
Abstract
14-3-3σ is a p53-inducible gene with tumor suppressive properties and SMAD4 is a transcription factor encoded by a tumor suppressor gene, which is commonly inactivated in colorectal cancer (CRC). Here, 14-3-3σ was characterized as direct transcriptional target of SMAD4. TGF-β treatment of tumoroids derived from CRC patients and mouse models resulted in a SMAD4-dependent induction of 14-3-3σ. In murine, intestinal epithelia, the apical expression of 14-3-3σ was dependent on Smad4. Ectopic SMAD4 or 14-3-3σ promoted mesenchymal-to-epithelial transition (MET) and suppressed invasion, migration, and autophagy of CRC cells. As experimental inactivation of 14-3-3σ abolished these tumor-suppressive functions of SMAD4, 14-3-3σ mediates these effects of SMAD4. Inhibition of autophagy and promotion of MET by SMAD4 was mediated by inhibition of TFEB via binding and sequestration of TFEB by 14-3-3σ. The association of 14-3-3σ and TFEB was dependent on phosphorylation of the TFEB serine 211 residue, which is a target of mTORC1. Taken together, the TGF-β/SMAD4/14-3-3σ/TFEB axes characterized here antagonizes epithelial plasticity and autophagy. Thereby, it may ultimately suppress the progression of CRC and other types of cancer.
Similar content being viewed by others


Introduction
Colorectal cancer (CRC), the third most prevalent cancer type world-wide, leads to the second highest number of cancer-related deaths with 1.9 million new cases and 935,000 deaths each year [1, 2]. Despite a decrease in the general incidence and mortality rates, CRC is being detected more frequently in younger individuals and at more advanced stages and tends to locate mostly in the left colon and rectum [3]. The considerable mortality associated with CRC is largely due to its metastatic potential. Even with treatment advances, the five-year survival rate of patients with metastatic CRC (mCRC) remained at approximately 14% [1, 4]. Therefore, gaining deeper insights into the molecular pathways governing CRC metastasis is fundamental to improving mCRC prevention and treatment.
The tumor suppressor gene SMAD4/DPC4 maps to human chromosome 18q21 [5, 6]. Loss of chromosome 18q occurs in more than 60% of colorectal and other cancer types [5, 7]. Notably, loss of SMAD4 accounts for ~60% of the 18q21.1 allelic losses in CRC [8]. The SMAD4 transcription factor is a central mediator of the canonical transforming growth factor-beta (TGF-β) pathway [9, 10]. When TGF-β binds to its receptor, receptor-bound SMADs (R-SMADs) are phosphorylated, activated and form a complex with SMAD4 which translocates to the nucleus, and binds to SMAD-binding elements (SBEs) in the promoters or enhancers of target genes [9]. Moreover, SMAD4 associates with other DNA-binding proteins and co-factors to regulate genes involved in cell growth, proliferation, differentiation, apoptosis, and matrix production [11].
EMT and the reverse process, mesenchymal-epithelial transition (MET), facilitate cellular invasion and migration, and ultimately metastasis formation. EMT promotes the spread of cancer cells and MET is necessary for their colonization at secondary sites [12]. SMAD4 is a key tumor suppressor that can mediate anti-metastatic effects of TGF-β signaling in CRC by limiting migration and invasion [13]. The frequency of SMAD4 gene mutations is elevated in CRC patients with distant metastases, particularly in liver metastases, and is associated with poor prognosis [14, 15]. Furthermore, patients with CRCs expressing high SMAD4 levels display significantly improved overall and disease-free survival when compared to patients with low SMAD4 levels in CRCs [16]. The clinical significance of SMAD4 inactivation is not limited to CRC, e.g. in pancreatic ductal adenocarcinoma it is associated with metastatic dissemination and poor patient prognosis [17]. Concomitant deletion of Smad4 and Apc synergistically accelerates intestinal tumorigenesis and progression when compared to loss of Apc alone, indicating that loss of Smad4 function promotes the advancement and metastatic potential of CRC in vivo [18].
The 14-3-3σ protein (also known as Stratifin/SFN) belongs to the highly conserved 14-3-3 family and has been implicated in cell cycle control, apoptosis, and signal transduction [19]. In contrast to other 14-3-3 family members, 14-3-3σ exclusively forms homodimers [19]. This dimerization is important as it creates a pocket that mediates binding to ligand proteins via a motif with phosphorylated serine/threonine residues [20]. 14-3-3σ is directly induced by the p53 tumor suppressor after its activation by DNA damage [21]. 14-3-3σ sequesters cell cycle regulators, such as the CDC2-cyclin B1 complex, in the cytoplasm, resulting in a G2/M arrest [21]. 14-3-3σ is commonly down-regulated in cancers after p53 inactivation and by epigenetic silencing due to CpG methylation [22,23,24]. Deletion of 14-3-3σ significantly increases intestinal tumor formation in ApcMin/+ mice and shortens their life-span, suggesting a critical tumor-suppressive role of 14-3-3σ in CRC [25].
TFEB, member of the MiT/TFE family of basic helix-loop-helix-leucine-zipper (bHLH-Zip) transcription factors, is a master regulator of lysosomal biogenesis and autophagy [26]. Nuclear TFEB promotes transcription of genes within the coordinated lysosomal expression and regulation (CLEAR) motif, driving lysosomal function and cellular clearance [27]. Accelerated tumorigenesis is characterized by high energy consumption and scarcity of biosynthetic precursors relative to normal cells [28]. Thus, increased autophagolysosomal flux is a common feature of tumors, and makes TFEB an attractive subject in the study of cancer. Interestingly, TFEB is sequestered in the cytoplasm through phosphorylation and binding to 14-3-3 proteins (such as 14-3-3α/β and γ) [29]. However, it is unknown whether 14-3-3σ can bind to TFEB and block its function by cytoplasmic sequestration.
We had previously shown that 14-3-3σ expression increases apically in villi of intestinal epithelia [25]. As TGF-β signaling also increases in a similar pattern, we hypothesized that 14-3-3σ may represent a direct target and effector of the TGF-β pathway. We found that TGF-β induces 14-3-3σ via activating SMAD4, which directly binds to the 14-3-3σ promoter. Furthermore, SMAD4 promoted MET and inhibited autophagy through 14-3-3σ, which retained the transcription factor TFEB in the cytoplasm and thereby inhibited autophagy. Taken together, this pathway is likely to represent an important mediator of the suppressive effect of SMAD4 on CRC invasion and metastasis.
Results
14-3-3σ is a direct target gene of SMAD4 in human CRC cells
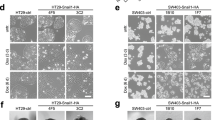
In order to determine, whether 14-3-3σ may represent an effector of the TGF-β pathway, we first determined whether SMAD4 induces the expression of 14-3-3σ. Therefore, SMAD4 was ectopically expressed in the colorectal cancer cell line SW620, which is known to be SMAD4-deficient [8], using the conditional, doxycycline (DOX)-inducible pRTR vector system described before [30]. As expected, treatment with doxycycline (DOX) resulted in a time-dependent increase in 14-3-3σ mRNA and 14-3-3σ protein expression (Fig. 1A, B). TGF-β1 treatment further enhanced SMAD4-induced 14-3-3σ mRNA and protein levels in SW620 cells (Fig. 1C, D), presumably by inducing the phosphorylation of SMAD. Similar results were obtained in SW480 cells (Fig. S1A-D). Furthermore, deletion of SMAD4 in a patient-derived tumor organoid (PDTO) abolished the TGF-β1-induced up-regulation of 14-3-3σ expression (Fig. 1E, F). Similar results were obtained in another PDTO (Fig. S1E). These results imply that the induction of 14-3-3σ by TGF-β is mediated by SMAD4. To evaluate the regulatory effect of TGF-β on 14-3-3σ expression, we performed an integrative analysis of GEO transcriptomic datasets (Fig. 1G). Overall, TGF-β stimulation resulted in a consistent induction of 14-3-3σ in different cell types and species. Of note, TGF-β induced an elevated expression of 14-3-3σ in several mouse cell models. In the promoter region of 14-3-3σ we identified several potential SMAD binding sites (SBE) using the JASPAR algorithm (Fig. 1H). qChIP analysis confirmed that SMAD4 occupies two predicted SBEs up- and down-stream of the 14-3-3σ coding region (Fig. 1H, I). Taken together, these results demonstrate that human 14-3-3σ is a direct target gene of SMAD4.

A qPCR analysis of 14-3-3σ expression in SW620 cells after SMAD4 induction with DOX for the indicated periods. B Western blot analysis of 14-3-3σ in SW620 cells treated as in A. C qPCR analysis of 14-3-3σ expression in SW620 cells after treatment with DOX and/or 20 ng/mL recombinant TGF-β1 for 48 h. D Western blot analysis of 14-3-3σ in SW620 cells treated as in C. E qPCR analysis of 14-3-3σ expression in SMAD4 wild-type and SMAD4 knockout PDTO2 after treatment with/without 20 ng/mL recombinant TGF-β1 for 72 h. F Western blot analysis of 14-3-3σ in PDTO2 cells treated as in E. G Expression changes of 14-3-3σ in response to TGF-β1 stimulations across multiple GEO datasets. Bars represent fold change values of 14-3-3σ relative to control, with datasets from both human and mouse cell types indicated on the y-axis. H The top panel shows SMAD4 potential binding site in jaspar.genereg.net. The bottom panel shows the map of human 14-3-3σ genomic regions with indicated SMAD4 binding sites. I qChIP analysis of SMAD4 occupancy at the human 14-3-3σ genomic regions in SW620 cells treated with DOX for 48 h. Human SMAD7 and 16q22 served as positive and negative controls, respectively. Results are presented as the mean ± SD (n = 3) for panel A, C, E and I with *p < 0.05, **p < 0.01, ***p < 0.001.
Murine 14-3-3σ is a direct target gene of Smad4
Next we determined, whether the regulation of 14-3-3σ by Smad4 is conserved between species. When the murine colorectal cell line CT26 was treated with TGF-β1 an increase in 14-3-3σ mRNA and protein levels was observed (Fig. 2A, B). Bioinformatics analysis hinted to a potential SBE approximately 3 kbp upstream of the murine 14-3-3σ promoter (Fig. 2C). qChIP analysis confirmed that Smad4 directly binds this SBE (Fig. 2C, D). To investigate whether Smad4 regulates 14-3-3σ expression in the mouse intestine, we generated Villin-CreERT2/Smad4fl/fl mice, which allow the conditional, intestinal epithelial cell-specific deletion of Smad4. From these mice we derived intestinal organoids. After exposure to 4-hydroxy-tamoxifen/4-OHT the expression of 14-3-3σ was significantly reduced on the mRNA and protein levels (Fig. 2E, F). Furthermore, 14-3-3σ expression was analysed in 8-week-old mice after intraperitoneal (IP) injection of Tamoxifen to induce deletion of Smad4 in vivo (Fig. S2A). Immunohistochemical analysis confirmed the absence of Smad4 expression in the small intestinal epithelium when compared to control Smad4fl/fl mice (Fig. 2G). Similar reductions were observed in the colonic epithelium (Fig. S2B). Deletion of Smad4 resulted in a lack of increased 14-3-3σ expression at mRNA and protein levels in the apical regions of epithelia in the small intestine (Fig. 2H, I) and the colon (Fig. S2C, D). To further explore the transcriptional response induced by TGF-β1, we re-analyzed the RNA-seq dataset published by Surakhy et al. (2025), which characterized the expression of Smad4-wildtype and Smad4-knockout adenoma organoids after 0, 1, and 12 h of TGF-β1 treatment [31]: 14-3-3σ expression was significantly up-regulated in Smad4 wild-type organoids after 12 h of TGF-β1 treatment, whereas no significant induction was observed in Smad4-deficient organoids (Fig. 2J). These results demonstrate that murine 14-3-3σ is a direct target gene of Smad4 and that intestinal epithelia display Smad4-dependent expression of 14-3-3σ during differentiation.

A PCR analysis of 14-3-3σ in CT26 cells after treatment with TGF-β1 for 72 h. B Western blot analysis of 14-3-3σ in CT26 cells after treatment with TGF-β1 for 72 h. C The top panel shows that the map of murine 14-3-3σ genomic regions with indicated Smad4 binding sites. The bottom panel is the map of mouse 14-3-3σ genomic regions with indicated Smad4 binding sites. D qChIP analysis of Smad4 occupancy at the murine 14-3-3σ genomic regions in CT26 after treatment with/without 20 ng/mL recombinant TGF-β1 for 48 h. Murine Smad7 and 16q22 served as positive and negative controls, respectively. E qPCR analysis of Smad4 and 14-3-3σ expression in organoids isolated from three male mice of each genotype treated with 4-OHT for the indicated days. F Western blot analysis of indicated protein levels in organoids treated as in E. G Immunohistochemical detection of Smad4 in 8 weeks old mice of the indicated genotype. n = 3 mice per genotype. Scale bar represent 100 μm. H 14-3-3σ mRNA was detected by ISH in 8 weeks old mice of the indicated genotype. n = 3 mice per genotype. Scale bar represents 100 μm. I Immunohistochemical detection of 14-3-3σ in 8 weeks old mice of the indicated genotype. n = 3 mice per genotype. Scale bar represents 100 μm. J Comparison of 14-3-3σ expression in Apc-/-/Smad4+/+ and Apc-/-/Smad4-/- organoids after TGF-β1 exposure for the indicated times. In panel A, D, E, and J mean values ± SD are shown (n = 3). **p < 0.01, ***p < 0.001. n.s. no significance.
Functional effects of SMAD4-induced MET
Ectopic expression of SMAD4 in SW620 CRC cells resulted in increased expression of the epithelial marker E-cadherin and decreased levels of the mesenchymal markers Vimentin and Snail1 (Fig. 3A). It also induced a morphology shift from spindle-shaped, mesenchymal-like cells to increased cell-cell contacts and loss of pseudopodia, indicating MET (ΔMET, Fig. 3B). In contrast, doxycycline (DOX) treatment of SW620 cells carrying an empty pRTR vector did not cause these changes in protein expression, nor morphological changes (Fig. 3A, B). Similar results were observed in SW480 cells upon ectopic SMAD4 expression (Fig. S3A, B). Furthermore, SMAD4 activation in SW620 cells resulted in a relocation of E-cadherin to the outer cell membrane, a marker specific for an epithelial state (Fig. 3C). Reorganization of the F-actin cytoskeletal and activation of β-catenin/TCF4 signaling are hallmarks of EMT/MET [30, 32, 33]. SMAD4 activation also induced rearrangement and reduction of cytoplasmic F-actin stress fibers (Fig. 3D). Additionally, SMAD4 re-expression induced relocation of β-catenin from the cytoplasm and nucleus to the plasma membrane, thereby presumably inhibiting β-catenin/TCF4 transcriptional activity (Fig. 3E). This is consistent with a previous report showing similar β-catenin re-localization in SW620 cells following ectopic SMAD4 expression [34]. Furthermore, ectopic SMAD4 expression significantly inhibited invasion, migration, and colony formation, while no inhibitory effects were observed in cells transfected with the empty pRTR vector (Fig. 3F–H). Similarly, activation of SMAD4 inhibited invasion and migration in SW480 cells (Fig. S3C, D). Therefore, ectopic SMAD4 induces MET and suppresses invasion, migration, and clonogenicity in mesenchymal-like CRC cell lines.

A Western blot analysis of indicated proteins after SMAD4 induction by DOX in SW620 cells for the indicated periods. As a control, SW620 cells containing a pRTR vector were also treated with DOX for the indicated periods. B Representative phase contrast images show the morphology of SW620 cells with or without the addition of DOX. eGFP induced by the addition of DOX served as a control. Scale bar: 50 μm. C–E E-cadherin, F-actin and β-catenin were detected by immunofluorescence after SW620 cells were treated with or without DOX for 48 h. Nuclear DNA was stained with DAPI. Scale bar: 20 μm. F Invasion was measured using a modified Boyden chamber assay. After 48 h of DOX treatment, the number of cells that invaded through the Matrigel was counted by crystal violet staining. G Wound healing assay after 48 h of treatment with or without DOX. Width of the wound was determined 24 h after scratching (top). The results represent the mean (%) of wound healing (bottom). Scale bar: 50 μm. H Colony formation assay of the indicated cells treated with or without DOX for 48 h. In panel F–H mean values ± SD are shown (n = 3). **p < 0.01, n.s. no significance.
Functional effects of 14-3-3σ-induced MET
Previous studies in adenomas from ApcMin/+ mice demonstrated that deletion of 14-3-3σ leads to up-regulation of mesenchymal markers (Snail1, ZEB1, and Vimentin) and down-regulation of epithelial markers (CDH1, EPCAM) [25]. These results implied that 14-3-3σ is required, at least to some extent, for keeping cells in an epithelial state. To determine whether 14-3-3σ is sufficient for inducing MET, we generated SW620 CRC cell pools with inducible, ectopic expression of 14-3-3σ. Ectopic expression of 14-3-3σ led to increased E-cadherin and reduced Vimentin levels (Fig. 4A), and induced a morphological shift from a mesenchymal to an epithelial phenotype (Fig. 4B). No corresponding changes were observed in cells transfected with the empty pRTR vector. In addition, ectopic 14-3-3σ resulted in a localization of E-cadherin to the outer membrane, loss and re-organization cytoplasmic F-actin, and translocation of β-catenin from the cytoplasm and nucleus to the outer cell membrane (Fig. 4C–E). Furthermore, ectopic 14-3-3σ inhibited cell invasion, migration, and colony formation, whereas no inhibitory effects were observed in control cells harboring the pRTR vector (Fig. 4F–H). These results show that 14-3-3σ is sufficient to induce MET and suppresses migration and invasion of mesenchymal-like CRC cells.

A Western blot analysis of indicated proteins after 14-3-3σ induction by DOX in SW620 cells for the indicated periods. As a control, SW620 cells containing a pRTR vector were also treated with DOX for the indicated periods. B Representative phase contrast images show the morphology of SW620 cells with or without the addition of DOX. eGFP induced with DOX served as a control. Scale bar: 50 μm. C–E E-cadherin, F-actin and β-catenin were detected by immunofluorescence after SW620 cells were treated with or without DOX for 48 h. Nuclear DNA was stained with DAPI. Scale bar: 20 μm. F Invasion was measured using a modified Boyden chamber assay. After 48 h of DOX treatment, the number of cells that invaded through the Matrigel was counted by crystal violet staining. G Wound healing assay after 48 h of treatment with or without DOX. Width of the wound was determined 24 h after scratching (top). The results represent the mean (%) of wound healing (bottom). Scale bar: 50 μm. H Colony formation assay of the indicated cells treated with or without DOX for 48 h. In F–H mean values ± SD are shown (n = 3). **p < 0.01, n.s. no significance.
14-3-3σ mediates induction of MET by SMAD4
To further investigate the role of 14-3-3σ in SMAD4-induced MET, we concomitantly silenced 14-3-3σ using siRNA pools during activation of a conditional SMAD4 allele (Fig. 5A). Ectopic SMAD4 increased E-cadherin expression, which was attenuated by silencing of 14-3-3σ. Conversely, a SMAD4-mediated decrease in Vimentin was reversed following knock-down of 14-3-3σ. Ectopic SMAD4 triggered MET, whereas cells with concomitant 14-3-3σ silencing displayed a mesenchymal morphology (Fig. 5B). In addition, ectopic SMAD4 resulted in recruitment of E-cadherin to the membranes which was prevented by concomitant silencing of 14-3-3σ (Fig. 5C). The SMAD4-induced redistribution and reduction of F-actin were dependent on 14-3-3σ, as simultaneous silencing of 14-3-3σ abrogated these effects (Fig. S4A). Ectopic SMAD4 expression promoted β-catenin membrane localization, which was blocked by knock-down of 14-3-3σ (Fig. 5D). 14-3-3σ silencing partially reversed the SMAD4-induced suppression of cell invasion, migration, and colony formation (Fig. 5E–G, Fig. S4B, C). These findings demonstrate that 14-3-3σ mediates SMAD4-induced MET and the resulting inhibition of invasion and migration.

A Lysates were subjected to immunoblot analysis of the indicated proteins 72 h after induction of SMAD4 by addition of DOX and simultaneous transfection with the indicated siRNAs in SW620 cells. B Representative phase contrast images show the morphology of SW620 cells treated as in A. Scale bar: 50 μm. C, D Indirect immunofluorescence detection of E-cadherin and β-catenin after treatment as shown in A. Scale bar: 20 μm. E Boyden chamber analysis was used to quantify cell invasion after cells were treated as indicated. DOX was treated for 48 h. F Quantification of wound healing after the indicated treatments. G Quantification of colony formation after the indicated treatments. In E–G mean values ± SD are shown (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001.
14-3-3σ mediates inhibition of autophagy by SMAD4
Increased autophagy has been associated with a more mesenchymal stem cell-like phenotype and is required for invasion and migration of glioblastoma stem cell lines [35]. To investigate the regulatory roles of SMAD4 or 14-3-3σ in autophagy, we assessed the expression of the autophagy effector LC3B-II. Ectopic SMAD4 resulted decreased LC3B-II levels, indicating an inhibition of autophagy by SMAD4 (Fig. 6A). Similarly, ectopic 14-3-3σ decreased LC3B-II, and reduced cytoplasmic LC3B puncta (Fig. 6B, S5). Furthermore, silencing of 14-3-3σ abrogated the inhibitory effect of SMAD4 on autophagy (Fig. 6C). Moreover, ectopic SMAD4 expression reduced the number of cytoplasmic LC3B puncta, whereas concomitant silencing of 14-3-3σ restored LC3B puncta formation in the presence of activated SMAD4 (Fig. 6D). Ectopic 14-3-3σ suppressed the autophagy-related genes TMEM55B, CTSA, CTSD, CLC7 and p62, without affecting TFEB (Fig. 6E). Similarly, SMAD4 activation inhibited the same set of genes; however, this inhibitory effect was abolished upon simultaneous 14-3-3σ silencing (Fig. 6F). Collectively, these results show that SMAD4 inhibits autophagy via inducing 14-3-3σ.

A Immunoblot analysis of indicated protein levels in SW620 cells after induction of ectopic SMAD4 by addition of DOX for the indicated periods. 20 μM of chloroquine was added for the last 4 h before harvesting cells. B Immunoblot analysis of indicated protein levels after addition of DOX for the indicated periods to induce ectopic 14-3-3σ. 20 μM chloroquine was added for the last 4 h before harvesting cells. C SW620 cells were transfected with control siRNA or 14-3-3σ-specific siRNA pools, and 24 h later, DOX was added for 48 h as indicated to induced ectopic SMAD4 expression. 20 μM chloroquine was added 4 h before harvesting cells. Immunoblot analysis of the indicated protein. D Indirect immunofluorescence detection of LC3B in SW620 cells transfected with the indicated siRNAs for 24 h and then treated with DOX for another 48 h. Nuclear DNA was stained with DAPI. Scale bar: 5 μm. The bar graph represents the quantification of puncta/cell. E qPCR analysis of the indicated mRNAs in SW620 cells treated with DOX for the indicated periods. F qPCR analysis of the indicated mRNAs after the treatments described in D. In E+F mean values ± SD are shown (n = 3). *p < 0.05, **p < 0.01, n.s. no significance.
SMAD4 inhibits TFEB via inducing 14-3-3σ
The transcription factor TFEB plays a central role in autophagy regulation [27]. The 14-3-3α/β/γ proteins are known to bind TFEB upon its phosphorylation by mTORC1 and subsequently inhibit TFEB by sequestration in the cytoplasm [29, 36, 37]. Therefore, we determined whether the SMAD4/14-3-3σ axis affects autophagy via regulating TFEB. Indeed, ectopic SMAD4 resulted in increased levels of TFEB in the cytoplasm while nuclear TFEB decreased (Fig. 7A, B). Also, ectopic expression of 14-3-3σ resulted in cytoplasmic sequestration of TFEB (Fig. 7C, D). After treatment with the nuclear export inhibitor leptomycin B, TFEB predominantly localized to the nucleus despite SMAD4 or 14-3-3σ activation, implying that SMAD4- or 14-3-3σ-induced TFEB nuclear-cytoplasmic translocation depends on nuclear export (Fig. S6A, B). Importantly, knockdown of 14-3-3σ abolished the SMAD4-induced cytoplasmic retention of TFEB, indicating that SMAD4 induced cytoplasmic localization of TFEB is mediated by the SMAD4-induced 14-3-3σ protein (Fig. 7E, F). Furthermore, gene set enrichment analysis (GSEA) revealed an enrichment of mRNAs up-regulated by TFEB (N = 414) in intestinal adenomas from 14-3-3σ-deficient mice (Fig. 7G). These findings indicate that inactivation of 14-3-3σ resulted in up-regulation of TFEB target genes, providing evidence that 14-3-3σ acts as a repressor of TFEB transcriptional activity in vivo.

A, C Western blot analysis of TFEB content in the cytoplasm and nucleus after DOX treatment for the indicated periods. B and D Immunofluorescence analysis of TFEB after DOX treatment for 48 h. Nuclear DNA was stained with DAPI. Scale bar: 20 μm. E and F SW620 cells were transfected with the indicated siRNAs for 24 h and then treated with DOX for 48 h as indicated. E Western blot analysis of TFEB in the cytoplasm and nucleus. F TFEB was then analyzed by indirect immunofluorescence. Nuclear DNA was stained with DAPI. Scale bar: 20 μm. G GSEA analysis of mRNAs up-regulated by TFEB (N = 414) in intestinal tissues from ApcMin/+ and ApcMin/+/14-3-3σ−/−.
Mechanisms and effects of 14-3-3σ-mediated sequestration of TFEB
To investigate how 14-3-3σ sequesters TFEB in the cytoplasm, we transiently co-transfected plasmids encoding 14-3-3σ with either HA-tagged, wild-type or S211A mutant TFEB expressing plasmids, respectively. Ectopic expression of 14-3-3σ retained wild-type TFEB in the cytoplasm, whereas the mutant S211A/TFEB protein primarily localized in the nucleus even in the presence of ectopic 14-3-3σ (Fig. 8A). Correspondingly, 14-3-3σ bound to wild-type TFEB but not to the S211A/TFEB mutant as determined by immune-precipitation analysis (Fig. 8B). Therefore, phosphorylation at position S211 is crucial for the binding of 14-3-3σ to TFEB. Concomitant ectopic expression of wild-type TFEB with 14-3-3σ decreased autophagy, whereas the S211A-TFEB mutant antagonized the inhibitory effect of 14-3-3σ (Fig. 8C). In addition, concomitant expression of 14-3-3σ and wild-type TFEB resulted in MET, whereas ectopic, mutant TFEB-S211A prevented 14-3-3σ-induced MET and resulted in a mesenchymal phenotype (Fig. 8D). The expression of MET markers changed accordingly (Fig. 8E). Moreover, concomitant ectopic expression of 14-3-3σ and TFEB-WT reduced cell invasion and migration, whereas ectopic expression of TFEB-S211A alleviated the effects of 14-3-3σ (Fig. 8F, G, Fig. S7A, B). These results demonstrate that SMAD4-induced 14-3-3σ inhibits autophagy, EMT, invasion, and migration by direct sequestration of TFEB.

A Cells were transiently transfected with the indicated plasmids following 24 h of treatment with or without DOX. After an additional 48 h, HA-tag expression was detected by indirect immunofluorescence analysis. Nuclear DNA was stained with DAPI. Scale bar: 20 μm. B Cells were treated with or without DOX for 24 h and then transfected with the indicated plasmids. Following an additional 48 h, cell lysates were subjected to immunoprecipitation (IP) with VSV-specific antibodies. After separation by gel electrophoresis, Western blotting analysis was used to detect the co-precipitated HA-tagged TFEB. “total lysates” indicate the expression of the proteins prior to immunoprecipitation. β-actin was used as a loading control. C Cells were transfected as in A, except that 20 μM of chloroquine was added 4 h before harvesting the cells. Immunoblotting was performed to assess the expression of the indicated protein. D After 24 h of treatment with or without DOX, cells were transfected with the indicated plasmids and then treated for another 48 h before analysis by phase-contrast microscopy. Scale bar: 50 μm. E Immunoblot assays analyze the indicated proteins after the treatments described in D. F Boyden chamber analysis was used to quantify cell invasion after cells were treated as described in D. G Quantification of wound healing after treatments as described in D. In F+G mean values ± SD are shown (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001. n.s. no significance.
Discussion
Here we show that SMAD4 directly induces expression of the 14-3-3σ gene in human and murine cells (see Fig. 9 for a summarizing scheme). The resulting increase in 14-3-3σ protein mediates down-stream effects of SMAD4, such as the induction of MET and inhibition of autophagy via sequestration of TFEB. These effects may ultimately contribute to SMAD4-mediated tumor suppression. For example, an increase in MET inhibits migration and invasion, and thereby prevents metastatic spread of cancer cells. Inhibition of autophagy may lower the threshold for apoptosis and prevent the survival of disseminated cancer cells. During therapy, a decrease of autophagy in tumor cells may increase their chemosensitivity. However, during tumor progression mutational inactivation of SMAD4 and/or down-regulation of 14-3-3σ may lead to activation of TFEB, which may have the opposite effect, and promote EMT and autophagy. Therefore, the molecular connections identified here are clinically relevant and may be exploited for therapeutic purposes in the future.

The scheme summarizes the findings of this study. Activation of SMAD4 by TGF-β results in its nuclear translocation and binding of SBEs within the 14-3-3σ promoter and transcriptional activation of 14-3-3σ. The resulting increase of cytoplasmic 14-3-3σ protein levels promotes sequestration of the transcription factor TFEB via binding to the phosphorylated Ser211 motif within TFEB. Therefore, the TFEB-/CLEAR-motif mediated transcription, which mediates autophagy and EMT, is inhibited, resulting in reduced invasion and migration, and presumably decreased survival of CRC cells. SBE SMAD-binding elements, CLEAR coordinated lysosomal and autophagic regulation.
Here, we showed that the SMAD4-induced 14-3-3σ facilitates the re-localization of β-catenin to the membrane which is known to decrease the transcriptional activity β-catenin/TCF complexes [38]. Given that hyperactivation of Wnt/APC/β-catenin pathway constitutes a major hallmark of CRC [39, 40], the loss of SMAD4 during CRC progression may therefore contribute to further activation of the transcriptional activity of β-catenin/TCF through down-regulation of 14-3-3σ. In addition, the SMAD4/14-3-3σ connection may also affect the PI3K/AKT, MAPK and Hippo/YAP signaling pathways, which have been shown to be down-stream of 14-3-3σ previously [41,42,43], and are involved in CRC tumorigenesis, therapeutic resistance and cellular plasticity.
Phosphorylation of TFEB at S211 by mTORC1 was shown to facilitate binding of the 14-3-3 family members 14-3-3α/β/γ next to a nuclear localization signal within the TFEB protein [29], thereby blocking nuclear translocation and preventing TFEB-driven expression of autophagy-related genes [44, 45]. Canonical SMAD signaling has been reported to precede mTORC1 activation in response to TGF-β1 stimulation [46]. Here, we show that 14-3-3σ specifically interacts with wild-type TFEB but not with the TFEB-S211A mutant, implying that phosphorylation of Ser211 is necessary for this interaction. mTORC1 is activated by multiple nutrient- and growth-related inputs, including amino acids via Rag GTPases [47], PI3K/AKT signaling [48], and cellular energy status sensed through AMPK-dependent regulation [49]. Since TFEB-induced autophagy has been implicated in promoting cancer progression through lysosomal biogenesis, EMT induction, cell cycle progression, and promotion of invasion in multiple cancer types [50,51,52], the TGF-β/SMAD4/14-3-3σ/TFEB axis may be a conserved tumor suppressive pathway in multiple types of cancers.
Consistent with the induction of MET by ectopic 14-3-3σ described here, deletion of the 14-3-3σ has been shown to promote EMT [53]. EMT is associated with enhanced autophagy, which allows cell-survival under conditions of stress [54]. In addition, autophagy was shown to induce EMT and anoikis resistance, and thereby enhance metastatic potential [54,55,56]. Recent studies indicate that TFEB activates Wnt/β-catenin signaling, thereby promoting EMT and enhancing the migratory and invasive capacities of gastric cancer cells [57]. Here, we show that cytoplasmic sequestration of TFEB by 14-3-3σ inhibits both MET and autophagy. It remains to be determined whether these processes are independently regulated in this context or whether they are linked. Since this study relies on in vitro and ex vivo models for functional analyses, future studies should extend these findings to animal models in order to validate the effects in vivo. Taken together, the TGF-β/SMAD4/14-3-3σ/TFEB axis described here facilitates the coordinated control of autophagy and EMT. This may ultimately contribute to suppression of mCRC. On the contrary, inactivation of its components may promote CRC progression.
Materials and methods
Generation and handling of mice
Smad4fl/fl mice (Stock No: 017462) were purchased from Jackson Laboratories. To generate mice with intestinal-specific knockout of Smad4, Vil-CreERT2 mice were crossed with Smad4fl/fl mice with a C57BL/6 background [58, 59]. All mice were housed in individually ventilated cages in the animal facility of the Biomedical Center (BMC) of the Ludwig-Maximilians-University (Munich, Germany). Mouse identification, healthy controls, and cage changes were performed under a laminar flow hood. Animal experimentation was approved by the local authorities. Genotyping primers are listed in Supplementary Table S1.
Cell culture and treatments
The human patient-derived tumor organoids (PDTOs) used here were initially described and maintained as in [60]. Murine intestinal crypt isolation and organoid culture were performed as previously described in [59]. The human colorectal cancer cell lines SW480 and SW620 were cultured in McCoy’s 5A medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (FBS) (Invitrogen) and 1% penicillin/streptomycin (Gibco, Thermo Fisher Scientific). Mouse colorectal cancer cells CT26 were cultured in RPMI1640 medium (σ-Aldrich, Gibco Life Technologies) with 10% FBS and 1% penicillin/streptomycin. All cells were cultured in an incubator with 20% O2, 5% CO2 and 37 °C. Doxycycline (DOX) (σ, St Louis, MO) was dissolved in water (100 μg/mL stock). The final concentration of DOX used for treatment was 100 ng/mL. To maintain stable cell pools containing the pRTR-vector plasmid [30], selection pressure was applied using puromycin at a final concentration of 8 μg/mL. Medium containing puromycin was replaced every 48 h to ensure continuous selection of transfected cells. FlexiTube GeneSolution GS2810 siRNA pools specific for 14-3-3σ (containing 4 different 14-3-3σ-specific siRNAs) and control siRNA pools were purchased from Qiagen (Hilden, Germany).
RNA isolation and real-time polymerase chain reaction (qPCR) analysis
Total RNA was isolated from cultured cells according to the manufacturer’s instructions of the High Pure RNA Isolation Kit (Roche). cDNA was generated using the Verso cDNA Kit (Thermo Scientific), and mRNA was analyzed by qPCR using Fast Green Master Mix (Applied Biosystems) and LightCycler 480 II (Roche Diagnostics). For human genes, gene expression was normalized to GAPDH or β-actin using the 2-ΔΔCt method [61], while murine gene expression was normalized to Cyclophilin or mouse β-actin. The primers used for qPCR are listed in Supplementary Table S2.
Analysis of expression from public databases
Transcriptomic datasets were obtained from the NCBI Gene Expression Omnibus (GEO) using the search term “TGF-β” combined with relevant cell or tissue terms. The inclusion criteria for each dataset were either publicly available raw data or publicly available processed expression matrices with sufficient annotation or with appropriate sample information. Normalized expression values (RMA) and RPKM values were used from microarray and RNA-seq datasets, respectively. From each dataset, we extracted the expression values of 14-3-3σ mRNA to compare fold changes between TGF-β-treated and control groups. The final dataset with the fold changes of 14-3-3σ was plotted as bar plots with controls set to 1. Gene Set Enrichment Analysis (GSEA) was performed using the GSEA software provided by the Broad Institute. First, public GEO gene expression profiling datasets of cell lines/tissues with ectopic TFEB expression were analyzed to generate a TFEB signature of 414 genes, that were consistently up-regulated by TFEB. Next, the enrichment of the TFEB signature in the previously published RNA-seq dataset from ApcMin/+ and ApcMin/+/14-3-3σ−/− mice [25] were analyzed. From GSEA, we obtained normalized enrichment scores (NES), FDR adjusted q-values, and generated enrichment plots showing the distributions of TFEB target genes.
Chromatin immunoprecipitation
Cells were subjected to chromatin immunoprecipitation according to the instructions in the iDeal ChIP-qPCR kit (Diagenode, Belgium). Briefly, cells were cross-linked with 11% formaldehyde in fixation buffer for approximately 10 min at room temperature, followed by the addition of glycine at 1:10 volume ratio with gentle shaking for 5 min at room temperature. After washing with PBS, samples were collected and lysed using the buffers provided in the kit (Buffer iL1B and iL2). Chromatin was fragmented into 200–500 bp fragments by sonication using shearing buffer iS1b, and shearing efficiency was verified by agarose gel electrophoresis. Equal amounts of sheared chromatin were incubated with antibodies against SMAD4 or negative control IgG overnight at 4 °C, and the immune complexes were captured using protein A-coated magnetic beads. Wash with appropriate wash buffers iW1 to iW4 to eliminate non-specific binding. Reverse crosslinking was performed with complete DIB buffer containing proteinase K. Separated DNA was then quantified by qPCR to determine enrichment at selected promoters or regulatory regions. The relative amount of immunoprecipitated DNA compared to input DNA for the control regions was calculated using the following formula: %recovery = 2^[(Ctinput – 6.64) - Ctsample] * 100%. Positive and negative control areas were included to assess the specificity and efficiency of the ChIP assay. Sequences of qChIP primers and antibodies are provided in Supplementary Tables S3.
Modified Boyden-chamber assay
To measure the invasion ability, Matrigel (Corning) and serum-free culture medium were mixed at a ratio of 1:6 and added to the culture chamber. The membrane was incubated at 37 °C for 3 h to form a coating. 1 × 105 cells were seeded in the upper chamber of serum-free culture medium (8.0 μM pore size filter, Corning), and the culture medium containing 10% fetal bovine serum as a chemo-attractant was seeded in the lower chamber. After 48 h, the invasive cells located on the lower surface of the chamber membrane were fixed with methanol for 30 min, stained with 0.5% crystal violet for 20 min, and then photographed and counted under a microscope. The fold change of invasive cells was calculated and normalized by comparing it with the corresponding control group.
Wound healing assay
The cells were seeded and cultured in culture-inserts (80241; IBIDI, Martinsried, Germany) until cells became confluent and formed a cell monolayer without intercellular spaces. The cells were treated with 10 μg/mL Mitomycin C (M4287, Sigma-Aldrich, Germany) for 2 h, and then the culture-inserts were removed. After washing twice with HBSS, fresh culture medium was added. The cells were photographed immediately with a phase contrast microscope and observed and photographed with a phase contrast microscope 24 h later.
Protein immunoprecipitation analysis
Protein immunoprecipitation analysis was performed according to the instructions of the Classic Magnetic IP/Co-IP Kit (Thermo Scientific, catalog number: 88804). Cells were seeded in 100 × 100 mm culture dishes. After the designated treatments, each dish was supplemented with 1 mL of IP Lysis/Wash Buffer and incubated on ice 5 min. The lysates were then collected and centrifuged at 4 °C for 10 min. The supernatant was transferred to a new tube, and protein concentration was determined using a BCA assay. 600 μg of total protein was incubated with 2 μg of the specific antibody. The antibody-lysate mixture was adjusted to a final volume of 500 μL with IP Lysis/Wash Buffer and rotated overnight (12–16 h) at 4 °C. 25 μL (0.25 mg) of Protein A/G magnetic beads were pre-washed with IP Lysis/Wash Buffer and added to each antibody-lysates mixture. Samples were rotated at room temperature for 1 h to allow immune complex binding. Beads were then collected with a magnetic rack and washed three times with 500 μL of IP/Wash Buffer, followed by one rinse with ultrapure water. Bound proteins were eluted with 100 μL of elution buffer at room temperature for 10 min with gentle mixing, and the eluates were subsequently denatured at 95 °C before immunoblot analysis. The list of antibodies is shown in Table S4.
Western blot analysis
Cells were washed with HBSS and lysed with RIPA lysis buffer containing mini protease inhibitor (Roche) and PhosSTOP Phosphatase Inhibitor Cocktail Tablets (Roche). The cell lysate was sonicated for 5 s and then centrifuged at 13,000 rpm for 20 min at 4 °C to collect the supernatant containing proteins, which were then quantified according to the instructions of the PierceTM BCA protein Assay Kit (Thermo Fisher Scientific). Protein samples were separated by electrophoresis on 10% or 12% SDS-PAGE gels and transferred to PVDF membranes (Millipore). The ECL (Millipore) system was used and images were obtained with a LI-COR Odyssey FC Imaging System (Bad Homburg, Germany). Western blots were quantified using Image J software. A list of antibodies is provided in Table S4.
Colony formation assay
Cells were seeded in 6-well plates and treated as indicated. The treated cells were seeded into 12-well plates at a density of 1000 cells/well, and colony formation was measured after 2 weeks. After methanol fixation and crystal violet staining, colonies were recorded with a digital camera (Nikon, Japan) and counted using Image J software.
In situ hybridization analysis (ISH)
The analysis was performed as described previously [62]. In brief, formaldehyde-fixed tissue samples were placed on glass slides. Cell membranes were permeabilized by 30 μg/mL proteinase K treatment. 14-3-3σ-specific in situ RNA probes were generated as described previously [25]. The probes were added to the samples, hybridized at 65 °C, and washed to remove unbound probes. The signals were detected using the BCIP/NBT Liquid Substrate System (Sigma), and the signal locations were observed under a microscope to show the spatial distribution of the target nucleic acid. The slides were scanned using a Vectra® PolarisTM Imaging System.
Immunohistochemical analysis
Mouse small intestine tissues were fixed with formalin. Paraffin-embedded sections (5 μm) were deparaffinized in xylene and rehydrated through a graded ethanol series. Antigen retrieval was performed by microwaving the slides in Target Retrieval Solution Citrate (pH 6.0, Dako) at 750 W for 25 min. After cooling to room temperature, endogenous peroxidase activity was blocked with 3% hydrogen peroxide for 10 min. Blocking was then carried out following the protocol of the ImmPRESS HRP Horse Anti-Rabbit (or Anti-Mouse) IgG Polymer Kit (Vector Laboratories). Primary antibodies were diluted in Dako Antibody Diluent with Background Reducing Components, applied to slides and incubated overnight at 4 °C. After washing with PBS, the sections were treated with an HRP-conjugated antibody for 1 h at room temperature. The Dako Liquid DAB Substrate Chromogen System was then used for visualization and sections were counterstained with hematoxylin (Vector). Cover-slips were applied using Roti®-Histokitt II (Roth). Slides were scanned using a Vectra® PolarisTM Imaging System. Data analysis was conducted using Phenochart version 2.0.0 software. A list of antibodies is provided in Table S4.
Immunofluorescence analysis by confocal laser-scanning microscopy
Cells were cultured on round glass slides in 12-well plates and then fixed in 4% paraformaldehyde for 15 min, permeabilized with 0.2% Triton X-100 for 10 min, and blocked with 5% BSA in PBS for 1 h at the room temperature. Next, primary antibodies were incubated at 4 °C for 12 to 16 h in a humidified chamber, washed three times with PBS-Tween 20, and then incubated with appropriate secondary antibodies for 1 h at room temperature. Cell chromatin was stained with DAPI (Roche) for 5 min. After final washes, coverslips were mounted with ProLong Gold anti-fluorescent dye (Invitrogen). Images were acquired using a confocal microscope (LSM 700, Zeiss) with 63x Plan-Apochromat oil immersion lens and ZEN 2009 software (Zeiss). Puncta quantification of LC3B was performed using ImageJ software. For each condition, at least three microscopic fields encompassing a total of no fewer than 200 cells were analyzed. Antibodies are listed in Table S4.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 10 software (USA). Statistical differences between the two groups were calculated using Student’s t-test (two-tailed; unpaired). Asterisks generally indicate: *p < 0.05, **p < 0.01, and ***p < 0.001, n.s. no significant.
Data availability
All data, analytic methods, and study materials will be made available to other researchers upon reasonable request.
References
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021;71:209–49.
Arnold M, Sierra MS, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global patterns and trends in colorectal cancer incidence and mortality. Gut. 2017;66:683–91.
Siegel RL, Wagle NS, Cercek A, Smith RA, Jemal A. Colorectal cancer statistics, 2023. CA Cancer J Clin. 2023;73:233–54.
Shin AE, Giancotti FG, Rustgi AK. Metastatic colorectal cancer: mechanisms and emerging therapeutics. Trends Pharmacol Sci. 2023;44:222–36.
Thiagalingam S, Lengauer C, Leach FS, Schutte M, Hahn SA, Overhauser J, et al. Evaluation of candidate tumour suppressor genes on chromosome 18 in colorectal cancers. Nat Genet. 1996;13:343–6.
Hahn SA, Schutte M, Hoque AT, Moskaluk CA, da Costa LT, Rozenblum E, et al. DPC4, a candidate tumor suppressor gene at human chromosome 18q21.1. Science. 1996;271:350–3.
Cho KR, Oliner JD, Simons JW, Hedrick L, Fearon ER, Preisinger AC, et al. The DCC gene: structural analysis and mutations in colorectal carcinomas. Genomics. 1994;19:525–31.
Woodford-Richens KL, Rowan AJ, Gorman P, Halford S, Bicknell DC, Wasan HS, et al. SMAD4 mutations in colorectal cancer probably occur before chromosomal instability, but after divergence of the microsatellite instability pathway. Proc Natl Acad Sci USA. 2001;98:9719–23.
Massague J. TGFbeta signalling in context. Nat Rev Mol Cell Biol. 2012;13:616–30.
Heldin CH, Miyazono K, ten Dijke P. TGF-beta signalling from cell membrane to nucleus through SMAD proteins. Nature. 1997;390:465–71.
Heldin CH, Moustakas A. Signaling Receptors for TGF-beta Family Members. Cold Spring Harb Perspect Biol. 2016;8:a022053.
Nieto MA, Huang RY, Jackson RA, Thiery JP. Emt: 2016. Cell. 2016;166:21–45.
Zhang B, Halder SK, Kashikar ND, Cho YJ, Datta A, Gorden DL, et al. Antimetastatic role of Smad4 signaling in colorectal cancer. Gastroenterology. 2010;138:969–80. e1-3.
Miyaki M, Iijima T, Konishi M, Sakai K, Ishii A, Yasuno M, et al. Higher frequency of Smad4 gene mutation in human colorectal cancer with distant metastasis. Oncogene. 1999;18:3098–103.
Losi L, Bouzourene H, Benhattar J. Loss of Smad4 expression predicts liver metastasis in human colorectal cancer. Oncol Rep. 2007;17:1095–9.
Alazzouzi H, Alhopuro P, Salovaara R, Sammalkorpi H, Jarvinen H, Mecklin JP, et al. SMAD4 as a prognostic marker in colorectal cancer. Clin Cancer Res. 2005;11:2606–11.
Iacobuzio-Donahue CA, Fu B, Yachida S, Luo M, Abe H, Henderson CM, et al. DPC4 gene status of the primary carcinoma correlates with patterns of failure in patients with pancreatic cancer. J Clin Oncol. 2009;27:1806–13.
Takaku K, Oshima M, Miyoshi H, Matsui M, Seldin MF, Taketo MM. Intestinal tumorigenesis in compound mutant mice of both Dpc4 (Smad4) and Apc genes. Cell. 1998;92:645–56.
Hermeking H. The 14-3-3 cancer connection. Nat Rev Cancer. 2003;3:931–43.
Muslin AJ, Xing H. 14-3-3 proteins: regulation of subcellular localization by molecular interference. Cell Signal. 2000;12:703–9.
Hermeking H, Lengauer C, Polyak K, He TC, Zhang L, Thiagalingam S, et al. 14-3-3sigma is a p53-regulated inhibitor of G2/M progression. Mol Cell. 1997;1:3–11.
Lodygin D, Hermeking H. The role of epigenetic inactivation of 14-3-3sigma in human cancer. Cell Res. 2005;15:237–46.
Lodygin D, Hermeking H. Epigenetic silencing of 14-3-3sigma in cancer. Semin Cancer Biol. 2006;16:214–24.
Ferguson AT, Evron E, Umbricht CB, Pandita TK, Chan TA, Hermeking H, et al. High frequency of hypermethylation at the 14-3-3σ locus leads to gene silencing in breast cancer. Proc Natl Acad Sci USA. 2000;97:6049–54.
Winter M, Rokavec M, Hermeking H. 14-3-3sigma Functions as an Intestinal Tumor Suppressor. Cancer Res. 2021;81:3621–34.
Sardiello M, Palmieri M, di Ronza A, Medina DL, Valenza M, Gennarino VA, et al. A gene network regulating lysosomal biogenesis and function. Science. 2009;325:473–7.
Settembre C, Di Malta C, Polito VA, Garcia Arencibia M, Vetrini F, Erdin S, et al. TFEB links autophagy to lysosomal biogenesis. Science. 2011;332:1429–33.
Yang S, Wang X, Contino G, Liesa M, Sahin E, Ying H, et al. Pancreatic cancers require autophagy for tumor growth. Genes Dev. 2011;25:717–29.
Xu Y, Ren J, He X, Chen H, Wei T, Feng W. YWHA/14-3-3 proteins recognize phosphorylated TFEB by a noncanonical mode for controlling TFEB cytoplasmic localization. Autophagy. 2019;15:1017–30.
Jackstadt R, Roh S, Neumann J, Jung P, Hoffmann R, Horst D, et al. AP4 is a mediator of epithelial-mesenchymal transition and metastasis in colorectal cancer. J Exp Med. 2013;210:1331–50.
Surakhy M, Matheson J, Barnes DJ, Carter EJ, Hughes J, Buhnemann C, et al. Smad4 and TGFbeta1 dependent gene expression signatures in conditional intestinal adenoma, organoids and colorectal cancer. Sci Rep. 2025;15:16330.
Moreno-Bueno G, Peinado H, Molina P, Olmeda D, Cubillo E, Santos V, et al. The morphological and molecular features of the epithelial-to-mesenchymal transition. Nat Protoc. 2009;4:1591–613.
Brabletz T, Hlubek F, Spaderna S, Schmalhofer O, Hiendlmeyer E, Jung A, et al. Invasion and metastasis in colorectal cancer: epithelial-mesenchymal transition, mesenchymal-epithelial transition, stem cells and beta-catenin. Cells Tissues Organs. 2005;179:56–65.
Tian X, Du H, Fu X, Li K, Li A, Zhang Y. Smad4 restoration leads to a suppression of Wnt/beta-catenin signaling activity and migration capacity in human colon carcinoma cells. Biochem Biophys Res Commun. 2009;380:478–83.
Galavotti S, Bartesaghi S, Faccenda D, Shaked-Rabi M, Sanzone S, McEvoy A, et al. The autophagy-associated factors DRAM1 and p62 regulate cell migration and invasion in glioblastoma stem cells. Oncogene. 2013;32:699–712.
Zhang W, Li X, Wang S, Chen Y, Liu H. Regulation of TFEB activity and its potential as a therapeutic target against kidney diseases. Cell Death Discovery. 2020;6:32.
Roczniak-Ferguson A, Petit CS, Froehlich F, Qian S, Ky J, Angarola B, et al. The transcription factor TFEB links mTORC1 signaling to transcriptional control of lysosome homeostasis. Sci Signal. 2012;5:ra42.
Li FQ, Mofunanya A, Harris K, Takemaru K. Chibby cooperates with 14-3-3 to regulate beta-catenin subcellular distribution and signaling activity. J Cell Biol. 2008;181:1141–54.
Clevers H, Nusse R. Wnt/beta-catenin signaling and disease. Cell. 2012;149:1192–205.
Dzialo E, Tkacz K, Blyszczuk P. Crosstalk between the TGF-beta and WNT signalling pathways during cardiac fibrogenesis. Acta Biochim Pol. 2018;65:341–9.
Morrison DK. The 14-3-3 proteins: integrators of diverse signaling cues that impact cell fate and cancer development. Trends Cell Biol. 2009;19:16–23.
Tzivion G, Avruch J. 14-3-3 proteins: active cofactors in cellular regulation by serine/threonine phosphorylation. J Biol Chem. 2002;277:3061–4.
Yu FX, Zhao B, Guan KL. Hippo Pathway in Organ Size Control, Tissue Homeostasis, and Cancer. Cell. 2015;163:811–28.
Settembre C, Zoncu R, Medina DL, Vetrini F, Erdin S, Erdin S, et al. A lysosome-to-nucleus signalling mechanism senses and regulates the lysosome via mTOR and TFEB. EMBO J. 2012;31:1095–108.
Martina JA, Chen Y, Gucek M, Puertollano R. MTORC1 functions as a transcriptional regulator of autophagy by preventing nuclear transport of TFEB. Autophagy. 2012;8:903–14.
Woodcock HV, Eley JD, Guillotin D, Plate M, Nanthakumar CB, Martufi M, et al. The mTORC1/4E-BP1 axis represents a critical signaling node during fibrogenesis. Nat Commun. 2019;10:6.
Sancak Y, Bar-Peled L, Zoncu R, Markhard AL, Nada S, Sabatini DM. Ragulator-Rag complex targets mTORC1 to the lysosomal surface and is necessary for its activation by amino acids. Cell. 2010;141:290–303.
Dibble CC, Cantley LC. Regulation of mTORC1 by PI3K signaling. Trends Cell Biol. 2015;25:545–55.
Gwinn DM, Shackelford DB, Egan DF, Mihaylova MM, Mery A, Vasquez DS, et al. AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol Cell. 2008;30:214–26.
Astanina E, Bussolino F, Doronzo G. Multifaceted activities of transcription factor EB in cancer onset and progression. Mol Oncol. 2021;15:327–46.
Zhu X, Zhuo Y, Wu S, Chen Y, Ye J, Deng Y, et al. Corrigendum: TFEB Promotes Prostate Cancer Progression via Regulating ABCA2-Dependent Lysosomal Biogenesis. Front Oncol. 2021;11:750277.
Blessing AM, Rajapakshe K, Reddy Bollu L, Shi Y, White MA, Pham AH, et al. Transcriptional regulation of core autophagy and lysosomal genes by the androgen receptor promotes prostate cancer progression. Autophagy. 2017;13:506–21.
Raychaudhuri K, Chaudhary N, Gurjar M, D’Souza R, Limzerwala J, Maddika S, et al. 14-3-3sigma Gene Loss Leads to Activation of the Epithelial to Mesenchymal Transition Due to the Stabilization of c-Jun Protein. J Biol Chem. 2016;291:16068–81.
Gugnoni M, Sancisi V, Manzotti G, Gandolfi G, Ciarrocchi A. Autophagy and epithelial-mesenchymal transition: an intricate interplay in cancer. Cell Death Dis. 2016;7:e2520.
Mowers EE, Sharifi MN, Macleod KF. Autophagy in cancer metastasis. Oncogene. 2017;36:1619–30.
Babaei G, Aziz SG, Jaghi NZZ. EMT, cancer stem cells and autophagy; The three main axes of metastasis. Biomed Pharmacother. 2021;133:110909.
Li S, Liu F, Xu L, Li C, Yang X, Guo B, et al. Wnt/beta-Catenin Signaling Axis Is Required for TFEB-Mediated Gastric Cancer Metastasis and Epithelial-Mesenchymal Transition. Mol Cancer Res. 2020;18:1650–9.
el Marjou F, Janssen KP, Chang BH, Li M, Hindie V, Chan L, et al. Tissue-specific and inducible Cre-mediated recombination in the gut epithelium. Genesis. 2004;39:186–93.
Jaeckel S, Kaller M, Jackstadt R, Gotz U, Muller S, Boos S, et al. Ap4 is rate limiting for intestinal tumor formation by controlling the homeostasis of intestinal stem cells. Nat Commun. 2018;9:3573.
Loevenich LP, Tschurtschenthaler M, Rokavec M, Silva MG, Jesinghaus M, Kirchner T, et al. SMAD4 loss induces c-MYC-mediated NLE1 upregulation to support protein biosynthesis, colorectal cancer growth, and metastasis. Cancer Res. 2022;82:4604–23..
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402–8.
Gregorieff A, Clevers H. In situ hybridization to identify gut stem cells. Curr Protoc Stem Cell Biol. 2010;Chapter 2:Unit 2F 1.
Acknowledgements
We thank Ursula Götz for technical support and, Peter Jung and Leon P. Loevenich for providing PDTO2 and PDTO4 tumoroids and cDNA derived from these. Fangteng Liu for assistance with in situ hybridization.
Funding
XC is a recipient of a Chinese Scholarship Council fellowship. The project was funded by grants from the Deutsche Krebshilfe to HH. Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
HH conceived, planned, and supervised the project; HH, MW, and XC designed experiments; XC performed experiments and analyzed results. MW and JK designed the mouse crossing. MR performed the bioinformatics analyses. HH and XC wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
Animal experimentations and analyses were approved by Government of Upper Bavaria, Germany (ROB-55.2Vet-2532.Vet-02-20-154). All methods were performed in accordance with the relevant guidelines and regulations.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by Dr Gerry Melino
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chen, X., Winter, M., Rokavec, M. et al. TGF-β/SMAD4/14-3-3σ/TFEB axis promotes mesenchymal-epithelial transition and inhibits autophagy in colorectal cancer. Cell Death Dis 17, 397 (2026). https://doi.org/10.1038/s41419-026-08733-x
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41419-026-08733-x
