
Abstract
Congenital hypogonadotropic hypogonadism (CHH) is a rare and genetically heterogeneous disorder characterized by absent or incomplete puberty due to impaired gonadotropin-releasing hormone (GnRH) function. A subset of individuals with CHH also present with developmental anomalies, including midline defects such as cleft lip and/or palate (CLP). This study investigates the genetic overlap between CHH and CLP. A total of 336 individuals diagnosed with CHH were clinically assessed for associated phenotypes, including CLP. High-throughput sequencing was performed using a targeted gene panel encompassing known CHH- and CLP-related genes. Variants were analyzed and classified according to the American College of Medical Genetics and Genomics (ACMG) criteria for pathogenicity. CLP was present in 21 patients with CHH (6%). Pathogenic or likely pathogenic variants in genes associated with both CHH and CLP—such as FGFR1 and CHD7—were identified in eight individuals. Furthermore, 17% of the patients with CHH without CLP harbored deleterious variants in genes implicated in clefting, including DVL3, PLCB4, NIPBL, and EDNRA. Evidence of digenic inheritance involving both CHH- and CLP-related genes was observed in multiple cases. FGFR1 variants were the most frequently detected and were commonly associated with anosmia and additional developmental anomalies. These findings highlight a genetic and phenotypic continuum between CHH and CLP, underscoring the involvement of shared developmental pathways. The high prevalence of FGFR1 variants in patients with CHH and CLP supports its role as a pleiotropic gene. Understanding the overlapping genetic mechanisms may enhance diagnostic precision and inform personalized management strategies for affected individuals.
Similar content being viewed by others

Introduction
Congenital hypogonadotropic hypogonadism (CHH) is a rare heterogeneous genetic disorder occurring in 1 in 10,000 births [1]. It is characterized by the absence of puberty and subsequent infertility due to gonadotropin-releasing hormone (GnRH) deficiency [2]. The developmental link between the GnRH and olfactory systems accounts for the frequent association of CHH with anosmia, a condition known as Kallmann syndrome (KS) [3]. Over the past 30 years, genetic studies in both humans and mice have identified more than 40 loci harboring rare, high-impact variants implicated in CHH pathogenesis [4, 5]. Pathogenic (P) or likely pathogenic (LP) variants in known CHH genes are found in around 50% of patients, highlighting the need to identify additional genetic contributors [6]. Oligogenic inheritance—mutations in two or more genes—likely contributes to the variable expressivity seen both within and between affected families carrying the same mutation. It is presently observed in approximately 15% of patients, this landscape is likely to increase as more genes are discovered [7].
Patients with CHH often present with additional developmental anomalies, most commonly anosmia in around 50% of cases, but also including cleft lip and/or palate (CLP) [5, 8]. Our previous work has shown phenotypic and genetic overlap between CHH and several other syndromic conditions, such as septo-optic dysplasia (SOD), combined pituitary hormone deficiency (CPHD), and CHARGE syndrome (characterized by coloboma, heart defects, choanal atresia, growth retardation, genital anomalies, and ear abnormalities) [9, 10].
CLP is among the most common congenital malformations, with a complex etiology and substantial impact on morbidity and quality of life. It affects approximately 1 in 700 live births, with prevalence varying significantly by ethnicity [11]. Based on anatomical, embryological, and genetic evidence, orofacial clefts are typically classified into two major categories: cleft lip with or without cleft palate (CLP) and cleft palate only (CP). The clinical burden of these anomalies depends on severity and may include feeding difficulties, speech impairments, ear, nose and throat complications, dental and orthodontic issues, aesthetic concerns, and psychosocial challenges that can persist into adulthood [11]. Clefts may occur as part of a broader syndromic presentation involving additional developmental anomalies or, in isolation, referred to as non-syndromic CLP. Approximately 70% of CLP and 50% of CP cases are classified as non-syndromic [12]. The etiology of CLP is multifactorial, involving both genetic and environmental determinants. The genetic architecture is complex, with mutations in several genes already been described. Genome-wide association studies also contributed in the dentification of several genes and variants that are associated with CLP susceptibility [11]. Five to ten percent of probands with CHH also exhibit CLP [8, 13,14,15]. This prevalence increases to 30–40% among patients with CHH carrying FGFR1 mutations, highlighting a notable clinical and genetic overlap between CHH and CLP [8, 16]. Moreover, FGFR1 mutations have also been implicated in isolated cases of CLP [17]. While prior studies have evaluated the role of CHH-associated genes—such as FGFR1 and FGF8—in cohorts with CLP [18], the reverse approach, namely, the investigation of CLP-related genes in patients with CHH, has not been explored.
The aim of this study was to expand our understanding of the clinical and genetic overlap between CHH and CLP. Through high-throughput sequencing and comprehensive variant analysis, we sought to identify shared molecular mechanisms, uncover novel CLP-related genes implicated in CHH, investigate potential digenic inheritance patterns, and gain deeper insight into the developmental pathways connecting these two conditions.
Material and methods
Patients and clinical evaluation
The CHH cohort was comprised 336 unselected probands. The patients are part of Prof. N. Pitteloud’s cohort that she has assembled over more than 2 decades. This cohort comprises samples from patients coming from several medical centers all over the world, yet mainly from Europe. All participants provided written informed consent, and the study was conducted in accordance with the Declaration of Helsinki. Diagnosis of CHH was established based on the criteria outlined in the European Consensus Statement on CHH [19]. Specifically, CHH was defined by: (i) absent or incomplete puberty by age 17; (ii) low or inappropriately normal gonadotropin levels in the context of low serum sex steroids (testosterone in males or estradiol in females); (iii) normal hypothalamic and pituitary anatomy on imaging; (iv) otherwise normal anterior pituitary function, and (v) absence of iron overload.
Olfactory function was assessed through self-report and/or formal testing using the University of Pennsylvania Smell Identification Test [16]. All participants underwent a comprehensive medical examination and review of clinical history. Magnetic resonance imaging was used to evaluate the olfactory bulbs and tracts, as well as the hypothalamic-pituitary region. Diagnosis of CLP was based on clinical evaluation, supplemented by radiographs and computed tomography imaging when necessary.
Genetic analysis
DNA extraction and sequencing
Genomic DNA was extracted from blood samples of the 336 patients with CHH and their available relatives using standard protocols provided by the manufacturer. WES and WGS were performed using the Agilent SureSelect v5 exome capture kit and the DNBSEQ-T20×2 platform, respectively. Sequencing was carried out on the Illumina HiSeq 2500 platform, following previously described protocols [7].
The resulting raw sequencing data (FASTQ files) were processed using an in-house bioinformatics pipeline built around the Sentieon DNASeq toolkit, a GATK-compliant platform [20, 21]. Reads were aligned to the human reference genome (GRCh38), and single-nucleotide variants (SNVs) as well as small insertions and deletions (Indels <50 bp) were identified. Variant annotation included population allele frequencies from gnomAD v4.1 [22] and in silico predictions of pathogenicity using tools such as CADD [23], SpliceAI [24], REVEL [25] and Alphamissense [26] facilitated by ANNOVAR [27]. Sequencing was performed to a minimum average coverage depth of 30× per sample.
Annotated variants were subsequently visualized, filtered, and prioritized using GenMasterTable, a variant interpretation platform developed in-house [28].
SNVs analysis
Variants were selected based on one or more of the following criteria: (i) predicted loss-of-function variants, including nonsense (stop-gain), frameshift, and canonical splice site mutations (± 2 bp from an exon); (ii) missense variants; (iii) in-frame insertions or deletions; or (iv) a SpliceAI-predicted probability > 0.8 of causing a splicing defect [24]. All variants met GATK quality control thresholds, including a minimum quality score of 50.
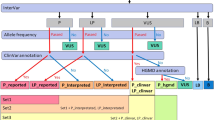
Filtered variants were assessed against two panels: panel A—a curated CLP gene panel, which includes 162 genes and panel B—a CHH-specific gene panel with 41 genes (Supplementary Table 1). There are two overlapping genes in both panels (A + B), FGFR1 and CHD7, since they have already been associated with both phenotypes. Panel A was assembled using PanelApp, a publicly accessible, expert-curated virtual gene panel resource [29], downloaded on April 20, 2023. Panel B comprises 41 genes known to be associated with CHH as documented in the OMIM database (Supplementary Table 1).
Consistent with inheritance patterns commonly observed in rare diseases, variants were considered potentially pathogenic according to the following thresholds: (i) autosomal recessive, MAF < 1% (ii) autosomal dominant MAF < 0.1% and X-linked MAF < 0.1%. Variants were further annotated and classified for pathogenicity using VarSome [30], based on the guidelines of the American College of Medical Genetics and Genomics [31]. A variant was considered novel if it had not been previously reported in the medical literature or was absent from the human gene mutation database [32] and ClinVar https://www.ncbi.nlm.nih.gov/clinvar/. All prioritized variants were confirmed by Sanger sequencing, using previously published protocols [33], and, when possible, familial segregation analysis was performed.
Results
Probands with CHH and CLP
Among the 336 patients with CHH, 21 (6%) presented with CLP. Heterozygous P or LP variants in CLP-related genes were identified in 10 of these individuals (5 males; Table 1); among these 10 individuals, eight harbored variants in genes known to be implicated in both CLP and CHH individually. Of note, FGFR1 and CHD7 are both CLP and CHH-related genes and are listed in panels A and B (Supplementary Table S1).
Among the patients with CHH and CLP, the most frequently observed deleterious variants were in FGFR1 (n = 7). Among these, five patients exhibited anosmia or hyposmia, and three had complex associated phenotypes: patient 4 presented with synkinesia, dental agenesis, growth retardation, and myasthenia gravis; Patient 5 had SOD, hearing loss, and intellectual disability; and patient 7 was diagnosed with congenital ocular motor dyspraxia in childhood. In six cases, the variant was inherited from one parent. In all but two families (patients 3 and 5), the transmitting parent was asymptomatic, consistent with the incomplete penetrance previously described for FGFR1 mutations [28]. For patient 7, parental DNA was not available for genetic analysis.
The remaining three patients carried heterozygous deleterious variants in three different genes: CHD7, SOX9, and CTNND1 (Table 1). Patient 8, a female with a LP CHD7 variant (p.Ala1107Val), exhibited anosmia and a complex phenotype including hearing loss, skeletal anomalies, synkinesia, mitral valve prolapse, atrioventricular nodal reentry tachycardia, astigmatism, choroidal nevus, vesicoureteral reflux, and unilateral renal hypoplasia. The variant was inherited from her mother, who had hearing loss. Her sister, who also carried the variant, presented with both CHH and CLP; patient 9, a female, harbored a de novo pathogenic SOX9 variant (p.Pro170Arg). She exhibited anosmia and multiple anomalies, including ear malformations, growth retardation, hearing loss, intellectual and developmental disability (ID/DD), bicuspid aortic valve, synkinesia, strabismus, metatarsus varus, brachydactyly, and low-set ears; and Patient 10, a male, carried a LP splice-site variant in CTNND1 (c.2809-1 G > C), inherited from his affected father, who had delayed puberty and CLP. The proband exhibited hypertelorism, double permanent teeth, hearing loss, unilateral cryptorchidism, and brachydactyly. He had four brothers: three with facial clefts who also carried the variant, and one with delayed puberty who did not carry the variant.
Overall, among the 10 index patients, only two had a normal sense of smell, while five presented with complex associated phenotypes—hearing loss being the most common, observed in four individuals.
Probands with CHH without CLP harbor P/LP variants in CLP genes
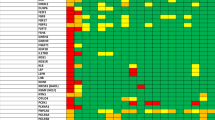
Among the remaining 315 probands with CHH without CLP, 50 harbored P and LP variants in CLP-associated genes, representing 15.8% of the cohort (Fig. 1). Notably, four cases exhibited potential digenic inheritance involving both CLP-related and CHH-associated genes (Fig. 1).

A LP/P variants in CLP genes in 50 CHH patients without CLP. B 35 FGFR1 mutations in 336 probands with CHH. FGFR1 mutations in patients with CHH and CLP are depicted below.
Probands with CHH without CLP harboring deleterious variants in overlapping genes for CHH and CLP
FGFR1 and CHD7 are individually implicated in both CHH and CLP. Among 315 patients with CHH without CLP, 32 patients (22 males) carried P or LP variants in FGFR1 (Supplementary Table S2). The identified FGFR1 variants included a range of deleterious types: missense, nonsense, frameshift, and splice-site-altering mutations. Inheritance could be evaluated in five cases, and four variants were confirmed as de novo (Supplementary Table S2, patients 19, 24, 26, and 30).
Phenotypically, 27 of the 32 patients were anosmic or hyposmic (Fig. 1A and Supplementary Table S2). Several individuals exhibited complex associated anomalies: patient 33 had agenesis of the septum pellucidum, consistent with a diagnosis of SOD in the context of CHH; dental agenesis was observed in patients 39 and 48; patient 24 had an atrial septal defect; patient 29 exhibited growth retardation; patient 49 had a diagnosis of autism spectrum disorder; and patient 47 presented with ear anomalies, type 2 diabetes, enuresis, and genu valgum (Supplementary Table S2).
For CHD7, the major causative gene for CHARGE syndrome, we identified P/LP variants in 10 patients with CHH (7 males) (Supplementary Table S3). These included eight missense variants, one nonsense (stop-gain) variant, and one splice-site variant. KS was diagnosed in seven of these individuals, while the remaining three were classified as having normosmic CHH (nCHH). Three patients (patients 54, 58, and 59) met clinical criteria for CHARGE syndrome. Other associated phenotypes among CHD7 variant carriers included hearing loss, SOD, and congenital cataracts.
Probands with CHH without CLP harboring deleterious variants in genes exclusively associated with CLP
In our cohort of 315 CHH patients without cleft, we found 4 patients (1 female) with heterozygous deleterious variants in genes exclusively associated with CLP—DLV3, PLCB4, NIPBL and EDNRA. Three variants were frameshifts leading to a premature stop codon, and one was splice-site altering (Table 2). Two patients had KS, and 2 patients were normosmic. As associated phenotypes, patient 12 had obesity and gynecomastia, and patient 13 had Cornelia de Lange Syndrome. The inheritance pattern was possible to be established only in patient 13, de novo. Pathogenic variants in these genes were previously associated with genetic syndromes (Table 3). Notably, three of these syndromes have clinical features in common with CHH, such as cryptorchidism, micropenis and hypoplasia of the corpus callosum.
Probands with CHH without CLP harboring digenic P/LP variants
We identified four patients with CHH in our cohort who carried more than one deleterious variant in the CLP gene panel and/or CHH gene panel (Table 4), suggesting possible digenic or oligogenic inheritance. In three of these cases, FGFR1 variants co-occurred with variants in TP63, TGFBR2, and INTS1, respectively. Patient 18 has compound heterozygous LP variants in PNPLA6 and an additional deleterious variant in PIEZO2. Regarding associated phenotypes: patient 16 exhibited dental agenesis; patient 17 presented with oxycephaly (craniosynostosis of the sagittal and coronal sutures); and patient 18 showed evidence of an empty sella on neuroimaging, along with vermis hypoplasia and ataxia.
Discussion
CHH presents with a wide spectrum of associated phenotypes, including well-defined syndromic forms, such as KS, SOD, and CHARGE syndrome [19]. This clinical heterogeneity reflects the shared embryological origins of the cranial placodes and the GnRH neuronal system, which contribute to both olfactory and neuroendocrine development [3, 19]. While midline craniofacial anomalies are reported in approximately 9% of patients with CHH [15], prior studies have not systematically distinguished between subtypes, such as cleft lip vs. cleft palate, bifid uvula, ectopic posterior pituitary, or midline brain anomalies, including agenesis of the corpus callosum and septum pellucidum.
In this study, we analyzed a large cohort of 336 individuals with CHH to investigate the frequency and spectrum of pathogenic variants in genes associated with CLP. Clinical features were thoroughly reviewed, and a curated CLP gene panel was applied to identify P or LP variants. “Overall, 60 patients with P/LP variants were identified in approximately 17.8% of the cohort, highlighting a significant genetic overlap between CHH and CLP.”
Among the 21 patients with CHH who presented with CLP (6%), 10 harbored P/LP variants in CLP-related genes, five of whom exhibited complex associated phenotypes. Notably, 8 of these 10 P/LP variants were in genes known to be implicated in both CHH and CLP. The most frequently implicated gene was FGFR1 (n = 7), followed by CHD7, SOX9, and CTNND1. Both FGFR1 and CHD7 are also CHH-related genes. In the remaining 315 CHH patients without CLP, we identified 54 P/LP variants in CLP-related genes in 50 individuals, including four cases suggestive of digenic inheritance. Across the full cohort with and without CLP, FGFR1 (n = 42) and CHD7 (n = 11) emerged as the most affected genes, reinforcing their central role in the overlapping genetic architecture of CHH and CLP. In the 10 families, it is possible to observe two phenomena: incomplete penetrance and variable clinical expressivity, the latter manifested as constitutional delay of growth and puberty (CPHD). Both events have been described by our group and others [5, 7].
FGFR1 is a pleiotropic gene involved in the development of GnRH neurons as well as in craniofacial and limb morphogenesis [34, 35]. This explains its association with diverse syndromes such as CHH, CLP, split-hand/foot malformation, and Hartsfield syndrome [36, 37]. Likewise, CHD7—the primary gene implicated in CHARGE syndrome—is known to contribute to both CHH and CLP phenotypes [10, 38]. Murine models have demonstrated that Chd7 plays a key developmental role in GnRH neuron migration and olfactory system formation, further supporting its relevance in the shared pathogenesis of these conditions [39, 40]. In two patients with CHH and CLP, deleterious variants were found in SOX9 and CTNND1. Although both are clear linked with the CLP, this is not the case for CHH. Both genes are involved in complex phenotypes. Of note, SOX9 in implicated in skeletal dysplasia, hearing loss and absence of the olfactory bulbs, resembling patient 9 [41]. Regarding CTNND1, patients can manifest tooth abnormalities as observed in our patient [42]. The pathophysiological mechanism underlying CHH associated with these genes remains to be established.
Of particular interest, we identified four patients with CHH without CLP who carried P/LP variants in genes previously associated exclusively with CLP, including DLV3, PLCB4, NIPBL, and EDNRA. These genes are known to underlie various congenital anomaly syndromes, some of which include reproductive features, such as micropenis or cryptorchidism, or neurodevelopmental anomalies such as corpus callosum hypoplasia—findings that partially overlap with the CHH phenotype [43,44,45]. One exception is EDNRA, where pathogenic variants cause mandibulofacial dysostosis with alopecia, a condition not typically linked to reproductive dysfunction [40, 46]. The contribution of these genes to the pathophysiology of CHH remains unclear, and functional studies are needed to determine whether they disrupt GnRH neuron development, pituitary function, or downstream hormonal regulation. Integrated approaches using transcriptomics and model organisms may help elucidate novel developmental links between craniofacial and reproductive systems.
Additionally, our observation of patients harboring multiple deleterious variants—including combinations of CHH- and CLP-related genes—supports an oligogenic model of inheritance. This may help explain the clinical variability.
Our findings underscore the importance of heightened clinical awareness among craniofacial teams, pediatricians, and geneticists regarding the potential reproductive and neuroendocrine sequelae in patients with CLP—especially those carrying variants in FGFR1 or CHD7. Because CLP is typically diagnosed at birth, clinical attention is often directed toward surgical and structural concerns, while endocrine abnormalities such as delayed or absent puberty may remain undetected until adolescence or adulthood. Early genetic screening of CLP patients for mutations in overlapping genes (i.e., FGFR1 and CHD7) may enable timely identification of individuals at risk for CHH, facilitating early endocrine evaluation and intervention. Moreover, future studies are warranted to assess the true prevalence of CHH within CLP cohorts, particularly in those with relevant genetic variants, to further define at-risk populations and optimize clinical management.
In conclusion, this study highlights a significant genetic and phenotypic overlap between CHH and CLP, driven in part by shared developmental pathways and pleiotropic genes, such as FGFR1 and CHD7. The identification of pathogenic variants in CLP-related genes—both in patients with CHH with and without craniofacial anomalies—supports a broader view of CHH as a developmental disorder with variable expressivity and potential oligogenic inheritance. These findings emphasize the need for integrated clinical and genetic screening strategies in both CHH and CLP populations to enable earlier diagnosis, improve genetic counseling, and guide personalized care.
Data availability
The data generated and analyzed during this study can be found within the published article and its supplementary files. Additional data is available from the corresponding author on reasonable request.
References
Fromantin M, Gineste J, Didier A, Rouvier J. [Impuberism and hypogonadism at induction into military service. Statistical study]. Probl Actuels Endocrinol Nutr. 1973;16:179–99.
Sykiotis GP, Pitteloud N, Seminara SB, Kaiser UB, Crowley WF Jr. Deciphering genetic disease in the genomic era: the model of GnRH deficiency. Sci Transl Med. 2010;2:32rv2.
Schwanzel-Fukuda M, Bick D, Pfaff DW. Luteinizing hormone-releasing hormone (LHRH)-expressing cells do not migrate normally in an inherited hypogonadal (Kallmann) syndrome. Brain Res Mol Brain Res. 1989;6:311–26.
Bianco SD, Kaiser UB. The genetic and molecular basis of idiopathic hypogonadotropic hypogonadism. Nat Rev Endocrinol. 2009;5:569–76.
Cangiano B, Swee DS, Quinton R, Bonomi M. Genetics of congenital hypogonadotropic hypogonadism: peculiarities and phenotype of an oligogenic disease. Hum Genet. 2021;140:77–111.
Young J, Xu C, Papadakis GE, Acierno JS, Maione L, Hietamaki J, et al. Clinical management of congenital hypogonadotropic hypogonadism. Endocr Rev. 2019;40:669–710.
Cassatella D, Howard S, Acierno J, Xu C, Papadakis G, Santoni FA, et al. Congenital hypogonadotropic hypogonadism and constitutional delay of growth and puberty have distinct genetic architectures. Eur J Endocrinol. 2018;178:377–88.
Costa-Barbosa FA, Balasubramanian R, Keefe KW, Shaw ND, Al-Tassan N, Plummer L, et al. Prioritizing genetic testing in patients with Kallmann syndrome using clinical phenotypes. J Clin Endocrinol Metab. 2013;98:E943–53.
Correa FA, Trarbach EB, Tusset C, Latronico AC, Montenegro LR, Carvalho LR, et al. FGFR1 and PROKR2 rare variants found in patients with combined pituitary hormone deficiencies. Endocr Connect. 2015;4:100–7.
Xu C, Cassatella D, van der Sloot AM, Quinton R, Hauschild M, De Geyter C, et al. Evaluating CHARGE syndrome in congenital hypogonadotropic hypogonadism patients harboring CHD7 variants. Genet Med. 2017;20:872-81.
Dixon MJ, Marazita ML, Beaty TH, Murray JC. Cleft lip and palate: understanding genetic and environmental influences. Nat Rev Genet. 2011;12:167–78.
Jugessur A, Murray JC. Orofacial clefting: recent insights into a complex trait. Curr Opin Genet Dev. 2005;15:270–8.
Quinton R, Duke VM, Robertson A, Kirk JM, Matfin G, de Zoysa PA, et al. Idiopathic gonadotrophin deficiency: genetic questions addressed through phenotypic characterization. Clin Endocrinol. 2001;55:163–74.
Bonomi M, Vezzoli V, Krausz C, Guizzardi F, Vezzani S, Simoni M, et al. Characteristics of a nationwide cohort of patients presenting with isolated hypogonadotropic hypogonadism (IHH). Eur J Endocrinol. 2018;178:23–32.
Federici S, Cangiano B, Goggi G, Messetti D, Munari EV, Amer M, et al. Genetic and phenotypic differences between sexes in congenital hypogonadotropic hypogonadism (CHH): large cohort analysis from a single tertiary centre. Front Endocrinol. 2022;13:965074.
Dodé C, Levilliers J, Dupont JM, De Paepe A, Le Dû N, Soussi-Yanicostas N, et al. Loss-of-function mutations in FGFR1 cause autosomal dominant Kallmann syndrome. Nat Genet. 2003;33:463–5.
Riley, Mansilla MA BM, Ma J, Daack-Hirsch S, Maher BS, Raffensperger LM, et al. Impaired FGF signaling contributes to cleft lip and palate. Proc Natl Acad Sci USA. 2007;104:4512–7.
Stanier P, Pauws E. Development of the lip and palate: FGF signalling. Front Oral Biol. 2012;16:71–80.
Boehm U, Bouloux PM, Dattani MT, de Roux N, Dode C, Dunkel L, et al. Expert consensus document: European consensus statement on congenital hypogonadotropic hypogonadism-pathogenesis, diagnosis and treatment. Nat Rev Endocrinol. 2015;11:547–64.
DePristo MA, Banks E, Poplin R, Garimella KV, Maguire JR, Hartl C, et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat Genet. 2011;43:491–8.
Kendig KI, Baheti S, Bockol MA, Drucker TM, Hart SN, Heldenbrand JR, et al. Sentieon DNASeq variant calling workflow demonstrates strong computational performance and accuracy. Front Genet. 2019;10:736.
gnomAD. Genome Aggregation Database 2014 [v4 data set (GRCh38): Available from: https://gnomad.broadinstitute.org/.
Rentzsch P, Schubach M, Shendure J, Kircher M. CADD-splice-improving genome-wide variant effect prediction using deep learning-derived splice scores. Genome Med. 2021;13:31.
Jaganathan K, Kyriazopoulou Panagiotopoulou S, McRae JF, Darbandi SF, Knowles D, Li YI, et al. Predicting splicing from primary sequence with deep learning. Cell. 2019;176:535–48.
Ioannidis NM, Rothstein JH, Pejaver V, Middha S, McDonnell SK, Baheti S, et al. REVEL: an ensemble method for predicting the pathogenicity of rare missense variants. Am J Hum Genet. 2016;99:877–85.
Cheng J, Novati G, Pan J, Bycroft C, Žemgulytė A, Applebaum T, et al. Accurate proteome-wide missense variant effect prediction with AlphaMissense. Science. 2023;381:eadg7492.
Wang K, Li M, Hakonarson H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic acids Res. 2010;38:e164.
Zhai J, Pitteloud N, Santoni FA. GenMasterTable: a user-friendly desktop application for filtering, summarising, and visualising large-scale annotated genetic variants. bioRxiv: 10.1101/2025.04.10.648172 [Preprint]. 2025 [cited 2025 Apr 16]. Available from https://doi.org/10.1101/2025.04.10.648172.
Martin AR, Williams E, Foulger RE, Leigh S, Daugherty LC, Niblock O, et al. PanelApp crowdsources expert knowledge to establish consensus diagnostic gene panels. Nat Genet. 2019;51:1560–5.
Kopanos C, Tsiolkas V, Kouris A, Chapple CE, Albarca Aguilera M, Meyer R, et al. VarSome: the human genomic variant search engine. Bioinformatics. 2019;35:1978–80.
Richards S, Aziz N, Bale S, Bick D, Das S, Gastier-Foster J, et al. Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet Med. 2015;17:405–24.
HGMD. The Human Gene Mutation Database Cardiff University. 1996. Available from: https://www.hgmd.cf.ac.uk/ac/index.php.
Sanger F, Nicklen S, Coulson AR. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA. 1977;74:5463–7.
Chung WCJ, Tsai PS. Role of fibroblast growth factor signaling in gonadotropin-releasing hormone neuronal system development. Front Horm Res. 2010;39:37–50.
Verheyden JM, Lewandoski M, Deng C, Harfe BD, Sun X. Conditional inactivation of Fgfr1 in mouse defines its role in limb bud establishment, outgrowth and digit patterning. Development. 2005;132:4235–45.
Villanueva C, Jacobson-Dickman E, Xu C, Manouvrier S, Dwyer AA, Sykiotis GP, et al. Congenital hypogonadotropic hypogonadism with split hand/foot malformation: a clinical entity with a high frequency of FGFR1 mutations. Genet Med. 2015;17:651–9.
Simonis N, Migeotte I, Lambert N, Perazzolo C, de Silva DC, Dimitrov B, et al. FGFR1 mutations cause Hartsfield syndrome, the unique association of holoprosencephaly and ectrodactyly. J Med Genet. 2013;50:585–92.
Vissers LE, van Ravenswaaij CM, Admiraal R, Hurst JA, de Vries BB, Janssen IM, et al. Mutations in a new member of the chromodomain gene family cause CHARGE syndrome. Nat Genet. 2004;36:955–7.
Layman WS, Hurd EA, Martin DM. Reproductive dysfunction and decreased GnRH neurogenesis in a mouse model of CHARGE syndrome. Hum Mol Genet. 2011;20:3138–50.
Layman WS, McEwen DP, Beyer LA, Lalani SR, Fernbach SD, Oh E, et al. Defects in neural stem cell proliferation and olfaction in Chd7 deficient mice indicate a mechanism for hyposmia in human CHARGE syndrome. Hum Mol Genet. 2009;18:1909–23.
Houston CS, Opitz JM, Spranger JW, Macpherson RI, Reed MH, Gilbert EF, et al. The campomelic syndrome: review, report of 17 cases, and follow-up on the currently 17-year-old boy first reported by Maroteaux et al. in 1971. Am J Med Genet. 1983;15:3–28.
Ghoumid J, Stichelbout M, Jourdain AS, Frenois F, Lejeune-Dumoulin S, Alex-Cordier MP, et al. Blepharocheilodontic syndrome is a CDH1 pathway-related disorder due to mutations in CDH1 and CTNND1. Genet Med. 2017;19:1013–21.
White JJ, Mazzeu JF, Hoischen A, Bayram Y, Withers M, Gezdirici A, et al. DVL3 alleles resulting in a -1 frameshift of the last exon mediate autosomal-dominant Robinow syndrome. Am J Hum Genet. 2016;98:553–61.
Kido Y, Gordon CT, Sakazume S, Ben Bdira E, Dattani M, Wilson LC, et al. Further characterization of atypical features in auriculocondylar syndrome caused by recessive PLCB4 mutations. Am J Med Genet A. 2013;161A:2339–46.
Boyle MI, Jespersgaard C, Brøndum-Nielsen K, Bisgaard AM, Tümer Z. Cornelia de Lange syndrome. Clin Genet. 2015;88:1–12.
Gordon CT, Weaver KN, Zechi-Ceide RM, Madsen EC, Tavares AL, Oufadem M, et al. Mutations in the endothelin receptor type A cause mandibulofacial dysostosis with alopecia. Am J Hum Genet. 2015;96:519–31.
Funding
This work was supported by the Swiss National Science Foundation (grant number IZCOZ0_182958) to Professor Nelly Pitteloud. WSD is supported by the NIHR Imperial Clinical Research Facility and NIHR Imperial Biomedical Research Centre (BRC) and an NIHR Senior Investigator Award (NIHR202371).
Author information
Authors and Affiliations
Contributions
FAC drafted the manuscript; acquired data; interpreted the results. IH helped write the manuscript; acquired data. JZ acquired data; provided analysis tools. MA acquired data: provided analysis tools. YW acquired data; provided analysis tools. AB acquired data; provided analysis tools. YZ acquired data; provided analysis tools. AR acquired data; revised the manuscript. SP acquired data; revised the manuscript. RQ acquired data; revised the manuscript. MB acquired data; revised the manuscript. BC acquired data; revised the manuscript. WSD acquired data; revised the manuscript. CEF acquired data; revised the manuscript. AN acquired data; revised the manuscript. PMB acquired data; revised the manuscript. JMF acquired data; revised the manuscript. DP acquired data; revised the manuscript. ZH acquired data; revised the manuscript. CPL acquired data; revised the manuscript. AM acquired data; revised the manuscript. NJN helped write the manuscript; interpreted the results, acquired data; revised the manuscript. FS interpreted the results; revised the manuscript. JSA interpreted the results; helped write the manuscript; revised the manuscript. NP conceived and designed the work; interpreted the results; revised the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
The study was approved by the Institutional Review Board–Commission cantonale (VD) D’Éthique de la recherche (CER-VD)–number of the project: PB_2018-00247. I confirm that informed consent was obtained from all subjects, including for publication of individual patient data.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Correa, F.d.A., Habibi, I., Zhai, J. et al. Pleiotropic genes linking congenital hypogonadotropic hypogonadism and cleft lip/palate: evidence from a genomic CHH cohort study. Eur J Hum Genet 34, 340–347 (2026). https://doi.org/10.1038/s41431-025-02005-6
Received:
Revised:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1038/s41431-025-02005-6
