolites precursor compounds (such as pyruvate glyceraldehyde phosphate, phosphoenolpyruvate and etc.)33,34. Similarly, Dabaghkar et al.35 noted that melatonin application enhanced the carvacrol content of Satureja rechingeri Jamzad plant by approximately 18.09%.
Conclusion
The obtained results exhibited that drought stress affects negatively different aspect of the plant growth characteristics and productivity of grapefruit mint. However, the secondary metabolites such as essential oils, phenol and flavonoid content enhanced under drought stress conditions. Additionally, melatonin application, as anti-stress elicitor, reduced the negative impact of drought stress conditions by decreasing the ROS compound (such as H2O2) concentration. Moreover, application of melatonin enhanced phenol and flavonoid content. Optimization analysis predicted that under FC 50% irrigation level and application of 240 mM melatonin would positively assist in achieving the maximum essential oil content (2.39%). The mentioned analysis showed that application of 155 mM melatonin under FC 75% irrigation level would positively assist in achieving the highest dry yield of grapefruit mint. Interestingly, melatonin application, improve essential oil quality of grapefruit mint through increasing the main essential oil constitutes such as linalool, linalool acetate and grapefruit mint. Overall, the exogenous application of melatonin could be suggested as an effective strategy for improving plant performance, essential oil quantity and quality as well as other secondary metabolites such as phenol and flavonoid content under drought stress conditions.
Materials and methods
This investigation was performed under greenhouse condition at the University of Maragheh, Maragheh, Iran. Two factors consist on irrigation levels (25, 35, 50, 65 and 75% of field capacity) and melatonin concentration (0, 70, 155, 240 and 300 mM) were arranged in a five-level factorial scheme of central composite design which was a 2n fractional factorial scheme, 2×n axial by m central points. The axial form is identical to this scheme except for one factor, which will take on amounts either above the high or low magnitudes of the 2n factorial scheme. The required runs coupled with 5 central positions in the runs. The collected data were subjected to response surface methodology to identify the best-fitted models for the factors under examination. The suitability of each model was assessed using ANOVA, lack of fitness, and coefficient of determination (R2, using the Expert-Design 13.0 (Stat-Ease, MN, SC, USA).
The grapefruit-mint rhizomes were planted in 24-cell transplant trays filled with a 50:50 (v/v) cocopeat: perlite medium. When the seedlings reached the ten-pair leaf stage, they were transferred to 10 L pots filled with a 2:1:1 mixture of sieved field soil, sand and leaf mold. Temperatures between 18 and 30 °C, a 16-hour photoperiod and relative humidity between 60 and 80% were used to maintain seedlings. One month after planting, grapefruit mint seedlings were subjected to five different irrigation levels including 75, 65, 50, 35 and 25% of soil field capacity (FC). For precise application of these water deficit stress levels, pots were weighed daily, and water replacement was calculated based on the technique by Morshedloo et al.36. Specifically, soil moisture content was measured 24 h after watering by collecting a 100 g soil sample from each pot, oven-drying it, and reweighing. Based on this, any required water was added to maintain the intended soil moisture level within each specific treatment group. Melatonin was purchased from Sigma–Aldrich company (USA). To investigate the impact of melatonin on drought resistance, exogenous melatonin was applied to the plants six days before applying drought stress levels. Melatonin, sourced from Sigma–Aldrich (USA), was sprayed at concentrations of 0 (control), 70, 155, 240, and 300 mM. Control plants received distilled water. To prevent nutrient deficiencies, each pot was irrigated four times throughout the growth period with Hoagland nutrient solution. Subsequent foliar applications of melatonin were administered every 10 days to reinforce plant defenses. Finally, at flowering stage, fully expanded leaves of grapefruit mint were harvested and stored at − 80 °C to preserve them for subsequent biochemical analyses4.
Measurements
At the full flowering stage, all grapefruit mint seedlings were harvested. To measure the aerial biomass, harvested samples were left at room temperature for two weeks without direct sunlight to allow for thorough drying.
For chlorophyll and carotenoid analysis, 0.5 g of fresh grapefruit mint leaf tissue was homogenized in 10 mL of 80% acetone, then centrifuged at 12,000 rpm for 15 min. Absorbance of the resulting solution was recorded at wavelengths of 646.8, 663.2, and 470 nm using a Shimadzu UV-1800 spectrophotometer (Japan). Chlorophyll a (Chl a), chlorophyll b (Chl b), and carotenoid (Car) content were calculated in mg per gram fresh weight (FW) using equations from Lichtenthaler37:
For essential oil extraction, 50 g of dried ground grapefruit mint samples were combined with 300 mL of water and subjected to distillation for 3 h using a Clevenger-type apparatus. Essential oil content was determined by calculating the volume of oil obtained relative to the dry weight, using the method described by Haghaninia et al.1.
The essential oil yield was calculated by multiplying the dry weight yield by the essential oil percentage7. Furthermore, essential oil components were analyzed using gas chromatography-mass spectrometry (GC-MS; Agilent 5977 A) and gas chromatography with flame ionization detection (GC-FID; Agilent 7990B), as described by Ahmadi et al.4.
Malondialdehyde (MDA) concentration was measured by grinding 0.5 g of fresh leaves in 1.5 mL of 1% (w/v) trichloroacetic acid (TCA), followed by centrifugation at 12,000 rpm for 10 min at 4 °C. To 500 µL of the supernatant, 1000 µL of 0.1% (w/v) thiobarbituric acid (TBA) was added. The mixture was heated at 95 °C for 30 min, then rapidly cooled on ice, and absorbance was measured at 532 nm and 600 nm38.
To determine the proline content, fresh leaves were extracted using a ninhydrin reagent. The toluene phase was then separated for spectrophotometric examination in a 96-well quartz plate, with absorbance measured at 520 nm39.
To measure the total phenolic content, 0.2 g of dried, powdered grapefruit mint leaves were combined with 10 mL of 80% methanol. After 12 h of shaking at 200 rpm, the samples were separated from the supernatant. With absorbance at 765 nm, total phenol was determined using a 3% Na2HCO3 solution and a 10x diluted Folin-Ciocalteu reagent. Using the AlCl3reagent, the total flavonoid content was evaluated similarly to Ahmadi et al.4, with 415 nm being the absorbance.
Data availability
Correspondence and requests for materials should be addressed to AJ.
References
Haghaninia, M. et al. Adoption of arbuscular mycorrhizal fungi and Biochar for alleviating the agro-physiological response of Lavander (Lavandula angustifolia L.) subjected to drought stress. Plant. Stress. 12, 100461. https://doi.org/10.1016/j.stress.2024.100461 (2024).
Ćavar Zeljković, S. et al. Phenolic compounds and biological activity of selected Mentha species. Plants 10 (3), 550. https://doi.org/10.3390/plants1003055 (2021).
Machiani, M. A., Javanmard, A., Morshedloo, M. R. & Maggi, F. Evaluation of competition, essential oil quality and quantity of peppermint intercropped with soybean. Ind. Crop Prod. 111,743–754. https://doi.org/10.1016/j.indcrop.2017.11.052).
Ahmadi, H. et al. Introducing three new fruit-scented mints to farmlands: insights on drug yield, essential-oil quality, and antioxidant properties. Antioxidants 11 (5), 866. https://doi.org/10.3390/antiox11050866 (2022).
Akbarzadeh, S., Morshedloo, M. R., Behtash, F., Mumivand, H. & Maggi, F. Exogenous β-aminobutyric acid (BABA) improves the growth, essential oil content, and composition of grapefruit mint (Mentha Suaveolens× piperita) under water deficit stress conditions. Sci. Hortic. 9 (3), 354. https://doi.org/10.3390/horticulturae9030354 (2023).
de Cássia da Silveira e Sá, Andrade, R., de Sousa, D. P. & L.N. & A review on anti-inflammatory activity of monoterpenes. Molecules 18 (1), 1227–1254. https://doi.org/10.3390/molecules18011227 (2013).
Ostadi, A., Javanmard, A., Amani Machiani, M. & Kakaei, K. Optimizing antioxidant activity and phytochemical properties of peppermint (Mentha Piperita L.) by integrative application of biofertilizer and stress-modulating nanoparticles under drought stress conditions. Plants 12 (1), 151. https://doi.org/10.3390/plants12010151 (2022).
Haghaninia, M., Javanmard, A., Mahdavinia, G. R., Shah, A. A. & Farooq, M. Co-application of biofertilizer and Stress-Modulating nanoparticles modulates the physiological, biochemical, and yield responses of Camelina (Camelina sativa L.) under limited water supply. Soil. Sci. Plant. Nutr. 23 (4), 6681–6695. https://doi.org/10.1007/s42729-023-01521-y (2023).
Gao, S. et al. Effects of drought stress on growth, physiology and secondary metabolites of two Adonis species in Northeast China. Sci. Hortic. 259, 108795. https://doi.org/10.1016/j.scienta.2019.108795 (2020).
Zeng, X. et al. Decline in tree-ring growth of Picea Mongolica and its intra-annual eco-physiological responses to drought and CO2 enrichment in semi-arid China. J. Res. 35, 66. https://doi.org/10.1007/s11676-024-01716-8 (2024).
Asgari-Lajayer, B., Ghorbanpour, M. & Nikabadi, S. Heavy metals in contaminated environment: destiny of secondary metabolite biosynthesis, oxidative status and phytoextraction in medicinal plants. Ecotoxicol. Environ. Saf. 145, 377–390. https://doi.org/10.1016/j.ecoenv.2017.07.035 (2017).
Murch, S. J. & Erland, L. A. A systematic review of melatonin in plants: an example of evolution of literature. Front. Plant. Sci. 12, 683047. https://doi.org/10.3389/fpls.2021.683047 (2021).
Sun, C. et al. Melatonin. A master regulator of plant development and stress responses. J. Integr. Plant. Biol. 63 (1), 126–145. https://doi.org/10.1111/jipb.12993 (2021).
Mazrou, R. M., Hassan, F. A., Mansour, M. M. F. & Moussa, M. M. Melatonin Enhanced drought stress tolerance and productivity of Pelargonium graveolens L. (Herit) by regulating physiological and biochemical responses. Horticulturae 9(11), 1222. https://doi.org/10.3390/horticulturae9111222 (2023).
Hussain, H. A. et al. Chilling and drought stresses in crop plants: implications, cross talk, and potential management opportunities. Front. Plant. Sci. 9, 393. https://doi.org/10.3389/fpls.2018.00393 (2018).
Ahluwalia, O., Singh, P. C. & Bhatia, R. A review on drought stress in plants. Implications, mitigation and the role of plant growth promoting rhizobacteria. Resour. Environ. Sustain. 5, 100032. https://doi.org/10.1016/j.resenv.2021.100032 (2021).
Zia, R., Nawaz, M. S., Siddique, M. J., Hakim, S. & Imran, A. Plant survival under drought stress: implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Microbiol. Res. 12, 126626. https://doi.org/10.1016/j.micres.2020.126626 (2021).
Amani Machiani, M., Javanmard, A., Ostadi, A. & Alizadeh, K. Improvement in essential oil quantity and quality of thyme (Thymus vulgaris L.) by integrative application of Chitosan nanoparticles and arbuscular mycorrhizal fungi under water stress conditions. Plants 12 (7), 1422. https://doi.org/10.3390/plants12071422 (2023).
Sadak, M. S., Abdalla, A. M., Abd Elhamid, E. M. & Ezzo, M. Role of melatonin in improving growth, yield quantity and quality of Moringa oleifera L. plant under drought stress. Bull. Natl. Res. Cent. 44, 1–13. https://doi.org/10.1186/s42269-020-0275-7 (2020).
Ahmad, I. et al. The role of melatonin in plant growth and metabolism, and its interplay with nitric oxide and auxin in plants under different types of abiotic stress. Front. Plant. Sci. 14, 1108507. https://doi.org/10.3389/fpls.2023.1108507 (2023).
Sharma, P., Thakur, N., Mann, N. A. & Umar, A. Melatonin as plant growth regulator in sustainable agriculture. Sci. Hortic. 323, 112421. https://doi.org/10.1016/j.scienta.2023.112421 (2024).
Mohammadi, H., Amirikia, F., Ghorbanpour, M., Fatehi, F. & Hashempour, H. Salicylic acid induced changes in physiological traits and essential oil constituents in different ecotypes of Thymus Kotschyanus and Thymus vulgaris under well-watered and water stress conditions. Ind. Crop Prod. 129, 561–574. https://doi.org/10.1016/j.indcrop.2018.12.046 (2019).
Dalal, V. K. & Tripathy, B. C. Modulation of chlorophyll biosynthesis by water stress in rice seedlings during Chloroplast biogenesis. Plant. Cell. Environ. 35 (9), 1685–1703. https://doi.org/10.1111/j.1365-3040.2012.02520.x (2012).
Muhammad, I. et al. A meta-analysis of photosynthetic efficiency and stress mitigation by melatonin in enhancing wheat tolerance. BMC Plant. Biol. 24 (1), 427. https://doi.org/10.1186/s12870-024-05132-2 (2024).
Zoufan, P., Zare Bavani, M. R., Tousi, S. & Rahnama, A. Effect of exogenous melatonin on improvement of chlorophyll content and photochemical efficiency of PSII in Mallow plants (Malva parviflora L.) treated with cadmium. Physiol. Mol. Biol. 29(1), 145–157. https://doi.org/10.1007/s12298-022-01271-8 (2023).
Khosravi, S., Haghighi, M. & Mottaghipisheh, J. Effects of melatonin foliar application on hot pepper growth and stress tolerance. Plant. Stress 9, 100192. https://doi.org/10.1016/j.stress.2023.100192 (2023).
Imran, M. et al. Exogenous melatonin induces drought stress tolerance by promoting plant growth and antioxidant defence system of soybean plants. AoB Plants. 13 (4), plab026. https://doi.org/10.1093/aobpla/plab026 (2021).
Hayat, S. et al. Role of proline under changing environments: a review. P Plant. Signal. Behav. 7 (11), 1456–1466. https://doi.org/10.4161/psb.21949 (2012).
Hosseinifard, M. et al. Contribution of exogenous proline to abiotic stresses tolerance in plants: a review. Int. J. Mol. Sci. 23 (9), 5186. https://doi.org/10.3390/ijms23095186 (2022).
Hasanuzzaman, M. et al. Reactive oxygen species and antioxidant defense in plants under abiotic stress: revisiting the crucial role of a universal defense regulator. Antioxidants 9 (8), 681. https://doi.org/10.3390/antiox9080681 (2020).
Khan, Z. et al. Exogenous melatonin induces salt and drought stress tolerance in rice by promoting plant growth and defense system. Sci. Rep. 14 (1), 1214. https://doi.org/10.1038/s41598-024-51369-0 (2024).
Javanmard, A. et al. Optimizing phytochemical and physiological characteristics of Balangu (Lallemantia iberica) by foliar application of Chitosan nanoparticles and Myco-Root inoculation under water supply restrictions. Horticulturae 8 (8), 695. https://doi.org/10.3390/horticulturae8080695 (2022).
Sandoval, Y., Tighe-Neira, R., Inostroza-Blancheteau, C., Soto-Cerda, B. & González-Villagra, J. Melatonin improves plant water status, photosynthetic performance, and antioxidant defense system in highbush blueberry (Vaccinium corymbosum L.) plants subjected to drought stress. Sci. Hortic. 323, 112528. https://doi.org/10.1016/j.scienta.2023.112528 (2024).
Calsamiglia, S., Busquet, M., Cardozo, P., Castillejos, L. & Ferret, A. Invited review: essential oils as modifiers of rumen microbial fermentation. JDS 90 (6), 2580–2595. https://doi.org/10.3168/jds.2006-644 (2007).
Dabaghkar, Y. et al. Exploring the impact of exogenous melatonin on agro-morphological characteristics, carvacrol, and Rosmarinic acid production in Satureja Rechingeri Jamzad under drought stress. Chem. Biol. Technol. Agric. 11, 112. https://doi.org/10.1186/s40538-024-00643-4 (2024).
Morshedloo, M. R. et al. Effect of prolonged water stress on essential oil content, compositions and gene expression patterns of mono-and sesquiterpene synthesis in two oregano (Origanum vulgare L.) subspecies. PPB 111, 119–128. https://doi.org/10.1016/j.plaphy.2016.11.023 (2017).
Lichtenthaler, H. K. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. In: Methods in Enzymology. vol. 148: Elsevier. 350–382. https://doi.org/10.1016/0076-6879(87)48036-1 (1987).
Heath, R. L. & Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 125 (1), 189–198. https://doi.org/10.1016/0003-9861(68)90654-1 (1968).
Bates, L. S., Waldren, R. & Teare, I. Rapid determination of free proline for water-stress studies. Plant. Soil. 39, 205–207. https://doi.org/10.1007/BF00018060 (1973).
Acknowledgements
The present study was carried out by the use of facilities and materials at the University of Maragheh and the paper is published as the part of a Ph.D. Student research at University of Maragheh, research affairs office.
Funding
The current research has received no funding from agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
S. R., writing—review and editing, writing-original draft, software, visualization, methodology, investigation, conceptualization. A. J., writing—review and editing, writing—original draft, software, data curation. N.S., methodology, investigation, review and editing, conceptualization. M. M., writing—review and editing. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
The manuscript is an original work that has not been published in other journals.
Consent for publication
All authors agreed to the publication.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Rezaei, S., Javanmard, A., Sabaghnia, N. et al. Exogenous melatonin modulates physiological responses, phytochemical profiles and essential oil production in grapefruit mint under drought stress. Sci Rep 15, 14650 (2025). https://doi.org/10.1038/s41598-025-99281-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-025-99281-5
