
Abstract
Alcohol-associated liver disease (ALD), characterized by gut barrier disruption and microbial dysbiosis, is associated with significant depletion of the genus Bifidobacterium in patients, as evidenced by our cohort of 127 subjects. Functional screening revealed B. pseudocatenulatum as a protective strain. In a murine ALD model established with a Lieber–DeCarli ethanol diet, oral administration of B. pseudocatenulatum for 8 weeks ameliorated hepatomegaly, steatosis, and serum transaminase levels. Probiotic intervention restored intestinal barrier function, as indicated by reduced lipopolysaccharide-binding proteins and upregulated tight junction protein expression. Microbiome analysis revealed a mitigation of dysbiosis, with a reduction in pathogenic Escherichia-Shigella and Parabacteroides and an enrichment of beneficial Bifidobacterium and Blautia, concomitant with shifts in lipid metabolism. Mechanistically, B. pseudocatenulatum-derived short-chain fatty acids downregulated the expression of hepatic lipogenic genes (Cd36, Fasn, Accα) and pro-inflammatory cytokines (Il-1β, Ccl2, Tnf-α). These results suggest that B. pseudocatenulatum is a promising probiotic candidate for ALD management.
Similar content being viewed by others


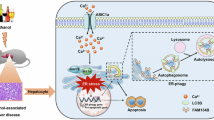
Introduction
Alcohol-associated liver disease (ALD) is a common and widespread chronic liver disease that includes three histologic stages—liver steatosis, alcoholic hepatitis and cirrhosis—and leads to 3.3 million deaths annually1,2. Accumulating evidence suggests that gut leakage components should not be neglected3 as additional potential factors3, given that hepatic immune cells respond to pathogen-associated molecular patterns and produce damage-associated molecular patterns.
Disruption of intestinal barrier function is an essential factor that promotes the onset and progression of ALD. The functional intestinal barrier comprises a mucus layer, an epithelial monolayer cell barrier, and mucosal immune cells that prevent bacterial infiltration and product penetration4. Bacteria, fungi, archaea and viruses are important components of the gut microbiota5,6,7. The detrimental effects of ethanol on the gut‒liver axis are well established and are mediated primarily through the disruption of intestinal barrier integrity and chronic gut microbiota dysbiosis8,9. Ethanol and its metabolites directly impair intestinal tight junctions, compromising the gut barrier. This breach facilitates bacterial translocation and alters the gut milieu, thereby driving microbial dysbiosis—which is characterized by reduced microbial diversity, a decline in beneficial taxa (such as Akkermansia, Bacteroides and the butyrate producer Faecalibacterium), and an expansion of potentially pathogenic ones (including Streptococcus, Proteobacteria, and Enterobacteriaceae). These alterations in microbial community structure and metabolic output drive the progression of liver inflammation6,10,11,12,13.
Probiotics have beneficial effects on liver function6,14. Bifidobacterium is the main genus that improves nutrient absorption and activates host immunity15. Notably, the reduction in short-chain fatty acids (SCFAs) caused by alcohol consumption can negatively affect gut health. Bifidobacterium, an SCFA-producing bacterium, may be beneficial for the maintenance of the gut microbiota, intestinal barrier and pathogenesis of ALD, making it a potentially useful probiotic to promote a healthy microbiota balance6. Many studies have explored the probiotic effect of B. pseudocatenulatum. For example, supplementation with a B. pseudocatenulatum strain in obese mice has been shown to significantly attenuate intestinal inflammatory responses16. B. pseudocatenulatum CECT 7765 administration attenuated colitis, CCl-4-induced liver fibrosis and immune-related diseases, enhancing the gut barrier and eliminating bacterial translocation and endotoxin levels in serum. The strain ameliorated the imbalance of the gut microbiota, further increasing the abundance of probiotics and suppressing harmful bacteria17. Although B. pseudocatenulatum has emerged as a promising therapeutic candidate for modulating the gut microbiota, its specific therapeutic potential and molecular mechanisms in ALD pathogenesis remain to be elucidated. In this investigation, the hepatoprotective effects of human-derived B. pseudocatenulatum were systematically evaluated through a multiomics approach involving 16S rRNA sequencing, metabolomic profiling, and histopathological analysis. Our experimental design is schematically presented in Fig. 1.

Schematic representation of the experimental design for restoring Bifidobacterium pseudocatenulatum to ameliorate alcohol-associated liver disease.
Results
Alcohol-induced microbiome dysbiosis and Bifidobacterium pseudocatenulatum isolated from healthy human feces alleviate alcohol-associated liver injury
Subject inclusion and exclusion were conducted in accordance with the European Association for the Study of the Liver (EASL) Clinical Practice Guidelines for the Management of ALD. 51 ALD patients, 35 healthy subjects and 41 patients with nonmetabolic liver disease (hepatitis B virus, HBV) were recruited and age- and BMI-matched. The mean ages of the participants were 56.23 years (ALD group), 53.53 years (HBV group) and 55.16 years (healthy group, HC), with a normal range of BMI. Ethanol exposure resulted in elevated levels of liver injury parameters, including alanine transaminase (ALT) and aspartate aminotransferase (AST). Bilirubin-related indices, such as total bilirubin (TB), direct bilirubin (DBIL), and indirect bilirubin (IBIL), were elevated in ALD patients (P < 0.01). The levels of nutritional status-related indicators, namely, albumin, high-density lipoprotein (HDL), and low-density lipoprotein (LDL), were greater in the HC group than in the ALD group (Table 1), indicating better nutritional status in the healthy group. The aforementioned parameters did not significantly differ between the ALD and HBV groups. The levels of multiple cytokines and chemokines significantly differed in patients with ALD (Table 2).
16S rRNA sequencing was performed, and we found that ethanol intake induced microbiome dysbiosis. The overall microbial diversity and community richness, as calculated by the Chao1 index, observed species, and coverage of goods, significantly differed across the experimental groups between the ALD and HC groups, but no difference was observed between the ALD and HBV groups (Fig. 2A). On the basis of the UniFrac distance, our analysis using the unweighted method revealed distinct separations in the principal coordinate analysis (PCoA) plot (PERMANOVA, R2 = 0.0515; P = 0.001). Specifically, the HC group and ALD group exhibited significant microbial community separation (ANOSIM unweighted, r = 0.142, P = 0.001; Adonis unweighted, P < 0.01). Even when disease severity was comparable between the HBV and ALD groups, their microbial communities remained significantly distinct (ANOSIM unweighted, r = 0.0799, P = 0.001; Adonis unweighted, P < 0.01). PCoA was employed to elucidate structural variations in the microbial communities. Under the unweighted algorithm, PC1 accounted for 12% of the total variation, and PC2 explained 4% (Fig. 2A). In addition, alcohol significantly reduced the relative abundance of the genus Bifidobacterium in humans, which was correlated with several clinical parameters (Fig. 2B). Positive associations were observed with hepatic reserve markers such as cholinesterase, nutrition-related indicators such as albumin and cholesterol, osteocalcin—a therapeutic agent targeting metabolic disorders—and the inflammatory cytokine TNFsf12 (all P < 0.01)18. The observed reduction in TNFsf12 expression in ALD patients may result from bacteria modulation, potentially impairing immunomodulatory and tissue repair processes19,20,21. Furthermore, there was a negative correlation with liver pathophysiology indicators such as AST, γ-GT, total bilirubin and CHI3L1 (P < 0.05, P < 0.05, P < 0.01, P < 0.01) (Fig. 2C)3.

A Alpha diversity and unweighted UniFrac PCoA of the gut microbiota in the healthy control (HC), alcohol-associated liver disease (ALD), and nonmetabolic liver disease (HBV) groups. In PCoA, box plots are superimposed above and to the right of the main plot, displaying the distribution of samples along the respective principal coordinates. Different lowercase letters on the box plots indicate statistically significant differences in beta diversity distances between groups based on Tukey’s HSD test. The results of the PERMANOVA are shown in the upper right corner, indicating the proportion of community structure variation explained by the grouping (N: HC = 32, ALD = 35, and HBV = 39). B MASLIN2 analysis of the differentially enriched genus Bifidobacterium between groups (N: HC = 32, ALD = 35, and HBV = 39). C Spearman correlation analysis between the genus Bifidobacterium and clinical characteristics/cytokines. The asterisks indicate the statistical significance of the correlation coefficients. The final sample sizes for the correlation analysis reflect participants with matched blood biochemical indices and fecal microbiome sequencing data (N: HC = 28, ALD = 35). D Functional differences of Bifidobacterium species: Animal experiment schematic of the chronic-binge ethanol feeding model (NIAAA model). The mice were gradually adapted to a 5% (vol/vol) ethanol Lieber–DeCarli diet for 1 week and maintained for 8 weeks. The mice were subsequently administered a single binge of ethanol (5 g/kg body weight) and sacrificed 9 h later. Different viable bacterial preparations were administered concurrently via oral gavage (N = 8 per group). E Efficacy evaluation of the intervention with three different viable bacterial preparations. F KEGG functional classification of the Bifidobacterium pseudocatenulatum genome. The data were analyzed using one-way ANOVA followed by Dunnett’s T3 test. All the data are presented as the mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001.
We successfully isolated three Bifidobacterium strains—Bifidobacterium longum, Bifidobacterium pseudocatenulatum and Bifidobacterium adolescentis—from healthy human feces. A functional screening approach was employed to identify the most promising candidate for ameliorating ALD (Fig. 2D). Among the tested strains, only B. pseudocatenulatum significantly reduced both serum hepatic enzymes (ALT and AST) and hepatic triglyceride levels. In contrast, B. longum decreased ALT and AST but did not markedly affect hepatic lipids, whereas B. adolescentis showed no significant improvement in either parameter. On the basis of these results, B. pseudocatenulatum was selected for subsequent in-depth mechanistic investigations in an animal model (Fig. 2E). The selected B. pseudocatenulatum strain, which was isolated from a healthy donor, has been deposited in the China General Microbiological Culture Collection Center under accession number CGMCC 10388. Genomic sequencing of this strain revealed a genetic repertoire enriched in pathways related to lipid metabolism, glycan biosynthesis and metabolism, energy metabolism, and carbohydrate metabolism (Fig. 2F).
B. pseudocatenulatum supplementation alleviated experimental ethanol-induced liver injury
The mice were fed with the Lieber–DeCarli alcohol diet for 8 weeks to induce the ALD model as described above. An experimental scheme illustrating the dietary protocols for all groups is presented in Fig. 3A. The mice’s status and body weight were assessed on a weekly basis. No accidental death was observed during the experiment. There was no difference in basal body weight among the groups. Owing to the pair-feeding protocol implemented for the NC and PC groups, which ensured that the two liquid diets were isocaloric, no significant difference in body weight gain was detected. B. pseudocatenulatum decreased body weight and percentage of weight gain (P < 0.05; P < 0.05) (Fig. 3B, Supplementary Fig. 1). The liver weight and liver-to-body weight ratio at week 8 were significantly greater in the PC group than in the NC group. Conversely, B. pseudocatenulatum supplementation effectively reversed these alcohol-induced increases (Fig. 3B, Supplementary Fig. 1). Although B. pseudocatenulatum supplementation significantly ameliorated this increase compared to the PC group (P < 0.05), the liver-to-body weight ratio in the probiotic-treated group remained significantly higher than that in the NC group (P < 0.05) (Fig. 3B). This result suggests a partial restoration of liver mass following probiotic intervention.

A Schematic of the chronic-binge ethanol feeding model (NIAAA model) and the experimental design for Bifidobacterium pseudocatenulatum intervention (same as in Fig. 1D). B Body weight and liver-to-body weight ratio. C Assessment of hepatic histopathology: Hematoxylin and Eosin (H&E), Oil Red O, and Myeloperoxidase (MPO) staining. D Hepatic levels of cholesterol and triglycerides, and serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST). E Serum concentrations of inflammatory cytokines and chemokines.
H&E staining revealed distinct histological features among the groups. In the NC group, hepatocytes were radially arranged around the central vein with clear and intact lobular architecture; only occasional small lipid droplets were observed. In contrast, the model group exhibited severe hepatic steatosis and injury, characterized by hepatocytes containing variably sized lipid droplets that displaced the nuclei to the periphery. These cells appeared enlarged, rounded, and disorganized and were accompanied by marked inflammatory cell infiltration. Consistent with these morphological changes, the Nanji score was significantly greater in the model group than in the NC group (P < 0.01). Treatment with B. pseudocatenulatum significantly alleviated these pathologies, reducing both intracellular lipid accumulation and inflammatory cell infiltration, as reflected by a markedly lower Nanji score in the B. pseudocatenulatum-treated group than in the model group (P < 0.05). The lipid status of the mouse liver was checked by Oil Red O staining. We found that the PC group exhibited significant fat accumulation, and microscopic observation revealed infiltration of a vast number of red lipid droplets (P < 0.01; Fig. 3C). However, this fatty infiltration was significantly reduced by treatment with B. pseudocatenulatum (P < 0.01). Less steatosis was observed in the NC group. Analysis of cholesterol and triglyceride levels in liver tissue revealed that the Lieber–DeCarli diet significantly increased lipid deposition in liver cells (P < 0.01, P < 0.05). B. pseudocatenulatum intervention resulted in significant reductions in tissue triglyceride and cholesterol levels (P < 0.01 and P < 0.05, respectively) (Fig. 3D). Neutrophil infiltration was revealed by myeloperoxidase (MPO) staining. MPO (+) cell quantity in the PC group was significantly greater than that in the NC and B. pseudocatenulatum groups (P < 0.05; P < 0.05) (Fig. 3C).
Fat accumulation in liver cells can cause steatosis and damage liver tissue. Assessment of liver function revealed that compared with the NC group, the PC group had significantly elevated serum ALT and AST levels (P < 0.01 for both). Importantly, B. pseudocatenulatum treatment significantly counteracted this effect, lowering ALT (P < 0.01) and AST (P < 0.05) levels (Fig. 3D). Inflammatory processes are integral to the pathogenesis and progression of alcohol-associated liver disease22. In this study, inflammatory cell infiltration was also observed in liver pathological sections. Inflammatory factors and chemokines were detected to assess the level of the inflammatory response in the peripheral circulation. The levels of pro-inflammatory cytokines, including IL-1α, IL-1β, IL-5, IL-6, IL-12 (p70), and IL-17, were significantly increased (P < 0.01, P < 0.01, P < 0.05, P < 0.01, P < 0.01, P < 0.01; respectively), while the serum levels of IL-1α, IL-1β, IL-5, and IL-6 were significantly decreased after intervention with B. pseudocatenulatum (P < 0.01, P < 0.01, P < 0.01, P < 0.01; respectively). The reduction in IL-10 following probiotic intervention may be a consequence of the attenuated inflammatory environment, thereby reducing the host’s compensatory need for elevated anti-inflammatory responses (P < 0.01) (Fig. 3E).
Similarly, the serum chemokine levels also showed a consistent trend. In the alcoholic liquid diet group, the expression levels of G-CSF, KC, MCP-1, MIP-1α, MIP-1β, and TNF-α increased significantly (P < 0.01, P < 0.01, P < 0.01, P < 0.01, P < 0.01, P < 0.01; respectively). Moreover, the RANTES level decreased (P < 0.01). Compared with the PC group, the B. pseudocatenulatum group had lower levels of the alcohol-induced chemokines G-CSF, KC, MCP-1, and MIP-1α (P < 0.05, P < 0.01, P < 0.01, P < 0.01) and higher levels of RANTES (P < 0.01). Compared with the NC group, the B. pseudocatenulatum group exhibited markedly lower levels of MIP-1α expression (P < 0.01) (Fig. 3E).
B. pseudocatenulatum restores the gut barrier in ALD
Intestinal barrier integrity was assessed using a combination of techniques: histopathological observation, electron microscopy, measurement of mucus layer thickness, and analysis of the expression of junction-related proteins, serum LPS-binding protein (LBP), and pattern recognition receptors23. H&E staining of the colon revealed that the Lieber–DeCarli alcohol diet significantly increased subepithelial Gruenhagen’s space as well as epithelial elevation and that some crypts almost disappeared. Consistent with these observations, the Chiu score significantly increased in the model group (P < 0.01). However, B. pseudocatenulatum administration reduced this abnormal histological structure such that the crypts and villi were neatly arranged and the structure was more complete. Accordingly, the Chiu score was significantly lower in the B. pseudocatenulatum-treated group than in the model group (P < 0.05). The structure of intestinal microvilli was assessed by transmission electron microscopy, and the intestinal microvillus structure in the NC group was complete, with dense and regular brush borders. The intestinal microvilli in the PC group were ruptured, sparse, short, and scattered. Consequently, B. pseudocatenulatum significantly improved the destruction of the brush border (Fig. 4A).

A Histological alterations and mucosal ultrastructure in the terminal ileum, as assessed by hematoxylin and eosin (H&E) staining and electron microscopy. B Assessment of goblet cells and mucus layer by Alcian blue/Periodic Acid-Schiff (AB-PAS) staining, and tight junction integrity by Zonula Occludens-1 (ZO-1) immunofluorescence; mRNA expression levels of tight junction proteins (ZO-1, Occludin). C mRNA expression levels of lipopolysaccharide-binding protein (LBP) and Toll-like receptor (TLR) pathway components (MYD88, TLR-2, TLR-4, TLR-5, TLR-7) as determined by quantitative PCR. D Protein localization and expression of TLR4 and MYD88 evaluated by immunohistochemistry. Data are presented as the mean ± SEM (N: Normal Control [NC] = 8, Positive Control [PC] = 10, B. pseudocatenulatum = 10). Statistical analysis was performed using one-way ANOVA followed by Bonferroni correction (for equal variances) or Dunnett’s T3 test (for unequal variances). *P < 0.05 and **P < 0.01 compared with the PC group.
Periodic acid-Schiff (PAS) and Alcian blue (AB) staining indicated that ethanol exposure impaired the intestinal mucosal chemical barrier, as evidenced by a significant reduction in the thickness of the intestinal mucus layer in the PC group compared with that in the normal control group (P < 0.01). This reduction, however, was notably reversed following supplementation with B. pseudocatenulatum (P < 0.05). Interestingly, we observed a significant increase in the goblet cell count in the mouse model of ALD (P < 0.05). In conjunction with the literature, these findings suggest that although the number of goblet cells is elevated, their function is likely compromised, leading to impaired mucus secretion and barrier integrity (Fig. 4B)24.
The expression of mouse intestinal Tjp was sequentially detected by qPCR and immunofluorescence. In terms of the mRNA expression level, the expression of intestinal occludin factor decreased in the PC group (P < 0.01). The ZO-1 factor also tended to decrease (P = 0.09). B. pseudocatenulatum supplementation notably increased the expression of occludin and ZO-1 (P < 0.01, P < 0.05). Immunofluorescence staining for Tjp revealed that the green compact protein ZO-1 was visible on the intestinal surface of the mice in the NC group. The fluorescence signal of the PC group weakened over a wide range, and the continuity of the protein structure was interrupted (P < 0.05). B. pseudocatenulatum significantly improved the fluorescence signal intensity (P < 0.05; Fig. 4B).
Compared with the NC group, the PC group had increased serum LBP content (P < 0.01), suggesting that the increased endotoxin content may be related to intestinal barrier function. However, the B. pseudocatenulatum group had a significantly decreased serum level of LBP (P < 0.01). Many studies have reported that LPS acts on the TLR-4-MyD88 signaling pathway to activate inflammatory responses25. Compared with that in the NC group, the intestinal expression of TLR4 and its linker MyD88 in the ALD group was greater (P < 0.05 and P < 0.05, respectively). In the intervention group, MyD88 expression significantly decreased (P < 0.01), and TLR4 expression significantly decreased (P < 0.1) (Fig. 4C). These findings were further confirmed at the protein level by immunohistochemistry (Fig. 4D). In addition to the expression of TLR4, the expression of other members of the TLR2 and TLR7 genes significantly increased in the ALD group (P < 0.05 and P < 0.05, respectively), whereas the expression of the TLR-5 factor increased with increasing B. pseudocatenulatum supplementation (P < 0.01) (Fig. 4C).
B. pseudocatenulatum improved microbiome imbalance and restored the gut microbiota
To explore the influence of ALD and probiotic strain pretreatment on the gut microbiota composition, the 16S rRNA V3-V4 region was sequenced using the Illumina platform. A total of 976,241 clean reads from 28 mouse samples were filtered for downstream analysis.
The alpha diversity of the gut bacterial community was evaluated using indices Chao1, the Shannon index, the Simpson index, and Observed species. The sequencing depth was evaluated with Good’s coverage. The alpha diversity indices did not differ significantly among the groups (Table 3).
β-Diversity was calculated on the basis of UPGMA clustering and PCoA. Compared with the mice fed with the Lieber–DeCarli control diet, the mice fed the Lieber–DeCarli alcoholic diet had significantly different colony compositions (ANOSIM Binary Jaccard P = 0.001, unweighted, P = 0.001) (Fig. 5A). Changes in the microbial community structure were revealed with PCoA. Under the unweighted algorithm, PC1 explained 13.3% of the overall difference, PC2 explained 9.6%, and PC3 explained 7.7% of the overall difference. The binary Jaccard algorithm explained 12%, 10.1% and 7.7% of the overall variance, respectively (Fig. 5A).

A Principal coordinate analysis (PCoA) plots based on unweighted and weighted UniFrac metrics, illustrating the beta diversity of the microbial communities. B Compositional profiles of the gut microbiota, showing the most abundant taxa at the phylum, family, and genus levels. C Cladogram generated from linear discriminant analysis (LDA) effect size (LEfSe) analysis, depicting the phylogenetically enriched taxa across different groups. Differences are indicated by the color of the most abundant class, with the circle diameter proportional to the taxon abundance and each ring representing a lower taxonomic level. D Hierarchical clustering analysis based on Euclidean distance, showing the intensity levels of characteristic metabolic pathways.
At the phylum level, compared with the NC group, the PC group exhibited a significant decrease in Firmicutes abundance (P < 0.01). Notably, B. pseudocatenulatum supplementation effectively restored Firmicutes levels (P < 0.05) while concurrently promoting significant increases in Actinobacteria and Verrucomicrobia populations (both P < 0.05). Importantly, the intervention demonstrated a suppressive effect on potential pathogenic taxa, as evidenced by reduced Proteobacteria abundance (Fig. 5B).
At the family level, the PC group demonstrated significant enrichment of potential pathogenic taxa, including Peptostreptococcaceae (P < 0.01), along with elevated levels of Enterobacteriaceae and Helicobacteraceae (both P < 0.05). Conversely, this group exhibited marked depletion of beneficial commensals, with reduced abundances of Bifidobacteriaceae (P < 0.05), Peptococcaceae (P < 0.01) and Lachnospiraceae (P < 0.05). Probiotic intervention effectively ameliorated these dysbiotic patterns, inducing marked increases in Bifidobacteriaceae and Peptococcaceae populations (both P < 0.01) but suppressing Peptostreptococcaceae abundance (P < 0.05). Furthermore, B. pseudocatenulatum supplementation selectively modulated specific bacterial families, such as Bacteroidaceae (P < 0.05), Ruminococcaceae, and Christensenellaceae (both P < 0.01), compared with those in the PC group. Intriguingly, the intervention concurrently reduced Rikenellaceae and Desulfovibrionaceae (represented by the Desulfovibrio genus) abundances (both P < 0.01) (Fig. 5B).
At the genus level (Fig. 5B), comparative analysis revealed distinct microbial shifts in the PC group, characterized by significantly reduced abundances of the beneficial genera Bifidobacterium (P < 0.05) and Blautia (P < 0.01) relative to those in the NC group. Notably, probiotic intervention markedly increased the abundance of these taxa (Bifidobacterium: P < 0.01; Blautia: P < 0.05). Pathogen-associated genera exhibited differential enrichment patterns, with Escherichia-Shigella (P < 0.01) and Parabacteroides (P < 0.05) showing significant proliferation in the PC group. Concurrently, we observed substantial depletion of SCFA-producing genera, including Alloprevotella, Anaerotruncus, Lachnoclostridium, Lachnospiraceae NK4A136 group, and Roseburia (all P < 0.01), along with Helicobacter (P < 0.05). B. pseudocatenulatum supplementation elicited profound microbial remodeling, manifested by enrichment of mucin-associated Bacteroides and fiber-degrading Christensenellaceae R-7 (P < 0.01, P < 0.05) (Fig. 5B).
The phylotype of the microbiota was assessed with LEfSe analysis, which identifies differential markers with biological associations26. LEfSe revealed that the abundance of potentially pathogenic bacterial taxa, including Enterobacteriaceae, Escherichia-Shigella, Peptostreptococcales_ Tissierllales, Rikenella, etc., was greater in the PC group than in the NC group, while the abundance of Roseburia and Ruminococcus was greater in the NC group than in the control group. The cecal contents of the mice showed increased abundances of Bifidobacteriales, Bifidobacteriaceae, and Bifidobacterium after 8 weeks of probiotic gavage. Compared with the B. pseudocatenulatum group, the PC group was enriched in more Deltaproteobacteria, Desulfovibrionales, Desulfovibrionaceae, Rikenella, etc. (Fig. 5C).
On the basis of the determined microbial genomes, the Kyoto Encyclopedia of Genes and Genomes (KEGG) database and PICRUSt software were used to analyze the composition of the genes, associated genes and functional information. We found that the functional differences in the flora were related mainly to digestion, absorption and metabolic processes related to carbohydrates (fructose and mannose, galactose, starch, sucrose and other glycans) and lipid metabolism (lipid metabolism, glycerolipid metabolism, linoleic acid metabolism, etc.) (Fig. 5D).
B. pseudocatenulatum alters the fecal metabolome composition in ALD mice
According to the functional prediction of the intestinal flora, the Lieber–DeCarli alcohol–supplemented liquid diet and probiotic intervention significantly affected the metabolic process of the gut microbiota. Therefore, untargeted metabolomics (GC‒MS detection) was carried out to analyze the characteristics of the fecal metabolites.
To evaluate the stability of the overall experiment, the analysis process and the distribution of the samples among the groups were used to analyze all the included samples and quality control (QC) samples, that is the mixed samples. In the PCA graph, t[1], t[2], and t[3] correspond to the initial three principal components. The degree of dispersion between samples reflects metabolite differences between groups. The QC samples were tightly aggregated and located in the center of the 3D map, indicating that the experimental and analytical processes were relatively stable. Among the samples between groups, the degree of discrimination between the NC group and the PC group was greater than that between the PC and B. pseudocatenulatum groups. However, there was still a significant clustering trend among the samples in each group, suggesting that there were large differences in metabolites among the groups (Fig. 6A).

A Principal component analysis (PCA) plot of fecal metabolomes from the normal control (NC), positive control (PC), and B. pseudocatenulatum supplement groups. B Volcano plots displaying differentially abundant metabolites between the PC and NC groups (left) and between the B. pseudocatenulatum and PC groups (right). C Spearman correlation analysis exploring the associations between differentially abundant gut microorganisms and metabolites. D Effects of B. pseudocatenulatum on the concentrations of key intestinal short-chain fatty acids (SCFAs). E Hepatic mRNA expression levels of key inflammatory cytokines, lipid metabolism-related factors, and Toll-like receptors. The data are presented as the mean ± SEM (N: Normal Control [NC] = 8, Positive Control [PC] = 10, B. pseudocatenulatum = 10). Statistical analysis was performed using one-way ANOVA followed by Bonferroni correction (for equal variances) or Dunnett’s T3 test (for unequal variances). *P < 0.05 and **P < 0.01 compared with the PC group.
A total of 271 metabolites were identified. The filter was based on a t test (Student’s t test) P-value < 0.05, VIP value > 1 and FC to obtain differentially abundant metabolites between groups. A total of 53, 37, and 48 metabolites were found to be differentially abundant between the PC and NC groups, B. pseudocatenulatum and PC groups, and B. pseudocatenulatum and NC groups, respectively (Supplementary Tables 1–3). These differentially abundant metabolites were enriched mainly in pathways such as those involving carbohydrates, alcohols and polyols, amino acids, and steroids and steroid derivatives (subclasses of bile acids). Metabolomic profiling via volcano plot analysis revealed distinct alterations in hepatic metabolites following 8 weeks of alcohol consumption intervention. The alcohol-exposed group exhibited the most pronounced alterations in three key metabolites: fructose, galacturonic acid and xanthosine. Notably, chronic alcohol intake significantly downregulated sterol metabolism, as evidenced by reduced levels of stigmasterol and galactitol. Probiotic supplementation partially reversed alcohol-induced metabolic disturbances: The increase in the level of Galacturonic Acid (P = 0.002) was normalized. 11-Eicosenoic acid (log2FC = −1.9; P = 0.008) and uridine (log2FC = −1.5; P = 0.03) significantly decreased lipid remodeling. The accumulation of cysteine-glycine and urea indicated increased ammonia detoxification capacity (Fig. 6B).
The gut ecosystem demonstrates dynamic microbiota–metabolite crosstalk. Gut microbial activity affects the composition of metabolites, and the metabolism of external substances also affects the microbial community. We combined metabolomic and 16S rRNA analyses by using Spearman correlation analysis to explore the interactions between microorganisms and their metabolites. Escherichia coli, which was enriched in the PC group, was negatively correlated with indoxyl sulfate (R = −0.88, P < 0.01) in the association analysis between the NC and PC groups. A significant positive correlation was observed between the Lachnospiraceae FCS020 group and the Ruminococcaceae NK4A214 group with respect to 4-hydroxybutyric acid levels (R = 0.83, P < 0.001; R = 0.84, P < 0.001). A statistically significant negative correlation was identified between the levels of Romboutsia and lutein (R = −0.86, P < 0.001) in this study. The enriched ambiguous taxa in the NC group were negatively correlated with 2-hydroxycaproic acid (R = −0.85; P < 0.001), Alloprevotella was positively correlated with luteinic acid and daidzein (R = 0.87; P < 0.001; R = 0.84; P < 0.001), and Ruminiclostridium exhibited a significant inverse relationship with galacturonic acid and isoleucine (R = −0.85, P < 0.001; R = −0.81, P < 0.001) (Fig. 6C).
A comparison of the B. pseudocatenulatum probiotic and PC groups revealed that in the probiotic group, the abundance of Ruminococcaceae UCG-013 was negatively correlated with the abundance of deoxycholic acid (R = −0.82, P < 0.01). Parvibacter was inversely correlated with nonadecanoic acid (R = −0.81, P < 0.001). While Ruminiclostridium was positively correlated with dodecanoic acid (R = 0.83, P < 0.001), Defluviitalea was positively correlated with 6-deoxyglucose and kynurenine (R = 0.81, P < 0.001; R = 0.83, P < 0.001).
On the basis of the microbiota and metabolome analyses, we found that ethanol exposure significantly affected lipid metabolic processes, especially the metabolism of fatty acids. B. pseudocatenulatum is considered a SCFA producer. A targeted metabolomic analysis of SCFAs revealed significantly reduced propionic and dimethylbutyric acid levels in the PC group. Additionally, the levels of acetic, propionic, dimethylbutyric, butyric, and isobutyric acids significantly increased in the gut in response to B. pseudocatenulatum (Fig. 6D).
B. pseudocatenulatum exhibited overall anti-inflammatory activity and improved lipid metabolism
Indicators of hepatic inflammation and lipid metabolism were detected, and compared with the NC group, the PC group exhibited higher expression of the inflammatory factor IL-1β and the chemokine CCL2 (P < 0.01, P < 0.05). The levels of TNF-α and the immunomodulatory cytokines IL-6 and IL-10 also significantly increased (P = 0.08; P = 0.06; P = 0.08), whereas the level of IL-4 significantly decreased (P < 0.01). The levels of TNF-α, IL-1β, IL-10 and the chemokine CCL2 in the liver of the B. pseudocatenulatum group were significantly lower than those in the ALD group (P < 0.01, P < 0.01, P < 0.05, and P < 0.01, respectively), and the level of IL-6 also tended to decrease (P = 0.09) (Fig. 6E).
TLRs activate inflammatory cytokines by recognizing pathogen-derived molecules such as bacteria, viruses and fungi. Compared with that in the NC group, the expression level of TLR4 was significantly greater (P < 0.01), whereas that of TLR-12 was notably lower (P < 0.01) in the livers of mice in the PC group. In the B. pseudocatenulatum group, liver TLR-2 (P < 0.01) and TLR-4 (P < 0.01) expression significantly decreased, and TLR-9 expression tended to decrease (P = 0.06), whereas TLR-12 expression significantly increased (P < 0.05) (Fig. 6E).
Significant changes in liver lipid synthesis-related factors were also detected in the PC group. ACC-1, CD36, FASN, etc., were markedly increased (P < 0.01, P < 0.01, and P < 0.05, respectively), and CPT1a levels were decreased (P < 0.05). There was no significant difference in the expression level of SREBP1c. The probiotic intervention triggered a decrease in the expression of the fatty acid synthase (FASN) factor, ACC-1 and CD36 (P < 0.05) and a significant increase (P < 0.01) in the CPT1a level. The peroxisome proliferator-activated receptor (PPAR) is a ligand-activated transcription factor with multiple domains. Studies have shown that PPARs are involved in regulating glucose and lipid metabolism and improving inflammatory responses. In our study, the alcohol diet significantly decreased the expression of PPAR-α and PPAR-γ (P < 0.01, P < 0.01). However, no significant difference was observed with respect to the effects of the probiotic intervention (Fig. 6E).
Safety assessment of B. pseudocatenulatum
A dedicated toxicological evaluation was conducted to assess the biosafety of the B. pseudocatenulatum strain. Throughout the 8-week oral administration period, no adverse effects on general health were observed. Clinical parameters, including fur condition, mental state, spontaneous activity, and behavior, remained normal, with no instances of diarrhea, constipation, anorexia, or mortality recorded. The levels of critical liver function biomarkers (ALT and AST) did not significantly differ. Concordantly, histopathological examination of liver tissues revealed no signs of toxicity or pathology. Although a trend toward body weight change was noted (P = 0.06), this finding—when considered alongside the absence of histological, biochemical, and clinical abnormalities and a high-energy liquid diet—collectively indicates a favorable safety profile for the administered strain (Supplementary Fig. 2).
Discussion
Recent research has indicated that ALD is linked to alterations in the composition and profile of the gut microbiota27. The microbiota composition influenced by alcohol consumption has a substantial effect on the development and progression of liver disease28. Targeting microbes may be an adjunctive strategy to prevent or treat alcoholic liver injury in the future29. B. pseudocatenulatum has been shown to have an effective regulatory effect on obesity-related metabolic and immune dysfunction in high-fat diet-fed mice and to prompt a transition toward an anti-inflammatory phenotype, morphology, and function in GM-CSF monocyte-derived and AF macrophages derived from patients with cirrhosis16. This transition may assist in managing persistent inflammation in individuals with decompensated cirrhosis. The strains used in this study have also been confirmed in previous studies, which revealed that B. pseudocatenulatum can reduce galactosamine-induced liver damage, systemic inflammatory responses, intestinal mucosal damage, bacterial translocation, and intestinal remodeling disorders, underscoring the role of the bacterial flora30. To investigate the therapeutic potential and mechanism of B. pseudocatenulatum in ALD, we established a mouse model using an 8-week Lieber–DeCarli ethanol diet. B. pseudocatenulatum intervention significantly alleviated ethanol-induced liver injury, as evidenced by reduced liver weight, decreased hepatic lipid deposition, and altered expression of inflammatory and lipid metabolic factors. Critically, the reduction in total body weight was interpreted as a marker of improved metabolic status. This conclusion is underpinned by the fact that the isocaloric, high-energy Lieber–DeCarli diet sustains body weight in the NC group but effectively promotes hepatic steatosis over time. Therefore, the concurrent attenuation of liver fat and normalization of liver function in probiotic-treated mice collectively confirm that weight loss signifies a beneficial shift toward metabolic homeostasis. B. pseudocatenulatum treatment significantly reversed the alcohol-induced hepatomegaly (PC vs. probiotic group); the liver mass in the supplemented group did not fully normalize and remained elevated compared to the NC group. This pattern indicates a partial restoration of liver mass, consistent with significant but incomplete resolution of alcohol-induced steatosis and injury and underscores that while B. pseudocatenulatum confers substantial therapeutic benefit, the restoration of total liver homeostasis to a naive state is a complex and potentially protracted process.
Chronic alcohol consumption is a well-established contributor to increased intestinal permeability, facilitating the translocation of gut-derived bacteria and their byproducts into the systemic circulation. This leads to the activation of hepatic Kupffer cells and the subsequent release of pro-inflammatory factors and chemokines, which ultimately contribute to liver injury31,32. Consistent with this established mechanism, our findings confirm that ethanol exposure reduced colonic mucus layer thickness and downregulated the expression of tight junction proteins (ZO-1 and occludin), thereby increasing gut permeability. Importantly, intervention with B. pseudocatenulatum effectively restored intestinal barrier integrity and significantly reduced serum endotoxin levels.
The LPS component in the cell wall outer membrane of gram-negative bacteria is thought to be significantly associated with pro-inflammatory and fibrotic signals in the liver33. LPS activates Kupffer cells by binding to TLR4 to promote liver inflammation. Following Kupffer cell activation, the primary inflammatory cytokines released are TNF-α, IL-1β and IL-6. The chemokine CCL2 is additionally involved in modulating the secretion of the aforementioned inflammatory cytokines34. Indeed, the function of IL-6 in ALD is intricate and unclear. Some researchers suggest that IL-6 may play a protective role in the liver. After alcohol-induced liver injury, IL-6 may inhibit hepatocyte apoptosis and contribute to the repair of mitochondrial DNA35. Other researchers believe that IL-6 potentially facilitates the differentiation of human Th17 cells and the subsequent production of IL-17, thereby playing a role in the development of ethanol-induced hepatitis22.
TLR acts as a ‘gatekeeper’ in ALD36. The present study demonstrates that B. pseudocatenulatum differentially modulates the expression of TLR family members across the gut-liver axis. In the intestine, alcohol exposure elicited upregulation of TLR2, TLR4, and TLR7, known to recognize bacterial lipoproteins, LPS, and viral single-stranded RNA, respectively. This pattern reflects widespread microbial ligand translocation and consequent innate immune activation. Following probiotic intervention, TLR4 expression was selectively suppressed, indicating a primary role in inhibiting the LPS-TLR4 axis that is central to alcohol-related inflammatory signaling. In contrast, expression of TLR5, a receptor for bacterial flagellin linked to microbial homeostasis and epithelial barrier integrity, was significantly enhanced37,38. In the liver, probiotic administration downregulated both TLR2 and TLR4, likely due to reduced exposure to intestinal-derived ligands and attenuated inflammatory feedback. Concurrently, it markedly elevated the expression of TLR12, a receptor implicated in tissue-specific immunoregulation39. These findings indicate that the probiotic does not induce broad TLR suppression but rather precisely recalibrates receptor activity, as inhibiting dominant pro-inflammatory pathways while promoting those associated with homeostasis, thereby orchestrating a coordinated reshaping of the host immune landscape.
More than 90% of individuals who consume alcohol over an extended period are likely to develop fatty liver disease1. In our study, a large accumulation of lipid droplets was observed in the liver in the PC group, indicating the presence of lipid metabolism disorders. Alcohol has a detrimental effect on the transcription factor SREBP1c and regulates the high expression of key lipogenic enzymes, such as ACC and FASN, which promote the incorporation of free fatty acids (FFAs) into lipid droplets40. Moreover, liver mitochondrial β-oxidation is reduced by the use of FFA substrates (CPT1a, PPAR-α, etc.). Additionally, alcohol exposure leads to increased uptake of circulating FFAs directly into hepatocytes41, and this increased trafficking of FFAs is correlated with upregulation of the expression of the cell surface FFA transporter CD3642. The metabolic intermediate acetaldehyde inhibits PPAR-α and PPAR-γ transcription, which increases the rate of fat accumulation compared with that of lipolysis in the liver. Our study revealed that B. pseudocatenulatum significantly reduced the accumulation of lipid droplets in the liver, but the expression of PPAR-γ was not significantly downregulated in the B. pseudocatenulatum group. However, factors associated with the incorporation of FFAs into lipid droplets, such as SREBP1c, ACC and FASN, varied significantly. These findings suggest that the main mechanism of B. pseudocatenulatum may involve not controlling fatty acid intake and promoting decomposition but rather integrating free fatty acids into lipid droplets.
Differential changes in the gut microbiota and metabolites induced by B. pseudocatenulatum intervention are also closely related to disease progression. At the gut bacterial taxonomic level, the abundance of Bifidobacterium and Blautia was significantly lower in the PC group than in the NC group. However, following probiotic intervention, the abundance of both genera significantly decreased. Bifidobacterium, a prominent member of the gut microbiota, plays diverse probiotic roles in healthy individuals, such as vitamin synthesis and carbohydrate fermentation, the restoration of the gut microbiota balance and the enhancement of host immune responses43. The Blautia genus is involved in the production of SCFAs, particularly acetic acid44,45. Mouse ingestion of alcohol results in the production of acetic acid under the action of ADH and ALDH. Therefore, it is speculated that substrate ethanol uptake leads to a higher acetic acid concentration, resulting in a reduced abundance of Blautia. Rodriguez et al.46 reported that the abundance of Blautia was positively correlated with a reduction in obesity and hepatic steatosis. These findings also corroborate our results. Probiotic modulation increases the abundance of the Blautia genus while improving hepatic steatosis. The abundance of the potentially pathogenic bacteria Escherichia-Shigella and Parabacteroides, which are closely related to inflammatory factors, was greater in the disease group than in the control group47. In our study, the results of the metabolomic analysis revealed that E. coli in cecal contents was negatively correlated with indoxyl sulfate. The genus E. coli exhibits pro-inflammatory properties, as evidenced by a significant increase in abundance in response to an alcoholic diet48. Indole sulfate is produced from the digestion by intestinal microorganisms of the essential amino acid substrate tryptophan and is then absorbed into the liver49. Alcohol intake often inhibits intestinal absorption of nutrients, such as amino acids. Therefore, changes in bacterial flora structure and insufficient nutrient intake may lead to a reduction in indole sulfate content.
An association analysis between the B. pseudocatenulatum and PC groups revealed that the Ruminococcaceae enriched in the probiotic group was negatively correlated with deoxycholic acid. Bile acids also significantly contribute to the remodeling of the microbiota, and the microbiota affects bile acid metabolism50. Deoxycholic acid is cytotoxic and can simultaneously disrupt glucose regulation by disrupting hepatic endoplasmic reticulum homeostasis in mice51, whereas Ruminococcaceae has been documented as a significant producer of short-chain fatty acids. thereby attenuating inflammatory responses52. After 8 weeks of being fed an alcoholic liquid diet, the cecal content exhibited a significant increase in galacturonic acid, as visualized through differentially abundant metabolite analysis using a volcano plot. The administration of probiotics led to a reduction in the levels of galacturonic acid. The primary origin of galacturonic acid is dietary pectin, which is predominantly metabolized by the microbial community Bacteroides. Galacturonic acid serves as a carbon substrate, facilitating the proliferation of pathogens, notably through the induction of colonic inflammation. This compound functions as both a nutritional resource and a crucial signaling molecule, delineating the interplay between the microbiota and pathogenic organisms within the gastrointestinal tract. Increased levels of cysteine-glycine, which is present in the B. pseudocatenulatum group, also function to regulate metabolism53.
The intestinal epithelium is shielded by a protective layer of mucus, which serves as a surface for microbiota colonization and provides a source of nutrients for their growth and metabolic functions, while the inner part of the mucus layer is virtually free of microbial colonization54,55. As a result, the majority of interactions between the host and its microbiota are indirect and occur through soluble factors such as bacterial products and metabolites, which are commonly referred to as prebiotics and postbiotics56. Current research indicates that the underlying mechanisms of long-term gut homeostasis are based on microbiota metabolites. Therefore, in addition to clarifying the protective effect of probiotic bacteria, we also used 16S rRNA detection combined with metabolome detection to identify bacterial metabolites that may have protective effects.
Our findings demonstrate that elevated acetate levels in the alcohol-induced liver injury model, particularly those enhanced by B. pseudolongum supplementation, are correlated with significant hepatoprotective effects. This finding aligns with prior reports that B. pseudolongum-derived acetate activates hepatic GPR43 to suppress pro-inflammatory IL-6/JAK1/STAT3 signaling57. Notably, while the referenced mechanism was established in NAFLD-HCC, our data extend this paradigm to ALD, suggesting a conserved protective role of acetate across etiologically distinct liver pathologies. Ki Tae Suk reported that blood acetate concentrations are elevated during ethanol consumption in ALD models58. Compared with that in the PC group, the concentration of acetate in the intestine further increased in the B. pseudocatenulatum group. This phenomenon may be attributed to several underlying mechanisms: (1) Metabolic compensation effect: The acetate generated from alcohol metabolism may not be efficiently converted into acetyl-CoA for energy metabolism because of mitochondrial dysfunction or insufficient activity of key enzymes. Supplementation with “exogenous acetate” may increase acetyl-CoA levels, thereby promoting fatty acid oxidation via the PPAR-related pathway and reducing lipid accumulation through alternative pathways. (2) Gut‒Liver Axis Modulation: Acetate strengthens intestinal barrier integrity via GPR43-dependent mucin secretion, reducing LPS translocation and TLR4/NF-κB-driven inflammation. (3) The potential indirect role of exogenously administered acetic acid is mediated through its interaction with other metabolites, such as butyric acid and propionic acid.
This study acknowledges limitations that also direct future research. First, the exclusive inclusion of male patients, while deliberate to control for sex as a confounding variable and to align with our animal model, precludes the generalizability of our findings to females. Future studies are warranted to explore potential sex-specific mechanisms in ALD. Inherent limitations of retrospectively collected data regarding detailed habits and over-the-counter medication use are difficult to quantify precisely. Future prospective studies with stringent dietary control are needed to validate our observations. Second, research on bacterial metabolites and their precise mechanisms of action remains in its infancy. A pertinent question regarding our intervention is the potential influence of B. pseudocatenulatum on alcohol absorption. Although blood alcohol concentrations were not measured, our data do not support the premise that the observed benefits stem from reduced ethanol uptake. The paired oral gavage protocol ensured equivalent alcohol exposure. Diminished absorption would likely exacerbate local mucosal injury because of prolonged luminal ethanol contact, yet we observed the opposite: a significant mitigation of gut leakiness and improved dysbiosis. These findings align with prior probiotic studies in ALD, which attribute efficacy to mechanisms such as enhanced barrier function without documented alterations in blood alcohol levels59. We therefore conclude that the protection conferred by B. pseudocatenulatum is independent of changes in alcohol absorption. In conclusion, our work systematically delineates the efficacy and mechanism of B. pseudocatenulatum against ALD. Our investigation originated from a clinical cohort showing Bifidobacterium depletion, led to the functional identification of B. pseudocatenulatum, and progressed to in vivo confirmation of its therapeutic effects in a murine model. Mechanistically, we elucidated that this strain protects against ALD by restoring intestinal barrier integrity, correcting microbial dysbiosis, and regulating metabolic and inflammatory pathways. These results establish B. pseudocatenulatum as a promising probiotic candidate for ALD management. While our results are promising, rigorous clinical trials are essential to validate the therapeutic potential of B. pseudocatenulatum and support its future translational application.
Methods
Study participants
Healthy controls were recruited from the Health Examination Center at the First Affiliated Hospital of Zhejiang University School of Medicine. Sample collection for both the ALD and HBV groups was conducted in the Department of Infectious Diseases–Hepatology at the First Affiliated Hospital of Zhejiang University School of Medicine. Fresh fecal and serum samples were obtained from all participants after written informed consent was provided. The study protocol was approved by the Ethics Committee of the First Affiliated Hospital of Zhejiang University (Approval No. 2019-981).
The inclusion criterion for all participants included individuals aged 18–60 years with a BMI of 18.5–28.0 kg/m². In light of epidemiological data showing that nearly two-thirds of ALD patients are male—and considering the established influence of sex on disease pathogenesis and the gut microbiota—this study enrolled an all-male cohort to control for these potential confounders60,61. Key exclusion criteria applied across all groups included a history of significant systemic diseases (e.g., metabolic or gastrointestinal disorders, diabetes, hypertension), recent antibiotic or probiotic use (within 2 months), excessive alcohol consumption, active infections, nonhepatic comorbidities affecting the gut microbiota, and recent immunosuppressive therapies. Additionally, participants in the healthy control and HBV groups were required to abstain from alcohol for at least 30 days prior to sample collection. Healthy controls had to meet specific laboratory criteria: ALT ≤ 50 U/L, AST ≤ 40 U/L, total bilirubin ≤ 17.1 μmol/L, serum creatinine within the normal range, normal vital signs, ECG and hematological parameters, and negative serology for HBsAg, HCV-Ab, and HIV.
For the ALD group, chronic alcohol consumption (≥40 g/day for men for more than 5 years), clinical or biochemical evidence of liver injury (elevated ALT, AST, GGT, TBil, or PT), imaging confirmation of the absence of cirrhosis, and the exclusion of viral or drug-induced liver diseases were included. For the hepatitis B virus (HBV) group, inclusion was based on confirmed HBV infection (HBsAg-positive or anti-HBc-positive with a chronic HBV history) along with histopathological or imaging evidence of no cirrhosis.
Strains and culture conditions
The B. pseudocatenulatum strain was sourced from healthy individuals and subsequently stored at the China General Microbiological Culture Collection Center (CGMCC15058). B. pseudocatenulatum was cultured under anaerobic conditions in Trypticase-Phytone-Yeast Broth Medium (manufactured by RiShui Co., Ltd., Qingdao, China) for 36 h at 37 °C and stored at −80 °C for a substantial period. Following resuspension in PBS, the bacterial concentration was standardized to 1.5 × 1010 CFU/ml by measuring the OD630. For oral administration, a 200 μl aliquot containing 3 × 109 CFU was prepared and administered to each mouse.
Experimental design
Six-week-old male specific pathogen-free C57BL/6J mice were fed a liquid control diet for 1 week for acclimation and housed in a specific pathogen-free facility with corncob bedding under a 12:12-h light–dark cycle. The room temperature was maintained at 22–24 °C. The experimental procedure comprised two sequential phases: an initial screening followed by an in-depth investigation. In the screening phase, the mice were randomly allocated into five groups (N = 8 per group): a negative control (NC) group, a positive control (PC) group, and three probiotic intervention groups that received Bifidobacterium longum, Bifidobacterium pseudocatenulatum, or Bifidobacterium adolescentis. B. pseudocatenulatum was selected for further evaluation. In the subsequent investigative phase, a new cohort of mice was randomly assigned to three groups: an NC group (N = 8) fed a pair-fed diet and administered normal saline, a PC group (N = 10) fed a Lieber–DeCarli ethanol diet and administered normal saline, and a B. pseudocatenulatum group (N = 10) fed the Lieber–DeCarli ethanol diet and treated daily with 3 × 10⁹ CFU of B. pseudocatenulatum via oral gavage.
The mice in the PC and probiotic groups were fed an alcohol-adapted diet in accordance with the principle of gradually increasing alcohol concentration. The mice were gradually fed the Lieber–DeCarli ethanol diet for a 1-week period: 1.5% (vol/vol) ethanol on days 1–2, 2.5% on days 3–4, 3.5% on day 5, and a final concentration of 5% (vol/vol) from day 7 onward for 8 weeks. In the B. pseudocatenulatum and PC groups, a mouse model was established of (8 weeks)–plus-binge ethanol feeding, and the mice were allowed free access to a 5% (vol/vol) ethanol TP-4030B diet. The NC group was fed an isocaloric liquid diet. A single dose of ethanol (5 g/kg body weight) was administered by gavage to the B. pseudocatenulatum and PC groups, and isocaloric maltose dextrin was administered to the NC group, which was sacrificed 9 h later. Anesthesia was induced by 5% isoflurane inhalation in a sealed chamber with 2 L/min oxygen flow. After loss of the righting reflex, the mice were transferred to a nose cone with 1.5% isoflurane maintenance. Blood collection was performed via the retro-orbital plexus using heparinized capillaries, with strict adherence to single-eye sampling. Immediate cervical dislocation ensured euthanasia prior to liver harvesting. This protocol complied with the AVMA Guidelines for the Euthanasia of Animals (2020 edition), minimizing suffering while preserving tissue integrity for downstream analyses. Animals were excluded from further experimentation and analysis if they exhibited severe adverse effects or died from causes unrelated to the experimental intervention.
Blinding
Personnel involved in different stages of the experiment were aware of the group assignment as follows: The principal investigator (Lanjuan Li), who was aware of group assignment, generated the randomization sequence and assigned mice to their respective groups. Personnel responsible for daily animal care and drug/vehicle administration were necessarily aware of group assignment because of the distinct procedures involved in preparing and administering the drug versus the vehicle control. To minimize potential bias arising from this knowledge, all treatments were administered using identical syringes at the same time daily following standardized protocols. Critically, all personnel responsible for assessing the primary outcomes (specific outcomes, histological scoring, etc.) were blinded to group allocation. Coded identifiers were used for all samples and data during assessment to enforce this blinding. The statistician performing the initial statistical analysis was blinded to group identities, which were labeled only as A, B, and C until after completion of the analysis and finalization of the results.
Ethics statement
All experiments involving live vertebrates were performed in strict accordance with the relevant national guidelines and institutional regulations. Specifically, the study adhered to the “Guidelines for the Care and Use of Laboratory Animals” issued by the Ministry of Science and Technology of China. The experimental protocols were reviewed and approved by the Institutional Animal Care and Use Committee of the First Affiliated Hospital, School of Medicine, Zhejiang University (Approval License Number: 2019-1075). The original ethics approval documentation is provided in the supplementary materials.
Biochemical indicators of hepatic function
ALT, AST, GGT, TBA, TBil, albumin and globulin levels in human serum were measured with a Hitachi 7600–210 automatic analyzer (Hitachi 7600–210; Hitachi, Tokyo, Japan). ALT and AST levels in mouse blood were measured with a dry chemistry analyzer. TG and TC levels in mouse hepatocytes were detected with a triglyceride or cholesterol assay kit (Applygen Technologies Inc., China).
Histopathological examination
Liver samples were acquired, fixed in 10% formalin for 1 d and subsequently embedded in paraffin for further analysis. The hepatic structure was assessed using hematoxylin and eosin (H&E) staining, and lipid accumulation was assessed with Oil Red O staining. Furthermore, to analyze neutrophil recruitment in the liver, immunohistochemical staining for myeloperoxidase (MPO) was performed using an antibody obtained from Abcam (Cambridge, UK). The histological evaluation was performed with Pannoramic Viewer software (Version 2.2; 3DHISTECH, Budapest, Magyarország, Hungary).
Plasma cytokine measurement
The concentrations of cytokines in the serum were measured using a Bio-Plex Cytokine Panel Assay kit (Bio-Rad, Hercules, CA, USA). Plasma (20 μl) was tested with a Bio-Plex 200 analyzer. Furthermore, the results were analyzed with Bio-Plex Manager 6.0 software (Rotstein 2014). The standard method and protocols that were used were established and described in the related literature62,63.
Examination of gut barrier function
The terminal ileum samples were fixed as indicated and stained with H&E. Mucosal thickness and goblet cells were assessed with Carnoy’s fluid fixative and stained with Alcian blue periodic acid-Schiff (AB-PAS). The length, diameter and density of the microvilli were measured to identify damage to the mucosal ultrastructure. The specimens were fixed in 2.5% glutaraldehyde, stained with uranyl acetate (3%) and lead citrate (0.2%), and subsequently viewed under a Philips Tecnai 10 electron microscope (Philips, Eindhoven, the Netherlands). Furthermore, immunohistochemical staining was performed to detect the expression of tight junction proteins (zonula occludens-1, ZO-1) and TLR4-MYD88 pathway components. All the slides were scanned and examined using Pannoramic Viewer software. Positive staining areas and histochemistry scores (H-scores) were analyzed. The H score was calculated as ∑(pi × i) = (percentage of weak intensity cells × 1) + (percentage of moderate intensity cells × 2) + (percentage of strong intensity cells × 3), where i represents the staining intensity grade: negative (no staining, 0 points), weakly positive (light yellow, 1 point), moderately positive (brownish yellow, 2 points), and strongly positive (brown, 3 points). pi denotes the percentage of positive cells at each respective intensity grade. Lipopolysaccharide (LPS) is an indicator of intestinal permeability impairment. As direct examination yielded inaccurate results, the concentration of LBP in the serum was quantified instead.
Nucleic acid extraction and 16S rRNA sequencing
Total genomic DNA was extracted from the stool samples using a QIAamp® Fast DNA Stool Mini Kit (Qiagen, Valencia, USA) according to the manufacturer’s instructions. To extract total RNA from liver samples and intestinal segments, an RNeasy Mini Kit (Qiagen, Valencia, USA) was used according to the manufacturer’s instructions. HiFi Hot Start Ready Mix (kk2601; Kapabiosystems, Beijing, China) was applied to amplify a 1 ng/μl DNA template. The V3-V4 regions of the 16S rRNA gene were amplified with the barcoded primers 343 F (5′-TACGGRAGGCAGCAG-3′) and 798 R (5′-AGGGTATCTAATCCT-3′). The libraries were subsequently sequenced on an Illumina MiSeq platform (Illumina, San Diego, CA). The resulting paired-end reads were merged using Fast Length Adjustment of Short Reads software. On the basis of the Silva database, chimeras were checked, and the raw tags were filtered with QIIME2 software. The gut microbiota composition was illustrated with alpha and beta diversity—including calculations of the Shannon index, Chao1, Simpson index, observed species, and Good’s coverage indices—as well as with PCoA and LEfSe analysis. The gene functions of the intestinal flora were predicted on the basis of the KEGG and GO databases.
Reverse transcription polymerase chain reaction
The comparative cycle threshold (Ct) method was used to determine the relative expression, with an Applied Biosystems VIIA7 real-time PCR system and a One-Step SYBR PrimeScript Plus RT‒PCR Kit (Takara Biomedicals, Kusatsu, Japan). The primer pairs used are listed in Supplementary Table 4.
Quantitative analysis of SCFAs in fecal samples
Preparation of the internal standard: The internal standard stock solution of hexanoic acid D3 was diluted to a ratio of 1:100, and the final concentration was 10 µg/ml.
Sample preparation: Samples (20 mg) were weighed and mixed with 400 µl of hexanoic acid standards and then homogenized at a speed of 5000 rpm for four cycles. The liquid portion was transferred into a microcentrifuge tube. Sulfuric acid (5%) was added at a ratio of 1:10 (v/v), and the mixture was subsequently vortexed. Afterward, 500 μl of ethyl acetate was injected into the tube. The derivatives were extracted by centrifugation at 14,000 rpm for 5 min. The derivatives were derivatized prior to GC–MS analysis.
Standard curve: The standard was diluted with ethyl acetate, and a 1 mg/ml stock solution (1:1000, v/v) was prepared. The acetic acid, propionic acid, butyric acid, valeric acid, isobutyric acid, 2-methylbutyric acid, etc., were diluted (1:9, v/v). The standard curves were constructed according to the gradient of the blank wells, 1000, 100, 10, 1, 0.1, and 0.01 µg/ml.
Parameters of GC‒MS and Data Analysis: High-purity helium was used as the carrier gas, with a constant flow rate of 1 ml/min and a −70 eV electron energy. The temperature of the ion source was 230 °C, and the quadrupole was 290 °C. The GC‒MS data were analyzed using Agilent MassHunter Workstation Qualitative Analysis software to determine the concentration of SCFAs on the basis of the standard curve.
Statistical analysis
The normality of the data distribution was assessed using the Kolmogorov‒Smirnov test. An unpaired t test or Mann‒Whitney U test was applied for two-group comparisons, whereas ANOVA or the Kruskal‒Wallis test was employed when three groups were involved according to the data distribution. Microbial community structure differences were evaluated through permutational multivariate analysis of variance and multiple response permutation procedures. Spearman rank correlation analysis was employed to assess associations between key indices. All the statistical analyses were performed using SPSS 20.0 (SPSS, Inc., Chicago, IL, USA) and GraphPad Prism 9.0 (GraphPad Software, Inc., CA, USA), with P < 0.05 considered to indicate statistical significance.
Data availability
The raw sequence data reported in this paper have been deposited in the Genome Sequence Archive in the National Genomics Data Center, China National Center for Bioinformation/Beijing Institute of Genomics, Chinese Academy of Sciences (GSA: CRA009220), and are publicly accessible at https://ngdc.cncb.ac.cn/gsa.
References
Seitz, H. K. et al. Publisher Correction: Alcoholic liver disease. Nat. Rev. Dis. Prim. 4, 18 (2018).
Patel, R. & Mueller, M. Alcoholic Liver Disease (StatPearls, 2022).
Wang, H., Mehal, W., Nagy, L. E. & Rotman, Y. Immunological mechanisms and therapeutic targets of fatty liver diseases. Cell Mol. Immunol. 18, 73–91 (2021).
Camilleri, M. Leaky gut: mechanisms, measurement and clinical implications in humans. Gut 68, 1516–1526 (2019).
Kapitan, M., Niemiec, M. J., Steimle, A., Frick, J. S. & Jacobsen, I. D. Fungi as part of the microbiota and interactions with intestinal bacteria. Curr. Top. Microbiol. Immunol. 422, 265–301 (2019).
Pohl, K., Moodley, P. & Dhanda, A. D. Alcohol’s impact on the gut and liver. Nutrients 13, 3170 (2021).
Wu, J. et al. Altered faecal microbiota on the expression of Th cells responses in the exacerbation of patients with hepatitis E infection. J. Viral Hepat. 27, 1243–1252 (2020).
Litwinowicz, K., Choroszy, M. & Waszczuk, E. Changes in the composition of the human intestinal microbiome in alcohol use disorder: a systematic review. Am. J. Drug Alcohol Abus. 46, 4–12 (2020).
Engen, P. A., Green, S. J., Voigt, R. M., Forsyth, C. B. & Keshavarzian, A. The gastrointestinal microbiome: alcohol effects on the composition of intestinal microbiota. Alcohol Res. 37, 223–236 (2015).
Bajaj, J. S. et al. Continued alcohol misuse in human cirrhosis is associated with an impaired gut-liver axis. Alcohol Clin. Exp. Res. 41, 1857–1865 (2017).
Llopis, M. et al. Intestinal microbiota contributes to individual susceptibility to alcoholic liver disease. Gut 65, 830–839 (2016).
Ciocan, D. et al. Characterization of intestinal microbiota in alcoholic patients with and without alcoholic hepatitis or chronic alcoholic pancreatitis. Sci. Rep. 8, 4822 (2018).
Bajaj, J. S. et al. Altered profile of human gut microbiome is associated with cirrhosis and its complications. J. Hepatol. 60, 940–947 (2014).
Wolfe, W. et al. The challenge of applications of probiotics in gastrointestinal diseases. Adv. Gut Microbiome Res. 2023, 1–10 (2023).
Nie, N. et al. Bifidobacterium plays a protective role in TNF-α-induced inflammatory response in Caco-2 cell through NF-κB and p38MAPK pathways. Mol. Cell. Biochem. 464, 83–91 (2020).
Cano, P. G., Santacruz, A., Trejo, F. M. & Sanz, Y. Bifidobacterium CECT 7765 improves metabolic and immunological alterations associated with obesity in high-fat diet-fed mice. Obesity 21, 2310–2321 (2013).
Chen, Y. et al. Bifidobacterium pseudocatenulatum ameliorates DSS-induced colitis by maintaining intestinal mechanical barrier, blocking proinflammatory cytokines, inhibiting TLR4/NF-κB signaling, and altering gut microbiota. J. Agric. Food Chem. 69, 1496–1512 (2021).
Mizokami, A., Kawakubo-Yasukochi, T. & Hirata, M. Osteocalcin and its endocrine functions. Biochem. Pharm. 132, 1–8 (2017).
Silva de Oliveira, R. et al. Bifidobacteria-derived exopolysaccharide promotes anti-tumor immunity. Cell Rep. 44, 116223 (2025).
Liu, L. et al. TWEAK-Fn14 signaling protects mice from pulmonary fibrosis by inhibiting fibroblast activation and recruiting pro-regenerative macrophages. Cell Rep. 44, 115220 (2025).
Liu, J. et al. TWEAK/Fn14 signals mediate burn wound repair. J. Invest. Dermatol. 139, 224–234 (2019).
Kawaratani, H. et al. The effect of inflammatory cytokines in alcoholic liver disease. Mediators Inflamm. 2013, 495156 (2013).
Muta, T. & Takeshige, K. Essential roles of CD14 and lipopolysaccharide-binding protein for activation of toll-like receptor (TLR)2 as well as TLR4 Reconstitution of TLR2- and TLR4-activation by distinguishable ligands in LPS preparations. Eur. J. Biochem. 268, 4580–4589 (2001).
Llorente, C. et al. mAChR4 suppresses liver disease via GAP-induced antimicrobial immunity. Nature 646, 180–189 (2025).
Jin, L. et al. Degradation products of polydopamine restrained inflammatory response of LPS-stimulated macrophages through mediation TLR-4-MYD88 dependent signaling pathways by antioxidant. Inflammation 42, 658–671 (2019).
Segata, N. et al. Metagenomic biomarker discovery and explanation. Genome Biol. 12, R60 (2011).
Sarin, S. K., Pande, A. & Schnabl, B. Microbiome as a therapeutic target in alcohol-related liver disease. J. Hepatol. 70, 260–272 (2019).
Canesso, M. C. C. et al. Comparing the effects of acute alcohol consumption in germ-free and conventional mice: the role of the gut microbiota. BMC Microbiol. 14, 240 (2014).
Gao, B. & Bataller, R. Alcoholic liver disease: pathogenesis and new therapeutic targets. Gastroenterology 141, 1572–1585 (2011).
Fang, D. et al. Bifidobacterium pseudocatenulatum LI09 and Bifidobacterium catenulatum LI10 attenuate D-galactosamine-induced liver injury by modifying the gut microbiota. Sci. Rep. 7, 8770 (2017).
Keshavarzian, A. et al. Leaky gut in alcoholic cirrhosis: a possible mechanism for alcohol-induced liver damage. Am. J. Gastroenterol. 94, 200–207 (1999).
Choudhry, M. A., Fazal, N., Goto, M., Gamelli, R. L. & Sayeed, M. M. Gut-associated lymphoid T cell suppression enhances bacterial translocation in alcohol and burn injury. Am. J. Physiol. Gastrointest. Liver Physiol. 282, G937–G947 (2002).
Seki, E. et al. TLR4 enhances TGF-beta signaling and hepatic fibrosis. Nat. Med. 13, 1324–1332 (2007).
Jiang, Y., Beller, D. I., Frendl, G. & Graves, D. T. Monocyte chemoattractant protein-1 regulates adhesion molecule expression and cytokine production in human monocytes. J. Immunol. 148, 2423–2428 (1992).
Sheron, N., Bird, G., Goka, J., Alexander, G. & Williams, R. Elevated plasma interleukin-6 and increased severity and mortality in alcoholic hepatitis. Clin. Exp. Immunol. 84, 449–453 (1991).
Gustot, T. et al. Differential liver sensitization to toll-like receptor pathways in mice with alcoholic fatty liver. Hepatology 43, 989–1000 (2006).
Vijay-Kumar, M. et al. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science 328, 228–231 (2010).
Chassaing, B., Ley, R. E. & Gewirtz, A. T. Intestinal epithelial cell toll-like receptor 5 regulates the intestinal microbiota to prevent low-grade inflammation and metabolic syndrome in mice. Gastroenterology 147, 1363–1377.e1317 (2014).
Koblansky, A. A. et al. Recognition of profilin by Toll-like receptor 12 is critical for host resistance to Toxoplasma gondii. Immunity 38, 119–130 (2013).
Ji, C., Chan, C. & Kaplowitz, N. Predominant role of sterol response element binding proteins (SREBP) lipogenic pathways in hepatic steatosis in the murine intragastric ethanol feeding model. J. Hepatol. 45, 717–724 (2006).
Zhong, W. et al. Chronic alcohol exposure stimulates adipose tissue lipolysis in mice: role of reverse triglyceride transport in the pathogenesis of alcoholic steatosis. Am. J. Pathol. 180, 998–1007 (2012).
Li, L. C. et al. Palmitate aggravates proteinuria-induced cell death and inflammation via CD36-inflammasome axis in the proximal tubular cells of obese mice. Am. J. Physiol. Ren. Physiol. 315, F1720–F1731 (2018).
Arboleya, S., Watkins, C., Stanton, C. & Ross, R. P. Gut bifidobacteria populations in human health and aging. Front. Microbiol. 7, 1204 (2016).
Shi, Z. et al. Anti-obesity effects of α-amylase inhibitor enriched-extract from white common beans (Phaseolus vulgaris L.) associated with the modulation of gut microbiota composition in high-fat diet-induced obese rats. Food Funct. 11, 1624–1634 (2020).
Koh, A., De Vadder, F., Kovatcheva-Datchary, P. & Backhed, F. From dietary fiber to host physiology: short-chain fatty acids as key bacterial metabolites. Cell 165, 1332–1345 (2016).
Rodriguez, J. et al. Discovery of the gut microbial signature driving the efficacy of prebiotic intervention in obese patients. Gut 69, 1975–1987 (2020).
Agudelo-Ochoa, G. M. et al. Gut microbiota profiles in critically ill patients, potential biomarkers and risk variables for sepsis. Gut Microbes 12, 1707610 (2020).
Mancuso, C. & Santangelo, R. Alzheimer’s disease and gut microbiota modifications: the long way between preclinical studies and clinical evidence. Pharmacol. Res. 129, 329–336 (2018).
Huc, T., Nowinski, A., Drapala, A., Konopelski, P. & Ufnal, M. Indole and indoxyl sulfate, gut bacteria metabolites of tryptophan, change arterial blood pressure via peripheral and central mechanisms in rats. Pharmacol. Res. 130, 172–179 (2018).
Albillos, A., De Gottardi, A. & Rescigno, M. The gut-liver axis in liver disease: pathophysiological basis for therapy. J. Hepatol. 72, 558–577 (2020).
Zaborska, K. E., Lee, S. A., Garribay, D., Cha, E. & Cummings, B. P. Deoxycholic acid supplementation impairs glucose homeostasis in mice. PLoS ONE 13, e0200908 (2018).
Astbury, S. et al. Lower gut microbiome diversity and higher abundance of proinflammatory genus Collinsella are associated with biopsy-proven nonalcoholic steatohepatitis. Gut Microbes 11, 569–580 (2020).
Grimble, R. F. et al. Cysteine and glycine supplementation modulate the metabolic response to tumor necrosis factor alpha in rats fed a low protein diet. J. Nutr. 122, 2066–2073 (1992).
Bajaj, J. S. et al. Fungal dysbiosis in cirrhosis. Gut 67, 1146–1154 (2018).
Forsyth, C. B. et al. Lactobacillus GG treatment ameliorates alcohol-induced intestinal oxidative stress, gut leakiness, and liver injury in a rat model of alcoholic steatohepatitis. Alcohol 43, 163–172 (2009).
Cassard, A. M. & Ciocan, D. Microbiota, a key player in alcoholic liver disease. Clin. Mol. Hepatol. 24, 100–107 (2018).
Song, Q. et al. Bifidobacterium pseudolongum-generated acetate suppresses non-alcoholic fatty liver disease-associated hepatocellular carcinoma. J. Hepatol. 79, 1352–1365 (2023).
Ganesan, R. et al. Characteristics of microbiome-derived metabolomics according to the progression of alcoholic liver disease. Hepatol. Int. 18, 486–499 (2024).
Shu, X. et al. Bifidobacterium lactis TY-S01 protects against alcoholic liver injury in mice by regulating intestinal barrier function and gut microbiota. Heliyon 9, e17878 (2023).
Iwanaga, S. et al. Chromogenic substrates for horseshoe crab clotting enzyme. Its application for the assay of bacterial endotoxins. Haemostasis 7, 183–188 (1978).
Askgaard, G., Kraglund, F., Kann, A. E., Vilstrup, H. & Jepsen, P. [Epidemiology for alcohol-related liver disease]. Ugeskr. Laeger 183, V11200893 (2021).
Li, Y. et al. Bifidobacterium adolescentis CGMCC 15058 alleviates liver injury, enhances the intestinal barrier and modifies the gut microbiota in D-galactosamine-treated rats. Appl. Microbiol. Biotechnol. 103, 375–393 (2019).
Jiang, X. W. et al. New strain of Pediococcus pentosaceus alleviates ethanol-induced liver injury by modulating the gut microbiota and short-chain fatty acid metabolism. World J. Gastroenterol. 26, 6224–6240 (2020).
Acknowledgements
This work was supported by the National Natural Science Foundation of China (82502783); the Zhejiang Provincial Natural Science Foundation of China (LQ22H030013;2022-KY1-001-039); the Fundamental Research Funds for the Central Universities (2025ZFJH03); the State Key Laboratory for Diagnosis and Treatment of Infectious Disease Independent Project (zz202320); the Young Innovative Talents Support Program of Zhejiang Medical and Health Science and Technology Project (2022499572, 2022RC243); and the Young Innovative Talents Support Program of Zhejiang Chinese Medicine Science Technology Project (2024025437, 2024ZR152).
Author information
Authors and Affiliations
Contributions
Y.L. conceptualized the study design and experimental framework. L.Y., H.X. and X.B. conducted the experimental investigations and coordinated the specimen acquisition. W.W. performed nucleic acid library preparation and high-throughput sequencing and established analytical pipelines. D.S. spearheaded the computational analyses and bioinformatics interpretation. L.L. synthesized the experimental findings and composed the initial draft of the manuscript. All the authors provided final approval for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Li, Y., Yang, L., Xu, H. et al. Restoration of ethanol-induced Bifidobacterium pseudocatenulatum depletion ameliorates alcohol-associated liver disease. npj Biofilms Microbiomes 12, 47 (2026). https://doi.org/10.1038/s41522-026-00913-z
Received:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1038/s41522-026-00913-z
