
Abstract
In 2022, Hanahan integrated polymorphic microbiomes to the hallmarks of cancer, resulting in 14 overarching features that are considered fundamental to initiation and progression of cancers. It is well acknowledged that genomic instability/genetic alterations together with tumor-associated inflammation are so called “enabling hallmarks” as they drive the acquisition of the other traits. The microbiome is a key component of the inflammatory tumor stroma. Pancreatic ductal adenocarcinoma (PDAC) in particular is characterized by a pronounced stromal compartment whose role in the acquisition of malignant properties is well documented. Recent studies indicate massive alterations of the microbiome in PDAC tissues compared to healthy pancreas or precursor lesions. However, the mechanistic role of the PDAC-associated microbiome, its influence on the hallmarks of cancer, and how this relates to PDAC malignancy remain poorly understood. This raises the question of whether the tumor-associated microbiome through its direct influence on PDAC cells, their precursors, and the surrounding non-neoplastic cells promotes the acquisition of other hallmarks that drive PDAC development and progression. This perspective article outlines the current knowledge of the impact of the PDAC-associated microbiome on the hallmarks of cancer in PDAC. These current findings support the altered microbiome as a third enabling hallmark of PDAC and emphasize that further mechanistic studies are urgently needed to further substantiate its fundamental importance for this tumor entity. This knowledge will provide the basis for clinical translation to develop more effective therapeutic approaches for PDAC.

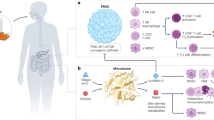
The intratumoral microbiome in PDAC exhibits numerous interactions with the hallmarks of cancer. Hallmarks indicated in blue have demonstrated interactions with the microbiome, while others still remain underexplored. These extensive interactions substantiate the role of the intratumoral microbiome in PDAC as an enabling hallmark, underlining its potential as a therapeutic target. Partially created with biorender.com.
Similar content being viewed by others


Introduction
Pancreatic ductal adenocarcinoma (PDAC) is still one of the most fatal cancers due to its aggressive nature and high therapy resistance. Besides characteristic genetic alterations (e.g., in KRAS), PDAC is marked by a pronounced tumor microenvironment (TME) composed of inflammatory and immune cells as well as the microbiome. The latter is altered in PDAC compared to the physiological pancreas, and is associated with different outcomes, depending on its composition [1]. Given these associations, the microbiome may be regarded as an enabling hallmark of PDAC [2]. This perspective article briefly outlines the current knowledge on how the tumor-associated microbiome impacts the other hallmarks of cancer in PDAC, referring to excellent reviews and some original articles. Finally, we provide a perspective on the further steps required to emphasize this role, providing the basis for improved therapeutic concepts. When mentioned in the text, the hallmarks of cancer are highlighted in bold.
The intratumoral microbiomes’ impact on hallmarks of cancer in PDAC
The induction of genomic instability, in conjunction with mutations, constitutes one carcinogenic mechanism of the intratumoral microbiome. Some bacteria synthesize substances capable of inducing DNA damage, like Escherichia coli (E. coli) colibactin [3]. The release of these substances can compromise the integrity of host DNA, which has been demonstrated to increase the risk of oncogenic changes in the respective colonized tissue [4]. For instance, the abundance of anaerobic Fusobacterium nucleatum (F. nucleatum), present in the oral cavity under physiological conditions, is highly increased in PDAC tissues, where it can induce DNA double strand breaks and increase the mutational burden [1, 5]. Furthermore, in other cancers like colorectal cancer (CRC), infection with F. nucleatum results in impairment of DNA repair mechanisms, leading to additional destabilization of DNA and increased mutation rates [6]. In a CRC mouse model, the bacterium promotes E-cadherin/β-catenin activation to upregulate checkpoint kinase 2, which induced additional DNA damage [7]. Since the various effects of F. nucleatum on PDAC are similar to its effects on CRC, it is reasonable to assume that the bacterium influences the genomic stability in PDAC as well [5, 8,9,10,11,12]. Studies investigating specifically this hallmark in PDAC in response to F. nucleatum infection have to be performed to confirm the assumption.
In addition to their impact on genomic instability, intratumoral bacteria can influence sustained proliferative signaling in tumor and stroma cells, as well as inflammation through induction of cytokine secretion [8]. F. nucleatum infection leads to increased IL-8, GM-CSF, and MIP-3a secretion, as well as stimulation of the JAK/STAT and MAP kinase pathways, resulting in increased proliferation and migration [13]. Furthermore, mutations caused by F. nucleatum stimulate the Ku70/p53 signaling pathway, and consequently the proliferative activity of infected tumor cells [5]. Besides pro-proliferative effects of F. nucleatum, bacterial infection with e.g., Helicobacter pylori (H. pylori) can lead to increased levels of IL-8 and VEGF, as well as an increase of the transcription factor NF-κB, further impacting cell proliferation and inflammation [14]. Similarly, Porphyromonas gingivalis (P. gingivalis), enriched in PDAC tissues compared to physiological controls, contributes to inflammation through activity of gingipain proteases [15]. Bacteria-induced inflammation is not limited to particular species; rather, it is induced by most bacteria bearing pathogen-associated molecular patterns. For instance, lipopolysaccharide (LPS) activates the PI3K-Akt-mTOR pathway in PDAC cells and activates the inflammasome [16, 17]. In KRAS-mutant mice, LPS stimulation promotes NF-κB-mediated development of pancreatic intraepithelial neoplasia, precursor lesions of PDAC [18]. Another study demonstrated that sterile bile juices inhibited PDAC cell growth. However, addition of bacteria to the bile juice restored cell growth, while bacterial conditioning did not [19]. In addition to these cancer promoting effects, some bacteria can have beneficial effects for patients. Pseudomonas aeruginosa produces mannose-sensitive hemagglutinin lectin, which leads to cell cycle arrest in PDAC, but not normal pancreatic epithelial cells [20]. Overall, these findings indicate that microbes can impact tumor cell growth and inflammation in a direct and paracrine fashion, as well as in a promoting and inhibiting manner, in dependence on the species.
The TME, as well as the tumor cells, also impact phenotype and activity of immune cells. Cancer cells evolve mechanisms to evade destruction by immune cells and promote an immunosuppressive and pro-tumorigenic environment, a process mediated by the intratumoral microbiome [1]. Returning to F. nucleatum and P. gingivalis, infection of PDAC cells in vitro or in vivo results in bacterial invasion to the gut and tumor, inducing a pro-inflammatory TME enriched with neutrophils, advancing further progression by chemokine release [8, 21]. Moreover, microbial diversity within PDAC tissues correlates with immune cell profiles. Tumors with more diverse microbiomes are associated with greater infiltration of activated cytotoxic T lymphocytes (CTL) and improved survival after tumor resection. Accordingly, fecal microbiome transfers (FMT) from human long-term survivors in PDAC mouse models enhance CTL invasion and pro-inflammatory cytokine expression, while microbiomes from short-term survivors promote regulatory T helper cells and myeloid-derived suppressor cells [22]. Gut microbiome depletion in PDAC-bearing mice increases effector T cell infiltration and IFNγ production, while the number of IL-17a+ and IL-10 + T cells decreases, underscoring the immunosuppressive function of the microbiome. Ablation also led to polarization of macrophages to an M1-like phenotype, resulting in slower tumor growth, driven primarily by T cells [1, 22].
Cancer cells not only evolve strategies to evade immune cells, but also to resist cell death [2]. It was shown that intracellular P. gingivalis protects PDAC cells from reactive oxygen species-induced cell death under nutrient stress conditions [21]. Additionally, this hallmark represents a key mechanism of therapy resistance [2]. Gemcitabine is a cornerstone chemotherapeutic agent in PDAC treatment, inducing cell death. However, the efficacy of Gemcitabine in PDAC patients is often limited due to the tumor’s intrinsic and acquired resistance mechanisms, potentially influenced by the intratumoral microbiome [23]. Geller et al. demonstrated that Gammaproteobacteria induce Gemcitabine resistance, converting Gemcitabine into its inactive form by expression of a long isoform of cytidine deaminase [24]. This resistance was reversed by the antibiotic Ciprofloxacin, underlining the influence of the bacteria. Even though these findings were obtained in a colon cancer mouse model, the study also revealed that 76% of analyzed PDAC tissues contained predominantly Gammaproteobacteria, supporting a role of these intratumoral bacteria in PDAC drug resistance. Indeed, antibiotic treatment prior to gemcitabine therapy, but not other cytostatic drugs, led to improved survival outcomes for PDAC patients [25]. This connection was further expanded to the evolution of therapy resistance, when E. coli rapidly adapted to gemcitabine treatment in a PDAC spheroid coculture model [26].
Besides the pronounced TME, PDAC is characterized by a high tumor cell plasticity contributing to tumor heterogeneity [27]. Tumor cell plasticity is mediated by Epithelial-to-Mesenchymal-Transition (EMT), a key mechanism to confer a motile and invasive phenotype, thereby promoting metastasis. Cancer stem cells (CSCs) further contribute to tumor cell plasticity and are regarded as essential for tumor initiation and progression [28]. It seems plausible that cancer cell plasticity in PDAC is influenced not only by CSCs and EMT, but also by additional factors such as stromal and immune cells and the distinct composition of the tumor microbiome. In different cancer entities, e.g., colorectal, bladder or breast cancer, a crucial association between microorganisms, EMT and CSCs was demonstrated [29]. It was reported that H. pylori infection induces EMT-like changes and high expression of the gastric CSC marker CD44 in vitro, leading to increased cell migration, invasion and tumor sphere formation [30]. Many PDAC tissues show presence of H. pylori, however the bacterium has not been clearly associated with patient outcome in this cancer entity [31]. Further knowledge about the mechanistic influence of the microbiome on PDAC progression is limited. Nevertheless, it was already shown that a tumor-associated microbiome can induce signaling pathways that cause activation of different EMT transcription factors and thus promote plasticity in PDAC [32]. It is reasonable to assume that the microbiome also influences CSCs, further impacting cancer cell plasticity. Regarding metastasis, Jeong et al. examined the microbiome detectable in extracellular vesicles derived from PDAC and corresponding non-cancerous pancreatic tissues [33]. They identified Tepidimonas enriched in tumor tissue and describe a positive correlation of lymph node metastasis with increased prevalence of Comamonas and Turicibacter in the tumor tissue. Treatment of PDAC cell lines with Tepidimonas-conditioned medium increased migration and upregulated expression of the EMT transcription factors SNAIL and TWIST. In another study, several bacterial species were enriched in the bile of patients with lymph node metastasis compared to non-metastasized PDAC [34]. Still, little mechanistic data exists elucidating the pathophysiological effects of intratumoral infection with individual bacterial species on invasion and potential metastasis capacities of PDAC cells. Udayasuryan et al. infected PDAC cells and non-malignant pancreatic epithelial cells with F. nucleatum, leading to elevated levels of cytokines in both cancerous and non-cancerous cells [8]. Interestingly, autocrine and paracrine exposure to these cytokines resulted in increased migration and invasion of malignant but not non-malignant pancreatic epithelial cells. Some aspects of this study were independently confirmed, as infection with F. nucleatum, but not with P. gingivalis, increased CXCL1 and IL-8 secretion by PDAC cell lines, resulting in increased migration and invasion in vitro [10]. In patient-derived tumor samples, CXCL1 expression was significantly higher in the F. nucleatum infected group, underlining a potential relevance of the in vitro findings for the clinical context. In a mouse model of peritoneal metastasis, bile juice pre-incubated with live Enterococcus fecalis (E. faecalis) or Streptococcus oralis markedly enhanced the growth of peritoneal metastasis, underscoring the promoting effect of certain bacteria on malignant phenotypes in PDAC [19].
Deregulation of cellular metabolism is another hallmark of cancer observed also in other pathological conditions such as obesity, metabolic syndrome and diabetes, all being associated with the incidence of PDAC [35]. The mechanism that connects these diseases involve promotion of chronic inflammation, the influence of adipose tissue-derived soluble mediators on KRAS signaling, and the disruption of metabolic pathways [36, 37]. These interlinked metabolic pathways, that may be initiated by one of the conditions, might lead to a higher susceptibility towards PDAC development. Of note, an altered microbiome has been implicated in the above-mentioned metabolic diseases [38]. High-fat diets, which are characteristically associated with metabolic dysregulation, can lead to a compromised intestinal barrier, resulting in leakage of bile acids into the circulatory system and initiating systemic inflammation and tumorigenesis [39]. Other bacterial metabolites affected by both metabolic dysregulation and PDAC tumorigenesis are short-chain fatty acids (SCFAs). SCFAs can reduce cancer incidence by inhibiting cell growth and migration and inducing apoptosis. In PDAC, probiotics and butyrate producing bacteria are reduced [39]. Infection with H. pylori, enriched in PDAC tissues, also result in metabolic dysregulation, as the bacteria produce nitrosamines and other carcinogens that can potentially induce genomic instability [40]. One of the key repercussions of metabolic dysregulation, compromised intestinal permeability, can lead to direct interlinking of the gut microbiome to the PDAC associated microbiome. The translocation of opportunistic pathobionts like E. faecalis from the gut to PDAC tissues, can ensure a continuous channel of colonization, resulting in continued influence on the tumorigenic conditions in PDAC [41]. E. faecalis is known to polarize colon macrophages into a pro-inflammatory phenotype, leading to endogenous genomic instability and cellular transformation in IL10-/- mice [42]. Interestingly, Pushalkar et al. observed bacterial migration of E. faecalis and E. coli to the pancreas upon oral administration in wild type mice, pointing towards the possibility of a similar mechanism in PDAC [43].
Future directions
This perspective article highlights the extensive and intricate web of interactions between the tumor-associated microbiome and other hallmarks of cancer in PDAC (Table 1). As integral part of the TME, the intratumoral microbiome induces genetic instability, impacts proliferative signaling and confers resistance to cell death. Through mechanisms of inflammation, it further shapes the TME to support cancer cell survival and progression, while impairing immune function and enabling immune evasion. In addition, bacterial interactions and metabolites have been shown to alter the tumor cell metabolism with implications for PDAC progression. Cancer cell motility is impacted by intratumoral bacteria, promoting metastasis. There is growing evidence that the microbiome also promotes cancer cell plasticity, thereby contributing to tumor heterogeneity and aggressiveness of PDAC, although further research is needed to translate key findings from other cancer entities to PDAC.
While the influence of the microbiome on these distinct hallmarks is established in PDAC, the impact on the other hallmarks remains underexplored. Cellular senescence can promote the progression of cancer through a senescence-associated secretory phenotype. Having demonstrated the influence of the microbiome on PDAC cell growth, it can be speculated that certain bacteria have a growth-inhibiting impact by which malignant cells acquire a senescent/quiescent phenotype. The association between the intratumoral microbiome and angiogenesis in PDAC remains to be elucidated. However, secretion of cytokines like IL-8 in response to bacterial infection may link these hallmarks. Connections to epigenetic reprofiling, replicative immortality, and evasion of growth suppressors are not yet well understood.
In summary, the rapidly expanding body of evidence strongly supports the view that the polymorphic microbiome has to be regarded as an enabling hallmark of cancer in PDAC. To further substantiate the enabling role of the microbiome in this disease, a deeper understanding of the underlying mechanistic interactions and the identification of beneficial and detrimental microbial communities are mandatory to transfer this knowledge into effective therapeutic concepts that could improve prognosis of PDAC patients.
To date, no microbiome-targeted therapy has been implemented in the clinical management of PDAC. However, data from preclinical models and retrospective analyses indicates its potential for future improvements in the treatment of PDAC patients.
A straightforward approach involves antibiotic therapy, ablating large portions of the microbiome across multiple organs. Retrospective analyses revealed an increase of overall survival in PDAC patients receiving gemcitabine, but not other cytostatic drug treatment [25]. In a mouse model, antibiotic-mediated microbiome ablation enhanced T cell infiltration to the TME and resolved immune checkpoint blockades [43, 44]. However, broad ablation comes with adverse side effects for the patients’ overall health. Furthermore, the resulting reduction in intratumoral alpha diversity has been associated with worse patient outcomes, and the loss of beneficial regulatory bacteria allows for uncontrolled recolonization [22]. This challenge could be addressed through targeted recolonization after ablation. Riquelme et al. demonstrated that orthotopic PDAC-bearing mice significantly benefited from FMT from human long-term survivors [22]. Indeed, two ongoing phase I clinical trials currently investigate the effect of FMT on survival of patients with either resectable (ID: NCT04975217) or non-resectable (ID: NCT06393400) PDAC [45, 46].
Additionally, studies have investigated the therapeutic potential of specific probiotic bacteria, particularly lactic acid bacteria. Oral administration of Lactobacillus or Megasphaera reduced progression of xenograft and orthotopic PDAC or pancreatic tumors in mice, respectively [47, 48]. However, use of singe-species probiotics can reduce microbial diversity, induce dysbiosis, and pose a risk of infection in immunocompromised patients [49]. Further research is required to develop complex formulation of probiotic bacteria before progressing to clinical trials.
Beyond antibiotics, bacteriophages offer potential to specifically target oncobacteria in PDAC. In a recent study, the F. nucleatum targeting bacteriophage ØTCUFN3 inhibited tumor cell growth in vitro and reduced progression in xenograft models of CRC [50]. Although no such studies have been performed in PDAC yet, this strategy seems to be also applicable to PDAC based on the pro-tumoigenic role of F. nucleatum in this tumor entity [5, 8, 10].
As a final note, the rapid adaptability of the microbiome can drive resistance to therapy regimens, as demonstrated in the context of gemcitabine resistance [26]. Therefore, data on the effects of treatments on the tumor microbiome are needed. In cases where patients relapse after primary tumor resection and subsequent treatment, and resection of the recurrent tumor becomes clinically feasible in the future, the microbiome of both specimens should be analyzed. In addition, in animal models, the compositional changes of the intratumor microbiome should be investigated and considered.
The therapeutic potential of microbiome-targeting strategies in PDAC is considerable. Their clinical implementation requires comprehensive research that further unravels the interactions between the tumor microbiome and the other hallmarks of cancer and how these impact PDAC progression and therapy responses.
References
Cruz MS, Tintelnot J, Gagliani N. Roles of microbiota in pancreatic cancer development and treatment. Gut Microbes. 2024;16:2320280.
Hanahan D. Hallmarks of cancer: new dimensions. Cancer Discov. 2022;12:31–46.
Wilson MR, Jiang Y, Villalta PW, Stornetta A, Boudreau PD, Carrá A, et al. The human gut bacterial genotoxin colibactin alkylates DNA. Science. 2019;363:eaar7785.
González-Sánchez P, DeNicola GM. The microbiome(s) and cancer: know thy neighbor(s). J Pathol. 2021;254:332–43.
Geng F, Zhang Y, Lu Z, Zhang S, Pan Y. Fusobacterium nucleatum caused DNA damage and promoted cell proliferation by the Ku70/p53 pathway in oral cancer cells. DNA Cell Biol. 2020;39:144.
Udayasuryan B, Zhou Z, Ahmad RN, Sobol P, Deng C, Nguyen TTD, et al. Fusobacterium nucleatum infection modulates the transcriptome and epigenome of HCT116 colorectal cancer cells in an oxygen-dependent manner. Commun Biol. 2024;7:1–15.
Guo P, Tian Z, Kong X, Yang L, Shan X, Dong B, et al. FadA promotes DNA damage and progression of Fusobacterium nucleatum-induced colorectal cancer through up-regulation of chk2. J Exp Clin Cancer Res. 2020;39:202.
Udayasuryan B, Ahmad RN, Nguyen TTD, Umaña A, Roberts LDM, Sobol P, et al. Fusobacterium nucleatum induces proliferation and migration in pancreatic cancer cells through host autocrine and paracrine signaling. Sci Signal. 2022;15:eabn4948.
Casasanta MA, Yoo CC, Udayasuryan B, Sanders BE, Umanã A, Zhang Y, et al. Fusobacterium nucleatum host-cell binding and invasion induces IL-8 and CXCL1 secretion that drives colorectal cancer cell migration. Sci Signal. 2020;13:eaba9157.
Hayashi M, Ikenaga N, Nakata K, Luo H, Zhong PS, Date S, et al. Intratumor Fusobacterium nucleatum promotes the progression of pancreatic cancer via the CXCL1-CXCR2 axis. Cancer Sci. 2023;114:3666–78.
Yin H, Miao Z, Wang L, Su B, Liu C, Jin Y, et al. Fusobacterium nucleatum promotes liver metastasis in colorectal cancer by regulating the hepatic immune niche and altering gut microbiota. Aging. 2022;14:1941–58.
Yang Y, Weng W, Peng J, Hong L, Yang L, Toiyama Y, et al. Fusobacterium nucleatum Increases proliferation of colorectal cancer cells and tumor development in mice by activating toll-like receptor 4 signaling to nuclear factor-κb, and up-regulating expression of microRNA-21. Gastroenterology. 2017;152:851–866.e24.
Liva SM, Kahn MA, Dopp JM, De Vellis J. Signal transduction pathways induced by GM-CSF in microglia: significance in the control of proliferation. Glia. 1999;26:344–52.
Takayama S, Takahashi H, Matsuo Y, Okada Y, Manabe T. Effects of Helicobacter pylori infection on human pancreatic cancer cell line. Hepatogastroenterology. 2007;54:2387-91.
Ogrendik M. The association between oral anaerobic bacteria and pancreatic cancer. World J Oncol. 2023;14:174–7.
Massoumi RL, Teper Y, Ako S, Ye L, Wang E, Hines OJ, et al. Direct effects of lipopolysaccharide on human pancreatic cancer cells. Pancreas. 2021;50:524–8.
Sivam HGP, Chin BY, Gan SY, Ng JH, Gwenhure A, Chan EWL. Lipopolysaccharide (LPS) stimulation of Pancreatic Ductal Adenocarcinoma (PDAC) and macrophages activates the NLRP3 inflammasome that influences the levels of pro-inflammatory cytokines in a co-culture model. Cancer Biol Ther. 2023;24:2284857.
Daniluk J, Liu Y, Deng D, Chu J, Huang H, Gaiser S, et al. An NF-κB pathway-mediated positive feedback loop amplifies Ras activity to pathological levels in mice. J Clin Investig. 2012;122:1519–28.
Shrader HR, Miller AM, Tomanek-Chalkley A, McCarthy A, Coleman KL, Ear PH, et al. Effect of bacterial contamination in bile on pancreatic cancer cell survival. Surgery. 2021;169:617–22.
Cheng X, Wang B, Jin Z, Ma D, Yang W, Zhao R, et al. Pseudomonas aeruginosa-mannose-sensitive hemagglutinin inhibits pancreatic cancer cell proliferation and induces apoptosis via the EGFR pathway and caspase signaling. Oncotarget. 2016;7:77916.
Saba E, Farhat M, Daoud A, Khashan A, Forkush E, Menahem NH, et al. Oral bacteria accelerate pancreatic cancer development in mice. Gut. 2024;73:770–86.
Riquelme E, Zhang Y, Zhang L, Montiel M, Zoltan M, Dong W, et al. Tumor microbiome diversity and composition influence pancreatic cancer outcomes. Cell. 2019;178:795–806.e12.
Zeng S, Pöttler M, Lan B, Grützmann R, Pilarsky C, Yang H. Chemoresistance in pancreatic cancer. Int J Mol Sci. 2019;20:4504.
Geller LT, Barzily-Rokni M, Danino T, Jonas OH, Shental N, Nejman D, et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science. 2017;357:1156–60.
Fulop DJ, Zylberberg HM, Wu YL, Aronson A, Labiner AJ, Wisnivesky J, et al. Association of antibiotic receipt with survival among patients with metastatic pancreatic ductal adenocarcinoma receiving chemotherapy. JAMA Netw Open. 2023;6:e234254–e234254.
Sayin S, Rosener B, Li CG, Ho B, Ponomarova O, Ward DV, et al. Evolved bacterial resistance to the chemotherapy gemcitabine modulates its efficacy in co-cultured cancer cells. Elife. 2023;12:e83140.
Aldag L, Beckinger S, Daunke T, Philipp LM, Surrow A, Yesilyurt UU, et al. The heterogeneity of the tumor microenvironment as essential determinant of development, progression, and therapy response of pancreatic cancer. Cancers. 2021;13:4932.
Philipp LM, Yesilyurt UU, Surrow A, Künstner A, Mehdorn AS, Hauser C, et al. Epithelial and mesenchymal-like pancreatic cancer cells exhibit different stem cell phenotypes associated with different metastatic propensities. Cancers. 2024;16:686.
Cao Y, Xia H, Tan X, Shi C, Ma Y, Meng D, et al. Intratumoural microbiota: a new frontier in cancer development and therapy. Signal Transduct Target Ther. 2024;9:1–24.
Bessède E, Staedel C, Acuña Amador LA, Nguyen PH, Chambonnier L, Hatakeyama M, et al. Helicobacter pylori generates cells with cancer stem cell properties via epithelial-mesenchymal transition-like changes. Oncogene. 2014;33:4123–31.
Kunovsky L, Dite P, Jabandziev P, Dolina J, Vaculova J, Blaho M, et al. Helicobacter pylori infection and other bacteria in pancreatic cancer and autoimmune pancreatitis. World J Gastrointest Oncol. 2021;13:835.
Basu M, Philipp LM, Baines JF, Sebens S. The microbiome tumor axis: how the microbiome could contribute to clonal heterogeneity and disease outcome in pancreatic cancer. Front Oncol. 2021;11:740606.
Jeong JY, Kim TB, Kim J, Choi HW, Kim EJ, Yoo HJ, et al. Diversity in the extracellular vesicle-derived microbiome of tissues according to tumor progression in pancreatic cancer. Cancers. 2020;12:1–15.
Kirishima M, Yokoyama S, Matsuo K, Hamada T, Shimokawa M, Akahane T, et al. Gallbladder microbiota composition is associated with pancreaticobiliary and gallbladder cancer prognosis. BMC Microbiol. 2022;22:1–12.
Chandana SR, Woods L, Maxwell F, Gandolfo R, Bekaii-Saab T. Risk factors for early-onset pancreatic ductal adenocarcinoma: a systematic literature review. Eur J Cancer. 2024;198:113471.
Fonteneau G, Redding A, Hoag-Lee H, Sim ES, Heinrich S, Gaida MM, et al. Stress granules determine the development of obesity-associated pancreatic cancer. Cancer Discov. 2022;12:1984–2005.
Srilatha M, Malla R, Adem MP, Foote JB, Nagaraju GP. Obesity associated pancreatic ductal adenocarcinoma: therapeutic challenges. Semin Cancer Biol. 2023;97:12–20.
Sasidharan Pillai S, Gagnon CA, Foster C, Ashraf AP. Exploring the gut microbiota: key insights into its role in obesity, metabolic syndrome, and type 2 diabetes. J Clin Endocrinol Metab. 2024;109:2709–19.
Guo X, Wang P, Li Y, Chang Y, Wang X. Microbiomes in pancreatic cancer can be an accomplice or a weapon. Crit Rev Oncol Hematol. 2024;194:104262.
Rabelo-Gonçalves EM, Roesler BM, Zeitune JM. Extragastric manifestations of Helicobacter pylori infection: possible role of bacterium in liver and pancreas diseases. World J Hepatol. 2015;7:2968–79.
Merali N, Chouari T, Kayani K, Rayner CJ, Jiménez JI, Krell J, et al. A comprehensive review of the current and future role of the microbiome in pancreatic ductal adenocarcinoma. Cancers. 2022;14:1020.
Wang X, Yang Y, Huycke MM. Commensal bacteria drive endogenous transformation and tumour stem cell marker expression through a bystander effect. Gut. 2015;64:459–68.
Pushalkar S, Hundeyin M, Daley D, Zambirinis CP, Kurz E, Mishra A, et al. The pancreatic cancer microbiome promotes oncogenesis by induction of innate and adaptive immune suppression. Cancer Discov. 2018;8:403–16.
Mohindroo C, Hasanov M, Rogers JE, Dong W, Prakash LR, Baydogan S, et al. Antibiotic use influences outcomes in advanced pancreatic adenocarcinoma patients. Cancer Med. 2021;10:5041.
Lehnan J. An open-label, single-arm, phase 1 study of the combination of FMT and Gemcitabine with Nab-paclitaxel as first-line therapy in patients with advanced pancreatic ductal adenocarcinoma. 2025. https://clinicaltrials.gov/study/NCT06393400.
McAllister F. Pilot study using fecal microbial transplants in patients with pancreatic cancer. 2021. https://www.clinicaltrials.gov/study/NCT04975217.
Huang Y, Zhu N, Zheng X, Liu Y, Lu H, Yin X, et al. Intratumor microbiome analysis identifies positive association between megasphaera and survival of Chinese patients with pancreatic ductal adenocarcinomas. Front Immunol. 2022;13:785422.
Chen SM, Hsu LJ, Lee HL, Lin CP, Huang SW, Lai CJL, et al. Lactobacillus attenuate the progression of pancreatic cancer promoted by porphyromonas gingivalis in K-rasG12D transgenic mice. Cancers. 2020;12:3522.
Zhang Z, Tang D. The huge clinical potential of microbiota in the treatment of pancreatic cancer: the next frontier. Biochim Biophys Acta Rev Cancer. 2022;1877:188733.
Lam HYP, Lai MJ, Wang PC, Wu WJ, Chen LK, Fan HW, et al. A novel bacteriophage with the potential to inhibit Fusobacterium nucleatum-induced proliferation of colorectal cancer cells. Antibiotics. 2025;14:45.
Acknowledgements
This project and its publication were supported by the Research and Training Group 2501 on Translational Evolutionary Research (RTG 2501 TransEvo) funded by the Deutsche Forschungsgemeinschaft. AMW was funded by the Clinician Scientist in Evolutionary Medicine Programme (by Deutsche Forschungsgemeinschaft (DFG, German Research Foundation)—Projektnummer 413490537) and by intramural funding of the Medical Faculty of Christian-Albrecht-University, Kiel. Large language models were used to improve grammar and language. The resulting content was reviewed and edited; the authors take full responsibility for the content of this article.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
Conceptualization: M.C. Investigation, writing (original draft and review, and editing): All authors. Visualization: M.C. (graphical abstract and table). Writing—abstract, future directions: S.Se. and M.C. Supervision: S.Se. and A.M.W.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Carstensen, M., Philipp, LM., Basu, M. et al. Intratumoral microbiome and pancreatic cancer: an enabling hallmark and path to novel treatments?. Br J Cancer 134, 843–848 (2026). https://doi.org/10.1038/s41416-025-03324-7
Received:
Revised:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41416-025-03324-7
