
Abstract
Floral induction in litchi is influenced by multiple environment cues including temperature and soil water condition. In the present study, we determined that a combined treatment consisting of 14-day drought imposed prior to exposure to 35-day low temperature (T3) significantly promoted litchi flowering relative to the low temperature alone (T2), suggesting an integrative effect of drought and low temperature on litchi floral initiation. Analysis of transcriptomic changes in leaves from different treatments showed that 2,198 and 4,407 unigenes were differentially expressed in response to drought and low temperature, respectively. 1,227 of these unigenes were expressed in response to both treatments, implying an interaction of drought and low temperature on expression of genes involved in litchi floral initiation. Additionally, 932 unigenes were consistently differentially expressed during floral induction between T2 and T3 plants, which potentially accounts for the difference of flowering time. Thirty-eight transcription factors out of these 932 unigenes were identified as hub genes with central roles in regulation of litchi floral induction. The expression of litchi homologs of well-known flowering genes was also investigated and one Flowering Locus T (FT) homolog may play a crucial role in litchi flowering in responses to drought and low temperature.
Similar content being viewed by others


Introduction
Litchi (Litchi chinensis Sonn.) is an economically important evergreen fruit tree in southern China and is widely distributed in the subtropics as well. The annual yield of litchi is mainly determined by successful floral initiation that is influenced by multiple endogenous and environmental cues, including temperature and soil water condition1,2. However, worldwide climate changes are likely to adversely affect floral initiation and flower development.
Low temperature plays an essential role in flowering of vernalization-dependent plants1 and is also required for litchi floral initiation3,4, although the cold requirement varies among the different varieties5. Exposure to cold temperature in winter or early spring promotes litchi floral initiation, while ambient temperature above 20 °C significantly reduces litchi flowering6. Unusual high temperatures in winter can result in insufficient cold accumulation for litchi floral initiation, leading to inadequate flowering. Previous studies in the field have showed that to a certain extent, drought prior to winter cold induction can promote litchi flowering by reducing the requirement of cold temperature7,8. However, drought treatment alone does not induce litchi flowering5. Despite the agronomic importance of this integrative regulation of litchi floral induction, little is known about the molecular basis of the interaction.
The genetic mechanism underlying the vernalization response has been intensively studied in the model plant Arabidopsis1,9,10. In Arabidopsis, Flowering Locus T (FT), encoding a florigen protein, plays a central role in promoting flowering by integrating various flowering pathways11. In contrast, Terminal flower 1 (TFL1), sharing a highly conserved amino acid sequence with FT, exhibits an antagonistic role in plant flowering due to a single amino acid mutation12,13. However, under non-inductive condition expression of FT is strongly inhibited by a MADS box transcription factor Flowering Locus C (FLC), a major floral repressor in the vernaliztion pathway, which also represses expression of another important flowering promoter SUPPRESSOR OF OVEREXPRESSION OF CO1 (SOC1)14,15,16,17. Prolonged exposure to low temperature (vernalization) can alleviate the repression of FT by decreasing expression of FLC, but stable repression of FLC requires activities of VERNALIZATION1 (VRN1)18,19. On the other hand, activation of FT can be facilitated by CONSTANS (CO), an important regulator in the photoperiodic flowering pathway20. Previous studies have shown that day length-dependent genes including CO can play complementary and important roles in temperature signaling in floral initiation21,22. However, the integrative effect of drought and temperature on plant flowering-genes has yet to be further studied.
In the present study, drought treatment, low temperature treatment and drought followed low temperature treatment were imposed on potted litchi plants to investigate the effects of water stress and low temperature on floral initiation. Next generation RNA-Sequencing was applied to obtain the overview of leaf transcriptome changes occurred during floral induction. The expression profiles of unigenes from different treatments were multiply compared and characterized to identify potentially important genes involved in litchi floral initiation in responses to drought and cold. Furthermore, the co-expression networks among important genes were established according to weighted gene co-expression network analysis (WGCNA). The hub genes that potentially play central regulative roles in litchi floral induction were also isolated. Together, the results provide evidence for regulation of litchi floral induction and establish a foundation for advanced research on functional flowering-genes in litchi or similar plants.
Results
Drought treatment promotes litchi flowering
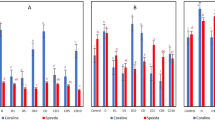
To study the effect of drought on litchi floral initiation, potted litchi trees were subjected to treatments as indicated in Fig. 1 to permit control and monitoring of soil moisture. When the plants were fully watered, the soil moisture was about 28% and the corresponding leaf water potential was near -1 MPa. Under the drought treatments, soil moisture levels were maintained at around 8% (T3) and 15% (T4 and T5) corresponding to about 25% and 50% of control and the corresponding water potentials were around −2.5 MPa and −1.5 MPa, respectively (Fig. 2). The trees were fully watered when the low temperature treatment started.

The experiment scheme of litchi floral induction conducted in a greenhouse.

The absolute soil moisture in the pots (A) and water potential of litchi leaves (B) under different treatments.
After the treatments, plants were transferred to room temperature, fully watered and the flowering time and the flower quantity were recorded. There were no flowers in T1 plants and T5 plants rarely flowered, suggesting that low temperature is required for litchi floral initiation. All of the plants under T2, T3 and T4 treatments bloomed, even though the flower quantity, such as rates of flowering shoots, length and width of panicle of each treatment varied (Table 1). Importantly, T3 and T4 plants were found to flower about six and three days earlier, respectively, than T2 plants, indicating that certain pre-treatment of drought prior to low temperature induction promoted litchi flowering, consistent with the previous study in open field8. In the next year, T2 and T3 treatments were repeated to validate the promotion of floral initiation by drought. The results showed that the T3 litchi plants flowered about four days earlier in comparison with T2 plants and the percentage of flowering plants of T2 plants decreased to 25% while 100% of T3 plants flowered.
Analysis of dynamic changes in leaf transcriptome with RNA-Seq
To investigate the causes of differential flowering time between T2 and T3 plants, the dynamic changes in leaf transcriptome profiles were obtained by high-throughput RNA-Seq. After removing possible rRNA reads, each library produced 19.9 to 32.0 million clean reads (Table 2). These clean reads were assembled into unigenes by using the litchi genome database as a reference (http://litchidb.genomics.cn) with the mapping rate ranging from 88.7% to 92.4% for each library. Between 34,067 to 41,173 unigenes were obtained with a mean length of 1,371 bp and an average N50 length of 1,735 bp. Approximately, 91% of these unigenes had top matches to 8 species based on Nr annotation (Figure S1). More than half of these unigenes could be annotated with gene sequences from two Citrus species, i.e. Citrus clementina and Citus sinensis. In comparison with our previous RNA-Seq data from litchi leaf and bud23, the average length of unigenes is more than doubled, likely due to advances in the sequencing technology.
Genes in responses to drought and low temperature during litchi floral initiation
The FPKM method was used to calculate the expression of unigenes and all the transcriptome files of leaves from different time points for the different treatments were pairwise compared. Spearman correlation between samples was calculated based on FPKM values using the function “cor” in R package and displayed in a heatmap (Fig. 3). Generally, these samples were categorized into three clusters: samples under room temperature and drought treatments, samples under low temperature treatment and samples transferred to room temperature after low temperature treatment. Notably, samples under the drought treatment were grouped together except for the sample from T3-14d, implying that drought treatment caused obvious molecular changes on the 14th day. Importantly, the Spearman correlation coefficient of T2-15d and T3-15d is lower than that of T1-14d and T3-14d, which means the molecular difference between T1-14d and T3-14d plants was enlarged, although both of the T2-15d and T3-15d plants underwent one-day low temperature treatment, suggesting that an integrative effect of drought and low temperature is required for induction of certain stress-responsive genes. Hence, the genes in response to drought, low temperature and even the combination need to be characterized.

Heat map showing the spearman correlation coefficient of pairwise comparison between samples.
The spearman correlation coefficient between two samples was calculated based on FPKM of each gene. The heatmap was constructed using R language with the function of heatmap.
According to the Venn diagram of pairwise comparisons of transcriptomes from T1-14d, T3-14d, T2-15d and T3-15d samples, 2,198 unigenes were differentially expressed in T3 plants after 14-day drought treatment (Fig. 4A). Notably, one-day low temperature treatment resulted in a different number of cold-responsible genes in T2 and T3 plants and totally 4,407 unigenes were differentially expressed in T2 and T3 plants in response to one-day low temperature treatment (Fig. 4A), which is consistent with the earlier analysis of Spearman correlation coefficient between samples in Fig. 3. These stress-genes are potentially involved in litchi floral initiation, responding to drought and low temperature. Among these differentially expressed genes (DEGs), 1,227 unigenes were expressed in response to both drought and low temperature, suggesting an interaction of low temperature and drought on genes involving in litchi floral initiation. The KEGG analysis of these 1,227 DEGs showed that genes related to Biosynthesis of secondary metabolites, Starch and sucrose metabolism and Plant hormone signal transduction represented the major proportion (Fig. 4B), indicating the intricate network between external stress cues and endogenous metabolism and signaling. The GO analysis classified these 1,227 unigenes into three categories: biological process, cellular component and molecular function (Fig. 4C). For the biological process category, metabolic process and cellular process represented the major proportion. Under the cellular component category, large numbers of unigenes were categorized as cell and cell part. Under the molecular function category, catalytic activity and binding were the top two most abundant sub-categories.

Identification and functional annotation of the differentially expressed genes (DEGs) in responses to low temperature and drought treatments.
(A) Venn diagram of the DEGs derived from pairwise comparisons. KEGG enrichment (B) and Gene Ontology classification (C) of the genes in responses to both floral induction treatments.
Genes accounting for the different flowering time between T2 and T3 litchi plants
T3 litchi plants flowered 3–7 days earlier than T2 plants did. To identify the gene(s) accounting for the difference, the transcriptomes of T2 and T3 plants at treatment time points of 15, 21 and 49-days were accordingly pairwise compared. Totally, there were 3,988 genes differentially expressed (Fig. 5), which might account for the difference of flowering time between T2 and T3 litchi plants. However, among these 3,988 DEGs, only 458 unigenes (11.5% of 3,988 unigenes) were differentially expressed in at least two pairwise comparisons across the low temperature treatment. Therefore, expression pattern and consistency of these potential candidate genes should be taken into account.

Venn diagram of the differentially expressed genes (DEGs) derived from pairwise comparisons between T2 and T3 plants at 15, 21 and 49-day time points under low temperature treatment.
A co-expression network was established on the basis of weighted gene co-expression network analysis (WGCNA), to analyze the gene expression pattern and linkage. Totally, 24,537 unigenes that were differentially expressed in at least one pairwise comparison of samples were used for WGCNA; 24,242 unigenes were categorized into 18 distinct modules and the other 295 genes were outliers (Fig. 6). Among these modules, the bottom three show an irregular expression trend. The top seven consisted of genes mainly expressed in leaves aging under room temperature are likely to be responsible for leaf development and senescence. Of particular interest is the identification of the flowering-related genes in responses to stress. Genes expressed in responses to drought and low temperature comprise the middle eight co-expression modules, of which four (cyan, steelblue, black and grey60) display drought-response and the other four (salmon, brown, turquoise and light_cyan) show low temperature-response. However, only black, grey60, turquoise and light_cyan modules show p-value ≤0.05 under drought and low temperature treatments, as indicated by cluster analysis in Figure S2. The eigengene expression of these 4 modules comprised of 8,029 genes is shown in Figure S3. 932 out of the 8,029 genes are shared in the DEGs between T2 and T3 at 15, 21 and 49-day time points (Figure S4). WGCNA was carried out on these 932 genes to construct the network, in which genes are indicated as nodes and the connecting lines (edges) with red and green color correspondingly represent the positive and negative co-expression correlations (Fig. 7). The statistical connection of all these 932 unigenes in the network suggested that these genes tend to be inner-correlated. In addition, 38 out of 932 unigenes are identified as transcription factors likely playing central roles in litchi floral initiation in responses to drought and low temperature.

Heatmap showing the weighted gene co-expression network analysis (WGCNA) of genes between different litchi leaf samples.
Each row corresponds to a module and each column corresponds to a time point of treatment. The top and bottom number in each cell indicate the correlation coefficient between the module and time point and p-value of the test, respectively.

The co-expression network of the genes potentially accounting for the difference of flowering time between T2 and T3 plants.
The co-expression network was constructed from 932 genes from 4 modules.
Expression profiles of flowering genes in leaves during litchi floral induction
The homologues of well-known flowering-related genes in litchi were also investigated in the present study (Fig. 8 and Figure S5). As revealed by the gene expression patterns obtained from RNA-Seq data, two CONSTANS genes (LcCO1 and 2) and LcSOC1-1 were up-regulated by drought treatment. Two VERNALIZATION genes (LcVRN-1 and 2) were induced while two TERMINAL FLOWERING LOCUS genes (LcTFL-1 and 2) were reduced by low temperature exposure. Two APETALA1 (LcAP1-1 and 2), one FLOWERING LOCUS T (LcFT1) and LcSOC1-3 were induced by both treatments. Of particular importance, the induction of LcFT1 by drought treatment was larger after exposure to low temperature resulting in significantly higher transcripts level of LcFT1 in T3 plants compared to T2 plants (Fig. 8).

Expression confirmation of eleven flowering-related genes using real time Q-PCR.
The relative expression of target genes relative to a control gene (LcActin) in real time Q-PCR was shown as yellow column with standard error and their corresponding fold changes (relative to the value of T1_0d) of FPKM value from RNA-seq were shown as blue lines. The Spearman correlation coefficient (r) between the gene expression values derived from the two methods and associated p value (n = 21) were shown accordingly.
To evaluate if and how the FPKM value represents the transcript level, expression of these eleven flowering-related genes was measured using real time Q-PCR. The relative expression of eight genes was consistent with their FPKM values obtained from RNA-Seq (r = 0.44 to 0.87). However, expression patterns of the other three genes were not strictly correlated with the RNA-Seq data (Fig. 8), possibly due to RNA alternative splicing. Alternative RNA splicing is common and leads to multiple transcripts (isoforms) per gene. The numbers and types of isoforms for genes are tissue-specific and vary during the life cycle24. Therefore, the isoforms can be accurately determined by RNA-Sequencing, but sometimes it is not easy to measure the expression by Q-PCR because it is difficult to design primers to quantify an isoform of a gene expressed in different tissue.
Discussion
Plant flowering time is determined by endogenous signals and environmental cues, including phytohormones, photoperiod, vernalization process and phase transition from juvenile to adult9,25,26. In nature, plants are usually exposed to multiple external cues and must facilitate the integration of this comprehensive information into a flowering network21,27,28,29.
In the case of litchi, low temperature is highly required for floral initiation, but unusually high temperature in winter can potentially lead to insufficient cold accumulation resulting in inadequate flowering. In an effort to enhance litchi flowering, agronomic regulation is sought to alleviate the adverse effect of climate change. In the present study conducted in a greenhouse, drought treatment followed by cold temperature induction caused earlier flowering in litchi compared to only cold temperature treatment (Table 1), which is consistent with studies in open field8 and other species30. This result is also supported by previous studies that plants tend to accelerate flowering and produce seeds in response to abiotic stress such as clod temperature, water deficit and UV-C exposure before severe stress leads to death1,31,32. However, drought treatment without subsequent low temperature exposure did not induce litchi flowering5 and a similar result was also observed in our study, where 49-day mild drought resulted in a small number of flowers (Table 1) suggesting that drought cannot replace the low temperature to induce litchi floral initiation33. Although a previous study showed a dose-dependent drought effect, i.e. the effect of drought depends on the severity, duration and even the genotype34, intensive or long-term drought stress may negatively affect flower development and fertility9,35, reducing crop productivity.
To investigate the molecular basis of the integrative effect of drought and low temperature on litchi floral initiation, the dynamic changes in transcriptomes in leaves under different treatments were studied using RNA-Seq technology36,37. According to the overview of the transcriptomes profiles (Fig. 3), the early responses of litchi plants to drought at a molecular level are not apparent suggesting the dosage-dependent effect of drought38 and thus we mainly analyzed the drought-responsive genes in leaves at the 14-day time point where 2,198 genes were found to be regulated by drought. Compared to drought, one-day-low temperature treatment caused remarkable changes in gene expression and about one-third of these genes responded to drought stress (Fig. 4) implying an interaction of low temperature and drought39. The KEGG analysis shows that these genes responding to both low temperature and drought treatment are mainly related to biosynthesis of secondary metabolites, starch and sucrose, as well as plant hormone signal transduction, which has been suggested to be associated with floral initiation and flower development. For example, phytohormone signaling is known to participate in various plant morphogenetic processes, including the switch from vegetative to reproductive growth40. In Arabidopsis, GA is considered to be the major promoting signal in flowering41,42, while ABA commonly antagonizes GA signaling40. However, ABA likely has a positive role in floral initiation in litchi plants43. Sugars are traditionally regarded as metabolic resources for energy supply and carbon skeleton construction, but recently sugars have been suggested to serve as signals in the plant life cycle. Plants containing mutations in starch biosynthesis or sugars transporter genes exhibit delayed-flowering44,45. Interestingly, one-day low temperature treatment induced different sets of cold-responsible genes in control and drought-treated plants (Fig. 4) indicates the integrative effect of drought and low temperature on gene expression. This result may indicate that either drought-mediated genes are required for activation of some low temperature-response genes, or that drought-treated plants and well-watered plants respond to low temperature in quite different manners.
As a consequence of the interaction of drought and low temperature, 3,988 unigenes were differentially expressed between T2 and T3 plants (Fig. 5), 932 out of these DEGs were consistently expressed in response to drought or low temperature based on further analysis using WGCNA (Fig. 6; Figure S3). Moreover, some litchi homologs of well-known flowering genes were found in the co-expression modules derived from the 932 genes (Fig. 8). For example, one litchi FT homolog (LcFT1) from light_cyan module was induced by drought and low temperature, similar to what was observed in Arabidopsis32. Epigenetically over-expression of this FT homolog in Arabidopsis results in significantly early flowering46, indicating the conserved functions of these genes in litchi plants47,48.
Generally, vernalizaiton or low temperature activate expression of FT via reducing the expression of FT-repressors including FLC and TFL. In the present study, low temperature did not alter the expression of FLCs, but did significantly decrease the expression of TFLs, which potentially alleviated the repression of expression and transport of FT; thus, TFL rather than FLC seems to be the major negative flowering regulator during floral induction (Fig. 8 and Figure S5). The expression of FT also requires upstream activators including CO that is induced not only by long photoperiod49, but also by low temperature and drought, as proposed by our data (Fig. 8 and Figure S5), leading to induction of FT and SOC150 and eventually promotion of flowering.
Materials and Methods
Plant materials
Three-year-old air layered potted litchi trees (Litchi chinenesis cv. Feizixiao) were grown in a green house on the campus of South China Agricultural University in Guangzhou City with loam, coconut chaff and mushroom cinder (3:1:1). The terminal growing flushes of twenty-one plants were removed one week prior to treatments for similar plant growth status for individual plant.
Experiment scheme
Trees were randomly divided into 5 groups with 4 or 5 plants per group and subjected to treatments as indicated in Fig. 1 in a growth chamber with adjustable temperature and humidity. In the beginning, all the plants were grown under room temperature (25 ± 2 °C); while T3, T4 and T5 plants were drought stressed with the soil moisture corresponding to 25%, 50% and 50% respectively of the T1 and T2 plants which were fully watered. For the drought treatments, the relative air humidity in the chamber was adjusted to 30% to accelerate the water evaporation. After 14 days, T2, T3 and T4 plants were transferred to a low temperature chamber (15 °C) and fully watered. However, T1 and T5 plants were kept under room temperature, with T5 plants maintained at 50% relative soil moisture. Thirty-five days later, plants were transferred to room temperature chamber and fully watered. These pots were moved daily to avoid the possible overlap of leaves. The leaf water potential and relative soil moisture were monitored when the drought treatment was carried out. The third to sixth compound leaves were sampled at 0, 1, 3, 7, 14, 15, 21 and 49-day time points named as T1-0…49d accordingly and final sampling was occurred when any panicle primordia emerged namely T1-F. The plants for T1 and T2 treatments were not separated until low temperature treatment was imposed on T2 plants.
Drought treatments, measurement of leaf water potential and soil moisture
Drought treatment was carried out as described by Su et al.51 with little modification. At the start, plants were fully watered and soil moisture was measured 2 h later by Theta probe type ML2x (Delta-T Devices Ltd; Cambridge England), which was indicated as soil water capacity. For the drought-treated plants, the soil water naturally evaporated until reaching the set level. Then the soil moisture was maintained by accurately watering according to the daily loss of water. Leaf water potential was measured with a pressure chamber (PMS Instrument Co., Corvallis, Oregon, USA). The soil moisture was measured three times per pot and leaf water potential was detected once per plant with the values indicated as the means ±SE of 4–5 plants.
RNA extraction, library construction and next generation RNA-Sequencing
Total leaf RNA was extracted using Hipure Plant RNA Mini Kit (Magen, China) according to the manufacturer’s instruction and treated with DNase (Takara, China). Equal quantities of RNA from three biological replicates were pooled to construct libraries that were sequenced with Illumina HiSeqTM 2500 using a 125 paired-end (PE) module at ANNOROAD Company (Beijing, China). The raw reads were trimmed and filtered using Trimmomatic software (v0.33) to remove adapters and low quality bases. The high-quality reads were aligned to the sillva SSU and LSU ribosome RNA (rRNA) database and the matched reads were removed to generate clean reads.
Assembly and annotation
Clean reads were assembled into transcripts using Cufflinks with the litchi genome as a reference. Cuffmerge was used to assemble the transcripts into non-redundant unigene sets. The functions of unigenes were annotated by blastXing to NCBI non-redundant (nr) database, Swiss-Prot protein database and Kyoto Encyclopedia of Genes and Genomes (KEGG) database with E-value threshold of 10−5 and minQueryCo (v0.33). To generate a broad overview of groups of genes, Gene Ontology (GO) classification was applied. GO annotation was obtained by id conversion from NCBI gene ID number to GO id leveraging the mapping relation downloaded from PIR (ftp://ftp.pir.georgetown.edu/databases/idmapping/idmapping.tb.gz) using TBtools software (http://www.cjchen.name/TBtools.jar).
Gene expression profiling
All sequences of transcripts were extracted from reference genome sequence using gffread from cufflinks pipeline. The expression calculation at transcript and gene level was conducted using RSEM (v1.2.21) with default parameters. “Fold change ≥2, p value ≤0.05” was set as the threshold for significant difference in expression.
Construction of gene co-expression networks and heat map
The ‘cor’ function in R programming language and software (http://www.r-project.org/) was used to calculate Spearman correlation coefficients between samples. A co-expression network was constructed using weighted gene co-expression network analysis (WGCNA, v1.48) package in R. Totally 24,537 genes that were differentially expressed in at least one pairwise comparison of samples were used for analysis. The log2 (FPKM + 1) values of these genes were imported into WGCNA to generate the modules with default setting except that the soft power is 5, min module size is 100, deep split is 1 and merged cut height is 0.25. The total connectivity and intra-modular connectivity (function soft Connectivity), eigengene-based connectivity (kME) and kME-P value were calculated for the 24,242 genes, which were clustered into different modules and remaining 295 genes were outliers. The heat map was constructed using the R package “pheatmap”.
Real time quantitative PCR (Q-PCR) analysis
Relative expression of flowering genes in leaf tissues was measured using Q-PCR performed on LightCycler 480 II (Roche, Germany) with iTaq Universal SYBR Green Supermix (BioRad, USA) according to the manufacturer’s instruction. For each sample three biological replicates were taken based on which the average value and standard error were calculated. PCR amplification was conducted under the following conditions: 95 °C for 10 min, followed by 40 cycles at 95 °C for 10 sec, then at 60 °C for 20 sec and finally 72 °C for 20 sec. PCR products were melted at 95 °C for 5 sec and then 65 °C for 1 min to detect the specificity. The gene expressions were normalized against an internal reference gene LcActin (HQ615689). Primers used in this study were designed on website of Primer 3 (v 0.4.0; http://bioinfo.ut.ee/primer3-0.4.0/primer3/) and showed in Figure S6.
Additional Information
How to cite this article: Shen, J. et al. Integrative effect of drought and low temperature on litchi (Litchi chinensis Sonn.) floral initiation revealed by dynamic genome-wide transcriptome analysis. Sci. Rep. 6, 32005; doi: 10.1038/srep32005 (2016).
References
Kim, D. H., Doyle, M. R., Sung, S. & Amasino, R. M. Vernalization: Winter and the timing of flowering in plants. Annu. Rev. Cell Dev. Biol. Palo Alto, Annual Reviews. p. 277–299 (2009).
Vinocur, B. & Altman, A. Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations. Curr. Opin. Biotech. 16, 123–132 (2005).
O’Hare, T. J. Interaction of temperature and vegetative flush maturity influences shoot structure and development of lychee (Litchi chinensis Sonn.). Sci. Hortic. -Amsterdam 95, 203–211 (2002).
O’Hare, T. J. Impact of root and shoot temperature on bud dormancy and floral induction in lychee (Litchi chinensis Sonn.). Sci. Hortic. – Amsterdam 99, 21–28 (2004).
Menzel, C. M. & Simpson, D. R. Effect of temperature on growth and flowering of litchi (Litchi chinensis Sonn.) cultivars. J. Hort. Sci. and Biotech. 63, 349–360 (1990).
Menzel, C. M. & Simpson, D. R. Temperatures above 20 degrees C reduce flowering in lychee (Litchi chinensis Sonn.). J. Hort. Sci. 70, 981–987 (1995).
Carr, M. K. V. & Menzel, C. M. The water relations and irrigation requirements of lychee (litchi chinensis Sonn.): A review. Exp. Agr. 50, 481–497 (2014).
Stern, R. A. et al. Effect of fall irrigation level in ‘Mauritius’ and ‘Floridian’ lychee on soil and plant water status, flowering intensity and yield. J. Am. Soc. Hortic. Sci. 123, 150–155 (1998).
Amasino, R. Seasonal and developmental timing of flowering. Plant J. 61, 1001–1013 (2010).
Turck, F. & Coupland, G. When vernalization makes sense. Science 331, 36–37 (2011).
Corbesier, L. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science 316, 1030–1033 (2007).
Hanzawa, Y., Money, T. & Bradley, D. A single amino acid converts a repressor to an activator of flowering. PNAS 102, 7748–7753 (2005).
Hanano, S. & Goto, K. Arabidopsis terminal flower1 is involved in the regulation of flowering time and inflorescence development through transcriptional repression. Plant Cell 23, 3172–3184 (2011).
Rouse, D. T., Sheldon, C. C., Bagnall, D. J., Peacock, W. J. & Dennis, E. S. FLC, a repressor of flowering, is regulated by genes in different inductive pathways. Plant J. 29, 183–191 (2002).
Immink, R. G. H. et al. Characterization of SOC1’s central role in flowering by the identification of its upstream and downstream regulators. Plant Physiol. 160, 433–449 (2012).
Searle, I. et al. The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis. Genes Dev. 20, 898–912 (2006).
Seo, E. et al. Crosstalk between cold response and flowering in Arabidopsis is mediated through the flowering-time gene SOC1 and Its upstream negative regulator FLC. Plant Cell 21, 3185–3197 (2009).
Levy, Y. Y., Mesnage, S., Mylne, J. S., Gendall, A. R. & Dean, C. Multiple roles of Arabidopsis VRN1 in vernalization and flowering time control. Science 297, 243–246 (2002).
Oliver, S. N., Deng, W. W., Casao, M. C. & Trevaskis, B. Low temperatures induce rapid changes in chromatin state and transcript levels of the cereal vernalization1 gene. J. Exp. Bot. 64, 2413–2422 (2013).
Tiwari, S. B. et al. The flowering time regulator constans is recruited to the flowering locus T promoter via a unique cis-element. New Phytol. 187, 57–66 (2010).
Ream, T. S. et al. Interaction of photoperiod and vernalization determines flowering time of Brachypodium distachyon. Plant Physiol. 164, 694–709 (2014).
Fernandez, V., Takahashi, Y., Le Gourrierec, J. & Coupland, G. Photoperiodic and thermosensory pathways interact through CONSTANS to promote flowering at high temperature under short days. Plant J. 86, 426–440 (2016).
Zhang, H. N. et al. Transcriptomic analysis of floral initiation in litchi (Litchi chinensis Sonn.) based on de novo RNA sequencing. Plant Cell Rep. 33, 1723–1735 (2014).
Pan, Q., Shai, O., Lee, L. J., Frey, J. & Blencowe, B. J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 40, 1413–1415 (2008).
Andres, F. & Coupland, G. The genetic basis of flowering responses to seasonal cues. Nat. Rev. Genet. 13, 627–639 (2012).
Verhage, L., Angenent, G. C. & Immink, R. G. H. Research on floral timing by ambient temperature comes into blossom. Trends Plant Sci. 19, 583–591 (2014).
Mouradov, A., Cremer, F. & Coupland, G. Control of flowering time: Interacting pathways as a basis for diversity. Plant Cell 14, S111–S130 (2002).
Song, Y. H., Ito, S. & Imaizumi, T. Flowering time regulation: photoperiod- and temperature-sensing in leaves. Trends Plant Sci. 18, 575–583 (2013).
Zhou, C. M. et al. Molecular Basis of Age-dependent vernalization in Cardamine flexuosa. Science 340, 1097–1100 (2013).
Sharp, R. G., Else, M. A., Cameron, R. W. & Davies, W. J. Water deficits promote flowering in Rhododendron via regulation of pre and post initiation development. Sci. Hortic. -Amsterdam 120, 511–517 (2009).
Martinez, C., Pons, E., Prats, G. & Leon, J. Salicylic acid regulates flowering time and links defence responses and reproductive development. Plant J. 37, 209–217 (2004).
Riboni, M., Galbiati, M., Tonelli, C. & Conti, L. GIGANTEA enables drought escape response via abscisic acid-dependent activation of the florigens and suppressor of overexpression of constans1. Plant Physiol. 162, 1706–1719 (2013).
Chaikiattiyos, S., Menzel, C. M. & Ramussen, T. S. Floral induction in tropical fruit trees: Effects of temperature and water supply. J. Hortic. Sci. 69, 397–415 (1994).
Xu, Z., Zhou, G. & Shimizu, H. Plant responses to drought and rewatering. Plant signal. Behav. 5, 649–654 (2010).
Cerekovic, N., Pagter, M., Kristerisen, H. L., Brennan, R. & Petersen, K. K. Effects of deficit irrigation during flower initiation of two blackcurrant (Ribes nigrum L.) cultivars. Sci. Hortic. –Amsterdam 168, 193–201 (2014).
Martin, L. B. B., Fei, Z. J., Giovannoni, J. J. & Rose, J. K. C. Catalyzing plant science research with RNA-seq. Front. Plant Sci. 4, 10 (2013).
Wang, Z., Gerstein, M. & Snyder, M. RNA-Seq: a revolutionary tool for transcriptomics. Nat. Rev. Genet. 10, 57–63 (2009).
Claeys, H., Van Landeghem, S., Dubois, M., Maleux, K. & Inze, D. What Is Stress? Dose-response effects in commonly used in vitro stress assays. Plant Physiol. 165, 519–527 (2014).
Shi, H. T., Chen, Y. H., Qian, Y. Q. & Chan, Z. L. Low Temperature-Induced 30 (LTI30) positively regulates drought stress resistance in Arabidopsis: Effect on abscisic acid sensitivity and hydrogen peroxide accumulation. Front. Plant Sci. 6, 9 (2015).
Davis, S. J. Integrating hormones into the floral-transition pathway of Arabidopsis thaliana. Plant Cell Environ. 32, 1201–1210 (2009).
Mutasa-Gottgens, E. & Hedden, P. Gibberellin as a factor in floral regulatory networks. J. Exp. Bot. 60, 1979–1989 (2009).
Yamaguchi, N. et al. Gibberellin acts positively then negatively to control onset of flower formation in Arabidopsis. Science 344, 638–641 (2014).
Cui, Z., Zhou, B., Zhang, Z. & Hu, Z. Abscisic acid promotes flowering and enhances LcAP1 expression in Litchi chinensis Sonn. S. Afr. J. Bot. 88, 76–79 (2013).
Corbesier, L., Lejeune, P. & Bernier, G. The role of carbohydrates in the induction of flowering in Arabidopsis thaliana: Comparison between the wild type and a starchless mutant. Planta (Berlin) 206, 131–137 (1998).
Wang, B. et al. UGT87A2, an Arabidopsis glycosyltransferase, regulates flowering time via flowering locus C. New Phytol. 194, 666–675 (2012).
Ding, F. et al. Promoter difference of LcFT1 is a leading cause of natural variation of flowering timing in different litchi cultivars (Litchi chinensis Sonn.). Plant Sci. 241, 128–137 (2015).
Kazan, K. & Lyons, R. The link between flowering time and stress tolerance. J Exp Bot. 67, 47–60 (2016).
Shinozaki, K. & Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 58, 221–227 (2007).
Valverde, F. et al. Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science 303, 1003–1006 (2004).
Hepworth, S. R., Valverde, F., Ravenscroft, D., Mouradov, A. & Coupland, G. Antagonistic regulation of flowering-time gene SOC1 by CONSTANS and FLC via separate promoter motifs. EMBO J. 21, 4327–4337 (2002).
Su, Z. et al. Flower development under drought stress: Morphological and transcriptomic analyses reveal acute responses and long-term acclimation in Arabidopsis. Plant Cell 25, 3785–3807 (2013).
Acknowledgements
We thank Professor Biyan Zhou for her critical advice on the design of this project. We acknowledge Professor Harry Klee for his critical comments on writing this paper. This work was supported by China Agricultural Research Service (CARS-33) and Guangdong Natural Science Foundation (2016A030310458).
Author information
Authors and Affiliations
Contributions
J.S. carried out the experiments, analyzed the RNA-Seq data and wrote the manuscript. Q.X. and H.Q. participated in doing the experiments. C.C. contributed to the data analysis. H.C. designed the project and reviewed the manuscript. All authors read and approved the final manuscript.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Shen, J., Xiao, Q., Qiu, H. et al. Integrative effect of drought and low temperature on litchi (Litchi chinensis Sonn.) floral initiation revealed by dynamic genome-wide transcriptome analysis. Sci Rep 6, 32005 (2016). https://doi.org/10.1038/srep32005
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep32005
This article is cited by
-
Transcriptomic and physiological analysis reveals interplay between salicylic acid and drought stress in citrus tree floral initiation
Planta (2022)
-
Species-specific genes account for the differences in floral transition between continuous-flowering and once-flowering roses
Journal of Plant Biochemistry and Biotechnology (2019)
