
Abstract
In eucaryotes, mitogen-activated protein kinase (MAPK) cascades are evolutionarily conserved signalling modules crucial for growth regulation and stress tolerance. However, the regulatory role of MAPK in nutrient sensing by plants remains largely unclear. Here we uncovered MEKK14 and its paralogue MEKK13 determine lateral root elongation via enhanced cell division and expansion. We further fine-mapped a naturally occurring histidine-to-glutamine substitution in MEKK14 that weakens protein kinase activity and attenuates lateral root growth and response to nitrate (NO3−). We further demonstrate that NO3− transcriptionally upregulates MEKK13/14 depending on NLP7 to activate a MKK3–MPK1/2/7/14 signalling module. Downstream of this signalling cascade, the core oscillator of the circadian clock CCA1 is phosphorylated and stabilized to feedback induce MEKK13/14 expression and to activate auxin signalling-dependent lateral root foraging for NO3−. Our findings reveal a positive-feedback phosphorylation–transcriptional regulatory loop in root NO3− foraging, extending the regulatory function of MAPK signalling in the nutrient sensing.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$32.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on SpringerLink
- Instant access to the full article PDF.
USD 39.95
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others
Data availability
Data are available within the Article and the Supplementary Information. Additional datasets are available from the corresponding author upon request. Source data are provided with this paper.
References
Wang, Y. Y., Cheng, Y. H., Chen, K. E. & Tsay, Y. F. Nitrate transport, signaling, and use efficiency. Annu. Rev. Plant Biol. 69, 85–122 (2018).
Liu, K. H. & Sheen, J. Dynamic nutrient signaling networks in plants. Annu. Rev. Plant Biol. 37, 341–367 (2021).
Liu, K. H. et al. Discovery of nitrate-CPK-NLP signalling in central nutrient-growth networks. Nature 545, 311–316 (2017).
Marchive, C. et al. Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat. Commun. 4, 1713 (2013).
Alvarez, J. M. et al. Transient genome-wide interactions of the master transcription factor NLP7 initiate a rapid nitrogen-response cascade. Nat. Commun. 11, 1157 (2020).
Ho, C. H., Lin, S. H., Hu, H. C. & Tsay, Y. F. CHL1 functions as a nitrate sensor in plants. Cell 138, 1184–1194 (2009).
Liu, K. H. et al. NIN-like protein 7 transcription factor is a plant nitrate sensor. Science 377, 1419–1425 (2022).
Jia, Z., Giehl, R. F. H. & von Wirén, N. Nutrient-hormone relations: driving root plasticity in plants. Mol. Plant 15, 86–103 (2022).
Jia, Z. & von Wirén, N. Signaling pathways underlying nitrogen-dependent changes in root system architecture: from model to crop species. J. Exp. Bot. 71, 4393–4404 (2020).
Liu, Y. & von Wirén, N. Integration of nutrient and water availabilities via auxin into the root developmental program. Curr. Opin. Plant Biol. 65, 102117 (2021).
Gifford, M. L., Dean, A., Gutierrez, R. A., Coruzzi, G. M. & Birnbaum, K. D. Cell-specific nitrogen responses mediate developmental plasticity. Proc. Natl Acad. Sci. USA 105, 803–808 (2008).
Vidal, E. A. et al. Nitrate-responsive miR393/AFB3 regulatory module controls root system architecture in Arabidopsis thaliana. Proc. Natl Acad. Sci. USA 107, 4477–4482 (2010).
Krouk, G. et al. Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev. Cell 18, 927–937 (2010).
Drew, M. C. Comparison of effects of a localized supply of phosphate, nitrate, ammonium and potassium on growth of seminal root system and shoot in barley. New Phytol. 75, 479–490 (1975).
Zhang, H. & Forde, B. G. An Arabidopsis MADS box gene that controls nutrient-induced changes in root architecture. Science 279, 407–409 (1998).
Remans, T. et al. The Arabidopsis NRT1.1 transporter participates in the signaling pathway triggering root colonization of nitrate-rich patches. Proc. Natl Acad. Sci. USA 103, 19206–19211 (2006).
Vidal, E. A., Moyano, T. C., Riveras, E., Contreras-López, O. & Gutiérrez, R. A. Systems approaches map regulatory networks downstream of the auxin receptor AFB3 in the nitrate response of Arabidopsis thaliana roots. Proc. Natl Acad. Sci. USA 110, 12840–12845 (2013).
Poitout, A. et al. Responses to systemic nitrogen signaling in Arabidopsis roots involve trans-Zeatin in shoots. Plant Cell 30, 1243–1257 (2018).
Ruffel, S. et al. Nitrogen economics of root foraging: transitive closure of the nitrate-cytokinin relay and distinct systemic signaling for N supply vs. demand. Proc. Natl Acad. Sci. USA 108, 18524–18529 (2011).
Jia, Z., Giehl, R. F. H. & von Wirén, N. Local auxin biosynthesis acts downstream of brassinosteroids to trigger root foraging for nitrogen. Nat. Commun. 12, 5437 (2021).
Jia, Z., Giehl, R. F. H., Meyer, R. C., Altmann, T. & von Wirén, N. Natural variation of BSK3 tunes brassinosteroid signaling to regulate root foraging under low nitrogen. Nat. Commun. 10, 2378 (2019).
Jia, Z., Giehl, R. F. H. & von Wirén, N. The root foraging response under low nitrogen depends on DWARF1-mediated brassinosteroid biosynthesis. Plant Physiol. 183, 998–1010 (2020).
Zhang, M. & Zhang, S. Mitogen-activated protein kinase cascades in plant signaling. J. Integr. Plant Biol. 64, 301–341 (2022).
Sun, T. & Zhang, Y. MAP kinase cascades in plant development and immune signaling. EMBO Rep. 23, e53817 (2022).
Shao, Y. M. et al. The YDA-MKK4/MKK5-MPK3/MPK6 cascade functions downstream of the RGF1-RGI ligand-receptor pair in regulating mitotic activity in root apical meristem. Mol. Plant 13, 1608–1623 (2020).
Lu, X. T. et al. RGF1-RGI1, a peptide-receptor complex, regulates Arabidopsis root meristem development via a MAPK signaling cascade. Mol. Plant 13, 1594–1607 (2020).
Zhu, Q. K. et al. A MAPK cascade downstream of IDA–HAE/HSL2 ligand–receptor pair in lateral root emergence. Nat. Plants 5, 414–423 (2019).
Yan, Z. W. et al. MPK3/6-induced degradation of ARR1/10/12 promotes salt tolerance in Arabidopsis. EMBO Rep. 22, e52457 (2022).
Zhou, F. L. et al. The MEKK1-MKK1/2-MPK4 cascade phosphorylates and stabilizes STOP1 to confer aluminum resistance in Arabidopsis. Mol. Plant 16, 337–353 (2023).
Sözen, C. et al. Wounding and insect feeding trigger two independent MAPK pathways with distinct regulation and kinetics. Plant Cell 32, 1988–2003 (2020).
Canales, J., Moyano, T. C., Villarroel, E. & Gutiérrez, R. A. Systems analysis of transcriptome data provides new hypotheses about Arabidopsis root response to nitrate treatments. Front. Plant Sci. 5, 22 (2014).
Blom, N., Gammeltoft, S. & Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 294, 1351–1362 (1999).
Obenauer, J. C., Cantley, L. C. & Yaffe, M. B. Scansite 2.0: proteome-wide prediction of cell signaling interactions using short sequence motifs. Nucleic Acids Res. 31, 3635–3641 (2003).
Colcombet, J., Sözen, C. & Hirt, H. Convergence of multiple MAP3Ks on MKK3 identifies a set of novel stress MAPK modules. Front. Plant Sci. 7, 1941 (2016).
Widmann, C., Gibson, S., Jarpe, M. B. & Johnson, G. L. Mitogen-activated protein kinase: conservation of a three-kinase module from yeast to human. Physiol. Rev. 79, 143–180 (1999).
Efeyan, A., Comb, W. C. & Sabatini, D. M. Nutrient-sensing mechanisms and pathways. Nature 517, 302–310 (2015).
Wang, H. et al. Regulatory functions of cellular energy sensor SnRK1 for nitrate signalling through NLP7 repression. Nat. Plants 8, 1094–1107 (2022).
Chardin, C., Schenk, S. T., Hirt, H., Colcombet, J. & Krapp, A. Review: mitogen-activated protein kinases in nutritional signaling in Arabidopsis. Plant Sci. 260, 101–108 (2017).
Voß, U. et al. The circadian clock rephases during lateral root organ initiation in Arabidopsis thaliana. Nat. Commun. 6, 7641 (2015).
Lv, B. et al. MPK14-mediated auxin signaling controls lateral root development via ERF13-regulated very-long-chain fatty acid biosynthesis. Mol. Plant 14, 285–297 (2021).
Chen, X. et al. The MKK3-MPK7 cascade phosphorylates ERF4 and promotes its rapid degradation to release seed dormancy in Arabidopsis. Mol. Plant 16, 1743–1758 (2023).
Zhang, L. et al. Mitogen-activated protein kinases MPK3 and MPK6 phosphorylate receptor-like cytoplasmic kinase CDL1 to regulate soybean basal immunity. Plant Cell 36, 963–986 (2024).
Sarma, U. & Ghosh, I. Oscillations in MAPK cascade triggered by two distinct designs of coupled positive and negative feedback loops. BMC Res. Notes 5, 287 (2012).
Jia, Z. et al. A spatially concerted epidermal auxin signaling framework steers the root hair foraging response under low nitrogen. Curr. Biol. 33, 3926–3941 (2023).
Brady, S. M. et al. Quantitative modeling of the short-term response to nitrogen availability that coordinates early events in lateral root initiation. Preprint at bioRxiv https://doi.org/10.1101/2023.12.05.570292 (2023).
Gutiérrez, R. A. et al. Systems approach identifies an organic nitrogen-responsive gene network that is regulated by the master clock control gene CCA1. Proc. Natl Acad. Sci. USA 105, 4939–4944 (2008).
Zhang, S. N. et al. Nitrogen mediates flowering time and nitrogen use efficiency via floral regulators in rice. Curr. Biol. 31, 671–683 (2021).
Heisler, M. G. et al. Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem. Curr. Biol. 15, 1899–1911 (2005).
Friml, J. et al. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 426, 147–153 (2003).
Yuan, L. et al. BBX19 fine-tunes the circadian rhythm by interacting with PSEUDO-RESPONSE REGULATOR proteins to facilitate their repressive effect on morning-phased clock genes. Plant Cell 33, 2602–2617 (2021).
Gruber, B. D., Giehl, R. F., Friedel, S. & von Wirén, N. Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiol. 163, 161–179 (2013).
Kang, H. M. et al. Efficient control of population structure in model organism association mapping. Genetics 178, 1709–1723 (2008).
Atwell, S. et al. Genome-wide association study of 107 phenotypes in Arabidopsis thaliana inbred lines. Nature 465, 627–631 (2010).
Horton, M. W. et al. Genome-wide patterns of genetic variation in worldwide Arabidopsis thaliana accessions from the RegMap panel. Nat. Genet. 44, 212–216 (2012).
Lampropoulos, A. et al. GreenGate—a novel, versatile, and efficient cloning system for plant transgenesis. PLoS ONE 8, e8304 (2013).
Zhang, X. Y. et al. Mediator subunit MED31 is required for radial patterning of Arabidopsis roots. Proc. Natl Acad. Sci. USA 115, E5624–E5633 (2018).
Acknowledgements
We thank J. Fuge and A. Bieber (Leibniz Institute of Plant Genetics and Crop Plant Research, Gatersleben) for excellent technical assistance; Z. Ding (Shandong University, China) and Y. Xiang (Chinese Academy of Agricultural Sciences) for sharing mpk1,2,14 and mpk7 mutants. This work was supported by grants to Z.J. by Biological Breeding-National Science and Technology Major Project (2023ZD04072), National Natural Science Foundation of China (32370270), Disciplinary Breakthroughs Project of Ministry of Education (JYB2025XDXIM702), Chinese Universities Scientific Fund (2023RC052, 1031-00114341, 2025RC027) and Pinduoduo-China Agricultural University Research Fund (PC2024B01009).
Author information
Authors and Affiliations
Contributions
Z.J., R.F.H.G. and N.v.W. conceived the project and designed the experiments. X.Z., S.Z. and Z.J. performed experiments; J.G. genotyped the mpk mutants; L.Y. and F.Z. contributed to helpful discussions; X.X. provided critical plant materials. Z.J., R.F.H.G. and N.v.W. wrote the manuscript with the support of X.X., L.Y. and M.J.B.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Plants thanks the anonymous reviewers for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
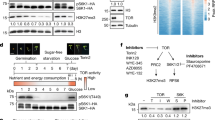
Extended Data Fig. 1 His to Gln substitution at position 172 does not impact the interaction of MEKK14 proteoforms with MKK3.
Y2H assay showing that MEKK14172H and MEKK14172Q interact similarly with MKK3. SD/-Leu-Trp, synthetic defined (SD) medium lacking Leu and Trp; SD/-Leu-Trp-His-Ade, SD medium lacking Leu, Trp, His and Ade.
Extended Data Fig. 2 Expression pattern of MEKK14 and MEKK13.
a-b Overview of promoter::GUS activity of MEKK14 (a) and MEKK13 (b) in 10-day-old light-grown seedlings. c-d Promoter activity of MEKK14 (c) and MEKK13 (d) in primary root tips. e Expression pattern of MEKK13 in early-stage LR primordia (i), LR primordia close to emergence (ii) and emerged LR (iii). 10-day-old seedlings grown on 11.4 mM N were sampled and subjected to GUS staining for 1 hour. Scale bars, 1 cm in a-b and 100 μm in c-e.
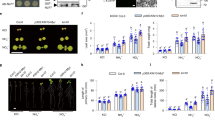
Extended Data Fig. 3 MEKK14 allelic variants determine phenotypic variation of NO3−-dependent LR growth.
a-c Average LR length of accessions from 172H haplotype (Col-0, Sha, Yo-0, Ty-1 and Rmx-A02) and 172Q haplotype (Ra-0, Uod-1, Ren-1, Pog-0, Van-0, Sg-1, Per-1, Wa-0, Ws-2 and Est-0) plants grown under 2 mM KCl (a) or KNO3 (b) and appearance of plants from representative genotypes (c). Symbols with crossbars represent means ± s.e.m. Individual symbols represent averaged LR length of respective accessions assessed from 6-10 independent plants. In a and b, n = 5 and 10 for 172H and 172Q haplogroup, respectively. d-e Appearance of plants (d), and average LR length (e) of wild types (Col-0, Sha, Ra-0 and Uod-1) and mekk14 crispr mutant plants grown on either 2 mM KCl or KNO3. Symbols with crossbars represent means ± s.e.m. Number of plants analyzed in KCl and KNO3: n = 16/14 (Col-0), 16/14 (cr2_Col-0), 16/14 (cr7_Col-0), 16/15 (Sha), 15/13 (cr7_Sha), 15/15 (cr10_Sha),16/14 (Ra-0), 16/16 (cr1_Ra-0), 16/16 (cr2_Ra-0), 16/15 (Uod-1), 15/16 (cr8_ Uod-1), 16/15 (cr11_Col-0), respectively. Scale bar, 1 cm. Different letters indicate significant differences at P < 0.01 according to one-way ANOVA and post-hoc Tukey’s test.
Extended Data Fig. 4 Loss-of-function of MKK3 reduces meristem size and cell length under NO3−.
a-b. Representative confocal images of root meristems (a) and mature cells (b) of Col-0 and mkk3 LRs grown on 2 mM KCl or KNO3. Red lines in a and arrowheads in b indicate meristem sizes (a) and the boundaries between two consecutive mature cortical cells (b). Scale bars, 100 μm. c-d Length of the meristems (c) and cortical cells (d) of LRs from Col-0 and mkk3 plants. Number of roots analyzed in KCl/KNO3: n = 12/12, 12/12 and 12/12 in e and 72/72, 72/72 and 72/72 in f for Col-0, mkk3-1 and mkk3-2, respectively. Blue symbols with crossbars represent mean ±s.e.m. Different letters indicate significant differences at P < 0.001 according to one-way ANOVA and post-hoc Tukey’s test.
Extended Data Fig. 5 MPK1, 2, and 14 single mutants exhibit inhibited lateral root elongation.
a-b Appearance of plants (a) and average LR length (b) of wild-type (Col-0), mpk1-1, mpk1-2, mpk14-1, mpk14-2 and mpk2-2 mutant plants grown on 2 mM KCl or KNO3. Scale bars, 1 cm. Number of roots analyzed in KCl/KNO3: n = 20/20, 19/19, 19/20,19/19, 20/20 and 19/20 for Col-0, mpk1-1, mpk1-2, mpk14-1, mpk14-2 and mpk2-2, respectively. c and e. Representative confocal images of root meristems (c) and mature cells (e) of Col-0 and mpk LRs. Red lines in c and arrowheads in e indicate meristem sizes (c) and the boundaries between two consecutive mature cortical cells (e). Scale bars, 100 μm. d and f. Meristem size (d) and cortical cell length (f) of LRs from Col-0 and mkks plants. Number of roots or cells analyzed in KCl/KNO3: n = 12/12, 12/12 and 12/12 in d and 72/72, 72/72, 72/72 and 72/72 in f for Col-0, mkk3-1 and mkk3-2, respectively. Blue symbols with crossbars represent mean ± s.e.m. P values related to differences between respective T-DNA KO and wild-type Col-0 were calculated according to Welch’s t-test.
Extended Data Fig. 6 Multiple deletion lines of MPK1/2/7/14 have decreased meristem size and cell length under NO3−.
a-b. Representative confocal images of root meristems (a) and mature cells (b) of Col-0, tko and qko (#1 and #2) LRs grown on 2 mM KCl or KNO3. Red lines in a and arrowheads in b indicate meristem sizes (a) and the boundaries between two consecutive mature cortical cells (b). c-d Meristem size (c) and cortical cell length (d) of LRs from Col-0, tko and qko (#1 and #2) plants. Number of LR analyzed in KCl/ KNO3: n = 13/12 (Col-0), 13/12 (tko), 15/10 (qko-#1) and 15/13 (qko-#2) in c; n = 72/76 (Col-0), 65/64 (tko), 70/60 (qko-#1), 66/63 (qko-#2) in d, respectively. Blue symbols with crossbars represent means ± s.e.m. Different letters indicate significant differences at P < 0.05 according to one-way ANOVA and post-hoc Tukey’s test.
Extended Data Fig. 7 NO3− activates a MEKK14-MKK3-MPK1/2/7/14 phospho-relay to promote LR growth.
a MEKK13/14- and MKK3-dependent activation of MPK1/2/7/14 phosphorylation by NO3−. 7-day-old seedlings precultured on high N were subjected to 4 days N starvation and then exposed to 10 mM KNO3. Recombinant GST-tagged MPK1/2/7/14 protein was incubated with total protein extracts from wild-type (Col-0), mekk13/14 and mkk3 seedlings for 1-4 h of nitrate treatment. Abundance of GST-MPK1/2/7/14 and phosphorylated GST-MPK1/2/7/14 proteins was assessed with Coomassie Brilliant Blue staining (CBB) and anti-thiophosphate ester-specific antibody (anti-TPE). Actin was used as internal control for equal protein loading. The band intensity of phosphorylated MPK was quantified with Image J (version 1.53e) and normalized to Actin with time 0 in Col-0 samples set to 1.0. b-c Appearance of plants (b), and average LR length (c) of wild-type (Col-0), mekk13/14 and two representative T3 transgenic lines expressing GFP-tagged MKK3DD under MEKK14 promoter grown under 2 mM KCl and KNO3. Number of plants analyzed in KCl/KNO3: n = 16/16, 16/15, 16/15 and 15/15 for Col-0, mekk14, MKK3DD-#1 and MKK3DD-#2, respectively. d-e Appearance of plants (d), and average LR length (e) of wild-type (Col-0), mkk3-2 and two representative T3 transgenic lines expressing GFP-tagged MPKsDD under MKK3 promoter grown under 2 mM KCl and KNO3. Number of plants analyzed in KCl/KNO3: n = 16/16, 16/16, 16/16, 15/15, 15/16, 16/15, 16/16, 16/15, 16/14 and 16/16 for Col-0, mkk3-2, MPK1DD-#1 and MPK1DD-#2, MPK2DD-#1, MPK2DD-#2, MPK7DD-#1, MPK7DD-#2, MPK14DD-#1 and MPK14DD-#2, respectively. Scale bars, 1 cm. Different letters indicate significant differences at P < 0.01 according to one-way ANOVA and post-hoc Tukey’s test.
Extended Data Fig. 8 CCA1 interacts with MPK1/2/7/14 in yeast two-hybrid and split luciferase complementation imaging assays.
a-d Y2H assay showing CCA1 interaction with MPK1 (a), MPK2 (b), MPK7 (c) and MPK14 (d). Co-transformation of CCA1-BD with empty AD or AD-MPK1/2/7/14, or of AD-MPK1/2/7/14 with empty BD vector. AD and BD vectors served as negative controls. SD/-Leu-Trp, synthetic defined (SD) medium lacking Leu and Trp; SD/-Leu-Trp-His-Ade, SD medium lacking Leu, Trp, His, and Ade. e-h Split-LUC complementation imaging assay in N. benthamiana testing interaction between CCA1 and MPK1 (e), MPK2 (f), MPK7 (g) and MPK14 (h). Indicated constructs were transiently expressed in N. benthamiana by Agrobacterium-mediated transformation, and the GUS (β-glucuronidase) protein served as negative controls. nLUC, N-terminal fragment of firefly luciferase; cLUC, C-terminal fragment of firefly luciferase.
Extended Data Fig. 9 Exogenous auxin supply restores the LR growth response to NO3−.
a-b Appearance of plants (a) and average LR length (b) of wild-type (Col-0) and cca1-1 mutant plants grown on 2 mM KCl or KNO3 supplied with or without 20 nM IAA. c-d Appearance of plants (c) and average LR length (d) of wild-type (Col-0), mekk13/14, mkk3-2 and qko-#1 mutant plants grown on 2 mM KCl or KNO3 supplied with or without 20 nM IAA. Scale bars, 1 cm. Symbols with crossbars represent means ± s.e.m. n = 17 and 16 for Col-0 and cca1-1 under respective N and auxin treatment in b; n = 16 for respective treatment and genotype in d. Different letters indicate significant differences at P < 0.001 according to one-way ANOVA and post-hoc Tukey’s test.
Extended Data Fig. 10 External auxin supply to LRs in a split-root system restores the LR growth response of cca1-1 to NO3− without penalizing primary root growth.
a Schematic representation of the spilt-agar experiment with auxin treatment. Col-0 and cca1-1 mutant were firstly precultured on 1/2 MS medium identical to sufficient N (11.4 mM) treatment but with N supplied exclusively as 1 mM Gln. After 6 days, seedlings of homogenous size from each genotype were transferred to either 2 mM KCl or KNO3 for 5 days enabling LR outgrowth and phenotypic characterization shown in figure panel c and d. Then, these seedlings were transferred to split-agar plates, so that only the branching zone containing emerged LRs was exposed to 10 or 20 nM IAA. After 3 days, roots were scanned and subjected to quantification of the length of individual LRs and primary root. b-d Appearance of plants (b), average LR length (c) and primary root length (d) of wild-type (Col-0) and cca1-1 mutant plants grown under either 2 mM KCl or KNO3 in the presence or absence of IAA in the split-agar plates for 3 days. Only LRs grown in the auxin-supplied compartments were considered for quantification. Scale bars, 1 cm. Number of roots analyzed given in KCl/KNO3 in c and d: n = 9/8 and 8/8 for Col-0 and cca1-1 in panel DAT5; 9/8 and 9/8 for Col-0 and cca1-1 in panel mock; 8/8 and 8/8 for Col-0 and cca1-1 in panel 10 nM IAA; 8/8 and 8/8 for Col-0 and cca1-1 in panel 20 nM IAA. Symbols with crossbars represent mean ± s.e.m. Different letters indicate significant differences at P < 0.01 according to one-way ANOVA and post-hoc Tukey’s test.
Supplementary information
Supplementary Information
Supplementary Figs. 1–12.
Supplementary Tables
Supplementary Tables 1 and 2.
Supplementary Data
Source data for Supplementary Figs. 1, 4, 5, 7, 10 and 12.
Source data
Source Data Figs. 1–7 and Extended Data Fig. 1–10
Statistical source data.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Zhang, X., Zhou, S., Guo, J. et al. A feedback regulatory loop by MAPK–CCA1 engages auxin signalling to stimulate root foraging for nitrate. Nat. Plants (2026). https://doi.org/10.1038/s41477-026-02225-8
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41477-026-02225-8
