
Abstract
Narcolepsy type 1 (NT1) is characterized by sleep-onset rapid eye movement periods (SOREMPs), reflecting dysregulated rapid eye movement (REM) sleep control. Treatment response variability in SOREMP persistence remains poorly understood, particularly regarding hypothalamic functional connectivity (FC) and depressive symptoms. This study investigated clinical, polysomnographic, and neuroimaging differences between NT1 patients with low (0–1) versus high (≥2) post-treatment SOREMPs, and explored whether hypothalamic FC mediates the relationship between depressive symptoms and SOREMPs outcomes. One hundred ten NT1 patients were categorized into low (n = 62) and high (n = 48) post-treatment SOREMPs groups. Demographic, clinical variables (symptoms and questionnaires), and polysomnography (PSG)/multiple sleep latency test (MSLT) parameters were compared. Resting-state fMRI assessed hypothalamic FC with whole-brain regions. LASSO regression modeled associations between FC, sleep latency, and clinical variables, while mediation analysis tested hypothalamic pathways as mediators of depression-SOREMP relationships. High post-treatment SOREMPs patients exhibited shorter pre/post-treatment REM sleep latency, lower post-treatment wakefulness index, and higher depressive symptom prevalence compared to low SOREMPs patients. Hypothalamic FC differed significantly between groups: low SOREMPs patients showed enhanced connectivity in right medial hypothalamus-right thalamus/left precuneus, left medial hypothalamus-left inferior parietal lobule (IPL), and right lateral hypothalamus-left IPL pathways, but reduced connectivity in left lateral hypothalamus-right insula/left anterior cingulate cortex pathways (p < 0.05, GRF-corrected). LASSO regression identified left medial hypothalamus-left IPL FC as a significant predictor of post-treatment MSLT mean sleep latency (β = 0.272, p = 0.001), alongside age (β = −0.256, p = 0.002) and pre-treatment sleep latency (β = 0.392, p < 0.001). Mediation analysis revealed complete mediation by two hypothalamic pathways: depressive symptoms predicted reduced right lateral hypothalamus-left IPL FC (indirect effect: 0.15–1.05), associated with fewer SOREMPs, and increased left lateral hypothalamus-right insula FC (indirect effect: 0.08–1.14), associated with more SOREMPs. Hypothalamic-parietal/insular FC abnormalities link depressive symptoms to post-treatment SOREMP variability in NT1, with specific pathways mediating opposing effects on REM sleep regulation. These findings highlight hypothalamic connectivity as a critical neural substrate for treatment response, integrating sleep-wake and emotional processing networks. Targeting these pathways may improve personalized management for NT1 patients with comorbid depression and treatment-resistant SOREMPs.
Similar content being viewed by others

Introduction
Narcolepsy type 1 (NT1), characterized by the loss of hypothalamic orexin neurons, is defined by excessive daytime sleepiness, cataplexy, and pathognomonic sleep-onset rapid eye movement periods (SOREMPs) on polysomnography (PSG) and multiple sleep latency tests (MSLT) [1, 2]. SOREMPs, indicative of dysregulated rapid eye movement (REM) sleep control, serve as a key diagnostic criterion and reflect the severity of REM sleep intrusion into wakefulness [3, 4]. While pharmacotherapies targeting hypocretin deficiency sleepiness (e.g., modafinil) or REM sleep dysregulation (e.g., selective serotonin reuptake inhibitors) have improved clinical outcomes, interindividual variability in treatment response remains pronounced, particularly in the persistence of SOREMPs after intervention [2, 5,6,7]. For instance, recent trials with the hypocretin receptor agonist demonstrated significant improvements in both wakefulness and cataplexy [8, 9] but did not have reports on the effects of treatment on SOREMPs, and the effects of stimulants (e.g., modafinil) and antidepressants on SOREMPs are not yet fully understood, underscoring the need to understand the neural and clinical factors driving this heterogeneity.
Previous research has highlighted the hypothalamus as a pivotal node in REM sleep regulation [10,11,12,13], with its functional connectivity (FC) to brain regions involved in sleep-wake homeostasis (e.g., thalamus, parietal lobe) and emotion processing (e.g., amygdala, cingulate cortex) playing a role in narcolepsy pathophysiology. For example, altered hypothalamic-parietal connectivity has been linked to sleep fragmentation and daytime sleepiness in NT1, while hypothalamic-amygdala interactions may contribute to emotional instability and cataplexy [14]. However, the specific mechanisms by which hypothalamic FC influences treatment-responsive SOREMPs remain unclear.
Depressive symptoms, which affect up to 50% of NT1 patients, further complicate clinical trajectories [15]. These symptoms are often comorbid with sleep dysfunction and may exacerbate REM sleep dysregulation through shared neural pathways. For instance, the hypothalamic-pituitary-adrenal (HPA) axis, a key regulator of stress responses, is dysregulated in both depression and NT1, potentially influencing sleep architecture and treatment outcomes [16]. Emerging evidence suggests that hypothalamic FC with emotion-processing regions (e.g., amygdala, prefrontal cortex) may mediate the relationship between depressive symptoms and sleep abnormalities [17,18,19]. However, the extent to which these pathways interact with treatment-induced changes in SOREMPs remains unexplored.
In this study, we sought to characterize differences in demographic, clinical, and polysomnographic parameters between NT1 patients with low (0–1) versus high (≥2) post-treatment SOREMPs. Leveraging resting-state functional magnetic resonance imaging (fMRI), we investigated group differences in hypothalamic FC with the whole brain, followed by statistical modeling to examine associations between FC, sleep latency, and depressive symptoms. Specifically, we hypothesized that altered hypothalamic connectivity would correlate with post-treatment SOREMPs and mediate the relationship between depressive symptoms and treatment response. By integrating clinical, polysomnographic, and neuroimaging data, this study aims to uncover neural mechanisms underlying heterogeneous treatment outcomes in NT1, with implications for precision medicine approaches targeting both sleep dysregulation and emotional comorbidities.
Participants and methods
Participants
A total of 110 drug-naïve NT1 patients from the Sleep Medicine Center in Peking University People’s Hospital participated in this study. NT1 was diagnosed according to the ICSD-3: the presence of excessive daytime sleepiness lasting at least 3 months, typical cataplexy, with mean sleep latency ≤ 8 min and 2 or more sleep-onset REM periods (SOREMPs) on the multiple sleep latency test (MSLT) or during the previous night polysomnography (PSG), or lower levels of orexin (≤110 pg/mL) in cerebral spinal fluid. Patients with cognitive dysfunction (measured by Mini-mental state examination, MMSE <27), psychiatric disorders, substance/alcohol abuse, or other serious diseases were excluded. Patients with congenital or inherited diseases and MRI contraindications were not recruited in this study. HLA-DQB1*06:02 genotyping was also determined for all NT1 patients. Out of 110 NT1 patients, 68 individuals underwent lumbar puncture to measure the level of hypocretin in their cerebrospinal fluid. All participants provided written informed consent. This study was conducted following the Declaration of Helsinki. The ethical committee of Peking University People’s Hospital approved the study (2022PHB089).
Polysomnography and multiple sleep latency test
All participants underwent a standardized protocol consisting of polysomnography (PSG, Respironics LE-Series Physiological Monitoring System, Alice 6 LE, FL, USA) and multiple sleep latency testing (MSLT) to confirm the diagnosis of narcolepsy, following the guideline of the American Academy of Sleep Medicine (AASM) [20]. On Day 1, participants were admitted to the sleep laboratory, and PSG was performed overnight (22:00–06:00) to assess nocturnal sleep architecture. The following morning (Day 2), MSLT was conducted every 2 h between 08:00 and 17:00 to measure mean sleep latency and rapid eye movement (REM) sleep onset latency. Then participants underwent functional magnetic resonance imaging (fMRI) immediately following MSLT. Pharmacological interventions were initiated sequentially post-fMRI. On the evening of Day 2, clomipramine (12.5 mg orally) was administered at 20:00 to address cataplexy. On the morning of Day 3, methylphenidate (9 mg orally) was given at 08:00 to promote wakefulness and clomipramine (12.5 mg) was continued at 20:00 on the evening of Day 3. On Day 4 and Day 5, methylphenidate (9 mg) was administered again at 08:00, followed by clomipramine (12.5 mg) at 20:00. A second PSG was performed overnight on Day 4(22:00–06:00) to assess treatment effects on sleep quality, followed by repeat MSLT between 08:00 and 17:00on Day 5. All medications were administered under clinical supervision, and vital signs were monitored throughout the protocol. This structured approach ensured temporal separation of diagnostic assessments and pharmacological interventions to minimize confounding effects on neuroimaging and sleep metrics. Detailed information about PSG and MSLT can be found in the supplementary materials.
Questionnaires
The degree of subjective sleepiness, depressed mood, and impulsivity in NT1 patients was measured by separate questionnaires.
Epworth Sleepiness Scale (ESS) was used to measure subjective sleepiness. ESS is a measure of a person’s general level of daytime sleepiness. For adolescents, ESS was modified slightly to be more applicable: the description regarding “taking alcohol after lunch” was deleted in item No. 7 and the subject was considered to be “a passenger in the car” instead of “a driver” in item No. 8 [21].
The Center for Epidemiologic Studies Depression Scale for Children (CES-DC) was used for measuring the depressed mood in adolescent (age ≤ 17) NT1 patients 11. A total score ≥20 suggests subjects with depressed symptoms [22].
The Self-Rating Depression Scale (SDS) was used for measuring depressed mood in adult (age ≥ 18) NT1 patients. A total standardized score ≥ 53 suggests subjects with depressed symptoms [23].
The Barratt impulse scale (BIS)-11 was used to measure impulsivity 13. The whole scale is composed of 30 items, using a 5-point Likert scale for each item, with a higher total score meaning higher impulsivity [24].
Imaging data acquisition and preprocessing
Resting fMRI scans and high-resolution T1-weighted structural images were acquired using a 3 T scanner (Siemens, Skyra, Germany) for NT1 patients, immediately after the MSLT. Participants were instructed to stay fully awake, minimize movement, and to keep their eyes open during the MRI scan, with continuous supervision. If sleep onset was detected via video, patients were alerted by a loud noise. They were also instructed in advance not to respond to any noises during the scan to prevent speaking. In addition, emotional triggers were avoided to reduce the risk of a cataplexy attack. Detailed information on the fMRI scanning procedure provided in supplementary materials.
fMRI data preprocessing
A total of 240 functional volumes were obtained during the resting-state fMRI scans for each subject. The preprocessing of fMRI data included slice timing correction, and head-motion correction, registration to 3D T1-weighted structural images, normalization to the Montreal Neurological Institute template, smoothing, and removal of linear trends, as detailed in our previously published reports. Further information on fMRI data preprocessing and head motion control can be found in supplementary materials.
Seed-based functional connectivity analyses
Two subregions of the hypothalamus were selected as the regions of interest (ROIs): the bilateral medial hypothalamus (MH) seed (MNI [x, y, z] = ±4, −2, −12 mm) and the lateral hypothalamus (LH) seed (MNI [x, y, z] = ±6, −9, −10 mm) with a 2 mm radius sphere, which had been used in previous research [25].
In the first-level analysis, the functional connectivity between the bilateral MH and LH, and the rest of the brain was computed. This was achieved by extracting the blood oxygen level-dependent time course from each ROI, separately, and calculating Pearson’s correlation coefficients between the time course in the LH/MH and every voxel in the whole brain. The correlation coefficients were then transformed to z-scores using Fisher’s z-transformation to improve their normality and enable better analysis using the General Linear Model in the second-level analysis.
Statistical analysis
We use descriptive statistics to summarize baseline population characteristics and clinical features in all NT1 patients. We report normally distributed continuous variables using means with standard deviations (SD), reported medians with interquartile ranges (IQR) for non-normal continuous variables, and raw numbers with corresponding percentages for categorical variables.
Step 1 Classify NT1 patients into two groups according to the SOREMPs after medication treatment
All NT1 patients were classified into two groups according to the number of SOREMPs on MSLT follow-up after medication treatment: one group with SOREMPs ≥ 2 episodes (high post-treatment SOREMPs group), and the other group with 0 or 1 SOREM episode (low post-treatment SOREMPs group). Differences between these two groups in terms of demographic characteristics, questionnaires results, PSG and MSLT parameters were compared. Continuous variables were compared using the two independent-samples t-test or Wilcoxon rank-sum test. Categorical variables were compared using the χ2 test. Statistical tests were two-tailed and the significance level was set at P < 0.05.
Step 2 Differentiating functional connectivity between two groups
A two-sample t-test was utilized to compare the inter-group differences in functional connectivity between LH/MH and the whole brain between two groups of NT1 patients. To address multiple comparisons, all statistical maps were assigned threshold at Gaussian random field correction (GRF), with voxel P = 0.001and cluster P = 0.05 [26, 27].
Step 3 Association functional connectivity with mean sleep latency after medication treatment
To identify factors associated with the mean sleep latency (MSL) on the MSLT after medication treatment, we employed a two-step statistical approach. The dependent variable was the post-treatment MSLT mean sleep latency, while the independent variables included: 1) demographic information (e.g., age, gender, body mass index); 2) clinical symptoms of the disease (e.g., age at onset, presence of hallucinations); 3) scores from questionnaires (e.g., Epworth Sleepiness Scale [ESS] score); 4) pretreatment PSG and MSLT parameters (e.g., total sleep time, pretreatment MSL); and 5) functional connectivity showing significant group differences previously identified in Step 2.
Step 3.1 Variable selection using LASSO regression
We applied Least Absolute Shrinkage and Selection Operator (LASSO) regression to screen for independent variables associated with post-treatment MSLT mean sleep latency. This method was chosen to handle potential high-dimensionality and collinearity among independent variables, leveraging a regularization penalty to shrink less relevant variables’ coefficients to zero, thereby selecting a parsimonious set of predictors. Independent variables with a regression coefficient equal to zero after the shrinkage process are excluded from the model while independent variables with nonzero regression coefficient are most strongly associated with the dependent variable. Based on the type measure of -2log-likelihood and binomial family, the LASSO regression analysis running in R software runs 10 times K cross-validation for centralization and normalization of included variables and then picks the best lambda value. “Lambda.lse” gives a model with good performance but the least number of independent variables.
Step 3.2 Model construction via forward stepwise linear regression
Using the subset of variables selected by LASSO, we then performed forward stepwise linear regression in SPSS to build a final model. This iterative procedure systematically added variables to identify the optimal set of predictors significantly associated with post-treatment MSLT mean sleep latency. The result was a linear regression model explicitly specifying which independent variables were independently related to the post-treatment MSLT mean sleep latency, accounting for potential confounding effects and ensuring statistical robustness.
Step 4 The relationship between depression symptoms, functional connectivity and SOREMPs after medication treatment
To investigate the mechanism through which depressive symptoms influence post-treatment SOREMPs via functional connectivity, a mediation analysis was conducted. Based on Step 1 results showing a higher proportion of depressive symptoms in patients with post-treatment SOREMP ≥ 2 episodes, functional connectivity with significant between-group differences identified in Step 2 served as the mediator variable. The independent variable was the presence of depressive symptoms before treatment (dichotomous: 0 = absent, 1 = present), and the dependent variable was post-treatment SOREMPs (dichotomous: 0 = < 2 episodes, 1 = ≥ 2 episodes). All analyses were performed using the PROCESS (Version 4.2) for SPSS, which allows specification of mediator models with dichotomous independent variables and dichotomous dependent variables through logistic regression. Direct and total effects were estimated using logistic regression due to the dichotomous dependent variable, reporting odds ratios (OR) and 95% confidence intervals (CI). The indirect effect was evaluated using bias-corrected bootstrap resampling with 5,000 resamples, a common method to handle non-normality in small-to-moderate samples. Significance was determined by whether the 95% bootstrap CI for the indirect effect excluded zero. A significant total effect and a significant indirect effect with a non-significant direct effect would indicate complete mediation. If both the direct effect and indirect effect are significant, partial mediation is supported. Non-significant indirect effect suggests functional connectivity does not mediate the relationship between depressive symptoms and post-treatment SOREMPs. This approach rigorously tests the hypothesized mediational mechanism, providing empirical evidence on whether functional connectivity serves as a critical pathway through which depressive symptoms influence medication treatment outcomes in SOREMPs.
Results
Among 110 NT1 patients, 62 patients showed 0 or 1 SOREMP episode (low post-treatment SOREMPs group) while the other 48 patients showed SOREMPs ≥ 2 episodes (high post-treatment SOREMPs group) after medication treatment.
Step 1 Differences between two groups of NT1 patients in demographic characteristics, disease symptoms, and polysomnography parameters
In terms of demographic characteristics, two-sample t-tests revealed no significant between-group differences in gender (χ² = 1.186, P = 0.278), age (t = 0.525, P = 0.601), or body mass index (BMI) (t = −1.106, P = 0.163) between the two groups of NT1 patients.
In terms of clinical symptoms, no significant group differences were observed in subjective sleepiness measured by the Epworth Sleepiness Scale (t = −1.109, P = 0.27), frequency of hallucinations (χ² = 1.402, P = 0.236), or frequency of sleep paralysis (χ² = 0.972, P = 0.324).
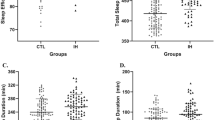
In terms of PSG and MSLT parameters, high post-treatment SOREMPs group patients showed significantly shorter REM sleep latency in pre-treatment PSG (Z = −2.729, P = 0.006), mean REM sleep latency in MSLT (Z = −2.593, P = 0.01), post-treatment REM sleep latency in PSG (Z = −2.267, P = 0.023), and mean REM sleep latency in post-treatment MSLT (Z = −2.146, P = 0.032) compared to those in low post-treatment SOREMPs group patients. Additionally, the wakefulness index in post-treatment PSG (Z = −3.011, P = 0.003) and mean sleep latency in post-treatment MSLT (Z = −2.146, P = 0.032) were also significantly lower in high post-treatment SOREMPs group.
Notably, the proportion of patients with depressive symptoms (assessed by questionnaire) was higher in high post-treatment SOREMPs group (56.3%) compared to those in low post-treatment SOREMPs group (34.4%; χ² = 5.191, P = 0.023). All the differences between two groups of NT1 patients in demographic characteristics, disease symptoms, and polysomnography parameters can be found in Table 1. Post-treatment SOREMPs counts among the 110 patients showed a bimodal distribution, with 56.4% exhibiting 0–1 SOREMPs and 43.6% exhibiting ≥ 2 SOREMPs. For mean REM latency from MSLT, pre-treatment mean REM latency from MSLT in the low post-treatment SOREMPs group had a median of 1.5 min (IQR: 0.7–2.5), while the high post-treatment SOREMPs group had a median of 0.85 min (IQR: 0.3–1.675). Post-treatment mean REM latency from MSLT in the low post-treatment SOREMPs group had a median of 5 min (IQR: 2–20), while the high post-treatment SOREMPs group had a median of 2.15 min (IQR: 1.23–3.95), detailed distributions are provided in supplementary materials.
Step 2 Differences in the connectivity between hypothalamus and whole brain in two groups of NT1 patients
Specifically, low post-treatment SOREMPs group patients showed increased FC in the following pathways compared to the high post-treatment SOREMPs group patients: right medial hypothalamus (rMH) with right thalamus and left posterior cingulate cortex, left MH (lMH) with left inferior parietal lobule (IPL), right lateral hypothalamus (rLH) with left IPL.
Conversely, low post-treatment SOREMPs group patients exhibited decreased FC in the left LH (lLH) with right insula and right middle cingulate cortex compared to the high post-treatment SOREMPs group patients. All findings survived after GRF for multiple comparisons correction, with voxel P = 0.001 and cluster P = 0.05. Detailed spatial patterns and statistical values are summarized in Fig. 1 and Table 2.

The red color indicates the functional connectivity between the right medial hypothalamus and the right thalamus, left posterior cingulate gyrus, the blue color indicates the functional connectivity between the left medial hypothalamus and the left inferior parietal lobule, the orange color indicates the functional connectivity between the right lateral hypothalamus and the left inferior parietal lobule, and the green color indicates the functional connectivity between the left lateral hypothalamus and the right insula, right middle cingulate gyrus. All findings survived after GRF for multiple comparisons correction, with voxel P = 0.001 and cluster P = 0.05. Abbreviation: MH.R, right medial hypothalamus; MH.L, left medial hypothalamus; LH.R, right lateral hypothalamus; LH.L, left lateral hypothalamus; THA.R, right thalamus; PCC.L, left posterior cingulate cortex; IPL.L, left inferior parietal lobule; INS.R, right insular; MCC.R, right middle cingulate cortex.
Step 3 Association functional connectivity with mean sleep latency after medication treatment
LASSO regression analysis identified five variables associated with post-treatment mean sleep latency on the MSLT from the initially included predictors: age, N3 sleep percentage from pre-treatment PSG, pre-treatment MSLT mean sleep latency, Epworth Sleepiness Scale (ESS) score, and functional connectivity (FC) between the left medial hippocampus (MH) and left inferior parietal lobule (IPL).
Subsequent forward stepwise linear regression further refined these associations, retaining three significant predictors of post-treatment MSLT mean sleep latency: age (β = −0.256, 95%CI (−0.238, −0.054), P = 0.002), pre-treatment MSLT mean sleep latency (β = 0.392, 95%CI (0.58, 1.39), P < 0.001), left MH-left IPL functional connectivity (β = 0.272, 95%CI (4.467, 17.422), P = 0.001). The linear regression model demonstrated moderate fit with an adjusted R² of 0.295 (F = 9.9, P = 0.002).
Step 4 The relationship between depression symptoms, functional connectivity and SOREMPs after medication treatment
Mediation analysis revealed a complete mediating effect of functional connectivity (FC) in the relationship between depressive symptoms and post-treatment SOREMPs among NT1 patients. Specifically, depressive symptoms influenced post-treatment SOREMPs exclusively through two distinct FC pathways:
Right LH-left IPL pathway: depressive symptoms were associated with reduced FC between the right LH and left IPL (β = −0.05, 95%CI (−0.09, −0.01), SE = 0.02, P = 0.0096), which in turn predicted a decline in post-treatment SOREMPs (β = −10.16, 95%CI (−15.42, −4.89), SE = 2.69, P = 0.0002). The indirect effect via this pathway was significant (95% CI: 0.15 to 1.05, bootstrap resampling = 5000), and the direct effect of depressive symptoms on SOREMPs became non-significant (P > 0.05) when the mediator was included.
Left LH-right insula pathway: depressive symptoms were also linked to increased FC between the left LH and right insula (β = 0.04, 95%CI (0.01, 0.08), SE = 0.02, P = 0.026), which independently predicted higher post-treatment SOREMPs (β = 11.81, 95%CI (6.2, 17.43), SE = 2.87, P < 0.001). The indirect effect through this pathway was robust (95% CI: 0.08 to 1.14), and again, the direct effect of depression on SOREMPs vanished (P > 0.05), confirming complete mediation for both pathways. No other FC metrics or direct effects emerged as significant. The mediating roles of these hypothalamic-parietal/insular connections are visually depicted in Fig. 2, illustrating the sequential relationships among depressive symptoms, FC alterations, and treatment response.

Abbreviation: rLH, right lateral hypothalamus; lIPL, left inferior parietal lobule; lLH, left lateral hypothalamus; rinsular, right insular; FC, functional connectivity.
Discussion
While pharmacological interventions aim to normalize sleep-wake cycles in NT1 patients, interindividual variability in post-treatment SOREMPs suggests uncharacterized neurobiological mechanisms. This study dissects clinical, polysomnographic, and functional connectivity between NT1 patients with two or more versus zero or one post-treatment SOREMPs, underscoring hypothalamic functional connectivity (FC) as a critical mediator linking depression and treatment response.
Rationale for grouping NT1 patients by post-treatment SOREMPs threshold
The classification of NT1 patients into low (0–1) versus high (≥2) post-treatment SOREMPs groups is anchored in the International Classification of Sleep Disorders (ICSD-3), which designates two or more SOREMPs during MSLT as a core diagnostic criterion for NT1 [20]. This threshold reflects the pathophysiological hallmark of NT1: hypocretin neuron loss leading to dysregulated REM sleep gating, where SOREMPs emerge from inappropriate rapid transition into REM sleep [28]. While treatment aims to normalize sleep-wake cycles, residual two or more SOREMPs suggest persistent REM sleep instability, indicating incomplete modulation of hypocretin-mediated neural circuits. This grouping strategy aligns with clinical and research practices, as SOREMPs serve as a validated biomarker for monitoring treatment response in NT1, reflecting both pharmacological efficacy and underlying neurobiological dysfunction [14]. The SOREMPs≥2 threshold mirrors diagnostic criteria, enabling direct comparison between pre-treatment pathophysiology and post-treatment outcomes. This approach is widely adopted in clinical trials and observational studies. For instance, Mignot et al. [29] emphasized the utility of SOREMPs in characterizing treatment responses in NT1. These findings underscore the neurobiological relevance of SOREMPs in NT1 patients’ treatment response.
Clinical and polysomnographic profiles: REM sleep dysregulation as a core endophenotype
Demographic and baseline symptoms (ESS scores, hallucination/sleep paralysis) did not distinguish between groups, highlighting that treatment response heterogeneity transcends clinical demographics. NT1 patients with two or more SOREMPs after treatment exhibited persistently shortened pre/post-treatment REM sleep latency and reduced post-treatment wakefulness, aligning with the hypothesis that SOREMPs reflect dysregulated REM sleep gating-likely due to hypocretin loss disrupting the orexinergic control of REM-on/REM-off neuronal networks [28]. SOREMPs reflect rapid transitions into REM sleep, and a higher number of such episodes inherently implies a greater likelihood of shorter REM latency. However, our findings extend beyond this inherent association by identifying neurobiological correlates that modulate this relationship. Specifically, the observed differences in hypothalamic functional connectivity (e.g., enhanced left medial hypothalamus-inferior parietal lobule connectivity in the low SOREMPs group and altered lateral hypothalamus-insula connectivity in the high SOREMPs group) provide mechanistic insights that transcend mere probability. These connectivity patterns, alongside the mediation of depressive symptoms through hypothalamic-parietal/insular pathways, suggest that neural circuit function actively regulates the expression of SOREMPs and REM latency, even when accounting for their inherent statistical linkage. In summary, while the shorter REM latency in the high SOREMPs group is partially consistent with probabilistic expectations, our neuroimaging and mediation analyses highlight that this phenomenon is not purely stochastic but is shaped by underlying hypothalamic network dynamics. This distinction reinforces the value of integrating functional connectivity measures with traditional sleep metrics to disentangle inherent associations from modifiable neurobiological mechanisms in NT1 treatment response. These findings reinforce SOREMP as a biomarker of residual REM sleep instability, even under pharmacological intervention, and suggest that patients with two or more SOREMPs may require more targeted modulation of REM regulatory circuits.
Hypothalamic functional connectivity: dissecting treatment-responsive neural networks
The observed FC differences map to key nodes in the sleep-wake regulatory axis. Patients in low post-treatment SOREMPs group showed enhanced connectivity between the right medial hypothalamus (rMH) and thalamus, a pathway critical for relaying wake-promoting signals [30, 31], and enhanced connectivity between the left medial hypothalamus (lMH) and left inferior parietal lobule (IPL), a region involved in attention and sensorimotor integration [32]. This may reflect compensatory reinforcement of hypothalamic-driven arousal pathways, facilitating better sleep latency control. Conversely, reduced left lateral hypothalamus (lLH)-right insula/anterior cingulate cortex (ACC) connectivity in low post-treatment SOREMPs group patients is notable, as the insula-ACC network mediates interoceptive awareness and emotional regulation [33]. Diminished connectivity here might signify improved dissociation of emotional arousal from sleep regulatory centers, reducing maladaptive hyperarousal that perpetuates excessive daytime sleepiness (EDS).
In contrast, patients in high post-treatment SOREMPs group likely retains disrupted hypothalamic-parietal/insular circuits, impairing integration of homeostatic sleep drive with cognitive-emotional processing. These findings align with resting-state fMRI studies in NT1, which report hypothalamic hypo-connectivity with prefrontal and parietal regions linked to EDS severity [34, 35], suggesting that treatment-responsive patients may exhibit restored connectivity in these sleep-attention networks.
Predictors of post-treatment sleep latency: age, baseline physiology, and connectivity
The regression analysis identified age, pretreatment MSLT mean sleep latency, and left MH-left IPL FC as key predictors for post-treatment MSLT mean sleep latency, offering mechanistic insights:
Age-related effects (β = −0.256) may reflect declining neuroplasticity or hypocretinergic reserve, consistent with longitudinal data showing age-dependent worsening of narcolepsy symptoms [28].
Pretreatment sleep latency (β = 0.392) as a robust predictor highlights the importance of baseline sleep-wake dysregulation in treatment outcomes, suggesting that patients with more severe baseline EDS may have limited response to medication therapies.
Left MH-left IPL connectivity (β = 0.272) underscores the role of hypothalamic-parietal circuits in sleep latency regulation. The IPL’s involvement in sensorimotor integration and the MH’s role in sleep homeostasis imply that stronger connectivity here facilitates better sleep-wake boundary maintenance, possibly by enhancing cortical arousal regulation.
Mediation analysis: depressive symptoms as a driver of connectivity-mediated treatment response (extended with pharmacological considerations)
The observed mediation via hypothalamic-parietal/insular pathways warrants contextualization within the specific pharmacological intervention used-clomipramine, a tricyclic antidepressant (TCA) with dual serotonin-norepinephrine reuptake inhibition. TCAs are known to modulate REM sleep architecture, primarily by suppressing REM sleep and prolonging REM latency through antagonism of histamine and muscarinic receptors, as well as enhancing monoaminergic neurotransmission [36]. These effects may intersect with the study’s findings in two critical ways:
Direct pharmacological impact on REM sleep and SOREMPs
Clomipramine’s REM-suppressive properties could theoretically reduce SOREMPs by directly stabilizing REM gating independent of depressive symptom improvement. However, our mediation analysis revealed that the relationship between depression and SOREMPs was fully mediated by functional connectivity, suggesting that even with a REM-modulating drug, the effect of depression on treatment response is channeled through neural circuit alterations rather than purely pharmacological REM suppression. This implies that residual SOREMPs in the two or more SOREMPs group may reflect insufficient modulation of hypothalamic-parietal/insular connectivity despite pharmacological REM suppression, highlighting a biological substrate resistant to TCA-induced sleep architecture changes.
Interaction between clomipramine’s antidepressant action and connectivity
As a serotonin-norepinephrine reuptake inhibitor, clomipramine may enhance synaptic plasticity in limbic-hypothalamic circuits, potentially normalizing the left LH-right insula hyperconnectivity associated with depression. The insula, a key node in the salience network, is rich in serotonin receptors (5-HT2A/C), and TCA-mediated serotonin elevation could dampen insular hyperactivity, reducing its aberrant coupling with the lateral hypothalamus [37]. Conversely, the right LH-left IPL pathway, where depression was associated with reduced connectivity, may be less sensitive to clomipramine’s monoaminergic effects, as the IPL’s role in attention-sleep integration relies more on glutamatergic or orexinergic signaling [38]. This differential pharmacologic impact may explain why only these two pathways emerged as mediators, underscoring the need to interpret connectivity findings in the context of specific drug mechanisms.
Clinical nuance: separating drug effects from disease biology
While clomipramine was likely prescribed to address comorbid depression, its off-target effects on sleep (e.g., increased wake after sleep onset, as seen in the post-treatment wakefulness index difference) must be considered. The two or more SOREMPs group exhibited lower post-treatment wakefulness, which could reflect either inadequate depression control (leading to persistent hyperarousal) or paradoxical REM dysregulation despite TCA use. This underscores the complexity of disentangling antidepressant efficacy, sleep pharmacology, and intrinsic neural connectivity in treatment response, particularly in a population with preexisting hypocretin dysfunction [13].
Clinical and translational implications
The identified FC pathways represent novel targets for precision medicine. For instance, non-invasive brain stimulation of the parietal lobule to enhance LH-IPL connectivity or insular modulation to reduce hyperarousal may augment pharmacological interventions. Additionally, integrating FC metrics into clinical assessments could predict treatment response, enabling stratified care for patients at risk of residual SOREMPs.
Limitations include cross-sectional design and lack of drug-specific analysis, as different medications (e.g., modafinil vs. pitolisant) may differentially affect connectivity. Future longitudinal studies with multimodal imaging and treatment-specific subgroups are needed to clarify causal relationships and optimize intervention strategies. The current analysis does not differentiate between clomipramine’s antidepressant effects, its REM-modulating properties, and its impact on hypothalamic connectivity. For example, while TCAs are less commonly used for narcolepsy compared to selective serotonin reuptake inhibitors (SSRIs) or sodium oxybate, their cholinergic antagonism may uniquely affect hypothalamic nuclei involved in REM regulation (e.g., sub-laterodorsal nucleus). Future studies should: compare connectivity changes across different antidepressant classes (e.g., TCAs vs. SSRIs vs. SNRIs) to isolate monoaminergic vs. cholinergic mechanisms; use longitudinal design to assess whether clomipramine-induced depression remission precedes connectivity changes or vice versa; incorporate polysomnographic measures of REM density/intensity to disentangle drug-induced REM suppression from connectivity-mediated SOREMP regulation. Dedicated self-report measures of cataplexy (e.g., frequency, triggers, severity) could provide additional granularity. Future studies incorporating such tools may further clarify the interplay between cataplexy experiences, neurobiological mechanisms, and treatment outcomes in NT1. Even with careful monitoring, the complexity of cataplexy triggers makes complete prevention challenging. Future studies could integrate physiological markers (e.g., electromyography during fMRI) to more sensitively detect subclinical cataplexy, enhancing our understanding of how such episodes interact with hypothalamic connectivity and treatment responses in NT1.
Conclusion
Results from this study extend our understanding of NT1 treatment response by linking objective SOREMPs to hypothalamic network integrity and depressive symptoms. The persistence of high SOREMPs may reflect unresolved REM sleep dyscontrol, which could be targeted through therapies modulating hypothalamic-parietal/insular connectivity. The identification of FC as a mediator between depression symptoms and SOREMPs suggests that addressing both neural circuit dysfunction and mood symptoms may be critical for optimizing treatment outcomes in NT1. These findings integrate objective sleep metrics, neuroimaging, and clinical phenotypes, providing a framework for personalized management of NT1. In summary, the actionable traits are clinically accessible measures of depressive symptoms and quantifiable hypothalamic-parietal/insular FC patterns, while personalized therapy could involve stratified depression management and targeted neuromodulation of these pathways.
Data availability
Anonymized data not published within this article will be made available by request from any qualified investigator.
References
Latorre D, Sallusto F. Narcolepsy: a model interaction between immune system, nervous system, and sleep-wake regulation. Semin Immunopathol. 2022;44:611–23.
Biscarini F, Barateau L, Pizza F, Plazzi G. Narcolepsy and rapid eye movement sleep. J Sleep Res. 2025;34:e14277.
Pizza F, Barateau L, Dauvilliers Y. The orexin story, sleep and sleep disturbances. J Sleep Res. 2022;31:e13665.
Thorpy MJ, Siegel JM, Dauvilliers Y. REM sleep in narcolepsy. Sleep Med Rev. 2024;77:101976.
Thorpy MJ, Hiller G. The medical and economic burden of narcolepsy: implications for managed care. Am health drug benefits. 2017;10:233–41.
Kukkonen JP, Jacobson LH. International union of basic and clinical pharmacology CXIV: orexin receptor function, nomenclature and pharmacology. Pharmacol Rev. 2024;76:625–88.
Ishikawa T, Kurimoto E, Joyal AA, Koike T, Kimura H, Scammell TE. An orexin agonist promotes wakefulness and inhibits cataplexy through distinct brain regions. Curr Biol. 2025;35:2088–99.e4.
Evans R, Kimura H, Alexander R, Davies CH, Faessel H, Hartman DS, et al. Orexin 2 receptor-selective agonist danavorexton improves narcolepsy phenotype in a mouse model and in human patients. Proc Natl Acad Sci Usa. 2022;119:e2207531119.
Dauvilliers Y, Mignot E, Del Río Villegas R. Oral orexin receptor 2 agonist in narcolepsy type 1. N Engl J Med. 2023;389:309–21.
Chen KS, Xu M, Zhang Z, Chang WC, Gaj T, Schaffer DV, et al. A hypothalamic switch for REM and non-REM sleep. Neuron. 2018;97:1168–76.e4.
Singh R, Biswas DA. Physiological role of orexin/hypocretin in the human body in motivated behavior: a comprehensive review. Cureus. 2023;15:e34009.
Hong J, Lozano DE, Beier KT. Prefrontal cortical regulation of REM sleep. Nat Neurosci. 2023;26:1820–32.
Adamantidis AR, de Lecea L. Sleep and the hypothalamus. Science. 2023;382:405–12.
Ballotta D, Talami F, Pizza F, Vaudano AE, Benuzzi F, Plazzi G, et al. Hypothalamus and amygdala functional connectivity at rest in narcolepsy type 1. NeuroImage Clin. 2021;31:102748.
Abenza-Abildua MJ, Suárez-Gisbert E, Lores-Gutiérrez V, Algarra-Lucas C, Gómez-Aceña Á, Navacerrada-Barrero FJ, et al. Anxiety and depression in patients with narcolepsy. J Sleep Res. 2023;32:e13812.
Wang Y, Liu L, Gu JH, Wang CN, Guan W, Liu Y, et al. Salt-inducible kinase 1-CREB-regulated transcription coactivator 1 signalling in the paraventricular nucleus of the hypothalamus plays a role in depression by regulating the hypothalamic-pituitary-adrenal axis. Mol psychiatry. 2024;29:1660–70.
Zhang S, Zhornitsky S, Wang W, Le TM. Resting state hypothalamic and dorsomedial prefrontal cortical connectivity of the periaqueductal gray in cocaine addiction. Addiction Biol. 2021;26:e12989.
Liu Y, Li A, Bair-Marshall C, Xu H, Jee HJ, Zhu E, et al. Oxytocin promotes prefrontal population activity via the PVN-PFC pathway to regulate pain. Neuron. 2023;111:1795–811.e7.
Zhu Z, Miao L, Li K, Ma Q, Pan L, Shen C, et al. A hypothalamic-amygdala circuit underlying sexually dimorphic aggression. Neuron. 2024;112:3176–91.e7.
Medicine AAoS The International Classification of Sleep Disorders. 3rd edn.. Darien, IL: American Academy of Sleep Medicine; 2014.
Fulong X, Spruyt K, Xiaosong D, Zhaolong C, Jun Z, Fang H. Morphological and age-related changes in the narcolepsy brain. Cereb cortex. 2021;31:5460–9.
Zhao Z, Ding N, Song S, Liu Y, Wen D. Association between depression and overweight in Chinese adolescents: a cross-sectional study. BMJ open. 2019;9:e024177.
Vilagut G, Forero CG, Barbaglia G, Alonso J. Screening for depression in the general population with the center for epidemiologic studies depression (CES-D): a systematic review with meta-analysis. PLoS one. 2016;11:e0155431.
Lu CF, Jia CX, Xu AQ, Dai AY, Qin P. Psychometric characteristics of Chinese version of barratt impulsiveness scale-11 in suicides and living controls of rural China. Omega. 2012;66:215–29.
Baroncini M, Jissendi P, Balland E, Besson P, Pruvo JP, Francke JP, et al. MRI atlas of the human hypothalamus. NeuroImage. 2012;59:168–80.
Tillikainen L, Salli E, Korvenoja A, Aronen HJ. A cluster mass permutation test with contextual enhancement for fMRI activation detection. NeuroImage. 2006;32:654–64.
Wu Y, Kong L, Yang A, Xin K, Lu Y, Yan X, et al. Gray matter volume reduction in orbitofrontal cortex correlated with plasma glial cell line-derived neurotrophic factor (GDNF) levels within major depressive disorder. NeuroImage Clin. 2023;37:103341.
Barateau L, Pizza F, Chenini S, Peter-Derex L, Dauvilliers Y. Narcolepsies, update in 2023. Rev neurologique. 2023;179:727–40.
Overeem S, Mignot E, van, Lammers DijkJG. GJ. Narcolepsy: clinical features, new pathophysiologic insights, and future perspectives. J Clin Neurophysiol : Publ Am Electroencephalographic Soc. 2001;18:78–105.
Yokoi R, Okabe M, Matsuda N, Odawara A, Karashima A, Suzuki I. Impact of sleep-wake-associated neuromodulators and repetitive low-frequency stimulation on human iPSC-Derived Neurons. Front Neurosci. 2019;13:554.
Falup-Pecurariu C, Diaconu Ș, Țînț D, Falup-Pecurariu O. Neurobiology of sleep (Review). Exp therapeutic Med. 2021;21:272.
Jiang JW, Narasimhan S, Johnson GW, González HFJ, Doss DJ, Shless JS, et al. Abnormal functional connectivity of the posterior hypothalamus and other arousal regions in surgical temporal lobe epilepsy. J Neurosurg. 2023;139:640–50.
Ji T, Li X, Xu Z, Zhao J, Wang G, Li Y, et al. Aberrant resting-state effective connectivity between the insula and other regions of the whole brain in children with obstructive sleep apnea. J Sleep Res. 2025;17:e70015.
Mao T, Guo B, Quan P, Deng Y, Chai Y, Xu J, et al. Morning resting hypothalamus-dorsal striatum connectivity predicts individual differences in diurnal sleepiness accumulation. NeuroImage. 2024;299:120833.
Qi J, Hu WL. Altered hypothalamic functional connectivity after partial sleep deprivation in young and elderly adults. Behavioural brain Res. 2025;488:115587.
Winkler A, Rheker J, Doering BK, Rief W. Conditioning of amitriptyline-induced REM sleep suppression in healthy participants: a randomized controlled trial. Psychophysiology. 2016;53:1560–6.
McDonald AJ. Functional neuroanatomy of monoaminergic systems in the basolateral nuclear complex of the amygdala: Neuronal targets, receptors, and circuits. J Neurosci Res. 2023;101:1409–32.
Li Y, Ma M, Shao Y, Wang W. Enhanced effective connectivity from the middle frontal gyrus to the parietal lobe is associated with impaired mental rotation after total sleep deprivation: an electroencephalogram study. Front Neurosci. 2022;16:910618.
Acknowledgements
We thank all the patients for their collaboration.
Funding
This work was supported by the National Natural Science Foundation of China (82470087, 82070091 and 82341243).
Author information
Authors and Affiliations
Contributions
Mengmeng Wang collected data, Haodong Zhang analyzed PSG data, Xiaosong Dong and Fang Han helped this study, Fulong Xiao designed and helped write the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Wang, M., Zhang, H., Dong, X. et al. Hypothalamic functional connectivity, depressive symptoms, and post-treatment SOREMPs in narcolepsy type 1: links to sleep latency and mediation mechanisms. Transl Psychiatry 15, 484 (2025). https://doi.org/10.1038/s41398-025-03670-3
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41398-025-03670-3
