
Abstract
Individuals at clinical high-risk (CHR) of developing psychosis, as well as patients with recent psychosis onset (RO), experience significant negative symptoms that detrimentally impact daily-life functioning and are associated with poor outcomes, even in those who do not convert to psychosis. Targeting negative symptoms may thus hold promise for the treatment of CHR and RO patients. Building from previous findings we examined whether the catechol-O-methyltransferase (COMT) Val158Met functional polymorphism and fasting peripheral proline concentration predicts the severity of negative symptoms experienced by adolescents and young adults at CHR or those with RO. As hypothesized, the interaction between fasting plasma proline and COMT predicted negative symptoms, as measured via the Scale for the Assessment of Negative Symptoms (SANS) total (n = 50, β = 0.066, adjusted p = 0.007) and global severity scores (n = 50, coefficient = 0.026, adjusted p = 0.003): Higher proline was beneficial for Val/Val subjects, but detrimental to those with the Met allele. In a secondary analysis, the COMT x proline interaction also predicted symptoms measured via the Clinical Assessment Interview for Negative Symptoms (CAINS) total scores (n = 50, β-coefficient = 0.035, adjusted p = 0.044), although this result did not reach the Benjamini-Hochberg’s threshold for significance. Further, there was a trend towards significance for an association with social and interpersonal function (Global Functioning-Social, coefficient = −0.005, adjusted p = 0.055). Negative symptoms are intractable and largely unaddressed by current medications. This study further supports a relationship between peripheral proline and COMT influencing negative symptoms such as anhedonia, in young CHR individuals and those with RO. That higher proline has converse effects on symptoms by COMT may have implications for the development of therapeutics to intervene early and specifically target the interaction pathway.
Similar content being viewed by others
Introduction
Individuals at clinical high-risk (CHR) for developing psychosis, as well as patients with recent psychosis onset (RO), exhibit significant negative symptoms such as anhedonia, avolition, blunted affect, and social withdrawal, which are associated with poor functional outcomes and severely impact quality of life [1, 2]. Negative symptoms are also associated with psychosis onset [3], however, even those CHR individuals who do not develop threshold psychosis often continue to exhibit negative symptoms that lead to substantial impairments in social and occupational functioning [2].
Early treatment of psychosis is associated with more favorable clinical outcomes [4, 5], and it has been suggested that intervention during the prodrome may delay the onset of full-blown psychotic symptoms [5]. That negative symptoms are associated with both the onset of psychosis and poor outcomes, even in those who do not convert, suggests that specifically targeting negative symptoms may hold promise for the treatment of both CHR and RO patients.
In previous work we demonstrated that fasting peripheral levels of the amino acid and likely CNS neuromodulator proline [6], coupled with differential activity of the catechol-O-methyltransferase (COMT) enzyme (as assessed via the COMT Val158Met functional polymorphism), which plays a primary role in the metabolism of catecholamine neurotransmitters (including dopamine) in the prefrontal cortex, interact to predict negative symptom outcomes in patients with schizophrenia and bipolar disorder. The COMT Val158Met polymorphism, has been extensively studied with regards to dopamine neurotransmission, because Val/Val homozygotes have pre-frontal COMT enzyme activity approximately 40% higher than Met/Met homozygotes and are considered to have concomitant lower cortical dopamine [7, 8], with Val/Met heterozygotes likely having intermediate levels. In our prior study, we observed that for patients with one or two copies of the low-activity enzyme (COMT Met allele), who are likely to have high frontal cortical dopamine, high levels of plasma proline were associated with significantly greater negative symptom severity or less symptom improvement over time. Conversely, for those with high COMT activity (Val/Val), high plasma proline levels were associated with less severe negative symptoms, or greater symptom improvement [6].
Given the high prevalence of negative symptoms in CHR and RO patients, we hypothesized that proline and dopamine metabolism also predict negative symptom outcomes in these subjects. Demonstrating that high proline has converse effects on symptoms by COMT genotype and thus activity, both prior to onset and in the early phase of psychosis, would advocate for the therapeutic potential of modulating proline levels to specifically target negative symptoms in CHR and RO patients.
Methods
Subjects
CHR and RO patients were recruited from New York State Office of Mental Health Facilities; The Center of Prevention and Evaluation at New York State Psychiatric Institute, and Rockland Psychiatric Center and its associated clinics (Orangeburg Service Center and Middletown Mental Health Clinic). The study protocol was reviewed and approved by the Western Institutional Review Board-Copernicus Group. All adult participants and parent(s) of minors provided written informed consent, and minors provided written informed assent. Eligibility for CHR was as previously described [9], and RO patients were within two-years of their first psychotic episode (see Supplementary Materials for additional details). Psychiatric diagnoses of CHR and RO patients were confirmed using the Structured Interview for Psychosis-Risk Syndromes and the Structured Clinical Interview for DSM-5 Disorders (Supplementary Table 1). In this cross-sectional study, negative symptoms were assessed via the Scale for the Assessment of Negative Symptoms (SANS, primary outcome), and the Clinical Assessment Interview for Negative Symptoms (CAINS, secondary outcome). Beck’s Depression Inventory (BDI), and the Global Functioning (GF) Social and Role Scales, were employed for assessing depression and function respectively. A fasting blood draw was obtained for measurement of plasma proline and COMT genotyping, as described [6].
Statistical analysis
Demographic and clinical characteristics were compared across genotypes and subject groups (CHR and RO), and genotype distributions tested for Hardy-Weinberg equilibrium (HWE). Linear regression was employed to model the main and interaction effects of fasting plasma proline and COMT genotype on negative symptoms, as assessed via the SANS (total and global scores). In secondary analysis the dependent variable was the CAINS. A priori exploratory outcomes were the GF:Role and GF:Social. Based upon our initial findings, additional exploratory outcomes were individual items of both the SANS and CAINS. Subject group was employed as a covariate in all analyses. Huber–White standard errors were utilized due to the presence of data outliers. To assess potential confounds, Least Absolute Shrinkage and Selection Operator (Lasso) regression for model inference was employed, with the double-selection feature for model fit of a priori defined variables (depression and gender, due to proline gender differences [6]), along with characteristics that significantly differed by genotype. Lasso regression was chosen because it is particularly useful in eliminating irrelevant features by adding penalties for large coefficients (resulting in more generalized models), and shrinking some coefficients to zero, thus efficiently performing feature selection in small datasets (https://www.stata.com/manuals/lassolassoinferenceintro.pdf) (and references therein). As a sensitivity analysis, a post-hoc stepwise selection procedure (criteria set to p < 0.1) was employed, starting with the independent variables of proline, COMT and the COMT x proline interaction, plus confounds. For all tests, the level of significance was fixed at p < 0.05, two-tailed. Benjamini-Hochberg’s corrections controlled the False Discovery Rate (FDR, <5%) for the primary, secondary, and exploratory outcomes. Analyses were conducted in STATA/BE v18 (Stata, College Station TX). STROBE reporting guidelines were employed.
Results
A total of 52 participants completed the study. Demographic and clinical characteristics are shown in Table 1. Subjects were well matched across genotype groups, although Val/Met participants were 3–4 years younger at study participation, and groups differed by their education level. Both CHR (n = 36) and RO (n = 16) patients exhibited negative symptoms at levels comparable to our previous inpatient schizophrenia population [6], and there were no differences between CHR and RO patients in the SANS total (sum of SANS items) or severity (sum of SANS global scores), nor the CAINS total score. However, CHR individuals had higher levels of depression, and a lower percentage of CHR subjects were prescribed neuroleptic medication (33% of CHR versus 94% of RO patients). Thus, subject group (CHR or RO) was employed as a covariate in regression models. Regarding biological outcomes, the sample was in HWE for Val158Met (p > 0.05), and two subjects had elevated plasma proline above standard references ranges (>383 µM), which were statistical outliers.
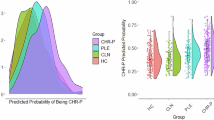
We observed a significant interaction between COMT and fasting plasma proline on SANS total (n = 52, interaction β-coefficient = 0.071, p = 0.006, partial η2 = 0.069) and global severity scores (n = 52, interaction β-coefficient = 0.028, p = 0.003, partial η2 = 0.11), Table 2. This interaction is depicted in Fig. 1. The interactions remained significant following adjustment for subject group. Potential confounds were assessed; however, the Lasso procedure did not select for retention in the final models the variables of depression (BDI score), gender, education, and age (Table 2). In a post-hoc sensitivity analysis via a stepwise selection procedure, the interaction remained significant (see Supplementary Materials Table 2), suggesting a robust finding.

The graphs depict the interaction between COMT Val158Met genotype and proline on SANS Total (A), and Global Scores (B). The data is plotted for those with the Met allele (left (A, B): Met/Met (n = 5, red) and Val/Met (n = 24, blue)), as well as the Val/Val genotype (right (A, B): n = 21, green). The n = 2 proline outliers are excluded from the figure. Lines represent the predicted values from the simple regression models, with 95% confidence intervals. In those with the Met allele, high proline levels are associated with higher scores. Conversely there is a significant negative relationship in Val/Val patients, with high proline associated with fewer and less severe negative symptoms.
In a secondary analysis, we tested for an interaction between COMT and proline on a second measure of negative symptoms, the CAINS total score; however, although supportive of our primary analysis, the significant effect (Table 2, adjusted interaction β-coefficient = 0.035, p = 0.044) did not remain after correction for multiple testing. Subsequent to these initial findings, and seeking to determine the individual symptom items that proline and COMT may influence, in exploratory analysis we investigated the five SANS and two CAINS subscales (Supplementary Table 3). While the interaction significantly predicted after Benjamini-Hochberg’s correction, the SANS items Anhedonia (covariate adjusted p = 0.002) and Alogia (covariate adjusted p = 0.007), the SANS item Attention (covariate adjusted p = 0.026), and the Motivation and Pleasure (MAP) subscale from the CAINS (covariate adjusted p = 0.04) did not retain significance after correction. Moreover, the interaction did not predict the SANS items Affective Flattening/Blunting (p = 0.208) or Avolition-Apathy (p = 0.187), nor the CAINS Expression subscale (p = 0.783).
Finally, in an a priori determined exploratory analysis, we tested whether COMT and fasting plasma proline interact to predict function (Table 2). The interaction did not predict functioning with regards to school, work, or within the home (GF-Role, p = 0.614), but interestingly, we observed a trend towards significance for the interaction predicting social and interpersonal functioning (GF-Social, covariate adjusted p = 0.055), which is consistent with the exploratory findings of an association with the Anhedonia and MAP subscales of the SANS and CAINS, and suggests that high fasting proline may have opposite effects on a patient’s ability to be interested in, or experience, social and pleasurable activities, based upon COMT genotype.
Discussion
In this study, we tested the hypothesis that proline and COMT significantly interact to predict the severity of negative symptoms in patients with recent psychosis onset, and those at clinical high-risk for developing psychosis. Given that both CHR and RO patient groups experienced substantial negative symptoms, at severities that did not differ by subject group, we tested our hypothesis using the non-stratified sample. Replicating our original finding, CHR and RO Met allele carriers demonstrated opposite associations between negative symptoms and fasting plasma proline as compared to those with the Val/Val genotype: Subjects with one or two copies of the low COMT activity enzyme exhibited markedly high levels of negative symptoms in the presence of high proline, while those with the high activity enzyme had less symptoms and lower global severity scores, when proline levels were high.
Elevated plasma proline (hyperprolinemia) has been associated with phenotypes including developmental delay and intellectual disability, autism spectrum disorder, and psychosis spectrum disorders [10]. Moreover, our finding of a detrimental effect of high proline (which would likely reflect high CNS levels [6] and references therein), in combination with the Met allele in adolescents and young adults at high-risk and those with recent psychosis onset, are supported by studies of 22q11DS patients, who carry a hemizygous deletion on chromosome 22q11 that is associated with a high-risk for developing psychosis [11]. The deletion encompasses both the COMT and PRODH genes [11] and, as such, 22q11DS patients also have significantly reduced COMT enzyme activity and routinely exhibit elevated peripheral proline due to loss of the PRODH encoded enzyme proline oxidase (POX), which metabolizes proline. Studies have confirmed that the majority of young people with 22q11DS experience one or more negative symptoms, of moderate to severe intensity [12], even in the absence of a psychosis diagnosis. Furthermore, 22q11DS patients, when carrying a single Met versus Val allele, have an increased risk of psychosis [13], as well as increased neurophysiological visual sensory deficits [14], autism spectrum symptoms [15], and amotivation [16], in the presence of high proline. Conversely, high proline levels coupled with the Val allele predicts higher mismatch negativity amplitudes, which themselves are associated with less severe negative symptoms in 22q11DS patients [17].
The findings reported here further support the opposing influences of proline and COMT enzyme activity on negative symptom severity, perhaps via prefrontal dopamine neurotransmission. In one proposed model, in those with one or two copies of the Met allele, the enhanced dopamine transmission anticipated in the prefrontal cortex as a result of excess proline, such as observed in the Prodh-null mouse [18], is exacerbated by low COMT enzyme activity, ultimately resulting in a frontal hyperdopaminergic state. It has been speculated that the similar hyperdopaminergic state observed in 22q11DS patients [19] may also contribute to psychosis-related symptoms, including negative symptoms. Conversely, in individuals with the Val/Val genotype, high prefrontal COMT activity would likely reduce dopamine, limiting D1 receptor-mediated excitation [20]. Again, as suggested by studies of the Prodh-null mouse [18], proline elevation may increase prefrontal dopamine signaling through interference with glutamatergic pathways, reducing vulnerability to an otherwise prefrontal hypodopaminergic state. In this model, and as previously speculated [6, 19], negative symptoms would be induced in conditions of both hyper- and hypo-dopaminergia, reflecting the inverted U-shape curve proposed for cognitive dysfunction in psychosis [20].
Although a limitation of this study is the small sample size, this investigation of CHR and RO patients builds positively from our previous study of schizophrenia and bipolar disorder inpatients [6], while also seeking to reduce the potential impact of confounding factors such as extended psychosis illness duration and long-term antipsychotic use. Moreover, this study of early psychosis and those at-risk may inform future strategies for early intervention and treatment. This is particularly important given that there are currently no effective treatments for negative symptoms. On the other hand, there are medications that are known to modulate proline levels [6], which may hold promise in pharmacological approaches to intervene early and target this symptom domain.
Data availability
Data will be available from the NIMH Data Archives. https://nda.nih.gov/
References
Vesterager L, Christensen TO, Olsen BB, Krarup G, Melau M, Forchhammer HB. et al. Cognitive and clinical predictors of functional capacity in patients with first episode schizophrenia. Schizophr Res. 2012;141:251–6.
Meyer EC, Carrión RE, Cornblatt BA, Addington J, Cadenhead KS, Cannon TD, et al. The relationship of neurocognition and negative symptoms to social and role functioning over time in individuals at clinical high risk in the first phase of the North American Prodrome Longitudinal Study. Schizophr Bull. 2014;40:1452–61.
Nelson B, Yuen HP, Wood SJ, Lin A, Spiliotacopoulos D, Bruxner A, et al. Long-term follow-up of a group at ultra high risk (“prodromal”) for psychosis: the PACE 400 study. JAMA Psychiatry. 2013;70:793–802.
Marshall M, Lewis S, Lockwood A, Drake R, Jones P, Croudace T. Association between duration of untreated psychosis and outcome in cohorts of first-episode patients: a systematic review. Arch Gen Psychiatry. 2005;62:975–83.
Mei C, van der Gaag M, Nelson B, Smit F, Yuen HP, Berger M, et al. Preventive interventions for individuals at ultra high risk for psychosis: An updated and extended meta-analysis. Clin Psychol Rev. 2021;86:102005.
Clelland CL, Drouet V, Rilett K, Smeed J, Nadrich RH, Rajparia A, et al. Evidence that COMT genotype and proline interact on negative symptom outcomes in schizophrenia and bipolar disorder. Transl Psychiatry. 2016;6:e891.
Lachman HM, Papolos DF, Saito T, Yu YM, Szumlanski CL, Weinshilboum RM. Human catechol-O-methyltransferase pharmacogenetics: description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics. 1996;6:243–50.
Chen J, Lipska BK, Halim N, Ma QD, Matsumoto M, Melhem S, et al. Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): effects on mRNA, protein, and enzyme activity in postmortem human brain. Am J Hum Genet. 2004;75:807–21.
Brucato G, Masucci MD, Arndt LY, Ben-David S, Colibazzi T, Corcoran CM, et al. Baseline demographics, clinical features and predictors of conversion among 200 individuals in a longitudinal prospective psychosis-risk cohort. Psychol Med. 2017;47:1923–35.
Namavar Y, Duineveld DJ, Both GIA, Fiksinski AM, Vorstman JAS, Verhoeven-Duif NM, et al. Psychiatric phenotypes associated with hyperprolinemia: a systematic review. Am J Med Genet B Neuropsychiatr Genet. 2021;186:289–317.
Drew LJ, Crabtree GW, Markx S, Stark KL, Chaverneff F, Xu B, et al. The 22q11.2 microdeletion: fifteen years of insights into the genetic and neural complexity of psychiatric disorders. Int J Dev Neurosci. 2011;29:259–81.
Stoddard J, Niendam T, Hendren R, Carter C, Simon TJ. Attenuated positive symptoms of psychosis in adolescents with chromosome 22q11.2 deletion syndrome. Schizophr Res. 2010;118:118–21.
Raux G, Bumsel E, Hecketsweiler B, van Amelsvoort T, Zinkstok J, Manouvrier-Hanu S, et al. Involvement of hyperprolinemia in cognitive and psychiatric features of the 22q11 deletion syndrome. Hum Mol Genet. 2007;16:83–91.
Magnée MJ, Lamme VA, de Sain-van der Velden MG, Vorstman JA, Kemner C. Proline and COMT status affect visual connectivity in children with 22q11.2 deletion syndrome. PLoS ONE. 2011;6:e25882.
Hidding E, Swaab H, de Sonneville LM, van Engeland H, Vorstman JA. The role of COMT and plasma proline in the variable penetrance of autistic spectrum symptoms in 22q11.2 Deletion Syndrome. Clin Genet. 2016;90:420–7.
Schneider M, Van der Linden M, Glaser B, Rizzi E, Dahoun SP, Hinard C, et al. Preliminary structure and predictive value of attenuated negative symptoms in 22q11.2 deletion syndrome. Psychiatry Res. 2012;196:277–84.
Zarchi O, Carmel M, Avni C, Attias J, Frisch A, Michaelovsky E, et al. Schizophrenia-like neurophysiological abnormalities in 22q11.2 deletion syndrome and their association to COMT and PRODH genotypes. J Psychiatr Res. 2013;47:1623–9.
Paterlini M, Zakharenko SS, Lai WS, Qin J, Zhang H, Mukai J, et al. Transcriptional and behavioral interaction between 22q11.2 orthologs modulates schizophrenia-related phenotypes in mice. Nat Neurosci. 2005;8:1586–94.
van Duin EDA, Ceccarini J, Booij J, Kasanova Z, Vingerhoets C, van Huijstee J, et al. Lower [(18)F]fallypride binding to dopamine D(2/3) receptors in frontal brain areas in adults with 22q11.2 deletion syndrome: a positron emission tomography study. Psychol Med. 2020;50:799–807.
Tunbridge EM, Harrison PJ, Weinberger DR. Catechol-o-methyltransferase, cognition, and psychosis: Val158Met and beyond. Biol Psychiatry. 2006;60:141–51.
Acknowledgements
We would like to thank the Study Coordinators and Raters involved in this study. Most importantly, we would like to thank the subjects who participated in this study.
Author information
Authors and Affiliations
Contributions
Concept and design of the study: CLC and JDC. JDC, HH, KR, AT, RRG, and CLC contributed to the acquisition, analysis, and interpretation of data. The primary statistical analysis was performed by CLC, with additional statistical analysis performed by JA. Drafting of the initial manuscript was undertaken by CLC and JDC. All authors contributed to the critical revision and review of the final manuscript.
Corresponding author
Ethics declarations
Competing interests
This work was supported by grant R21MH116515 from the National Institute of Mental Health (CLC). The views expressed in this article are solely those of the authors and do not reflect the endorsement or the official policy or position of the National Institute of Mental Health. CLC and JDC are inventors on a patent that describes use of proline-modulators as a treatment for schizophrenia. The patent is owned by their respective institutions, and CLC and JDC may benefit financially in the future if these patents are licensed. R. Girgis acknowledges recent expert consulting for Noble Insights, Signant Health, Guidepoint, Clearview Healthcare Partners, Health Monitor, and IMS Consulting and Expert Services for which he received compensation, as well as royalties from books on mental health published by Wipf and Stock and Routledge/Taylor and Francis. He also provides expert legal consulting for individual law firms. No other conflicts are reported.
Ethics approval and consent to participate
All methods were performed in accordance with all guidelines and regulations of Columbia University Irving Medical Center, The Nathan Kline Institute, New York State Psychiatric Institute, and New York State Office of Mental Health. The study protocol was reviewed and approved by the Western Institutional Review Board-Copernicus Group (protocol number: 20182236). All adult participants and parent(s) of minors provided written informed consent, and minors provided written informed assent.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Clelland, J.D., Hesson, H., Ramiah, K. et al. The relationship between COMT, proline, and negative symptoms in clinical high risk and recent psychosis onset. Transl Psychiatry 14, 409 (2024). https://doi.org/10.1038/s41398-024-03099-0
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41398-024-03099-0
