
Abstract
Auditory verbal hallucinations [AVH] are one of the most common psychopathological symptoms in psychosis and schizophrenia [SZ]. While various studies demonstrate the neuronal features of AVH in specific regions and networks including changes in structural and functional connectivity, the relation of their neuronal topography to the organization or structure of the subjects’ experience, i.e. the phenomenology of their ‘mental topography’, remains yet unclear. Addressing this gap in our knowledge, we review recent findings to formulate two neurophenomenological hypotheses of AVH (which extend our earlier Resting state hypothesis). First, we hypothesize that structural and functional dysconnectivity in AVH with desynchronization and temporal fragmentation between sensory auditory, interoceptive somatic and cognitive linguistic regions [e.g., auditory cortex, insula and Broca’s area] relate to the experience of analogous temporal fragmentation on the mental level, that is, among the contents in perception and thought - this is manifest in ‘hyperreflexivity’, the attribution of abnormal salience and attention to single isolated objects or elements in perception. Secondly, we propose that the topographic distinction and gradient of unimodal and transmodal regions are reduced in individuals with AVH. We hypothesize that such ‘inner-outer neuronal topographic dedifferentiation and compression’ is manifest in the experience of overlaps and confusions between the subject’s outer interpersonal social and inner intrapersonal spaces – analogous to the neuronal level, this reflects an ‘inner-outer mental topographic dedifferentiation and compression’. Together, we propose two testable neurophenomenological hypotheses of AVH showing shared structural-topographic changes in both brain and experience as their “common currency”. This directly connects changes in the brain’s neuronal topographic organization with the structure of experience, that is, the mental topographic organization of the subjects’ experience of AVH in SZ.
Similar content being viewed by others

Introduction
Hallucination are a heterogeneous group of phenomena with a wide array of different presentations canonically considered as a fallacious perception of a conscious sensory experience in absence of a corresponding external stimulation. Such phenomena can be defined as subjectively experienced sensations in the absence of an appropriate stimulus that are perceived by the individual as real. They may be of organic origin, drug induced [by pharmacological agents such as psilocybin, LSD and others], occur in healthy subjects, and can be associated with a mental disorder [1] such as schizophrenia spectrum disorder [2] or bipolar disorder [3]. We therefore suppose a dimensional view of AVH in our paper, as they can occur across different causal disorders in a transdiagnostic way.
Although often concomitant in presentation, AVH are distinct from illusions in that the latter are considered to be a misperception of an external stimulus rather than a subjective conscious perception in the complete absence of any external stimulation. [1] Distortion of the sense of reality, as encountered in positive symptoms such as delusions and hallucinations, is a hallmark feature of the insurgence of psychosis. Importantly, the phenomenological variables of the AVH themselves such as content, context, agency, loudness, complexity, location and persistence might carry prognostic value [4] and could act as a marker of disease severity and predictor of disease progression [5]. This makes it even more urgent to develop specific biomarkers for auditory verbal hallucinations [AVH] which directly connect subjective experience [i.e. phenomenology] of AVH to their underlying neuronal changes [6].
The goal of our article is to bridge the gap between neuronal findings and phenomenological observations of AVH by formulating two testable neurophenomenological hypotheses. We directly connect both neuronal findings and phenomenological insights about the experiential nature of AVH in schizophrenia which, more generally, allows us to bridge the gap of brain and experience. For that purpose, we provide a narrative review of recent findings concerning neurophysiological alterations in third-person perspective to connect them to key features described by phenomenological accounts of AVH in first-person perspective. Such connection of first- and third-person perspectives is based on a novel framework, namely Spatiotemporal psychopathology [7, 8].
What is Spatiotemporal Psychopathology? Roughly, psychopathological symptoms like AVH are here primarily conceived as spatial-topographic and temporal-dynamic changes that are shared by, and manifest in, both neural and mental features as their “common currency” [9,10,11]. For instance, we will demonstrate in our second hypothesis how the distinction (or lack therefore in AVH) between uni- and transmodal regions with their neuronal distinction of outer perception from inner cognition relate to a corresponding mental or experiential distinction (or lack therefore in AVH) of outer inter-personal social space and inner intra-personal space – the inner-outer distinction (or its confusion in AVH) is thus shared by both brain and experience as their “common currency” [10, 11]. This can, for instance, be tested by applying analogous topographic measures of inner-outer relationship on both neuronal (gradient of uni-transmodal regions during both external perception and internal cognition) and mental (experience of outer environment and inner thoughts) levels in analogous ways (see below for details).
Focusing on the spatial side, e.g., topography in this paper (while leaving out the temporal and thus dynamic side), we will speak of both “neuronal and mental topography”: “neuronal topography” refers to the organization of the brain’s different regions and networks including their relations among each other. While the concept of “mental topography” designates the structure or organization of the subjective experience of the relationship among distinct spatial ‘locations’ like the experiential distinction and relationship of inner intra-subjective and outer inter-subjective social space.
Neurophenomenological Hypothesis of AVH I: Perception of fragmented contents relates to the brain’s structural and functional dysconnectivity among sensory, somatic and cognitive regions
Phenomenology of AVH – Fragmentation and hyperreflexivity of the contents in perception
Perception is a dynamic process with a temporal contour that offers an incomplete experience of a phenomena/percept/object in the first person perspective [12]. This takes place within a larger context, that is, a perceptual field that consists of a series of percepts interconnected by spatiotemporal relations, as well as relations of meaning and significance. In the early stages, subjects suffering from schizophrenia spectrum disorder typically experience auditory hallucinations consisting of single words or brief sentences [12]. This suggests fragmentation of contents in perception in AVH.
AVH are often initially perceived as the patients’ own thoughts with the peculiarity of being recognized as alien and non-self in their origin [13], [‘thoughts that I’m forced to think’]. Importantly, at the same time, they are experienced as being unrelated to the external environment due to their acontextual nature [12]. In later stages, the contents of AVH typically gain complexity: the perceptual contents become more ample and articulated and, ultimately, they may result in classic Schneiderian first rank phenomena [12] such as perceiving a dialogue between “voices” discussing and commenting patients‘ actions or behavior. Importantly, these phenomena might be perceived without any appreciable sensorial [“silent voices”] and/or experiential quality, that is, their “what it is like” is reduced if not absent [12]. Moreover, is not uncommon that hallucinatory phenomena exhibit continuity with other non-hallucinatory alterations of thoughts and perception [14].
A phenomenological variable commonly associated with AVH is hyperreflexivity; this term refers to an abnormality of thought processing with the attribution of increased salience to discrete elements and details in one’s perception [and thoughts] which usually are not aware and reflected upon [15]. Therefore, hyperreflexivity leads the subject to single out elements from a situational context in a non-volitional, non-intellectual and non-reflective way: the subject becomes suddenly aware of a single isolated fragment of the whole perception with a focus on an either its internal [thoughts, feelings, sensations, proprioceptive inputs] or external (events, objects, persons, etc.) elements. [16]
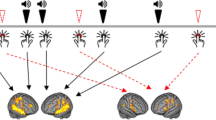
Taken together, the anomalies in the perception of AVH are characterized by fragmentation of the contents into single isolated elements. These fragmented contents are dissociated from experiential qualities [their “what is like”], and to which an abnormal salience is attributed, a process phenomenologically described as hyperreflexivity [See Fig. 1].

The left figure shows preserved connectivity, synchronization and temporal integration in both perception/experience and brain. While the right figure shows the fragmentation of perception in experience with the impairment in salience attribution/hyperreflexivity (upper part) and how that relates to the fragmentation of the auditory cortex’ functional connectivity with the insula and language areas (lower part).
Neuronal changes in AVH – Structural and functional dysconnectivity of the auditory cortex with sensory, somatic and cognitive regions
Structural connectivity
AVH in the schizophrenic patient are linked to anomalies in gray matter volume [GMV] localized in various brain areas. Analyzing whole-brain gray matter volume in patients experiencing auditory hallucinations, Zaira et al. [17] found hallucinatory phenomena to be correlated with reduction in GMV in a left lateralized cluster of brain areas including left anterior insula and left inferior frontal gyrus. The authors therefore conclude that structural impairment in areas involved in generating and perceiving speech, as well as in auditory signal processing, might be closely associated with AVH. This is further supported by the positive association of the loss of GMV in left insula and superior temporal gyrus [STG] with the severity of hallucination.
Sone et al. [18] analyzed a 204 patients pool comprising 58 schizophrenic patients suffering from AVH, 29 schizophrenic patients without a history of AVH, and 117 healthy controls. They found that the subjects with AVH showed significant reduction in gray matter volume in the left caudal middle frontal gyrus and precentral gyrus when compared to the non-AVH group. Moreover, they observed a smaller surface area in the left insula and reduced subcortical volume in the bilateral hippocampus when compared to the control group.
In line with these results, Ford et al. [19] observed a left-lateralized deficit in processing external auditory stimuli with decreased activation of primary auditory cortex during probe tones. Other studies suggest abnormal structural connectivity of the auditory cortex with other cortical or sub-cortical regions [20], reduction in GMV [21] reduction in cortical thickness [22], and increased activity of the auditory cortex [23, 24]. However, another study reports absent activation in auditory cortex while language and verbal short-term memory brain areas are activated during AVH [25].
Functional connectivity
In addition to structural connectivity, several resting state fMRI studies also analyzed alterations of functional connectivity [FC] in AVH. Recent studies [26, 27] found widespread changes in FC characterized by reduced connectivity between dorsal and ventral anterior insula and default mode network [DMN] as well as between salience network and central executive network. This suggests decreased functional specialization with reduced functional segregation between anterior and posterior insular cortices.
Xue et al. [28] examined alterations in dynamic functional connectivity [dFC] in 107 drug-naïve schizophrenia patients with AVH during resting state fMRI [rs-fMRI]. They observed an increase in dFC from left auditory association cortex to the right middle temporal gyrus and right middle occipital gyrus. Moreover, they show decreased dFC from left Heschl gyrus to various regions including the left superior occipital gyrus, left cuneus gyrus, and left precuneus gyrus; there was also decreased dFC from right Heschl gyrus to the posterior cingulate gyrus, as well as decreased dFC from left medial geniculate nucleus to the bilateral calcarine gyrus, bilateral cuneus gyrus, bilateral lingual gyrus. The authors conclude that AVH phenomena are closely related with dFC alterations particularly involving the occipital lobe, default mode network, and middle temporal lobe.
Consistent with these results, Panikratova et al. [29] demonstrated decreased FC between the left inferior frontal gyrus and the bilateral auditory association cortex in schizophrenia patients with history of AVH; this further strengthens the association between impairment in language processing areas and the occurrence of AVH. This is further supported by other studies also showing concurrent changes in both auditory cortex and language processing areas during AVH in SZ patients [22, 24, 30].
In another study comparing whole brain functional connectivity in SZ patients with and without AVH (and healthy controls), Zhang et al. [31] demonstrated abnormalities in functional connectivity localized among the frontal lobe, temporal regions and parietal regions. In particular compared to non AVH group, the AVHs group showed significantly enhanced functional connectivity among Frontal Gyrus, Inferior Parietal Lobule, and Hippocampus.
Alterations of connectivity between auditory regions, language processing areas and memory rgeions in SZ AVH are also reported by Gao et al. [32] Using Granger causality analysis, they showed significantly increased directionality in FC from Broca’s area to superior temporal gyrus which was positively associated with the severity of hallucinations. While the direction of FC, e.g., Granger causality, was decreased from the right homolog of Wernicke’s area towards the right homolog of Broca’s area, as well as from the right STG to the right hippocampus in the subjects suffering from AVH when compared with the non-AVH group.
Together, there are rather heterogenous results in the structural and functional connectivity studies focusing on AVH. We can nevertheless see a certain pattern of changes which concerns the auditory cortex and its structural and functional connectivity which is altered in relation various regions like; (i) sensory regions involved in the processing of exteroceptive inputs; (ii) somatic regions like the insula that are involved in processing interoceptive inputs; and (iii) higher-order cognitive regions like default mode network and prefrontal cortex that are involved in higher-order cognition including language. Put together, there is structural and functional dysconnectivity of the auditory cortex with sensory-exteroceptive, interoceptive and cognitive regions.
Neurophenomenological hypothesis of AVH I – Dysconnectivity, desynchronization and temporal fragmentation in both brain and experience/perception
What do the findings in both structural and functional connectivity among the various regions mean in phenomenological terms? To address this question, we first need to go back to the physiology underlying functional connectivity. Functional connectivity is calculated by correlating the timeseries’ of two or several regions activities which, physiologically, can be traced to their neuronal synchronization: the different regions’ activity timeseries synchronize with each other yielding high functional connectivity with high temporal integration among them. While they no longer synchronize with each other over time when their functional connectivity is low reflecting predominant temporal segregation over temporal integration.
Accordingly, neuronal synchronization and temporal integration are key features for establishing functional connectivity among auditory, somatic, sensory and cognitive regions. Correspondingly so in perception and cognition: the auditory contents processed in auditory cortex can then be synchronized and temporally integrated with contents that are processed in other regions including sensory, somatic and cognitive. One subsequently experiences the contents in one’s perception as integrated composites of auditory, somatic, sensory and cognitive elements. For instance, we experience the melody of a music piece (auditory) in conjunction with somatic feelings in our body like goosebumps, visual imagery and cognitive association of for instance memories of past events (like the encounter of a friend who played the same music).
Such integrated composite in one’s perception [and thoughts] is well compatible with the phenomenological observation that contents in perception and thought are neither experienced as purely sensory, somatic, or cognitive, but rather as an unified and integrated construct. In fact, we experience transient auditory contents in the foreground relative to our body’s somatic state and our ongoing thoughts [33]. This background-foreground relationship defines what is known as a “perceptual field,” that is characterized by the spatiotemporal integrations of various sensory, somatic, and cognitive components within a unified whole and unity in our perception [12].
We now assume that such background-foreground relations are disrupted in AVH which, as we assume, can be traced to structural/functional dysconnectivity of the auditory cortex with the sensory, [e.g. auditory], somatic [e.g. insula], and cognitive [e.g. DMN and language processing] regions. Let us explicate that in more detail.
The neuronal findings reveal structural and functional dysconnectivity, desynchronization and temporal fragmentation between the auditory cortex and sensory, somatic, and cognitive regions. As a result, the various contents—or more accurately, the individual elements—fail to be integrated into a cohesive, unified perceptual composite. Instead, the subject now perceives the single elements themselves isolated and independent of each other. This aligns closely with the phenomenology of AVH: percepts appear to be no longer naturally interwoven within the composite structure of the perceptual field but rather manifest with an extreme phenomenological poverty [12], devoid of any foreground-background contextualization and lacking temporal contour.
We consequently hypothesize that the structural/functional dysconnectivity of the auditory cortex with sensory, somatic and cognitive regions relates to the aberrant perception of isolated, individual elements without their structural integration within a unified experience as it is typical for AVH. Single elements and their details, now devoid of contextualization, become subsequently the main focus of the subject’s awareness and attention – this results in what phenomenologically is described as ‘hyperreflexivity’ (see above). Accordingly, we suppose corresponding processes on both neural and mental levels in AVH, that is, dysconnectivity, desynchronization and temporal fragmentation holding among different brain regions as well as among different perceptual and cognitive elements/contents.
Neurophenomenological hypothesis of AVH II – Disruption of local-to-global input integration leads to the disruption of the structure of experience (background-foreground, perceptual field, qualia)
Why and how do dysconnectivity, desynchronization, and temporal fragmentation in the neural activity of different regions transform into corresponding disruptions in the integration of contents in the experience of perception and cognition? This raises the question for neuronal mechanisms constituting the experience of a unified composite and unity of our perception and thoughts occurs. We suppose a key role for the neural integration of local-regional activity changes during their input processing within the brain’s global activity: the single input is processed within the brain’s global larger, more expanded, temporo-spatial context/framework which relates and integrates the various inputs within one single unity or composite, the brain’s global background activity. Importantly, we assume that such local-to-global integration of single inputs and elements into a unified whole is key for their association with subjective experiential qualities, their “what it is like” [34]. Let us explicate that.
The local-to-global integration among different regions is supposedly mediated by their structural-functional connectivity and neuronal synchronization: this enables the temporal integration of the auditory cortical inputs with the inputs of the somatic, sensory and cognitive regions. We now assume that such local-to-global integration is manifest in the experience of what phenomenologically is described as ‘background-foreground relation’ and ‘perceptual field’ : the single specific element or content in the foreground is perceived within the context of multiple other elements serving as background, resulting in the constitution of a perceptual field.
Given the findings of dysconnectivity, desynchronization and temporal fragmentation on the neuronal level of the brain, we now postulate that the local-to-global integration of different inputs is disrupted in AVH. The local inputs from for instance the auditory cortex are no longer connected with other inputs from sensory, somatic and cognitive regions – the former are not integrated within a unitary composite or whole. The lack of such unitary composite or whole makes impossible the constitution of both background-foreground relation and perceptual field in the subject’s experience. That leads consecutively to the loss of the experiential quality, the qualia or what it is like: the perceived contents are not only single isolated elements [rather than complete perceptual contents] but, at the same time, are also dissociated from their respective context/background – this makes impossible their association with an experiential quality [their “what is like”], a core feature of the experience of auditory contents in AVH.
Together, our first neurophenomenological hypothesis assumes that the dysconnectivity, desynchronization and temporal fragmentation of the auditory cortex‘ input processing with other inputs in sensory, somatic and cognitive regions leads to analogous changes on the mental level. As on the neuronal level, there is corresponding dysconnectivity, desynchronization and temporal fragmentation among the contents in perception and thoughts. Phenomenologically speaking, this disrupts both background-foreground relation and perceptual field with the subsequent dissociation between the perceptions and their experiential qualities [their ‘what it is like’]. In short, dysconnectivity, desynchronization and temporal fragmentation are shared by both neural and mental levels as their “common currency” [10] in AVH.
Neurophenomenological hypothesis of AVH II: Experience of hallucinatory space relates to dedifferentiation and compression of the brain’s unimodal-transmodal gradients
Phenomenology of AVH – Convergence with confusion of intrapersonal and interpersonal spaces within the hallucinatory space
AVH are typically perceived by the patient as embedded in a ‘hallucinatory space’ [35] that is, an intrasubjective space arising from a profound disturbance of most basic, immediate and pre-reflective relation to space, also termed primordial presence, originating from the pathological disruption of patients‘ perception of self [35]. The notion of space in this setting refers the subjectively and pre-reflectively experienced space within which the internal or external perceptual contents are embedded [36]. Such phenomenological notion relates to the concepts of interpersonal and intrapersonal space. The interpersonal space denotes a common space shared by different subjects within the external environment – this is manifest in the subjective perception of an interpersonal space by the subjects themselves. While the concept of intrapersonal space, refers to a subjectively experienced space localized and perceived within the individual itself where one experiences one’s own thoughts and emotions.
Usually, we can clearly distinguish between intrapersonal and interpersonal spaces in experience as we do not, for instance, confuse our inner thoughts with our outer perceptions. Such distinction, however, seems to be rather blurred during AVH, as manifest in the experience of the hallucinatory space. The concept of the hallucinatory space must be distinguished from abnormal space experience in mental imagery and illusions. Even if experiencing inner-outer connection in mental imagery or illusions, these subjects are still able to distinguish the space of the inner reality of their own self from the space of the outer environmental reality.
In contrast, this is no longer possible in the experience of a hallucinatory space: something that occurs in the inner self is associated with and located in the outer environment, e.g., the AVH. Similar so in the experience of passivity phenomena where for instance subjects experience that their own inner thoughts are manipulated by others as external environmental forces. We here leave it open whether and how such hallucinatory space in AVH with the typical inner-outer confusion also relates to delusions: this is even more pressing given that both delusions and AVH often co-occur together.
The hallucinatory space is often experienced by the patient to be deeply intimate and personal [living in the head, living in a glass bubble]. The AVH are consequently perceived as private and inaccessible to others [36] for which reason they are only rarely mistaken as ‘real’ voices coming from the surrounding intersubjective environment [37]. We suppose that the hallucinatory space of AVH arises from the progressive overlap between intra- and interpersonal spaces in one’s experience, a ‘phenomenological dedifferentiation’. Both inter- and intrasubjective spaces seem to converge and fuse within the hallucinatory space resulting in the experience of what has been described as “double ontological orientation” [12]: the hallucinatory space is oriented inwards towards the inner self and, at the same time, oriented outwards towards the outer world.
The experience of overlay, fusion and integration of intra- and interpersonal spaces carries major implications as the patient simultaneously experiences two different but subjectively equally significant “locations and origins” of perceptual contents: one is coming from a shared outer social space structured on laws of nature and the principle of non-contradiction [38]. While the other is arising from a more private inner sphere where the cardinal principles of non-contradiction and spatiotemporal continuity are somehow fluid or even completely absent [39]. This leads to confusion and distress, especially when the two spaces become completely indistinguishable during the acute exacerbation of psychosis [38, 39] with the typical confusion of inner and outer realities [40] [See Fig. 2].

The left figure shows the distinction of intra-personal and peripersonal spaces in the subject’s experience (upper) and the corresponding distinction of uni- and transmodal regions in their topographic gradient (lower part). While the right figure illustrates the overlay, fusion and integration of personal [orange] and peripersonal [blue] spaces with the subsequent experience of a hallucinatory [purple] space in AVH (upper part) which relates to decreased distinction of uni- and transmodal regions in their topographic gradient (lower part).
Neuronal changes in AVH – Topographic dedifferentiation and compression of unimodal-transmodal gradients
The global topography of the entire brain is represented by gradients that span across lower-order unimodal and higher-order transmodal regions describing their relative relations and connections. These uni-transmodal gradients of brain’s spontaneous activity [e.g. resting state] are grouped along a progressive continuum of cytoarchitecture, myeloarchitecture, chemo-architecture, excitation-inhibition balances, timescales, and activation patterns as well as functional and structural connectivity patterns. Taken in this sense, gradients provide an organizing principle for hierarchical interactions between brain areas [41] whose global relationships distribute along an information processing continuum across different functional systems [41].
Among the large scale gradients identified, two account for about half the variance in functional connectivity along the cortex. [42] The first gradient, also termed sensory-fugal gradient, spans the distance between unimodal specialized sensorimotor areas, involved in the perception of external stimuli, and higher-order transmodal association areas, that are related with a wide array of functions including processing of information, cognition, and self-referential thinking [41, 42]. The second gradient is anchored within the sensorimotor system itself by spanning the distance between visual and auditory and sensorimotors cortices [43].
Alterations of these gradients are reported in schizophrenia patients suffering from AVH. Using resting state fMRI, Dong et al. [44] observed reduced differentiation with compression of unimodal-transmodal gradients in AVH and schizophrenia. Such compression of the unimodal to transmodal gradient might impair effective integration of information through diminished differentiation between unimodal sensory systems, e.g., stimulus perception, and transmodal areas that support complex cognitive inferences. [44] Similarly, Wengler et al. [45] report that in schizophrenia patients the severity of AVH positively correlates with the degree of the hierarchical gradient’s compression within the auditory systems’ intrinsic neural timescales. Interestingly, they also observe converse relationship of the same neuronal features with the severity of delusions; this suggests partial overlap of AVH and delusions which may potentially explain their co-occurrence.
Further, He et al. [43] report alterations of a unimodal to transmodal hierarchy of cortical organization in first episode psychosis which affected the hierarchical distance between the sensorimotor and the default mode networks. They observe a global gradient compression in first episode patients, suggesting decreased functional distance between the two endpoints of the uni-transmodal cortical hierarchy. This suggests a disturbance in the balance between sensorimotor and higher-order association cortices in first episode psychosis. Specifically, the compression suggests reduced differentiation of unimodal and transmodal regions which affects their relative input processing, indicative of a decrease in their functional specialization.
These fMRI resting state findings entail that the default-mode network [DMN], usually at the apex of the hierarchy, is in a different topographic position relative to unimodal regions in the settings of AVH as it is now compressed with, and thus less differentiated from, the other networks. This is, for instance, manifest in decreased antagonistic behavior between the DMN and the language network [46] ;this is supported by reduced periods of time spent in, and reduced probability to switch to, a state of anticorrelation between the two networks in AVH + SZ patients compared to SZ patients without AVH. Decreased network-antagonism implies less differentiation between language network and DMN, and subsequently also dedifferentiation of their respectively associated functions, e.g., language processing and self-referential processing; this could engender confusion about the source and nature of the information processed. [46, 47]
Taken together, these findings clearly demonstrate topographic dedifferentiation between unimodal and transmodal regions, e.g., between sensory regions, language areas and the DMN. Decreased network-antagonism could imply increased overlap between these systems. For instance, language processing (language network) is less distinct from self-referential processing (DMN) – neuronal dedifferentiation of the networks may thus lead to subsequent dedifferentiation of their information processing.
Neurophenomenological hypothesis of AVH I – From the brain’s inner-outer neuronal topography to the inner-outer mental topography of experience
What exactly does the dedifferentiation and compression of uni-transmodal regions mean? Unimodal regions are specific brain areas dedicated to processing information from a single sensory or motor modality; sensory regions are specialized for processing specific types of external inputs from the outer environment while motor regions are involved in generating corresponding outputs in response to these inputs. In contrast, transmodal regions mainly process internal inputs related to the own self, memory, emotion and other higher-order functions originating from within the inner environment of the person’s self. In the healthy individual both unimodal and transmodal regions are clearly distinct as they occupy a precise hierarchical position along the primary sensoryfugal gradient.
Such hierarchical differentiation and functional segregation entail that their respective processing and therefore the exact origin of inputs —spanning both the inner self and outer environment—can be precisely distinguished at the neural level. We assume that this neural distinction is reflected in a corresponding differentiation at mental level as we can clearly distinguish the experience of our internal thoughts originating from the inner environment from our external perceptions arising from the outer environment. The neuronal findings clearly demonstrate that the brain regions distributed along the uni-transmodal gradient are no longer as differentiated in AVH – this is manifest in gradient compression and dedifferentiation with their reduced functional segregation. Reduced functional segregation among uni- and transmodal regions, in turn, suggests an impairment in their respective input processing: there may be reduced discrimination between inputs of distinct origins like internally and externally arising inputs from within inner brain/body and outer environment.
Following Dong et al. [44], reduced network differentiation, altered network dynamics and ineffective functional specialization could ultimately lead to confusion between outer environment and inner abstract cognitive processing [44]. That, in turn, could facilitate the intrusion of internally generated inputs, arising from the fragmented stream of consciousness, into the external perceptual field. Similarly, Weber et al. [47] suggests that “weak segregation between different functional networks could be related to impairments in differentiating between internal versus external contents” [47].
Together, these observations suggest that abnormal neuronal dedifferentiation and compression of the brain’s uni-transmodal topography lead to the confusion of inner and outer environments in subjective experience. Due to the pathological functional proximity between the two extremes (uni- and transmodal) of the hierarchy, an external input arising from the outer environment, received and processed by unimodal sensory regions is now also processed by transmodal regions; that functional overlap might entail that such input could be mistakenly designated as originating from the inner body and environment rather than being recognized as arising from the outer environment.
Conversely, an internally generated input from within the brain [and/or body itself] may no longer be processed primarily in transmodal regions, but could also involve unimodal sensory processing, such as that in the auditory cortex. As a result, this input may be perceived as originating from the outer rather than inner (brain/body) environment, and consequently experienced as non-self (rather than as self-specific), as we suppose might be the case in auditory verbal hallucinations.
Accordingly, the abnormal neuronal dedifferentiation and compression of the brain’s inner-outer/uni-transmodal topography may structure the experience in a corresponding way [9,10,11]. This is manifest in what we refer to as abnormal mental dedifferentiation and compression of the inner-outer topography of the subject’s experience with an overlap their inner thoughts and outer perceptions. Put briefly, the changes in the brain’s inner-outer neuronal topography relate to corresponding abnormalities in the structure of experience, that is, its inner-outer mental topography. How does that relate to the experience of a hallucinatory space in AVH? That shall be the focus in the next section.
Neurophenomenological hypothesis of AVH II – From the brain’s inner-outer neuronal topography to the experience of a hallucinatory space
How does the brain’s topographic unimodal-transmodal dedifferentiation and compression relate to the experience of the hallucinatory space in AVH? To better understand this, we need to distinguish what is described as peripersonal and personal spaces in psychology.
The peripersonal space is defined as the space surrounding one’s body where the subject can interact with external entities including other individuals and inanimate objects. [48] Both unimodal sensory and motor regions are closely related to the peripersonal space that demarcates and relates the subject to other subjects in the outer social world [48]; this is possible by merging interoceptive inputs from within the body with the exteroceptive environmental inputs/cues coming from the common shared intersubjective social space. [49,50,51] Conversely, the personal space, the physical distance a subject maintains from other subjects to avoid discomfort [52], is more anchored within the subject’s intimate sphere and thus restricted to within the subject itself, e.g., intra-personal. The findings suggest that the personal space relates more to the transmodal regions and their cognitive processing [48].
We now tentatively suppose that the distinction of peripersonal and personal spaces relates to the topographic gradient of uni- and transmodal regions. As described above, the peripersonal space is strongly mediated by unimodal regions including both sensory and motor cortices. While the personal space is more shaped by the transmodal regions with their integration of sensory inputs with cognitive processing. We now suppose that the distinction of uni- and transmodal regions by their gradient also allows for the corresponding distinction of peripersonal and personal space: the steeper the uni-transmodal gradient with their stronger differentiation, the more clearly the boundaries of peripersonal and personal spaces can be demarcated in the subject’s perception and experience.
The findings in AVH suggest the opposite, though. There is a lower gradient with topographic dedifferentiation and compression among uni- and transmodal regions. This should thus lead to decreased distinction with overlaps and confusions in the subject’s experience of their peripersonal and personal spaces. In fact, it has been observed that SZ patients experience alterations in the perception of both personal and peripersonal spaces. Different studies demonstrated that subjects perceive their peripersonal space as narrower, less demarcated, and more prone to intrusions by elements in the environment [48, 53, 54]. Concomitantly, other studies showed opposite changes in personal and peripersonal space: when compared to healthy controls, the SZ patients exhibit a narrowing of their peripersonal space while they perceive their personal space as more extended [52, 55].
These observations support the hypothesis of an overlap between the boundaries of peripersonal and personal spaces in subjects’ perception and experience – this suggests compression and dedifferentiation between interpersonal and intrapersonal spaces. In the most extreme case, subjects may perceive and experience no longer any difference between their inter- and intra-personal spaces. In that case, inputs arising in the intra-personal space are experienced in association with the inter-personal space – this is the moment where both spaces result in the experience of what phenomenologically is described as ‘hallucinatory space’, the confusion of inner-outer spaces in experience [56, 57] (see above).
We are now ready to formulate our second neurophenomenological hypothesis. We hypothesize that the topographic dedifferentiation and compression of the brain’s uni-transmodal topography leads to a corresponding topographic dedifferentiation and compression in the subject’s experience of their peripersonal and personal spaces. The less distinguished the uni-transmodal regions are from each other, the less subjects will experience a distinction of their personal and peripersonal spaces. In the most extreme case, uni- and transmodal regions are no longer distinguished at all which, as we postulate, should lead to the confusion and ultimately fusion of peripersonal and personal spaces, a hallucinatory space. Given that such hallucinatory space integrates and operates across the inner-outer boundaries, it makes possible for inner thoughts to be experienced as perceptions of the outer environment resulting in AVH.
Conclusions and future directions
Neuro-phenomenological hypotheses - Connecting brain, experience and symptoms
We here show how a more global topographic-dynamic view of the brain allows for direct connection of the neuronal and phenomenological levels of AVH. This led us to formulate two specific neurophenomenological hypothesis of AVH focusing on 1. Experience of fragmentation of contents and the brain’s structural-functional dysconnectivity; and 2. Experience of a hallucinatory space and the brain’s uni-transmodal gradients. While we here leave out several other phenomenological features of AVH, such as their commonly negative emotional content [36], our neurophenomenological hypotheses nevertheless could present a starting point for empirically testing and investigating the relations between brain, symptoms and experience of AVH [58].
Future investigation may also want to extend our neurophenomenological hypotheses to the biochemical level indagating imbalances in the levels of different transmitters such as dopamine and serotonin and how they modulate the brain’s uni-transmodal topography [59] in both physiological and pathological states. Furthermore, our neurophenomenological hypotheses do not stand in contradiction to other models of AVH but may rather be considered complementary [See Box 1]. Sensory perceptual [12, 60], cognitive [61, 62] and regional or network [63, 64] models of AVH may be well compatible with the here proposed spatiotemporal approach; Spatiotemporal Psychopathology [8, 58]: due to its global topographic-dynamic view of the brain, Spatiotemporal Psychopathology touches upon a more basic and fundamental level and thus provide a broader framework within which other models of AVH can be integrated.
From neurophenomenological hypotheses to clinical diagnosis and therapy
Our spatiotemporal approach opens the door for clinical differential diagnosis and, potentially, personalized therapy. First, it proposes direct connection from brain to symptom and subjective experience of AVH; such integrated brain-symptom-experience approach could then serve to investigate diagnostic markers that distinguishes AVH from other perceptual or cognitive disturbances. For instance, a recent study by Wengler et al. [45] observed distinct hierarchical gradients alterations in the presence of delusions and hallucinations in psychosis – the uni-transmodal gradient and its degree of dedifferentiation and compression may thus serve candidate diagnostic biomarker.
Presupposing a dimensional rather than categorical view of psychopathology, we suppose that the degree of uni-transmodal dedifferentiation and compression may serve as a strong candidate for a diagnostic biomarker of AVH independent of their origin, that is, whether organic, drug induced, occurring in healthy subjects, or associated with a mental disorder like schizophrenia or psychotic depression. Either transient (most common type in healthy subjects experiencing AVH) or more persistent (as in schizophrenia spectrum disorder) alterations in specific regions and networks may lead to a similar degree of topographic dedifferentiation and compression of the uni-transmodal gradient which, in turn, may lead to AVH. Additionally, it could be possible to assess and quantify the degree of uni-transmodal dedifferentiation and compression in each individual subject, which could offer a prognostic marker for, for example, the risk of AVH and the progression of its underlying pathology.
Finally, a more in depth understanding of the mechanism underling AVH could serve to guide the subsequent interventions, for example, in transcranial magnetic stimulation [65,66,67,68] which would need to change and re-balance the uni-transmodal topography (through stimulating key regions within that topography in an individualized subject-based way). Similarly, other forms of therapy could also target the uni-transmodal topographic balance and gradient like brain computer interface [69,70,71,72], alternative pharmacological delivery systems [73], and even psychotherapy, mindfulness, or other meditation techniques which are known to change the uni-transmodal topography [74]. In short, insight into the neurophenomenological mechanisms of AVH opens the door for precision treatment.
Data availability
This study did not involve the generation of new data.
References
Centers for Disease Control and Prevention. International classification of diseases, tenth revision, clinicalmodification (ICD-10-CM). (2023).
Sommer IE, Slotema CW, Daskalakis ZJ, Derks EM, Blom JD, van der Gaag M. The treatment of hallucinations in schizophrenia spectrum disorders. Schizophr Bull. 2012;38:704–14.
Smith LM, Yolland C, Rossell SL, Toh WL. The phenomenology of auditory verbal hallucinations in bipolar disorder. Psychol Psychother Theory Res Pr. 2023;96:399–409.
Toh WL, Thomas N, Hollander Y, Rossell SL. On the phenomenology of auditory verbal hallucinations in affective and non-affective psychosis. Psychiatry Res. 2020;290:113147.
De Loore E, Gunther N, Drukker M, Feron F, Sabbe B, Deboutte D, et al. Persistence and outcome of auditory hallucinations in adolescence: a longitudinal general population study of 1800 individuals. Schizophr Res. 2011;127:252–6.
Northoff G, Qin P, Feinberg T. Brain imaging of the self – conceptual, anatomical and methodological issues. Conscious cognition. 2010;20:52–63.
Northoff G, Stanghellini G How to link brain and experience? spatiotemporal psychopathology of the lived body. Front Hum Neurosci. 2016;10. https://doi.org/10.3389/fnhum.2016.00172
Northoff G, Daub J, Hirjak D. Overcoming the translational crisis of contemporary psychiatry – converging phenomenological and spatiotemporal psychopathology. Mol Psychiatry. 2023;28:4492–9.
Northoff G, Buccellato A, Zilio F. Connecting brain and mind through temporo-spatial dynamics: towards a theory of common currency. Phys Life Rev. 2025;52:29–43.
Northoff G, Wainio-Theberge S, Evers K. Is temporo-spatial dynamics the “common currency” of brain and mind? in quest of “spatiotemporal neuroscience. Phys Life Rev. 2020;33:34–54.
Georg Northoff From Brain Dynamics to the Mind, Academic Press, 2024, ISBN 9780128219355, https://doi.org/10.1016/B978-0-12-821935-5.00030-2
Parnas J, Yttri JE, Urfer-Parnas A. Phenomenology of auditory verbal hallucination in schizophrenia: an erroneous perception or something else?. Schizophr Res. 2024;265:83–88.
Frith C. The self in action: Lessons from delusions of control. Conscious Cogn. 2005;14:752–70.
Pienkos E, Giersch A, Hansen M, Humpston C, McCarthy-Jones S, Mishara A, et al. Hallucinations beyond voices: a conceptual review of the phenomenology of altered perception in psychosis. Schizophr Bull. 2019;45:S67–S77.
McCarthy-Jones S, Krueger J, Larøi F, Broome M, Fernyhough C. Stop, look, listen: the need for philosophical phenomenological perspectives on auditory verbal hallucinations. Front Hum Neurosci. 2013;7:127.
Sass L, Feyaerts J. Schizophrenia, the very idea: on self-disorder, hyperreflexivity, and the diagnostic concept. Schizophr Res. 2024;267:473–86.
Romeo Z, Spironelli C. Hearing voices in the head: Two meta-analyses on structural correlates of auditory hallucinations in schizophrenia. NeuroImage Clin. 2022;36:103241.
Sone M, Koshiyama D, Zhu Y, Maikusa N, Okada N, Abe O, et al. Structural brain abnormalities in schizophrenia patients with a history and presence of auditory verbal hallucination. Transl Psychiatry. 2022;12. 511.
Ford JM, Roach BJ, Jorgensen KW, Turner JA, Brown GG, Notestine R, et al. Tuning in to the voices: a multisite fMRI study of auditory hallucinations. Schizophr Bull. 2009;35:58–66.
Steinmann S, Leicht G, Mulert C. The interhemispheric miscommunication theory of auditory verbal hallucinations in schizophrenia. Int J Psychophysiol. 2019;145:83–90.
Modinos G, Costafreda SG, Van Tol MJ, McGuire PK, Aleman A, Allen P. Neuroanatomy of auditory verbal hallucinations in schizophrenia: a quantitative meta-analysis of voxel-based morphometry studies. Cortex. 2013;49:1046–55.
Mørch-Johnsen L, Nesvåg R, Jørgensen KN, et al. Auditory cortex characteristics in schizophrenia: associations with auditory hallucinations. Schizophr Bull. 2017;43:75–83.
Huang J, Zhuo C, Xu Y, Lin X. Auditory verbal hallucination and the auditory network: from molecules to connectivity. Neuroscience. 2019;410:59–67.
Kompus K, Westerhausen R, Hugdahl K. The “paradoxical” engagement of the primary auditory cortex in patients with auditory verbal hallucinations: a meta-analysis of functional neuroimaging studies. Neuropsychologia. 2011;49:3361–9.
Fuentes-Claramonte P, Soler-Vidal J, Salgado-Pineda P, et al. Auditory hallucinations activate language and verbal short-term memory, but not auditory, brain regions. Sci Rep. 2021;11:18890.
Sheffield JM, Rogers BP, Blackford JU, Heckers S, Woodward ND. Insula functional connectivity in schizophrenia. Schizophr Res. 2020;220:69–77.
Tian Y, Zalesky A, Bousman C, Everall I, Pantelis C. insula functional connectivity in schizophrenia: subregions, gradients, and symptoms. Biol Psychiatry Cogn Neurosci Neuroimaging. 2019;4:399–408.
Xue K, Chen J, Wei Y, Chen Y, Han S, Wang C, et al. Altered dynamic functional connectivity of auditory cortex and medial geniculate nucleus in first-episode, drug-naïve schizophrenia patients with and without auditory verbal hallucinations. Front Psychiatry. 2022;13:963634.
Panikratova YR, Lebedeva IS, Akhutina TV, Tikhonov DV, Kaleda VG, Vlasova RM. Executive control of language in schizophrenia patients with history of auditory verbal hallucinations: a neuropsychological and resting-state fMRI study. Schizophr Res. 2023;262:201–10.
Javitt DC, Sweet RA. Auditory dysfunction in schizophrenia: integrating clinical and basic features. Nat Rev Neurosci. 2015;16:535–50.
Zhang Y, Wang J, Lin X, Yang M, Qi S, Wang Y, et al. Distinct brain dynamic functional connectivity patterns in schizophrenia patients with and without auditory verbal hallucinations. Front Hum Neurosci. 2022;16:838181.
Gao J, Zhang D, Wang L, Wang W, Fan Y, Tang M, et al. Altered effective connectivity in schizophrenic patients with auditory verbal hallucinations: a resting-state fMRI study with granger causality analysis. Front Psychiatry. 2020;11:575.
Woods A, Jones N, Bernini M, Callard F, Alderson-Day B, Badcock JC, et al. Interdisciplinary approaches to the phenomenology of auditory verbal hallucinations. Schizophr Bull. 2014;40(Suppl 4):S246–S254.
Northoff G. Beyond task response—Pre-stimulus activity modulates contents of consciousness. Phys Life Rev. 2024;49:19–37.
Yttri JE, Urfer-Parnas A, Parnas J. Auditory verbal hallucinations in schizophrenia, part II: phenomenological qualities and evolution. J Nerv Ment Dis. 2022;210:659–64.
Henriksen MG, Raballo A, Parnas J. The pathogenesis of auditory verbal hallucinations in schizophrenia: a clinical–phenomenological account. Philos Psychiatry Psychol. 2015;22:165–81.
Larøi F, De Haan S, Jones S, Raballo A. Auditory verbal hallucinations: dialoguing between the cognitive sciences and phenomenology. Phenomenol Cogn Sci. 2010;9:225–40.
Parnas J, Urfer-Parnas A, Stephensen H. Double bookkeeping and schizophrenia spectrum: divided unified phenomenal consciousness. Eur Arch Psychiatry Clin Neurosci. 2021;271:1513–23.
Rasmussen AR. Anomalies of imagination and development of psychosis: a phenomenological account. Schizophr Res. 2024;264:204–10.
Northoff G, Gomez-Pilar J. Overcoming rest-task divide-abnormal temporospatial dynamics and its cognition in schizophrenia. Schizophr Bull. 2021;47:751–65.
Huang AS, Woodward ND. The brain and schizophrenia: from paradigm shifts to shifting gradients. Biol Psychiatry Cogn Neurosci Neuroimaging. 2023;8:1173–5.
Margulies DS, Ghosh SS, Goulas A, Falkiewicz M, Huntenburg JM, Langs G, et al. Situating the default-mode network along a principal gradient of macroscale cortical organization. Proc Natl Acad Sci. 2016;113:12574–9.
Cattarinussi G, Grimaldi DA, Sambataro F. Spontaneous brain activity alterations in first-episode psychosis: a meta-analysis of functional magnetic resonance imaging studies. Schizophr Bull. 2023;49:1494–507.
Dong D, Yao D, Wang Y, Hong SJ, Genon S, Xin F, et al. Compressed sensorimotor-to-transmodal hierarchical organization in schizophrenia. Psychol Med. 2023;53:771–84.
Wengler K, Goldberg AT, Chahine G, Horga G. Distinct hierarchical alterations of intrinsic neural timescales account for different manifestations of psychosis. eLife. 2020;9:e56151.
Geng H, Xu P, Sommer IE, Luo YJ, Aleman A, Ćurčić-Blake B. Abnormal dynamic resting-state brain network organization in auditory verbal hallucination. Brain Struct Funct. 2020;225:2315–30.
Weber S, Johnsen E, Kroken RA, Løberg EM, Kandilarova S, Stoyanov D, et al. Dynamic functional connectivity patterns in schizophrenia and the relationship with hallucinations. Front Psychiatry. 2020;11:227.
Rabellino D, Frewen PA, McKinnon MC, Lanius RA. Peripersonal space and bodily self-consciousness: implications for psychological trauma-related disorders. Front Neurosci. 2020;14:586605.
Bogdanova OV, Bogdanov VB, Dureux A, Farnè A, Hadj-Bouziane F. The peripersonal space in a social world. Cortex. 2021;142:28–46.
Ferri F, Tajadura-Jiménez A, Väljamäe A, Vastano R, Costantini M. Emotion-inducing approaching sounds shape the boundaries of multisensory peripersonal space. Neuropsychologia. 2015;70:468–75.
Serino S, Trabanelli S, Jandus C, Fellrath J, Grivaz P, Paladino MP, et al. Sharpening of peripersonal space during the COVID-19 pandemic. Curr Biol. 2021;31:R889–R890.
Zapetis SL, Nasiriavanaki Z, Luther L, Holt DJ. Neural correlates of variation in personal space and social functioning in schizophrenia and healthy individuals. Schizophr Bull. 2022;48:1075–84.
Stanghellini G, Fernandez AV, Ballerini M, Blasi S, Belfiore E, Cutting J, et al. Abnormal space experiences in persons with schizophrenia: an empirical qualitative study. Schizophr Bull. 2020;46:530–9.
Paredes R, Ferri F, Seriès P. Influence of E/I balance and pruning in peri-personal space differences in schizophrenia: A computational approach. Schizophr Res. 2022;248:368–77.
Holt DJ, Boeke EA, Coombs G 3rd, DeCross SN, Cassidy BS, et al. Abnormalities in personal space and parietal–frontal function in schizophrenia. NeuroImage Clin. 2015;9:233–43.
Sabbah SG, Northoff G. Basic self-disturbance in schizophrenia: from neuronal to mental topographic dedifferentiation. Schizophr Res. 2024;274:327–8.
Sabbah SG, Northoff G. Global neural self-disturbance in schizophrenia: a systematic fmri review. Schizophr Res. 2024;269:163–73.
Northoff G, Hirjak D. Spatiotemporal psychopathology – an integrated brain-mind approach and catatonia. Schizophr Res. 2024;263:151–9.
Conio B, Martino M, Magioncalda P, Escelsior A, Inglese M, Amore M, et al. Opposite effects of dopamine and serotonin on resting-state networks: review and implications for psychiatric disorders. Mol Psychiatry. 2020;25:82–93.
Powers AR, Kelley M, Corlett PR. Hallucinations as top-down effects on perception. Biol Psychiatry Cogn Neurosci Neuroimaging. 2016;1:393–400.
López-Silva P, Harrow M, Jobe TH, Tufano M, Harrow H, Rosen C. ‘Are these my thoughts?’: a 20-year prospective study of thought insertion, thought withdrawal, thought broadcasting, and their relationship to auditory verbal hallucinations. Schizophr Res. 2024;265:46–57.
McCleery A, Wynn JK, Green MF. Hallucinations, neuroplasticity, and prediction errors in schizophrenia. Scand J Psychol. 2018;59:41–48.
Hare SM. Hallucinations: a functional network model of how sensory representations become selected for conscious awareness in schizophrenia. Front Neurosci. 2021;15:733038.
Mallikarjun PK, Lalousis PA, Dunne TF, Heinze K, Reniers RL, Broome MR, et al. Aberrant salience network functional connectivity in auditory verbal hallucinations: a first episode psychosis sample. Transl Psychiatry. 2018;8:69.
Xie Y, Li C, Guan M, Zhang T, Ma C, Wang Z, et al. The efficacy of low frequency repetitive transcial magnetic stimulation for treating auditory verbal hallucinations in schizophrenia: Insights from functional gradient analyses. Heliyon. 2024;10:e30194.
Mehta DD, Siddiqui S, Ward HB, Steele VR, Pearlson GD, George TP. Functional and structural effects of repetitive transcranial magnetic stimulation (rTMS) for the treatment of auditory verbal hallucinations in schizophrenia: a systematic review. Schizophr Res. 2024;267:86–98.
Yuanjun X, Guan M, Zhang T, Ma C, Wang L, Lin X, et al. Targeting auditory verbal hallucinations in schizophrenia: effective connectivity changes induced by low-frequency rTMS. Transl Psychiatry. 2024;14:393.
Kubera KM, Barth A, Hirjak D, Thomann PA, Wolf RC. Noninvasive brain stimulation for the treatment of auditory verbal hallucinations in schizophrenia: methods, effects and challenges. Front Syst Neurosci. 2015;9:131.
Fernández-Sotos P, García-Martínez B, Ricarte JJ, Latorre JM, Sánchez-Morla EM, Fernández-Caballero A, et al. Electroencephalographic spectral analysis from a wireless low-cost brain-computer interface for symptom capture of auditory verbal hallucinations in schizophrenia. Schizophr Res. 2020;220:297–9.
García-Martínez B, Fernández-Sotos P, Ricarte JJ, et al. Detection of auditory hallucinations from electroencephalographic brain–computer interface signals. Cogn Syst Res. 2024;83:101176.
Fernández-Caballero A, Navarro E, Fernández-Sotos P, González P, Ricarte JJ, Latorre JM, et al. Human-avatar symbiosis for the treatment of auditory verbal hallucinations in schizophrenia through virtual/augmented reality and brain-computer interfaces. Front Neuroinform. 2017;11:64.
McCarthy-Jones S. Taking back the brain: could neurofeedback training be effective for relieving distressing auditory verbal hallucinations in patients with schizophrenia?. Schizophr Bull. 2012;38:678–82.
Ferreira MD, Duarte J, Veiga F, Paiva-Santos AC, Pires PC. Nanosystems for brain targeting of antipsychotic drugs: an update on the most promising nanocarriers for increased bioavailability and therapeutic efficacy. Pharmaceutics. 2023;15:678.
Cooper AC, Ventura B, Northoff G. Beyond the veil of duality – topographic reorganization model of meditation. Neurosci Conscious. 2022;2022:niac013.
Thakkar KN, Mathalon DH, Ford JM. Reconciling competing mechanisms posited to underlie auditory verbal hallucinations. Philos Trans R Soc B Biol Sci. 2021;376:20190702.
Waters F, Allen P, Aleman A, et al. Auditory hallucinations in schizophrenia and nonschizophrenia populations: a review and integrated model of cognitive mechanisms. Schizophr Bull. 2012;38:683–93.
Northoff G. Are auditory hallucinations related to the brain’s resting state activity? a “neurophenomenal resting state hypothesis”. Clin Psychopharmacol Neurosci. 2014;12:189–95.
Wolff A, Northoff G. Temporal imprecision of phase coherence in schizophrenia and n psychosis-dynamic mechanisms and diagnostic marker. Mol Psychiatry. 2024;29:425–38.
Author information
Authors and Affiliations
Contributions
AFC and GN contributed equally to the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no conflicts of interest.
Ethics approval and informed consent
This study did not involve human participants, animals, or the use of biological materials.
Consent for publication
This study does not contain any identifiable images or personal data from human participants.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Carluccio, A.F., Northoff, G. From neuronal to mental topography – Neurophenomenology of auditory hallucinations. Transl Psychiatry 15, 496 (2025). https://doi.org/10.1038/s41398-025-03664-1
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41398-025-03664-1
