
Abstract
On 13 April 2023, Cyclone Ilsa crossed Bedout Island in the Timor Sea off Western Australia’s remote north coast. Extensive damage was done to the entire island, with all coastal vegetation stripped clear from the island’s surface. Here we use repeated aerial and ground-based transect surveys of Bedout’s important seabird populations during 17 April – 21 July 2023 and Monte Carlo modelling to estimate total mortality for three species. We estimate that 80–90% of the populations of three seabird species (Masked Booby Sula dactylatra bedouti, Brown Booby S. leucogaster, and Lesser Frigatebird Fregata ariel) were killed in the storm, with only 40 breeding Masked Booby (a putative endemic subspecies) recorded on the island 15 weeks later. The frequency and intensity of such storms is likely approaching a threshold beyond which Bedout’s seabirds cannot readily recover, with cyclones hitting the island, on average, every seven years in recent decades. We discuss the implications for Bedout Island, and for wildlife conservation more broadly in the face of increasing, climate-driven storms and impacts documented worldwide.
Similar content being viewed by others

Introduction
Cyclones are the most destructive weather systems affecting Australia, especially for coastline bounded by the tropical waters of the Pacific and Indian Oceans1. The main hazards associated with tropical cyclones are intense winds, and heavy rain, and in coastal areas, storm surges can also have devastating impacts on wildlife, vegetation cover, and human infrastructure2,3. Low-lying islands (e.g., coral cays) commonly found throughout the tropics are refuge to unique and often threatened species4. Yet these habitats, including mangroves and fringing reefs, can be significantly damaged by cyclones, even when the storm is >800 km away2,5,6,7.
Outside the tropics, extreme weather events also cause substantial damage to wild species and spaces. For example, in the Atlantic, severe winter storms have resulted in starvation and mortality of thousands of seabirds across a vast area including the Labrador Sea, the waters off Iceland, and the Barents Sea8. In southern California, severe storms have repeatedly damaged or destroyed an important kelp (Macrocystis pyrifera) forest, with more than 10 such storms recorded in only a decade9. Storms also have significant implications for economies and livelihoods, with the global, annual costs of climate-driven storms estimated at US$143 billion10, and there is growing concern around the relationship between marine heat waves and storms, as warming waters facilitate more rapid intensification of cyclones11.
The extreme weather conditions created by cyclones and other types of storms can also reduce the fitness of wildlife in the path of the storm, either directly through mortality of individuals or reducing foraging and breeding opportunities of the survivors12. For example, while the immediate effects of the cyclone (e.g., big waves) are rarely the direct cause of fish mortality13, other consequences can impact population recovery or resilience. Deteriorating environmental conditions over subsequent weeks can suffocate fish or otherwise render their habitats unlivable (e.g., influx of freshwater from heavy rains creates anoxic conditions14,15). While some seabirds will actively flee their breeding areas during severe weather, and some will even fly directly into storms16,17, not all adult birds leave, and eggs/chicks are bound to the nest. In the tropics, many seabirds (including members of the family Sulidae) are adapted to low wind speeds, yet they often experience some of the strongest storms18. For many seabird species, their behaviour of building nests along the surface and shoreline means they receive little or no protection from severe storms. Not surprisingly, recent cyclones have contributed to significant mortality, breeding failure, reduced juvenile survival, and altered migration strategies in seabirds19,20,21.
On average, 11 tropical cyclones form or enter Australia’s waters every year22, however the frequency and intensity of tropical cyclones is increasing worldwide23,24. In Western Australia, tropical cyclones are a key factor degrading the ecological condition of reefs25, including those surrounding key seabird breeding sites, such as Christmas Island and the Abrolhos Islands26. The interval between cyclones classed as ‘big and strong’ (i.e., circulation >300 km and sustained wind speeds ≥33 m/s) is already less than 10 years for 14 ecoregions in Australia, including the north-west tropical zone which encompasses 1.43% of world coral reef area and multiple Important Bird and Biodiversity Areas such as Ashmore Reef and Bedout Island5. Together, these two sites account for ~2% of the world’s Kuyangarti/Lesser Frigatebird Fregata ariel and Purralyakura/Brown Booby Sula leucogaster populations27. As the climate continues to change rapidly, the occurrence of stretches of uninterrupted recovery time becomes shorter5 and the risk of significant disturbance to these islands and bird populations increases.
At present, there is little broad-scale information about the degree to which seabird species, populations, or ecosystems are vulnerable to climatic extremes, particularly in Australia, and whether plasticity in life-history characteristics, behaviour, or adaptation, is likely to moderate any of these effects28. Here we describe the impact of Cyclone Ilsa on the Jirli Waanyja/Masked Booby Sula dactylatra bedouti29, a putative subspecies that is only found on Bedout Island off Western Australia (Fig. 1). We also document the mortality of four additional species, the Lesser Frigatebird, Brown Booby, Marril/Crested Tern Thalasseus bergii and Brown Noddy Anous stolidus, and discuss the potential implications of the predicted climate-related increase in cyclone frequency and intensity on seabird populations, globally.

Aboriginal artwork depicting Bedout Island, a Purralyakura/Brown Booby and Cyclone Ilsa by a Wudjari artist Tanya Mead. © Tanya Mead.
Results
Counts of birds that survived the cyclone
No living seabirds of any species were observed in the video footage or images taken of the bare, sandy interior of the island during the helicopter fly-over of the island on 17 April (Fig. 2 and Supplementary S1). Roosting seabirds were noted along the northern shoreline (Supplementary Fig. S1), this group comprised 85 Brown Boobies and six Masked Boobies. Substantial effort was made to photograph and count the three focal species again during the July survey. In total, 40 Masked Boobies (one nest) and 2256 Brown Boobies (one nest) were recorded. Only three Lesser Frigatebirds were observed in thermals over a period of four hours (overlapping midday).

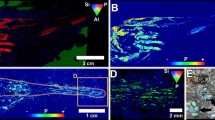
Photos of Bedout Island from the initial helicopter (aerial) survey four days after Cyclone Ilsa in April 2023. Panel b is a cropped (zoomed-in) section of panel a: in the bottom panel, at least 98 dead seabirds are visible, the area (approx. 25 × 13 m) is indicated by the dashed line. Both images are provided for reference only and were not used for data extraction as species identification was not possible.
The number of deceased seabirds detected within transects was consistently higher during the June survey of Bedout Island (average across all species 0.222 ± 0.155 birds/m2) compared with four weeks later in July (0.107 ± 0.092 birds/m2; Table 1). Masked and Brown Booby were the most frequently detected species (Table 1) and while carcasses were distributed across the entire island, the distribution was highly uneven. For example, 51 Brown Booby were detected in a single transect (53% of all individuals of this species recorded in June). Masked Booby carcasses were detected in two locations on the island with 15 birds recorded in a single transect along the south-west shoreline in June (0.125 birds/m2) and six near the clearing in July (0.050 birds/m2).
Overall densities for Brown Noddy and Lesser Frigatebird carcasses were low as these species were only detected in a single transect. However, within each of these transects, the number of carcasses was high (eight of both species; Table 1).
Estimated total mortality
In June, we estimated 26,235 (95% CI: 10,265–48,584) Brown Booby and 6941 (95% CI: 2221–14,165) Masked Booby carcasses, while in July the estimates were 10,272 (95% CI: 3602–21,384) for Brown Boobies and 3887 (95% CI: 1110–7218) for Masked Boobies. These estimates often exceed the known breeding population because of the limited sampling opportunity available and overdispersed data, but 95% CIs include maximum population estimates (Brown Boobies: 22,400 individuals, Masked Boobies: 800 individuals), including some juvenile mortality. We are therefore confident that nearly all Masked and significant numbers of Brown Boobies from Bedout Island died during Cyclone Ilsa.
Discussion
Seabird mortality on Bedout Island during Cyclone Ilsa was significant, impacting five species and around 80–90% of all individual birds nesting on the island. While cyclones are natural phenomena, this mortality event is notable for several reasons.
Firstly, cyclone frequency and intensity are increasing due to climate change23,24 and it is unclear whether seabird populations can sustain this level of mortality, particularly as many species are already in decline30 and/or facing multiple, cumulative pressures (e.g. ref. 31). On Bedout Island, at least three cyclones (George, Stan and Ilsa) have caused major damage in the past 15 years, each arriving after an interval of only seven years. The amount of time between environmental disturbance events like cyclones plays a significant role in overall ecosystem trajectory: if cyclone return intervals become shorter than those needed for community recovery, the classing OF individual storms as acute, pulse, or ‘natural’ disturbances requiring minimal intervention from wildlife managers is likely inaccurate, particularly given the many cumulative, synergistic and accelerating stressors associated with climate change32,33. For intertidal and shallow water systems, storm return intervals of 5–10 years already contribute to reduced fishery catch32, loss or inhibited recovery of some seagrass species33, and persistent degradation of coral reefs34 but comparable data and impacts are not yet available for birds.
Long-lived seabirds should prioritise their own survival over their current reproductive success35. Given this, adult birds would be expected to abandon their nests on Bedout given their own survival was compromised by Cyclone Ilsa. However, unlike many records of tropical cyclones elsewhere12,16, there was minimal evidence that adult seabirds fled Bedout Island prior to Cyclone Ilsa’s arrival, and instead, some species like Lesser Frigatebird appear to have died where they nested (carcasses of adult birds were only detected in a single transect). The reasons for this are not clear as frigatebirds and other seabird species can forecast a cyclone’s arrival based on meteorological conditions (e.g., wind, barometric pressure) and adjust their behaviour12,16. Cyclone Ilsa hit at a time when boobies were incubating both young chicks and eggs (Fig. 3c, d). The strong pair bond that develops between an adult and its chick may explain the reluctance of some birds to abandon the nest as the storm approached36, but this does not explain the behaviour of the frigatebirds, who would have been in the early stages of egg laying in April. In tropical systems, breeding seasons are less well demarcated than in temperate or polar regions because there is a less distinct shift in resource availability. In some species, subannual breeding or indeed continuous breeding can occur37,38,39. Regardless, our estimates of seabird mortality are conservative as detectability impacted our counts of deceased adult, breeding birds due to degradation of carcasses over time and re-growth of vegetation (mortality estimates in June were nearly double what was observed in July due to decreasing detection over time; Figs. 3 and 4, and Table 2). We were also unable to determine how many eggs or chicks were killed in the storm due to sediment erosion which buried or otherwise obscured from view the low-lying nest structures (Fig. 3d). Our limited time on the island precluded assessments of detection probability along transects. At a minimum, the mortality of adult birds from Cyclone Ilsa accounts for 80-90% of breeding booby populations on Bedout Island. The frequency of storms and potential recovery of these birds therefore has implications for the status of the island as an Important Bird and Biodiversity Area and conservation recognition of the Masked Booby globally listed as Least Concern but numerous populations known to be decreasing40,41,42.

Deceased adult boobies recorded during a transect completed in June (a). Many birds were difficult to detect as they were almost completely buried by sediment (b). Deceased booby chicks (c) were difficult to detect during surveys compared to larger, often darker-coloured adult birds. This was particularly the case in July due to degradation of the carcasses and re-growth of vegetation. Loss of eggs and nests during Cyclone Ilsa were widespread across Bedout Island, but detection was also difficult. For example, only a single booby egg was recorded (during the July survey; d): the nest structure and second egg were completely buried by sediment and not visible.

Estimates of Brown Booby (a) and Masked Booby (b) mortality on Bedout Island following Cyclone Ilsa in April 2023. The June (orange) and July (purple) surveys are presented separately due to issues with carcass detection and degradation (see Discussion and Fig. 5).
Nearly all the individual seabirds recorded dead during our surveys were adult, breeding birds. This also has important demographic implications as species that are long-lived and have low fecundity are reliant on high adult survivorship to sustain populations43. Red-tailed Tropicbird Phaethon rubricauda populations are robust to significant egg mortality resulting from cyclones, but only when no adult mortality is sustained19. Like Bedout’s Masked Boobies, the Critically Endangered Abbott’s Booby (Papasula abbotti) breeds on a single island off Australia’s northwest coast. Severe storms have been linked with the destruction of up to 30% of nests and fledglings in a single event, and a decline in the number of adults attempting to breed in subsequent years44. The consequences of storm damage for Abbott’s Booby are thought to be equivalent to activities associated with mining (e.g., habitat clearing)44. Overall, the capacity of long-lived species to recover from significant or repeated mortality events involving breeding adults is low and particularly worrisome for Bedout’s Masked Booby population with only 40 adult birds recorded alive in July 2023 (dry season). While some birds may have been at-sea during our surveys, or otherwise absent due to the damage to Bedout Island, the numbers appear substantially reduced. A recent genetic assessment identified six haplotypes unique to Masked Boobies on Bedout Island29, suggesting this population is unlikely to exchange genes with other colonies (the next nearest are the Lacepede Islands and Adele Island >450 km east; Fig. 5). Bedout’s Masked Booby population is therefore highly dependent on local recruitment for persistence, and at risk of a genetic bottleneck. A full taxonomic assessment using modern approaches is urgently needed. This combination of ecologically dependent reproductive isolation and depressed population size warrants careful monitoring of Bedout’s Masked Boobies and definitive resolution of its taxonomic status.

Transects completed in June (n = 5; orange) and July 2023 (n = 10; purple). Vegetation is visible on the island’s surface as the base map image is from May 2021. Adele Island and the Lacepede Islands have breeding Masked Boobies, but this species is not found on the Rowley Shoals or Scott Reef which are primarily home to terns and tropicbirds. The island shapefile was provided by ESRI World Imagery with all data available under Creative Commons Attribution 4.0 International license.
Conclusions
Many significant seabird breeding sites in the Pilbara region of Australia are regularly impacted by cyclones. For example, Cyclones Clare, Stan and Ilsa hit the Rowley Shoals (an important breeding site for Red-tailed Tropicbirds; Fig. 5, Supplementary Table S2) with peak winds of 91 km/h and 235 km/h recorded in 2016 and 2023, respectively. While precise estimates are only available for Bedout Island, bird mortality from Cyclone Ilsa is much more widespread. A significant storm surge has altered the shape of Bedout Island, and this undoubtedly impacted other marine species within the reef with sediment documented in the surrounding waters for at least four days post-cyclone (Supplementary Fig. S2). In 2004, Cyclone Fay struck nearby Scott Reef as a Category 5 storm, uprooting many large coral colonies down to a depth of 20 m and leading to a reduction in hard coral cover of up to 15% (no data are available regarding the cyclone’s impact on Scott Reef’s birds or other wildlife)45. It is therefore important to consider the impact of cyclones from a regional and multi-species context, particularly given predictions of increasing cyclone strength and frequency over coming decades.
As species with high adult survival, long generation times, and low fecundity, seabirds cannot often adapt in the face of rapidly changing environmental conditions. For example, seabirds (as a group) have not shifted breeding phenology in response to climate change which suggests that they either cannot, or that they are able to buffer these effects46, whereas phenological shifts in breeding are relatively common in terrestrial birds47.
Seabirds are integral to the health of islands and surrounding reef ecosystems. The birds connect marine and terrestrial systems by transporting vast quantities of marine-derived nutrients to their terrestrial breeding, roosting and nesting grounds via the deposition of guano and other inputs48. As ecosystem engineers, the removal of seabirds can lead to significant changes in island habitats and species complexes (e.g. ref. 49). Thus, the loss or major reduction in seabird colonies through significant storm events, such as Cyclone Ilsa documented for Bedout Island, could result in the alteration to ecosystem function or slow island recovery, particularly after severe or frequent storm events50. Given the paucity of basic count and life-history data for most seabird species in this increasingly cyclone-prone region of northern Australia, clear implications for demography and species persistence, and concurrent rapid expansion of offshore development (Supplementary Fig. S3), we encourage careful monitoring of seabird demography and recruitment so that we may better understand the capacity of these birds to adapt and recover from the rapid changes ahead.
Methods
Inclusion & ethics statement
All authors of this study have fulfilled the criteria for authorship required by Nature Portfolio journals and have been included as authors, as their participation was essential for the design and implementation of the study. This research includes findings that are locally relevant, being determined in collaboration with local Aboriginal community members and state government. Local and regional research and knowledge, including traditional species names, have been included in our citations and throughout the manuscript.
Study site and species
Bedout Island is a small (16.7 ha), low-lying coral cay located off the remote, tropical, north coast of Western Australia (−19.5878, 119.0995; Fig. 5). The island’s vegetation is primarily Spinifex longifolius and there are no trees or shrubs. The nearest town, Marapikurrinya/Port Hedland (91 km southwest), is Australia’s largest shipping port with 260 iron ore vessels (carrying 27 million tonnes of iron ore) passing through the port in May 2023 alone51. This area is also home to significant offshore oil and gas development (Supplementary Fig. S3), including the recent Dorado and Pavo discoveries with an estimated reserve of >480 million barrels of oil52; located within the Bedout basin.
Bedout Island is recognised as an Important Bird and Biodiversity Area as it is home to 1% of the world’s population of Lesser Frigatebirds53. Up to 2290 pairs of Lesser Frigatebird breed on the island during peak season (see below; authors’ pers. obs.), with the colony confined to an area around 3.34 ha along the north-central section of the island54. Found only on Bedout, the endemic S. d. bedouti is potentially one of Australia’s rarest seabirds with a total population of around 60–400 pairs, depending on the season (Supplementary Table S1). Most individuals breed in a small clearing near the lighthouse (approx. 1.68 ha; Supplementary Fig. S4) on the north-west side of the island, with occasional nests found along the eastern and northern coastline. Large numbers of Brown Booby also breed year-round on the island, with around 11,200 pairs in peak season see below53. Nests are commonly located within a few metres of the hightide mark along the entire coast covering 3.99 ha. Around 500 pairs of Crested Terns nest of Bedout Island in peak breeding season55. In recent years, this species has bred in a single colony (approx. 1.03 ha) on the eastern shore. Brown Noddy re-initiated breeding on the island following the removal of black rats Rattus rattus in 199127. Their population size and distribution on the island is not known.
Masked and Brown Boobies breed throughout the year on Bedout Island with eggs and chicks reported at various stages during Apr-June (often considered peak breeding) and again in Sep/Oct53. Lesser Frigatebirds nest in the Austral autumn with fluffy chicks observed in June and few fledglings remaining on the island by November (authors’ pers. obs.). Brown Noddy also breeds in autumn on Bedout Island53 however Crested Terns lay their eggs in November56 (authors’ pers. obs.). Thus, when Cyclone Isla hit the island in mid-April (towards the end of the cyclone season in Australia), four species were likely breeding.
Cyclone history of Bedout Island
Bedout Island has been impacted by five other cyclones in the past 15 years (older records, plus details for tropical low storms, are listed in Supplementary Table S2). First, Cyclone George hit the island as a Category 5 storm on 8 March 200757. Maximum windspeeds for this cyclone were recorded at Bedout Island (194 km/h; 53.9 m/s; lowest barometric pressure 941 hPa), and this remained a national windspeed record for Australia until Cyclone Ilsa hit Bedout in 2023 (detailed below).
In 2013, Bedout Island experienced a total of three cyclones. First, a tropical low (formerly Cyclone Peta) passed very close to the island on 22 January57. Weather data are incomplete, but winds reached at least 56 km/h and barometric pressure dropped to at least 991 hPa. Four weeks later Cyclone Rusty passed just north-west of the island as a Category 4 storm on 27 February (windspeeds 124 km/h, 34.4 m/s; pressure around 959 hPa57). Six months later, Cyclone Christine struck Bedout as a Category 3 storm on 30 December58. Notable for being a particularly large storm, the island sustained gale force winds (around 111 km/h, 30.8 m/s; lowest pressure 984 hPa) for 37 h.
Cyclone Stan hit Bedout Island as a Category 2 storm on 29–31 January 201659. The cyclone reached peak intensity of 102 km/h (28.3 m/s; lowest pressure 977 hPa) on 30 January when it was located ~90 km north of the island. During this time, southerly, gale force winds persisted with a peak wind gust of 91 km/h (25.3 m/s) recorded. To our knowledge, no surveys were conducted on Bedout Island to record the impact of any of these storms on the seabirds or other biodiversity.
Cyclone Ilsa impacted the same area of the Western Australian coastline as a Category 5 storm during 13–14 April 202359. This is the wet season along Australia’s north coast and a time of peak breeding for many seabird species. The eye of the cyclone passed directly over Bedout Island (Fig. 6) on the afternoon of 13 April 2023. The Bedout Island weather station ceased operation at that time, but immediately prior, it recorded sustained winds of 218 km/h (60.6 m/s) and gusts of 288 km/h (80 m/s), breaking the national record (see above; lowest pressure 915 hPa).

Cyclone Ilsa on 13 April 2023, with the eye of the storm immediately over Bedout Island (adapted from Australian Bureau of Meteorology, Port Hedland radar image).
Post-cyclone survey
Three surveys of Bedout Island were completed following Cyclone Ilsa. The first survey was completed four days after the storm passed on 17 April 2023. A helicopter flew over the island (permission to land was requested, but not approved); it spent approximately 20 min generating aerial imagery (photographs and video) using a GoPro 10 BLACK and iPhone 14 camera from an altitude of ~20 m. These images provided an initial snap-shot assessment of the overall condition of the birds and island but were not quantitative.
A second, ground-based survey was undertaken on 24 June 2023, ten weeks after Cyclone Ilsa. The island was accessed by boat, and a single person spent 2 h on shore. Deceased seabirds were counted and identified to species level within five transects (60 × 2 m) placed haphazardly across the island with an aim to cover as much of the central seabird breeding habitat as possible (Fig. 5). As this survey was limited in duration, a third and final assessment of seabird mortality was completed on 21 July 2023 to provide additional transect data, including from parts of the island not covered in June. During this final survey, the island was accessed by helicopter and a further 10 transects (60 × 2 m) were completed over four hours using a similar approach.
Birds still alive following Cyclone Ilsa were counted from aerial photographs taken on 17 April 2023 and again during ground-based surveys on 21 July 2023. Due to image constraints (low resolution), it was only possible to count the three larger species: Masked and Brown Booby and the Lesser Frigatebird.
To provide additional insight into overall, habitat-level changes to the island and surrounding reef from Cyclone Ilsa, satellite images were obtained from Sentinel-2 on 6 April 2023 (one week before the cyclone hit) and 16 April 2023 (three days after the cyclone passed; Supplementary Fig. S2).
Statistical analysis
To quantify seabird mortality, we used a Monte Carlo approach, treating June and July 2023 surveys separately because of issues around carcass detection and degradation. We used counts of dead birds and sampled five values (June) or ten values (July), with replacement, to generate a median density for the island (16.66 ha) and repeated this 10,000 times, drawing from a negative binomial distribution which was parameterised using a generalised linear intercept-only model for each survey period. From the resulting distributions, we calculated the mean, median, and 95% confidence interval (taken as the 2.5th and 97.5th percentile).
Data availability
Out of respect for Indigenous Data Sovereignty, the raw data are held by Esperance Tjaltjraak and are available upon request. The data contained in Tables 1 and 2 are publicly available here https://doi.org/10.6084/m9.figshare.24893376.
Code availability
The codes (in R language) used for our analyses are available here https://doi.org/10.6084/m9.figshare.24893376.
References
Wijnands, J. S. et al. Seasonal forecasting of tropical cyclone activity in the Australian and the South Pacific Ocean regions. 1, https://doi.org/10.1515/mcwf-2015-0002 (2015).
Taupo, T. & Noy, I. At the very edge of a storm: The impact of a distant cyclone on atoll islands. Econ. Disast. Clim. Change 1, 143–166 (2017).
Yuvaraj, E., Dharanirajan, K., Narshimulu, S. & Narshimulu, G. Post-disaster assessment of impact of Cyclone Lehar in South Andaman Island. Curr. Sci. 108, 85–90 (2015).
Fernández-Palacios, J. M. et al. Scientists’ warning – The outstanding biodiversity of islands is in peril. Glob. Ecol. Conserv. 31, e01847 (2021).
Puotinen, M. et al. Towards modelling the future risk of cyclone wave damage to the world’s coral reefs. Glob. Change Biol. 26, 4302–4315 (2020).
Riyas, C. A., Idreesbabu, K. K., Marimuthu, N. & Sureshkumar, S. Impact of the Tropical Cyclone Ockhi on ecological and geomorphological structures of the small low-lying Islands in the central Indian Ocean. Reg. Stud. Mar. Sci. 33, 100963 (2020).
Taillie, P. J. et al. Widespread mangrove damage resulting from the 2017 Atlantic mega hurricane season. Environ. Res. Lett. 15, 064010 (2020).
Clairbaux, M. et al. North Atlantic winter cyclones starve seabirds. Curr. Biol. 31, 3964–3971.e3963 (2021).
Seymour, R. J., Tegner, M. J., Dayton, P. K. & Parnell, P. E. Storm wave induced mortality of giant kelp, Macrocystis pyrifera, in southern California. Estuar. Coast. Shelf Sci. 28, 277–292 (1989).
Newman, R. & Noy, I. The global costs of extreme weather that are attributable to climate change. Nat. Commun. 14, 6103 (2023).
Rathore, S. et al. Interactions between a marine heatwave and Tropical Cyclone Amphan in the Bay of Bengal in 2020. Front. Clim. 4 https://doi.org/10.3389/fclim.2022.861477 (2022).
Wilkinson, B. P., Satgé, Y. G., Lamb, J. S. & Jodice, P. G. R. Tropical cyclones alter short-term activity patterns of a coastal seabird. Mov. Ecol. 7, 30 (2019).
Cheal, A. et al. Responses of coral and fish assemblages to a severe but short-lived tropical cyclone on the Great Barrier Reef, Australia. Coral Reefs 21, 131–142 (2002).
Bouchon, C., Bouchon-Navaro, Y. & Louis, M. Changes in the coastal fish communities following Hurricane Hugo in Guadelope Island (French West Indies). Atoll Res. Bull. 424, 1–19 (1994).
Wantiez, L., Chateau, O. & Le Mouellic, S. Initial and mid-term impacts of cyclone Erica on coral reef fish communities and habitat in the South Lagoon Marine Park of New Caledonia. J. Mar. Biolog. Assoc. U.K 86, 1229–1236 (2006).
Weimerskirch, H. & Prudor, A. Cyclone avoidance behaviour by foraging seabirds. Sci. Rep. 9, 5400 (2019).
Lempidakis, E. et al. Pelagic seabirds reduce risk by flying into the eye of the storm. Proc. Natl. Acad. Sci. 119, e2212925119 (2022).
Nourani, E. et al. Seabird morphology determines operational wind speeds, tolerable maxima, and responses to extremes. Curr. Biol. 33, 1179–1184.e1173 (2023).
Hennicke, J. C. & Flachsbarth, K. Effects of Cyclone Rosie on breeding Red-tailed Tropicbirds Phaethon rubricauda on Christmas Island, Indian Ocean. Marine Ornithol. 37, 175–178 (2009).
Thiebot, J.-B., Nakamura, N., Toguchi, Y., Tomita, N. & Ozaki, K. Migration of Black-naped Terns in contrasted cyclonic conditions. Marine Biol. 167, 83 (2020).
Nicoll, M. A. C. et al. Contrasting effects of tropical cyclones on the annual survival of a pelagic seabird in the Indian Ocean. Glob. Change Biol. 23, 550–565 (2017).
Dowdy, A. J. Long-term changes in Australian tropical cyclone numbers. Atmos. Sci. Lett. 15, 292–298 (2014).
Mei, W. & Xie, S.-P. Intensification of landfalling typhoons over the northwest Pacific since the late 1970s. Nat. Geosci. 9, 753–757 (2016).
Wu, L., Zhao, H., Wang, C., Cao, J. & Liang, J. Understanding of the effect of climate change on tropical cyclone intensity: A review. Adv. Atmos. Sci. 39, 205–221 (2022).
Zinke, J. et al. Gradients of disturbance and environmental conditions shape coral community structure for south-eastern Indian Ocean reefs. Divers. Distrib. 24, 605–620 (2018).
Gilmour, J. P. et al. The state of Western Australia’s coral reefs. Coral Reefs 38, 651–667 (2019).
BirdLife International. Important bird areas factsheet: Bedout Island, Downloaded from https://datazone.birdlife.org/site/factsheet/bedout-island-iba-australia (2023).
Chambers, L. E. et al. Observed and predicted effects of climate on Australian seabirds. Emu 111, 235–251 (2011).
Kingsley, M. R., Lavers, J. L., Steeves, T. E. & Burridge, C. P. Genetic distinctiveness of Masked Booby (Sula dactylatra) on Bedout Island, Western Australia. Emu. 120, 150–155 (2020).
Dias, M. P. et al. Threats to seabirds: A global assessment. Biol. Conserv. 237, 525–537 (2019).
Andersen, J. H. et al. Potential for cumulative effects of human stressors on fish, sea birds and marine mammals in Arctic waters. Estuar. Coast. Shelf Sci. 184, 202–206 (2017).
Zhang, Y. S. et al. Tropical cyclone impacts on seagrass-associated fishes in a temperate-subtropical estuary. PLoS One 17, e0273556 (2022).
O’Brien, K. R. et al. Seagrass ecosystem trajectory depends on the relative timescales of resistance, recovery and disturbance. Mar. Pollut. Bull. 134, 166–176 (2018).
Cheal, A. J., MacNeil, M. A., Emslie, M. J. & Sweatman, H. The threat to coral reefs from more intense cyclones under climate change. Glob. Change Biol. 23, 1511–1524 (2017).
Sæther, B.-E. & Bakke, Ø. Avian life history variation and contribution of demographic traits to the population growth rate. Ecology 81, 642–653 (2000).
Palestis, B. G. Nesting stage and nest defense by Common Terns. Waterbirds 28, 87–94 (2005).
Hamer, K. C., Schreiber, E. A. & Burger, J. Biology of marine birds (CRC Press, 2002).
King, B. R., Hicks, J. T. & Cornelius, J. Population-changes, breeding cycles and breeding success over six years in a seabird colony at Michaelmas Cay, Queensland. Emu 92, 1–10 (1992).
Ratcliffe, N., Pelembe, T. & White, R. Resolving the population status of Ascension Frigatebird Fregata aquila using a ‘virtual ecologist’ model. IBIS 150, 300–306 (2008).
de Korte, J. & Silvius, M. J. in Seabirds on islands, Threats, Case Studies and Action Plans Vol. BirdLife Conservation Series No. 1 (eds D. N. Nettleship, J Burger, & M Gochfeld) 77–93 (Birdlife International, 1994).
Feare, C. J. The decline of booby populations in the western Indian Ocean. Biology Conservation 14, 295–305 (1978).
BirdLife International. Species factsheet: Sula dactylatra, http://www.birdlife.org (2023).
Sandvik, H., Coulson, T. & Sæther, B. E. A latitudinal gradient in climate effects on seabird demography: Results from interspecific analyses. Glob. Change Biol. 14, 703–713 (2008).
Yorkston, H. D. & Green, P. T. The breeding distribution and status of Abbott’s booby (Sulidae: Papasula abbotti) on Christmas Island, Indian Ocean. Biol. Conserv. 79, 293–301 (1997).
Gilmour, J. P. & Smith, L. D. Category 5 cyclone at Scott Reef, northwestern Australia. Coral Reefs 25, 200–200 (2006).
Keogan, K. et al. Global phenological insensitivity to shifting ocean temperatures among seabirds. Nat. Clim. Change 8, 313–318 (2018).
Hällfors, M. H. et al. Shifts in timing and duration of breeding for 73 boreal bird species over four decades. Proc. Natl. Acad. Sci. 117, 18557–18565 (2020).
Grant, M. L., Bond, A. L. & Lavers, J. L. The influence of seabirds on their breeding, roosting, and nesting grounds: A systematic review and meta-analysis. J. Appl. Ecol. 91, 1266–1289 (2022).
Croll, D. A., Maron, J. L., Estes, J. A., Danner, E. M. & Byrd, G. V. Introduced predators transform subarctic islands from grassland to tundra. Science 307, 1959–1961 (2005).
Şekercioğlu, Ç. H., Daily, G. C. & Ehrlich, P. R. Ecosystem consequences of bird declines. Proc. Natl. Acad. Sci. 101, 18042–18047 (2004).
Pilbara Ports Authority. www.pilbaraports.com.au (Port Hedland, Western Australia, 2023).
Santos. Santos launches Dorado FEED (Perth, Western Australia, 2021) https://www.santos.com/news/santos-launches-dorado-feed/.
Fuller, P. J. & Burbidge, A. A. Bedout Island, Pilbara region, Western Australia. Corella 22, 113–115 (1998).
Lavers, J. L. Survey of seabirds breeding on Bedout Island during 22-24 November 2016 (Hobart, Tasmania, 2016).
Burbidge, A. A., Fuller, P. J., Lane, J. A. K. & Moore, S. A. Counts of nesting boobies and Lesser Frigatebirds in Western Australia. Emu, 87, 128–129 (1987).
Hartert, E. List of birds collected in north-western Australia and Arnhem Land by Mr J. T. Tunney. Novit. Zool 12, 194–242 (1905).
BOM. Severe Tropical Cyclone Rusty (Bureau of Meteorology, Melbourne, Australia, 2013).
BOM. Tropical Cyclone Christine (Bureau of Meteorology, Melbourne, Australia, 2014).
BOM. Severe Tropical Cyclone Ilsa (Bureau of Meteorology, Melbourne, Australia, 2023).
Acknowledgements
We acknowledge the traditional custodians of the land and waters where the data for this paper were collected and analysed, the Kariyarra, Ngarla, and Nyamal people of the Marapikurrinya region and Wudjari people of Kepa Kurl/Esperance. We celebrate the role that Indigenous people play as the first scientists and conservationists of this beautiful Country. We are grateful to Albert Burgman (linguist) and the Wangka Maya Pilbara Aboriginal Language Centre for providing cultural guidance on traditional names for seabirds and sites in the region. We thank Karl Lyons and Fortescue Helicopters Pty Ltd (pilots C. Mitchell and T. Chartres) for providing boat and helicopter access to Bedout Island that reflected the urgency of the situation, while ensuring bird and human safety were a priority. The Western Australian Department of Biodiversity, Conservation and Attractions (DBCA; permit no. FO2500047) and Charles Sturt University Animal Ethics Committee (permit no. A22382) provided permits and/or logistical support for this project, particularly A. Whittington, D. Stefoni, and H. Robertson. G.J. Robertson (Canadian Wildlife Service) provided useful statistical advice. Finally, our thanks to Merindah (Minnie) Bairnsfather-Scott (Noongar) for her generous assistance with Fig. 4 and Supplementary Fig. S3. Comments from two reviewers who kindly volunteered their time to provide valuable feedback on earlier versions of this manuscript.
Author information
Authors and Affiliations
Contributions
J.L.L.: project conception, data collection, data analysis, writing – original draft. T.M.M.: data collection, image production. A.L.F.: data collection, image production. A.L.B.: data analysis, writing – editing and reviewing.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Communications Earth & Environment thanks the anonymous reviewers for their contribution to the peer review of this work. Primary Handling Editor: Aliénor Lavergne. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lavers, J.L., Mead, T.M., Fidler, A.L. et al. Cyclone Ilsa in April 2023 led to significant seabird mortality on Bedout Island. Commun Earth Environ 5, 276 (2024). https://doi.org/10.1038/s43247-024-01342-6
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s43247-024-01342-6
